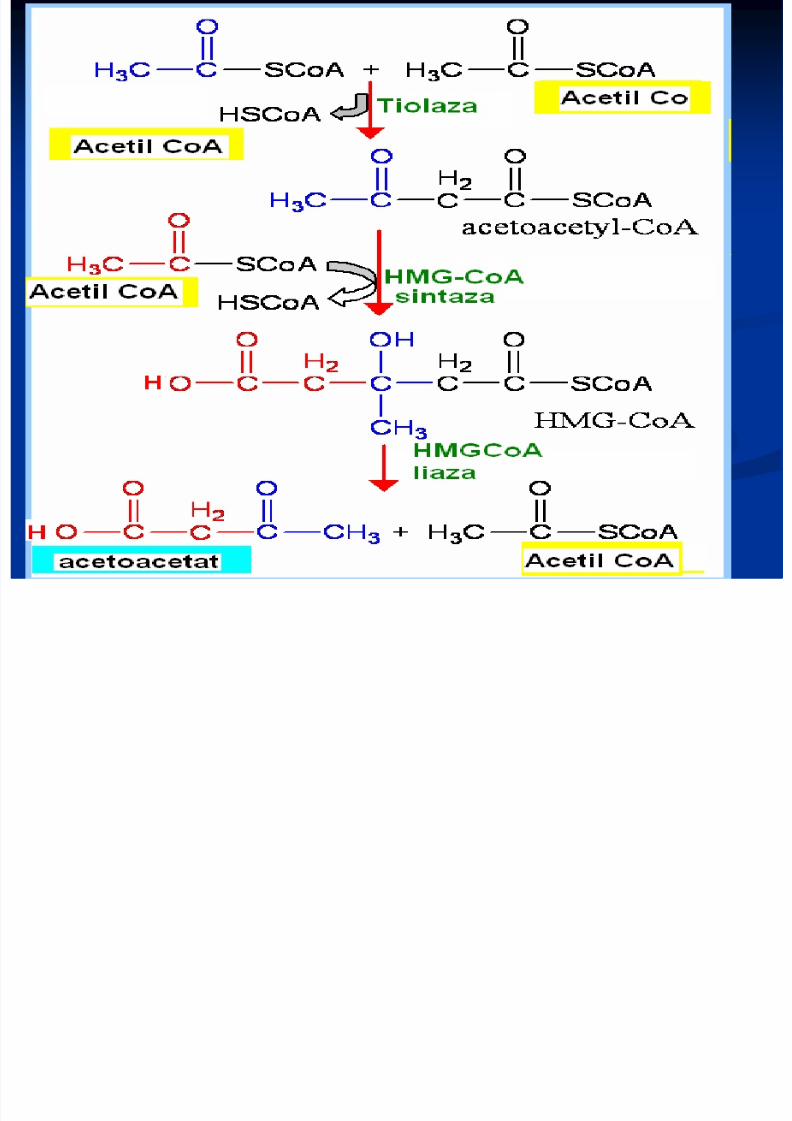
Sinteza corpilor cetonici (cetogeneza) Principala cale de metabolizare a acetil CoA–includerea în ciclul Krebs (în condiţiile în care scindarea lipidelor şi a glucidelor este echilibrată) - “lipidele ard în flacăra glucidelor”În lipsa glucidelor; inaniţie, diabet - OA se utilizează pentru generarea Gl. În lipsa OA, Acetil Co A recurge la formarea corpilor cetonici: acetoacetatul, β-hidrohibutiratul şi acetona Sinteza lor are loc în ficat, dar se utilizează de ţesuturile periferice Au rol energetic (muşchiul cardiac, stratul cortical al rinichilor)
Transcript
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 1/71
Sinteza corpilor cetonici
(cetogeneza)
Principala cale de metabolizare a acetil CoA – includerea în ciclul Krebs (în condiţiile în care scindarealipidelor şi a glucidelor este echilibrată) - “lipidele ard înflacăra glucidelor”
În lipsa glucidelor; inaniţie, diabet - OA se utilizeazăpentru generarea Gl. În lipsa OA, Acetil Co A recurge la formarea corpilor
Sinteza lor are loc în ficat, dar se utilizează de ţesuturileperiferice
Au rol energetic (muşchiul cardiac, stratul cortical alrinichilor)
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 2/71
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 3/71
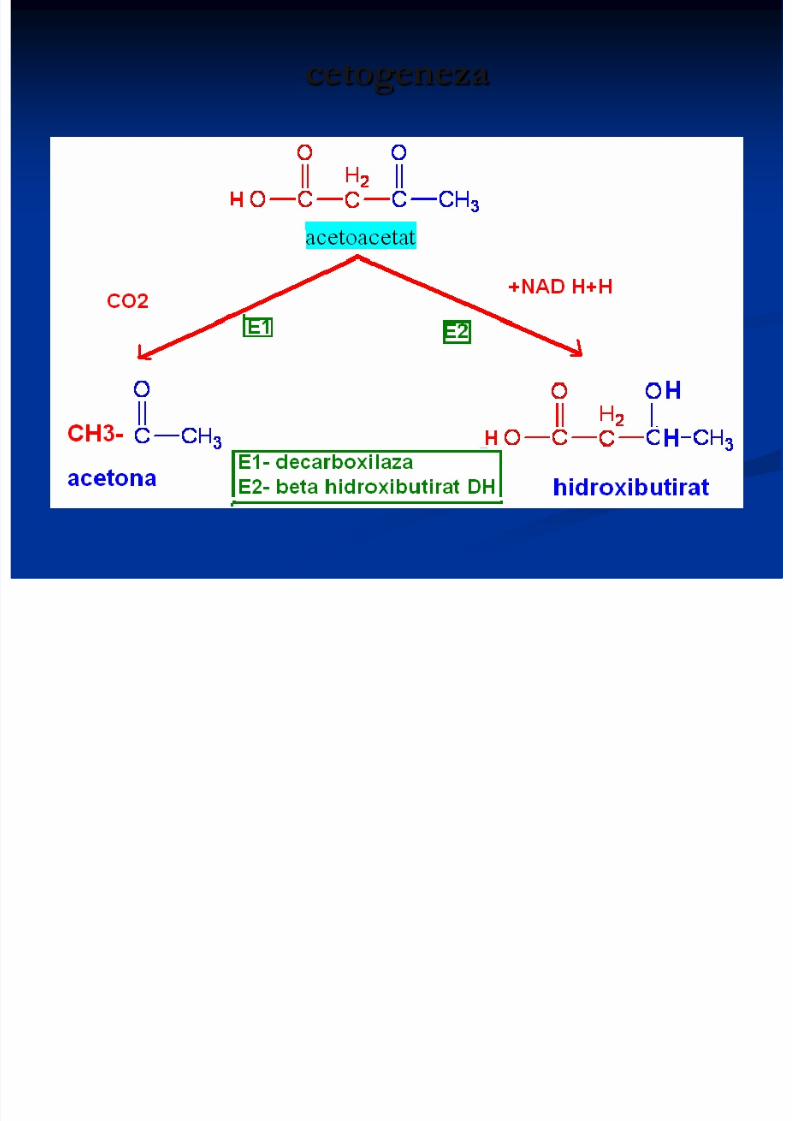
cetogeneza
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 4/71
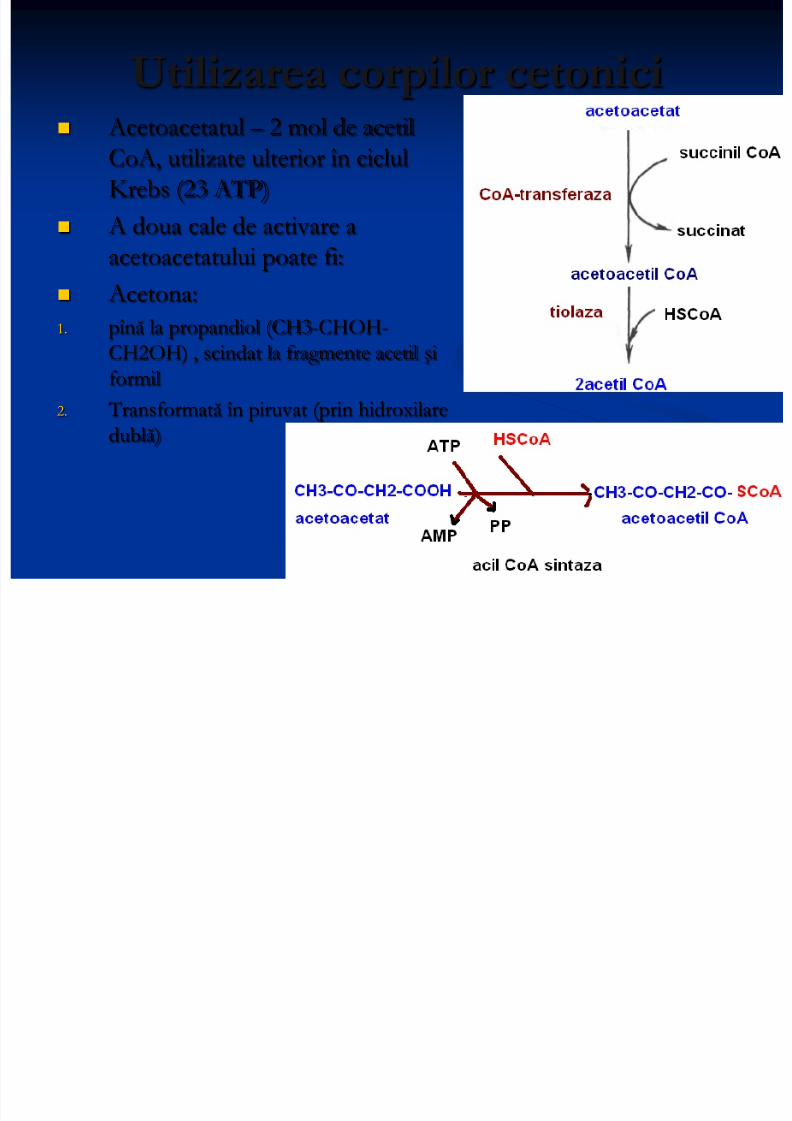
Utilizarea corpilor cetonici
Acetoacetatul – 2 mol de acetilCoA, utilizate ulterior în ciclulKrebs (23 ATP)
A doua cale de activare aacetoacetatului poate fi:
Acetona:1. pînă la propandiol (CH3-CHOH-
CH2OH) , scindat la fragmente acetil şiformil
2. Transformată în piruvat (prin hidroxilaredublă)
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 5/71
Cetonemie, cetonurie
Cetonemie- mărirea c% de corpi cetonici însînge Cetonurie – apariţia CC în urină
Diete bogate în lipide, sărace în glucide; inaniţie,diabet, dereglări gastrointestinale la copii saugravide; glucozurie renală
Eliminarea hidroxibutiratului şi acetoacetatuluidin organism (fiind anioni la excreţie) conduce lapierderea de cationi – Na- rezultă cetoacidoza
Pierderea H2O – dehidratarea organismului
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 6/71
Biosinteza lipidelor
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 7/71
Obiectivele: Biosintaza acizilor graşi: 1. saturaţi cu număr par de atomi de carbon; 2. nesaturaţi cu număr par de atomi de carbon; 3. saturaţi cu număr impar de atomi de carbon. Enzimele, coenzimele, reglarea. Biosinteza TAG: substanţele iniţiale, enzimele şi coenzimele,
reglarea. Biosinteza fosfogliceridelor: substratele, reacţiile parţiale ale I
substratele, etapele, reacţiile parţiale ale I etape (până la acidulmevalonic), enzimele, coenzimele, reglarea. Căile de utilizare şieliminare ale colesterolului.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 8/71
Sinteza AG
Sinteza AG şi încorporarea lor în Tg constituiemecanismul principal de stocare a excesului deglucide alimentare (Gl nu se mai transformă înglicogen dar în Tg)
Etapele:
Sinteza de novo cu formarea acidului palmitic
Elongarea acidului palmitic
Introducerea de legături duble în AG
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 9/71
Particularităţile sintezei AG
Are loc în citozol E – acid gras sintetaza – alcătuită din 8 proteine
(domenii)- 7 sunt enzime, a 8-a – proteina( purtătoare ) transportatoare de acil -ACP.
ACP cuprinde 2 grupe SH:1. – SH furnizat de un rest de cisteinil: SH-Cis2. - SH - fosfopanteteina, ataşată prin leg ătura
fosfat-Ser: SH-Pant Ca iniţiator este acetil CoA (rezultat din
glicoliză), pe cînd sursa majoră – malonil CoA rolul reducător îi revine NADPH+H
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 10/71
Sinteza de novo cu formarea acidului
palmitic
Etapele:
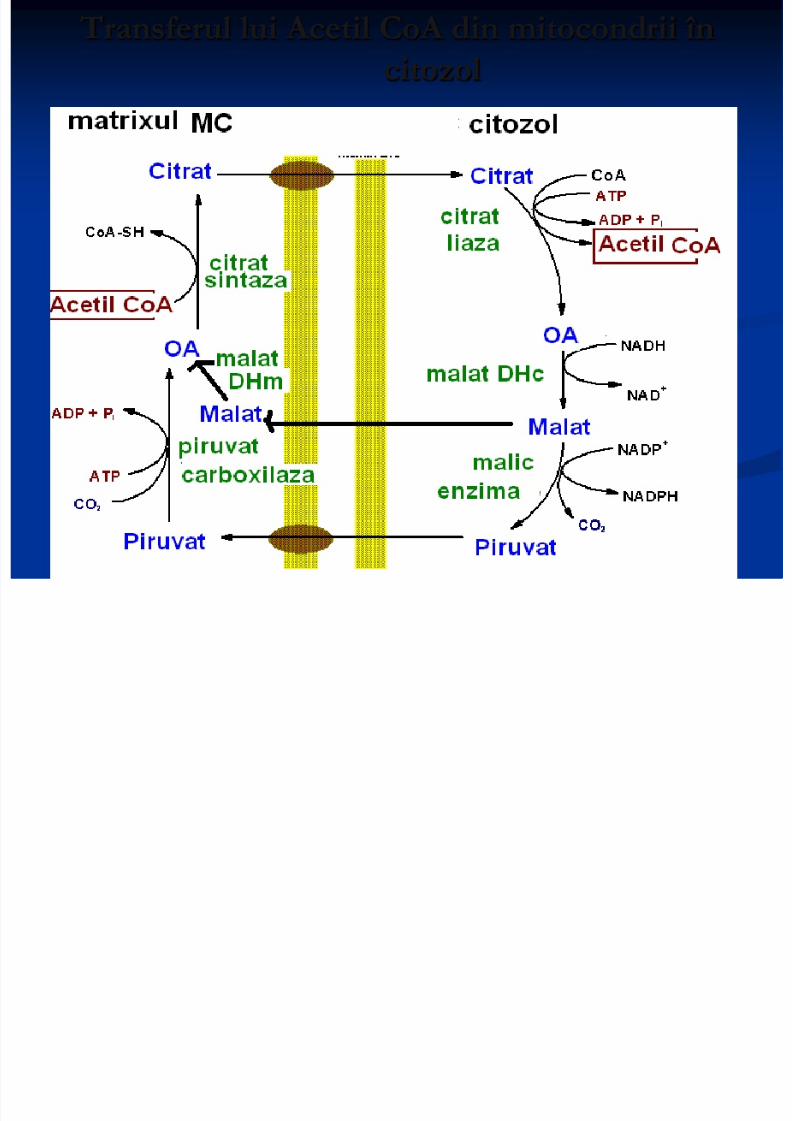
1. transferul lui Acetil CoA din mitocondrii încitozol
2. Sinteza de malonil CoA
3. Sinteza acidului palmitic
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 11/71
Transferul lui Acetil CoA din mitocondrii în
citozol
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 12/71
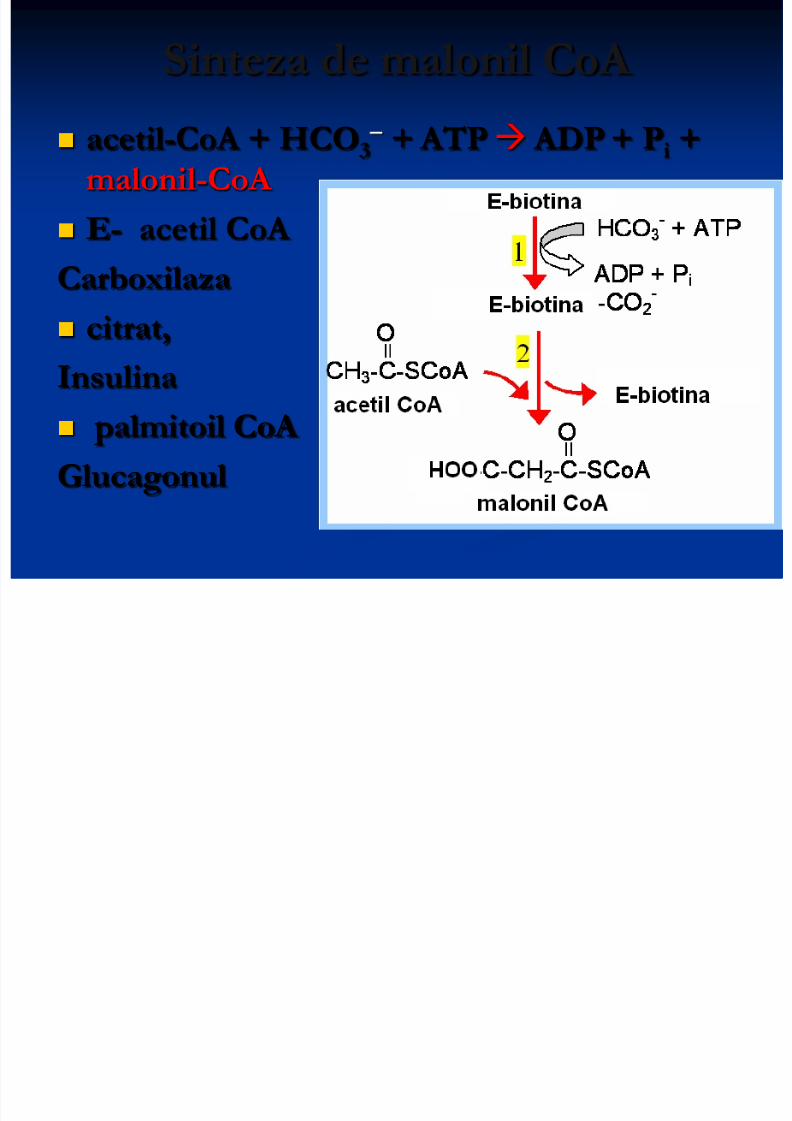
Sinteza de malonil CoA
acetil-CoA + HCO3 + ATP ADP + Pi + malonil-CoA
E- acetil CoA
Carboxilaza
citrat,
Insulina
palmitoil CoA
Glucagonul
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 13/71
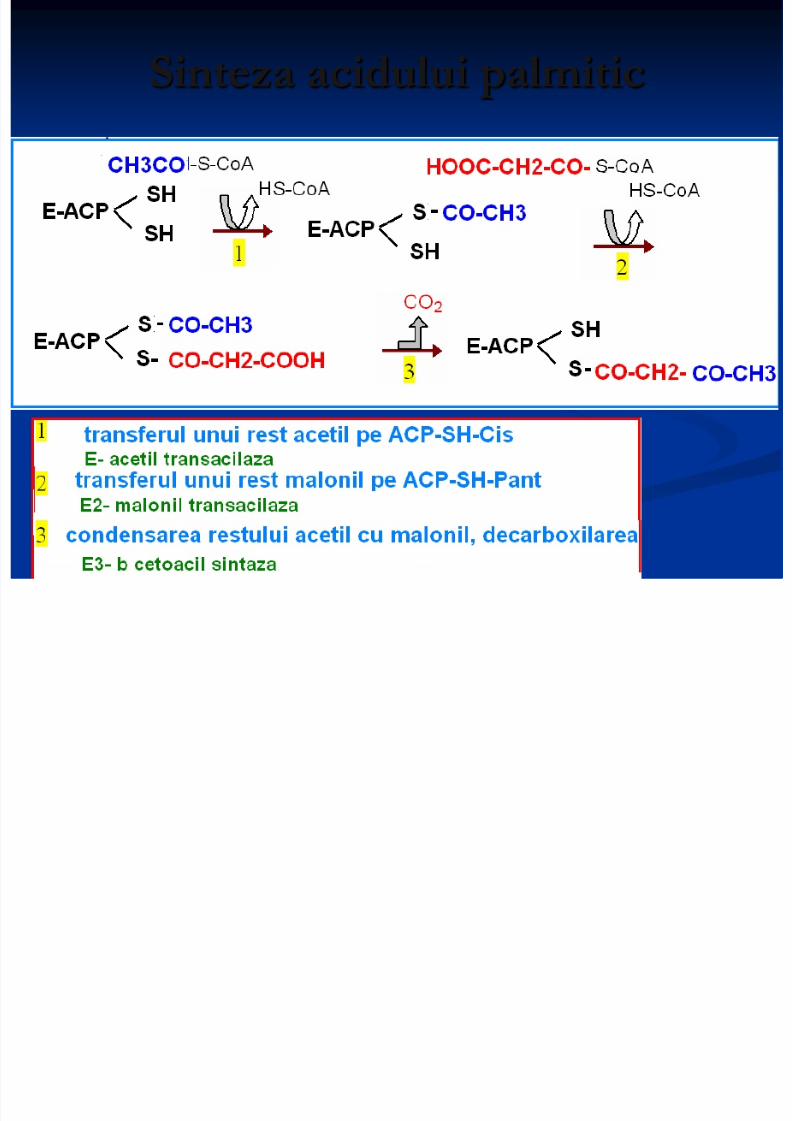
Sinteza acidului palmitic
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 14/71
Sinteza acidului palmitic
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 15/71
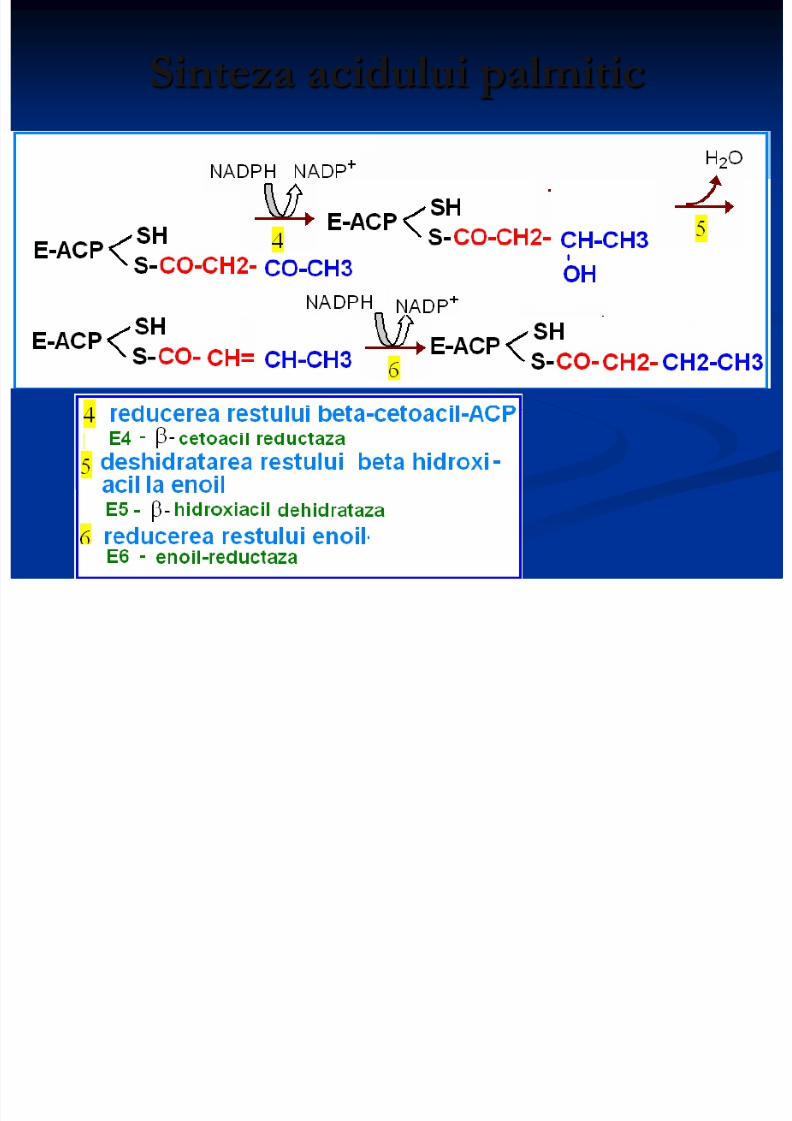
Sinteza acidului palmitic
Ciclu de reacţii este reluat: butiril+ACP secondensează cu malonil+ACP- formînd în final C6-acil ACP.
Catena AG creşte pînă la formarea palmitil-S-ACP
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 16/71
R eacţia sumară:
Acetil-ACP+7 malonil-CoA +14 NADPH+H
Palmitat +7CO2+14NADP + + 8HSCoA+6H2O
deoarece malonil CoA se sintetizează din acetilCoA:8 acetil-CoA + 14 NADPH +H + + 7 ATP
palmitate+ 14
NADP
+
+ 8HSCoA + 7
ADP
+
7
Pi
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 17/71
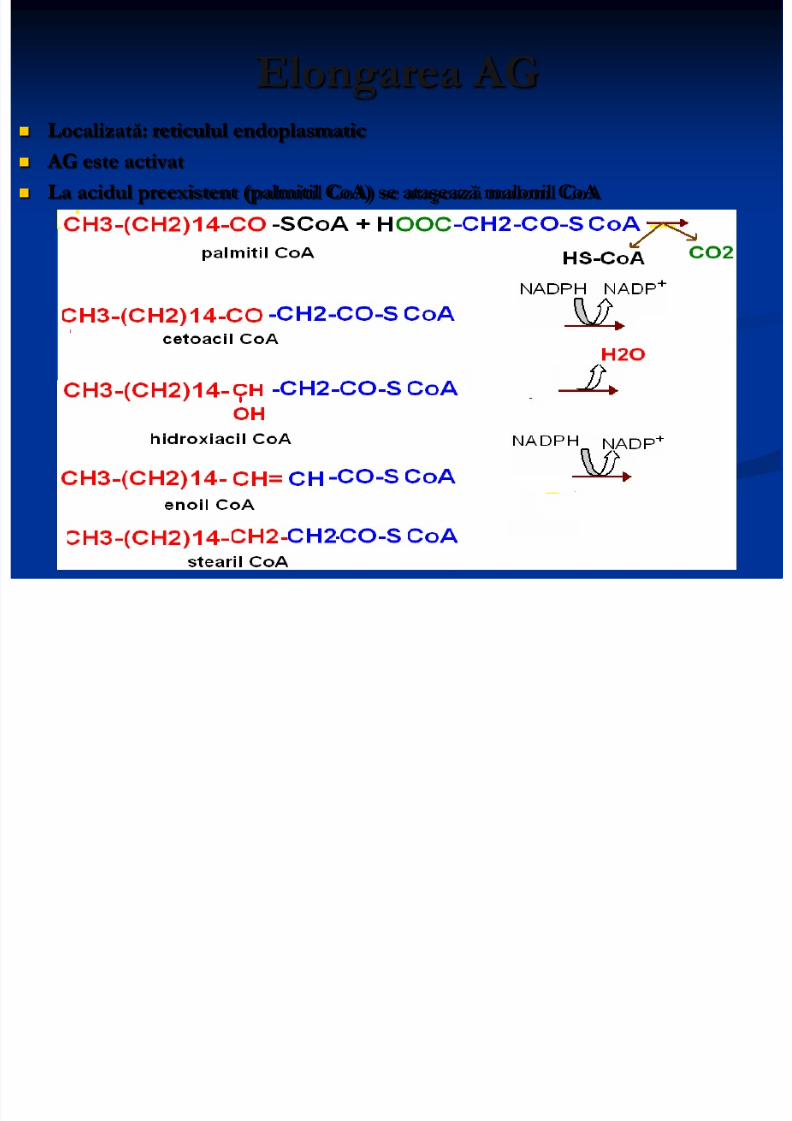
Elongarea AG
Localizată: reticulul endoplasmatic AG este activat
La acidul preexistent (palmitil CoA) se ataşează malonil CoA
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 18/71
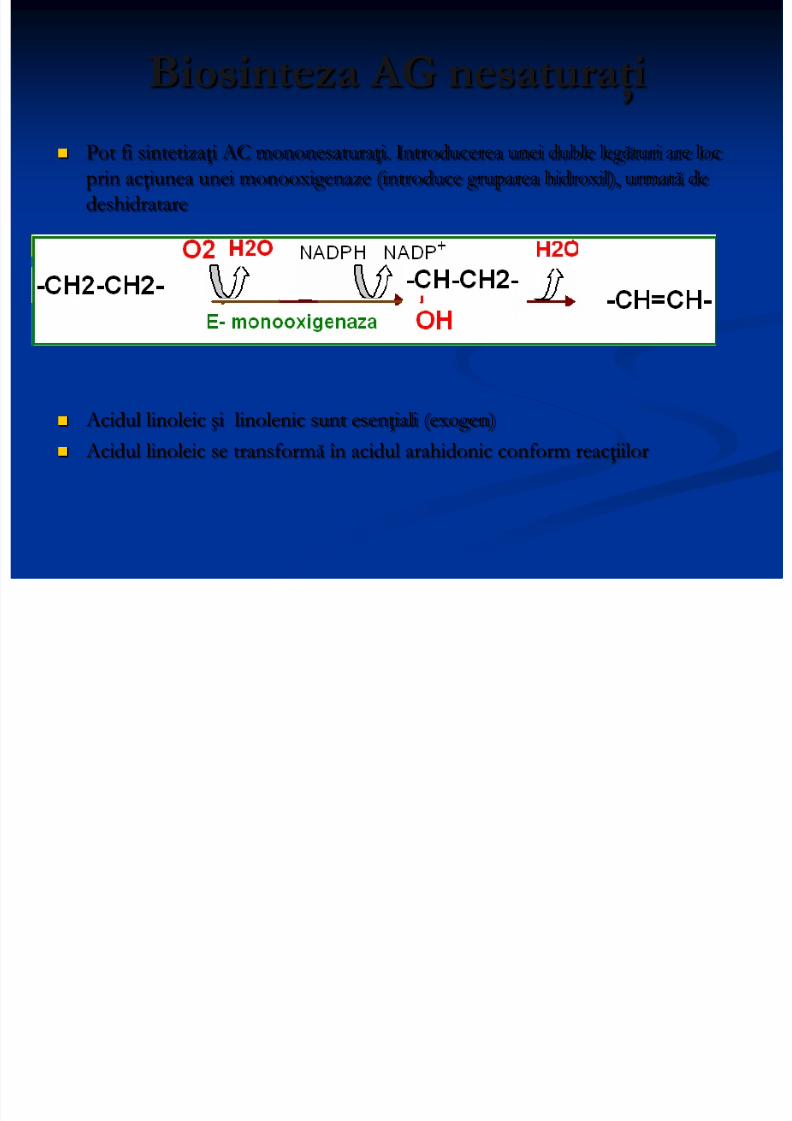
Biosinteza AG nesaturaţi
Pot fi sintetizaţi AC mononesaturaţi. Introducerea unei duble legături are locprin acţiunea unei monooxigenaze (introduce gruparea hidroxil), urmată dedeshidratare
Acidul linoleic şi linolenic sunt esenţiali (exogen) Acidul linoleic se transformă în acidul arahidonic conform reacţiilor
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 19/71
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 20/71
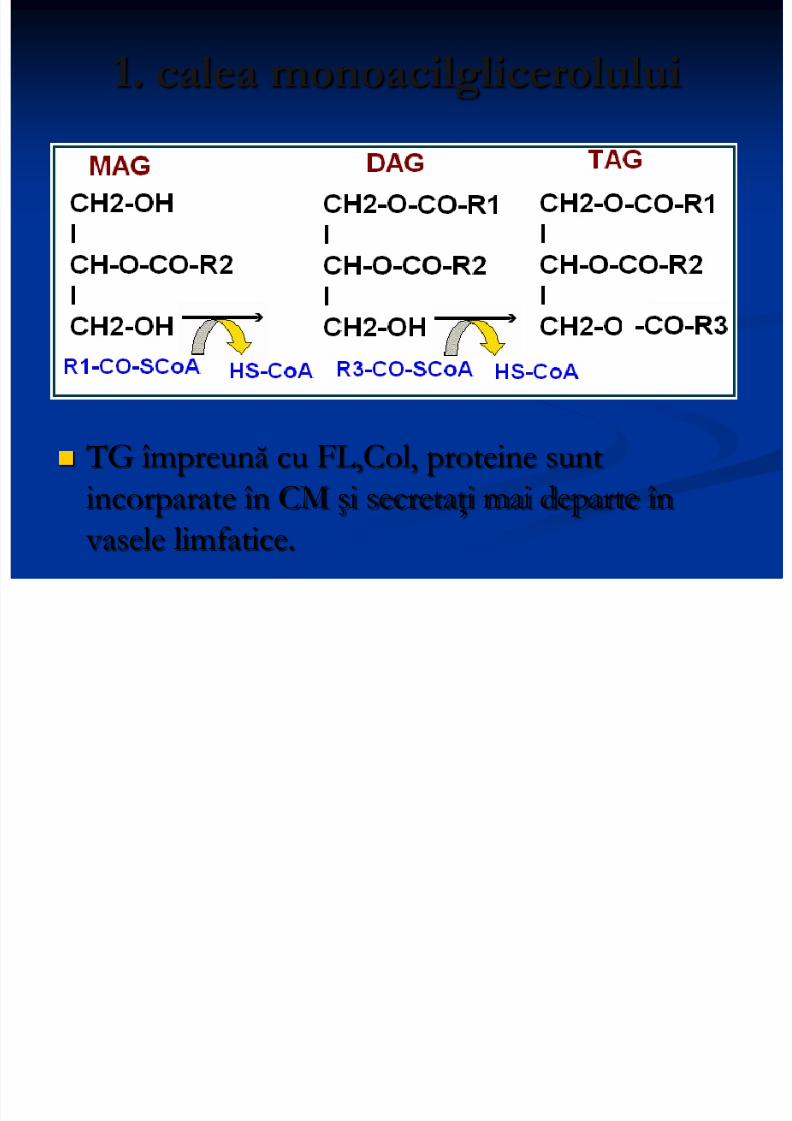
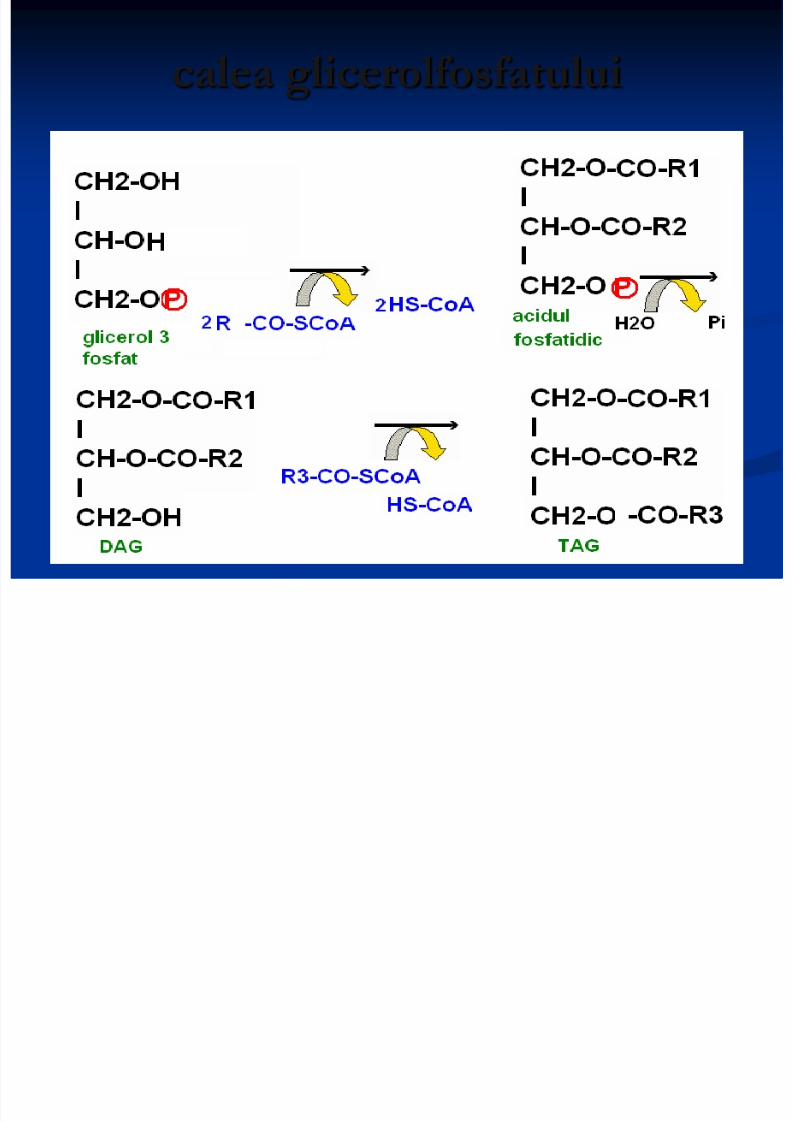
Sinteza TAG
2 căi: 1. calea monoacilglicerolului: are loc în peretele intestinal
(enterocite)din produşi absorbiţi (resinteza lipidelor). 2. calea glicerolfosfatului: în toate ţesuturile (activă: ţesutul adipos
şi ficat) AG sunt incorporaţi în TAG sub formă activă de acilCoA: R-COOH + ATP + HS-CoA +H2O R-CO~SCoA + AMP + 2 Pi
E- acil Co A sintetaza
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 21/71
1. calea monoacilglicerolului
TG împreună cu FL,Col, proteine suntincorparate în CM şi secretaţi mai departe în
vasele limfatice.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 22/71
calea glicerolfosfatului
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 23/71
originea glicerol fosfatului
În ficat:
În ţesut adipos, ficat
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 24/71
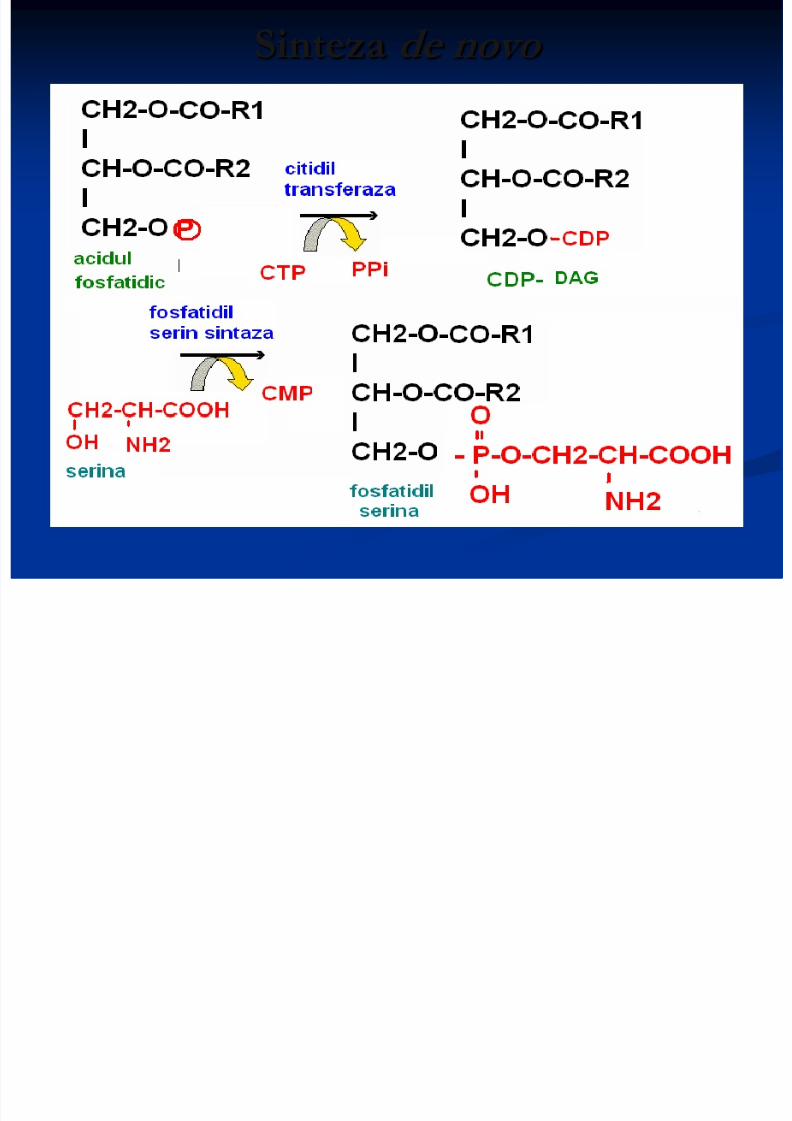
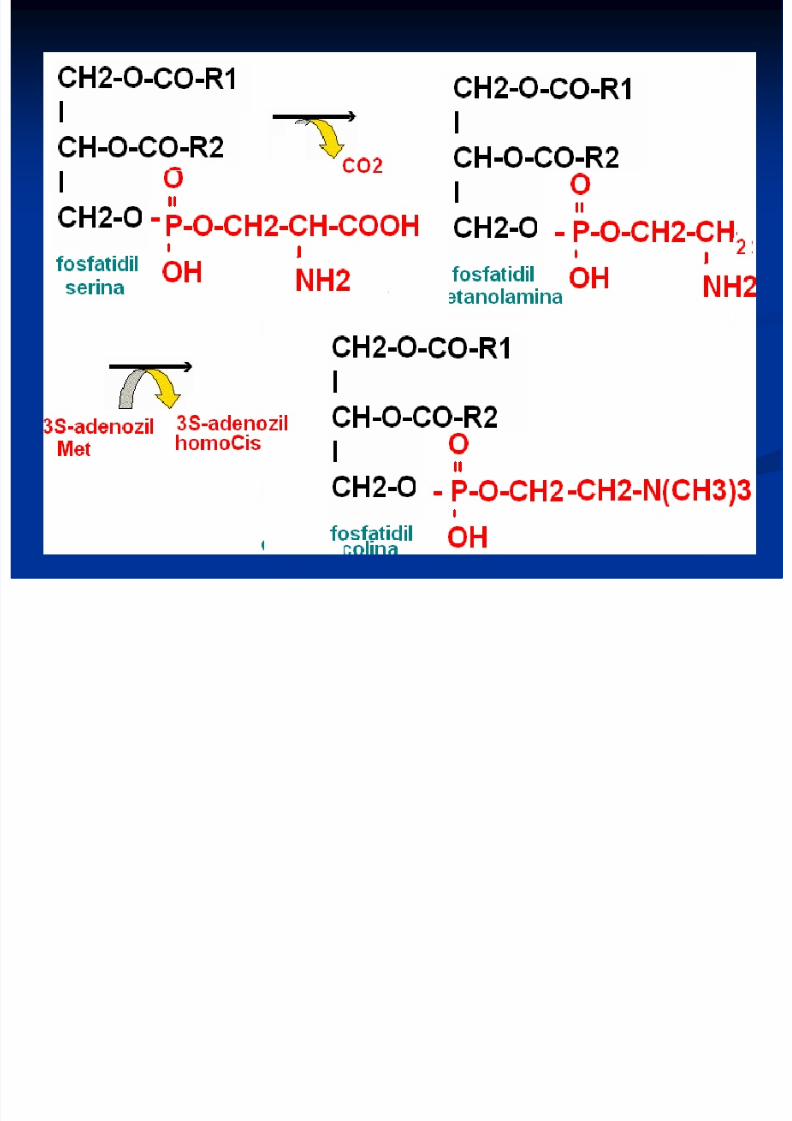
Sinteza glicerofosfolipidelor 2 căî de sinteză:
Sinteza de novo - utilizează ca intermediar comunacidul fosfatidic
Calea de rezervă – o sinteză din produse formate
Particularitatea biosintezei FL este participareaprecursorilor în forme active de derivaţi ai citidinfosfatului (CDP) ca CDP-colina, CDP-
etanolamina, CDP-diglicerid.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 25/71
Sinteza de novo
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 26/71
2 i t di d f t
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 27/71
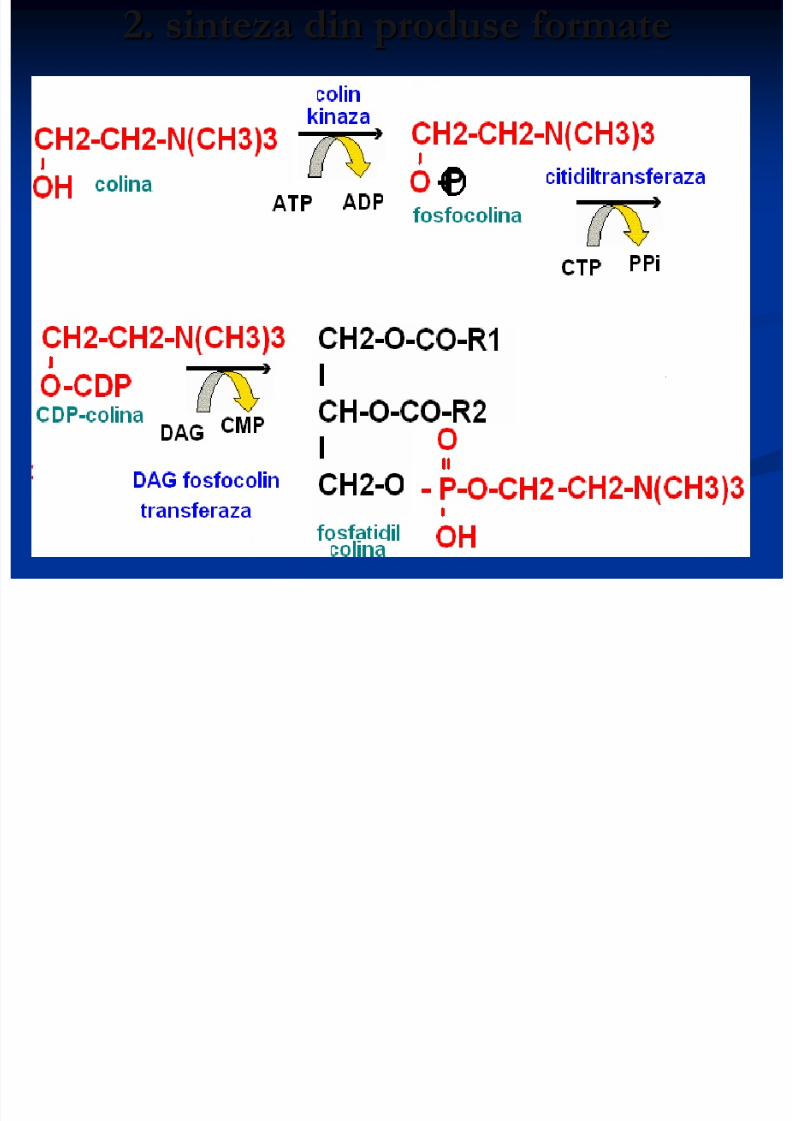
2. sinteza din produse formate
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 28/71
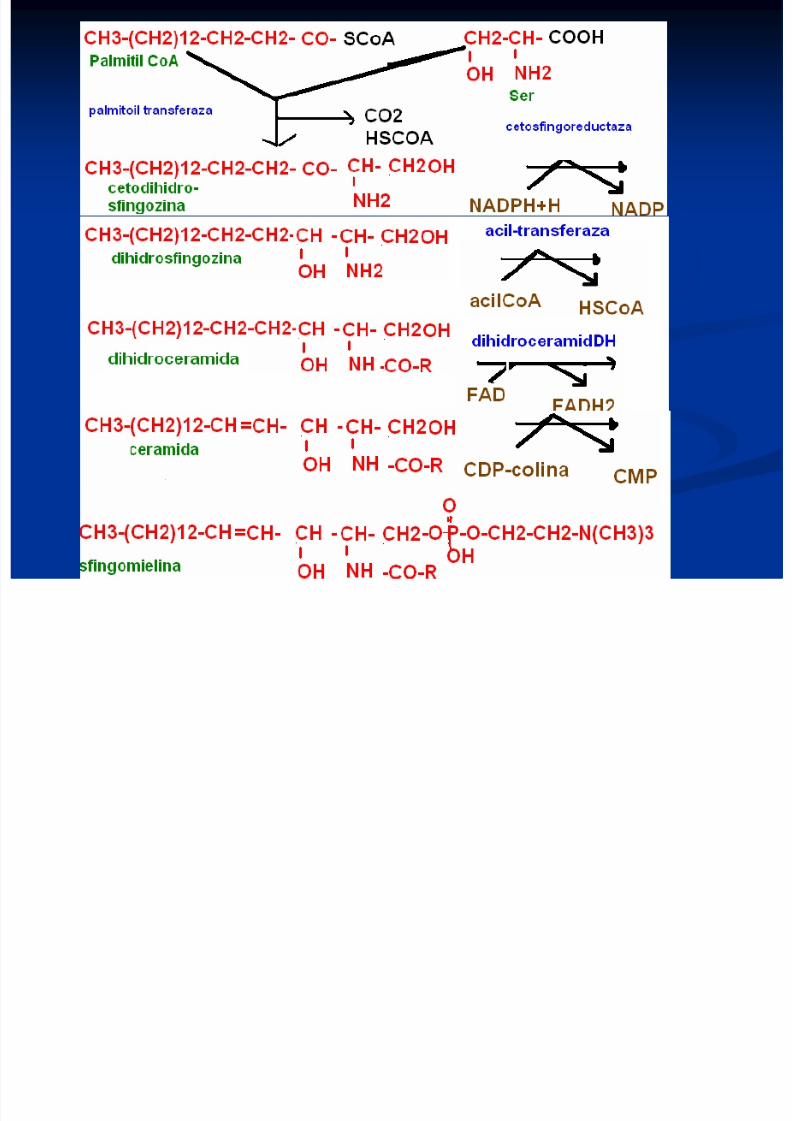
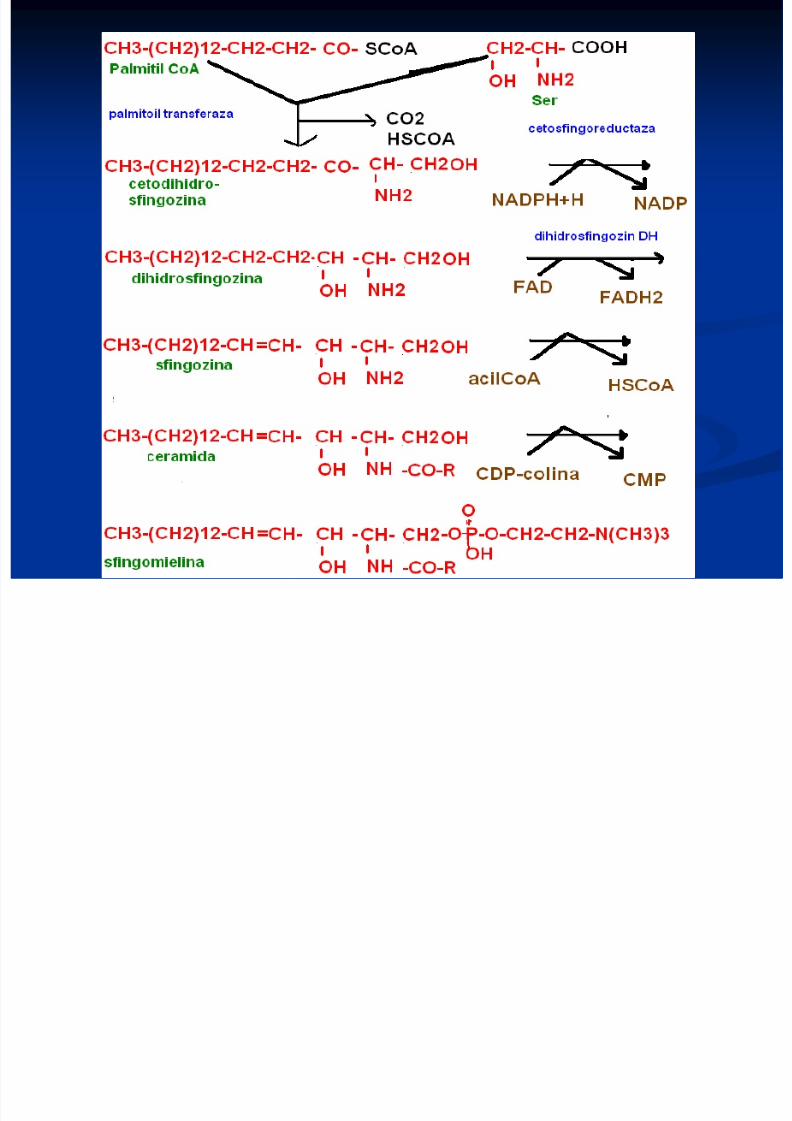
Sinteza sfingolipidelor
Se formează din palmitoil CoA şi Ser
Sfingozina liberă se formează din ceramidă
Sinteza are loc pe suprafaţa citozolică amembranelor reticulului endoplasmatic
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 29/71
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 30/71
Sinteza sfingolipidelor
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 31/71
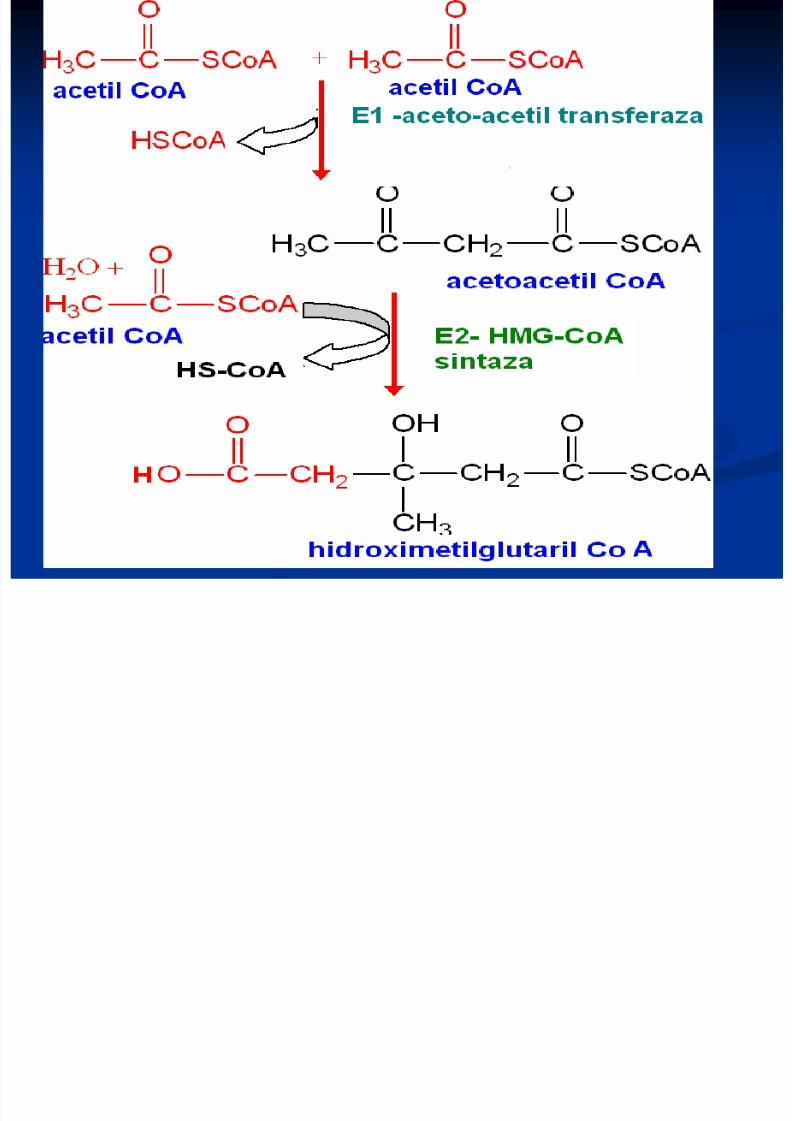
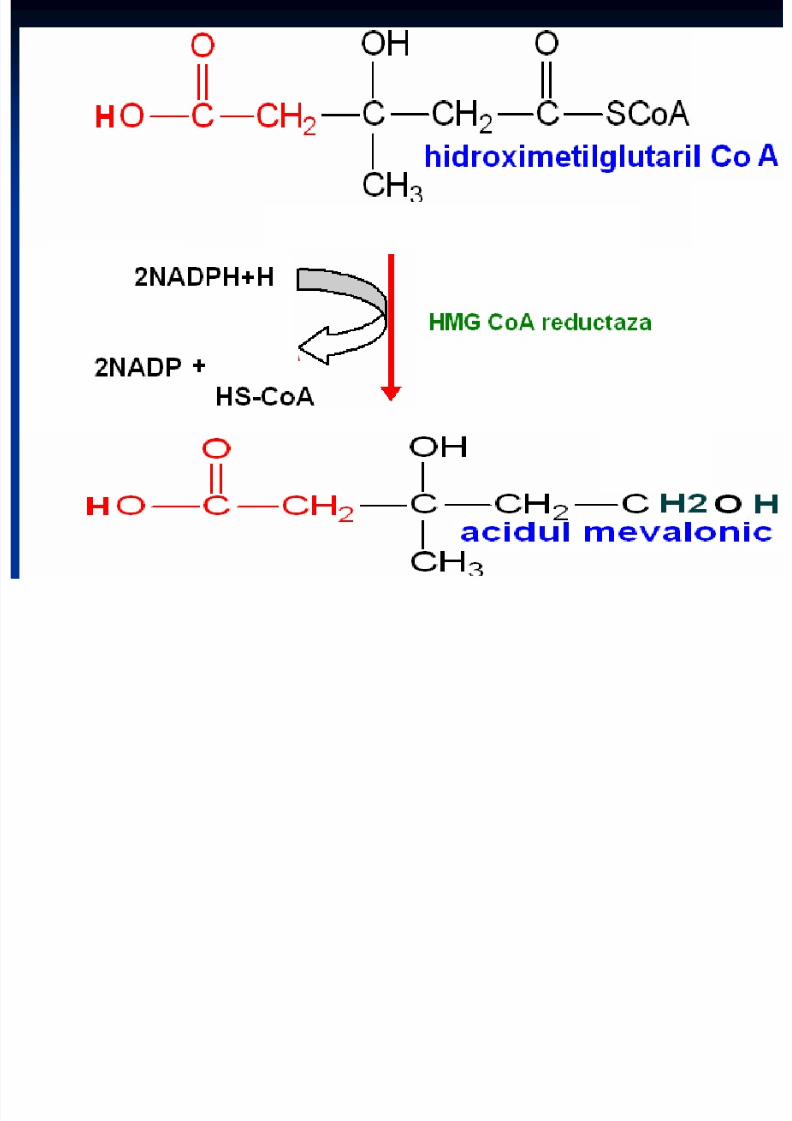
Sinteza Colesterolului
Se sintetizează din Acetil-CoA Necesită 18 moli de Acetil-CoA şi 18 de ATP Principalul organ de metabolizare este ficatul, dar are
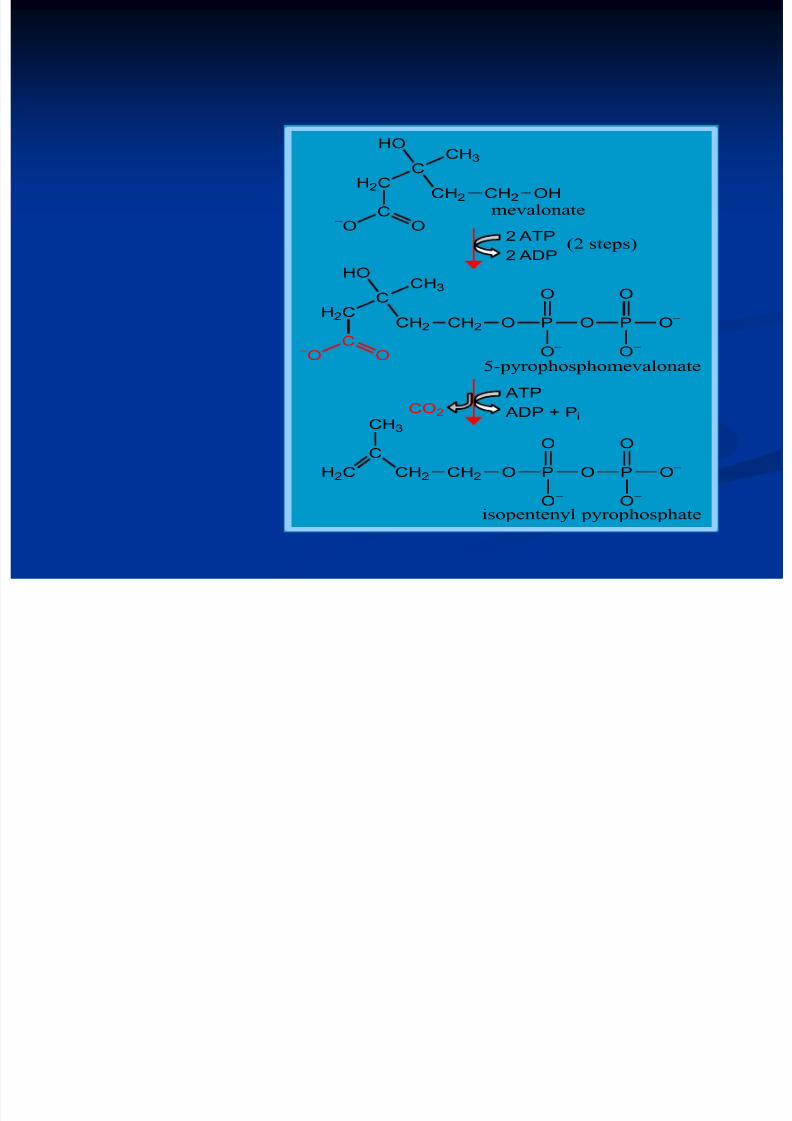
loc şi în intestin, suprarenale, tegumente Are loc în 3 etape:1. Sinteza acidului mevalonic2. mevalonatul prin mai multe reacţii - 3∆-izopentenil
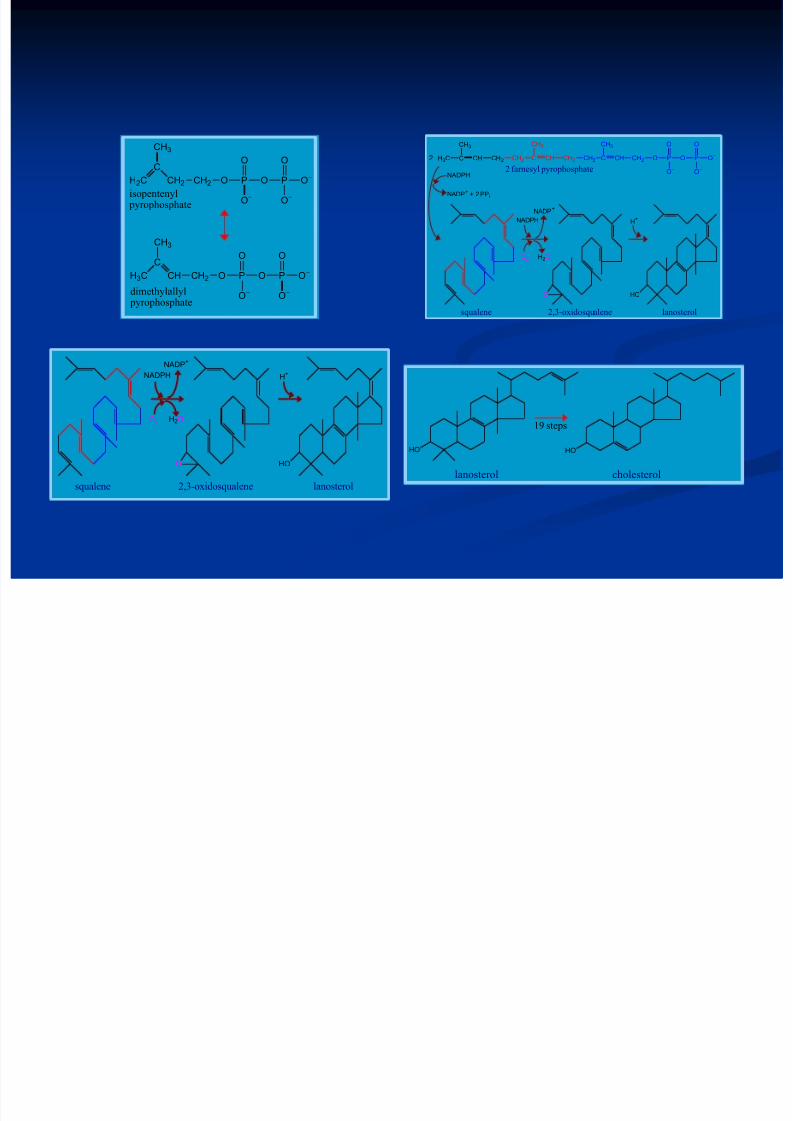
pirofosfat. 6 molecule de 3∆-izopentenil pirofosfat – scualen
3. Scualenul se supuine ciclizării – lanosterol -- Col
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 32/71
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 33/71
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 34/71
H2C
C
CH3
HO
CH2
C-
O O
CH2 OH
H2C
C
CH2 CH2 O P O P O-
O
O-
O
O-
CH3
H2C
C
CH3HO
CH2
C-
O O
CH2 O P O P O-
O
O-
O
O-
CO2
ATP
ADP + Pi
2 ATP
2 ADP
mevalonate
5-pyrophosphomevalonate
(2 steps)
isopentenyl pyrophosphate
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 35/71
H2C
C
CH2 CH2 O P O P O-
O
O-
O
O-
CH3
H3C
C
CH CH2 O P O P O-
O
O-
O
O-
CH3
isopentenyl pyrophosphate
dimethylallyl pyrophosphate
CH CH2CH3C
CH3
CH CH2CCH2
CH3
CH CH2 O P O P O-
O
O-
O
O-
CCH2
CH3
2
O
NADP+
O2 H2O
HO
H+
NADPH
NADP+ + 2 PPi
NADPH
2 farnesyl pyrophosphate
squalene 2,3-oxidosqualene lanosterol
O
NADP+
O2 H2O
HO
H+NADPH
squalene 2,3-oxidosqualene lanosterol
HO HO
lanosterol cholesterol
19 steps
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 36/71
REGLAREA ŞI
PATOLOGIAMETABOLISMULUI
LIPIDIC
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 37/71
Obiectivele Metabolismul eicosanoizilor. Căile ciclooxigenazică şi lipooxigenazică ale biosintezei lor.
Inactivarea. Metabolismul vitaminelor liposolubile: sursele alimentare, necesităţile diurne, transformările Reglarea metabolismului lipidelor la nivelul celulei. Reglarea neurohormonală a metabolismului lipidelor. Rolul lipotropinelor, ACTH, hormonilor
tiroizi, insulinei, glucagonului, glucocorticoizilor şi catecolaminelor. Relaţiile reciproce dintre metabolismul energetic, glucidic şi lipidic. Dereglările digestiei şi absorbţiei lipidelor. Steatoreea pancreatică, hepatică şi intestinală. Dislipidemiile: a) hipolipoproteinemiile familiale – afecţiunea Tangier, - şi -lipoproteinemia familială; b) hiperlipoproteinemiile primare şi familiale; c) hiperlipoproteinemiile secundare (dobândite) – în diabet zaharat, alcoolism, afecţiuni
metacromatică, gangliozidoza GM1; b) dobândite – obezitate, ateroscleroză, alcoolism. Cauze, mecanismele dereglării metabolismului lipidelor, manifestările biochimice. 7. A-, hipo- şi hipervitaminozele A, D, E, K – cauze, manifestări metabolice. 8. Rolul eicosanoizilor în procesele inflamatorii, reacţiile alergice, dereglările fluidităţii
sanguine.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 38/71
Metabolismul eicosanoizilor
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 39/71
ATP-dependent carboxylation provides energy input.
The CO2 is lost later during condensation with thegrowing fatty acid.
The spontaneous decarboxylation drives the condensationreaction.
H3C C SCoA
O
CH2 C SCoA
O
-OOC
acetyl-CoA
malonyl-CoA
The input to fatty acidsynthesis is acetyl-CoA ,
which is carboxylated tomalonyl-CoA .
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 40/71
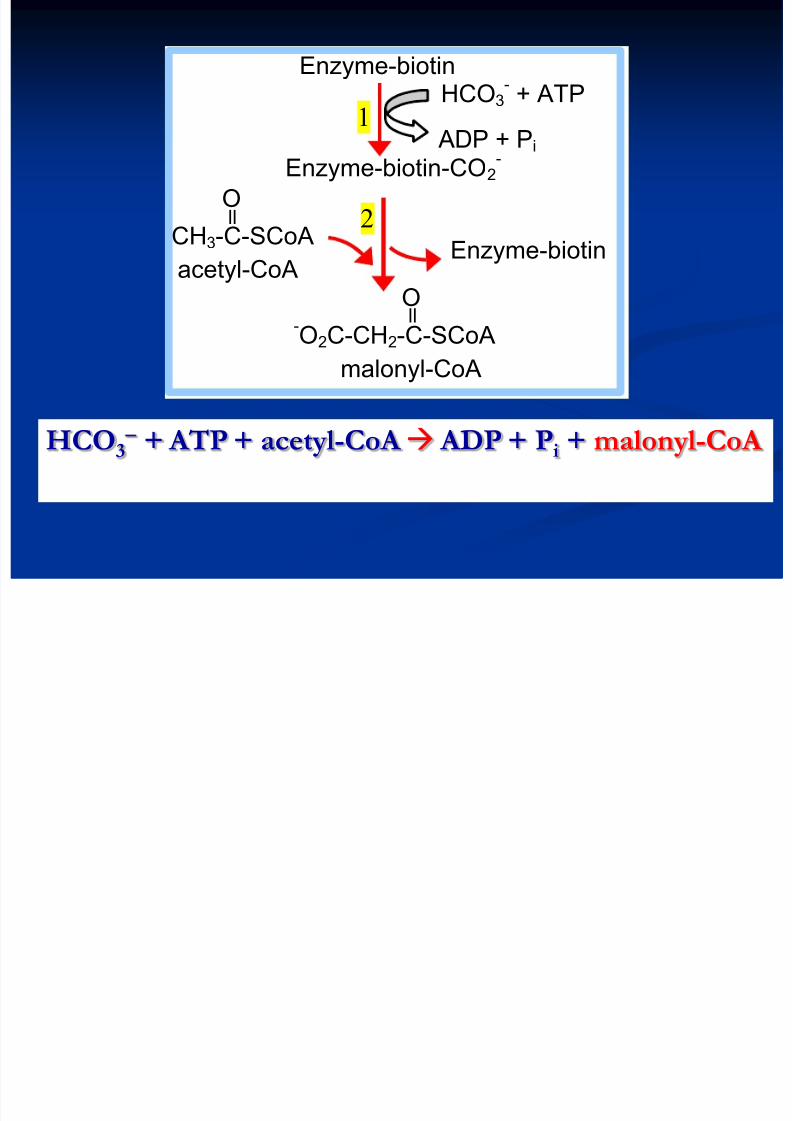
HCO3 + ATP + acetyl-CoA ADP + Pi + malonyl-CoA
ll
Enzyme-biotin HCO3
- + ATP
ADP + Pi
Enzyme-biotin-CO2-
O
CH3-C-SCoA
acetyl-CoA O
-O2C-CH2-C-SCoA
malonyl-CoA
ll
Enzyme-biotin
1
2
O
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 41/71
Biotin is linked to the enzyme by an amide bond between
the terminal carboxyl of the biotin side chain and thee-amino group of a lysine residue. The combined biotin and lysine side chains act as a longflexible arm that allows the biotin ring to translocate
between the 2 active sites.
CHCH
H2C
S
CH
NH
C
N
O
(CH2)4 C NH (CH2)4 CH
CO
NH
O
C
O
O-
Carboxybiotin lysineresidue
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 42/71
Acetyl-CoA Carboxylase, which converts acetyl-CoA tomalonyl-CoA, is the committed step of the fatty acidsynthesis pathway.
The mammalian enzyme is regulated, by
phosphorylation
allosteric control by local metabolites.Conformational changes associated with regulation:
In the active conformation, Acetyl-CoA Carboxylase
associates to form multimeric filamentous complexes. Transition to the inactive conformation is associated
with dissociation to yield the monomeric form of theenzyme (protomer).
Phosphorylated protomer of
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 43/71
The decreased production of malonyl-CoA preventsenergy-utilizing fatty acid synthesis when cellular energystores are depleted. (AMP is abundant only when ATP has
When AMP is high (ATP low), malonyl-CoA production isdiminished, releasing fatty acid oxidation from inhibition.
This will lead to increased ATP production.
AMP-Activated Kinase has a significant role evenin tissues (e.g., cardiac
muscle) that do notsignificantly synthesize fattyacids.
In such tissues malonyl-CoA , produced via oneisoform of Acetyl-CoACarboxylase, functionsmainly as an inhibitor offatty acid oxidation.
H3C C SCoA
O
CH2 C SCoA
O
-OOC
acetyl-CoA
malonyl-CoA
ATP + HCO3-
ADP + Pi
Acetyl-CoACarboxylase
(inhibited by
AMP-ActivatedKinase)
O
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 45/71
A cAMP cascade, activated by glucagon & epinephrine whenblood glucose is low, may also result in phosphorylation of
Acetyl-CoA Carboxylase via cAMP-Dependent Protein
Kinase. With Acetyl-CoA Carboxylase inhibited, acetyl-CoA remainsavailable for synthesis of ketone bodies, the alternativemetabolic fuel used when blood glucose is low.
H3C C SCoA
O
CH2 C SCoA
O
-OOC
acetyl-CoA
malonyl-CoA
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 46/71
The antagonistic effect of insulin, produced when bloodglucose is high, is attributed to activation of ProteinPhosphatase.
Palmitoyl-CoA (product of Fatty Acid Synthase) promotesthe inactive conformation, diminishing production ofmalonyl-CoA, the precursor of fatty acid synthesis.
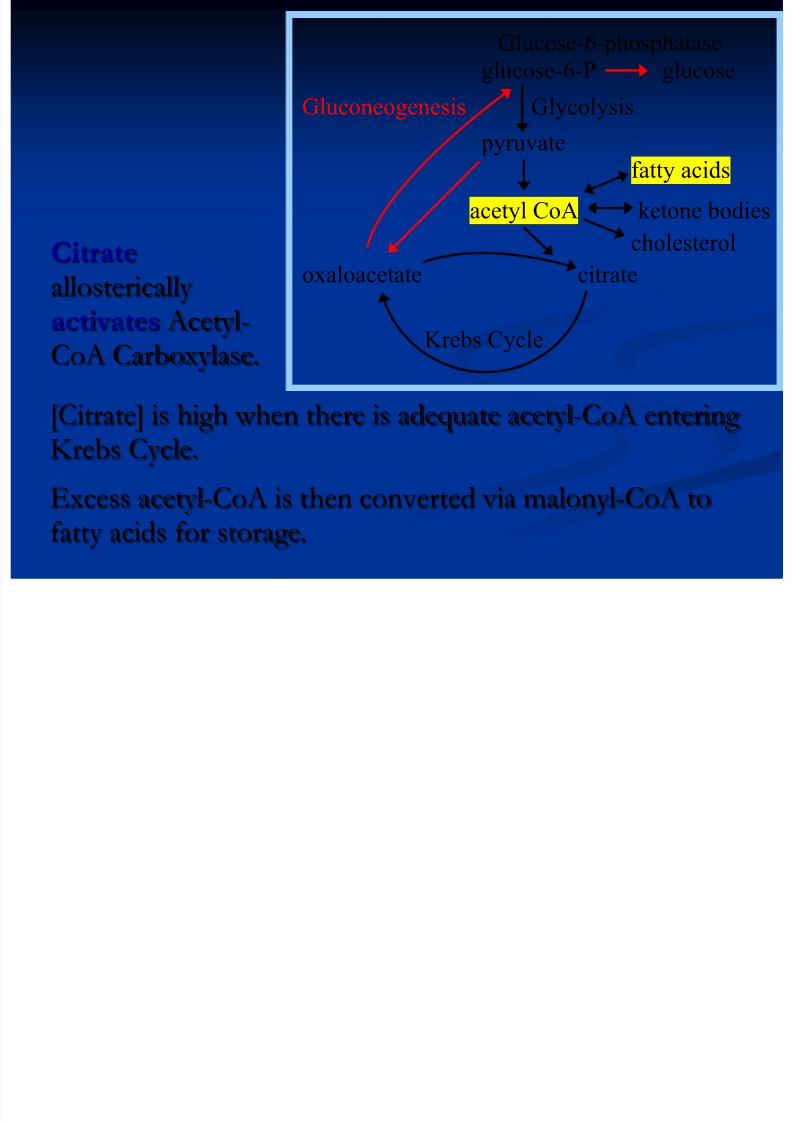
[Citrate] is high when there is adequate acetyl-CoA enteringKrebs Cycle.
Excess acetyl-CoA is then converted via malonyl-CoA tofatty acids for storage.
Glucose-6-phosphatase
glucose-6-P glucose
Gluconeogenesis Glycolysis
pyruvatefatty acids
acetyl CoA ketone bodies
cholesterol
oxaloacetate citrate
Krebs Cycle
Citrate
allostericallyactivates Acetyl-CoA Carboxylase.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 49/71
Fatty acid synthesis from acetyl-CoA & malonyl-CoAoccurs by a series of reactions that are:
in bacteria catalyzed by 6 different enzymes plus aseparate acyl carrier protein (ACP)
in mammals catalyzed by individual domains of a very
large polypeptide that includes an ACP domain.Evolution of the mammalian Fatty Acid Synthaseapparently has involved gene fusion.
NADPH serves as electron donor in the two reactionsinvolving substrate reduction.
The NADPH is produced mainly by the Pentose PhosphatePathway.
SH Coenzyme AH
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 50/71
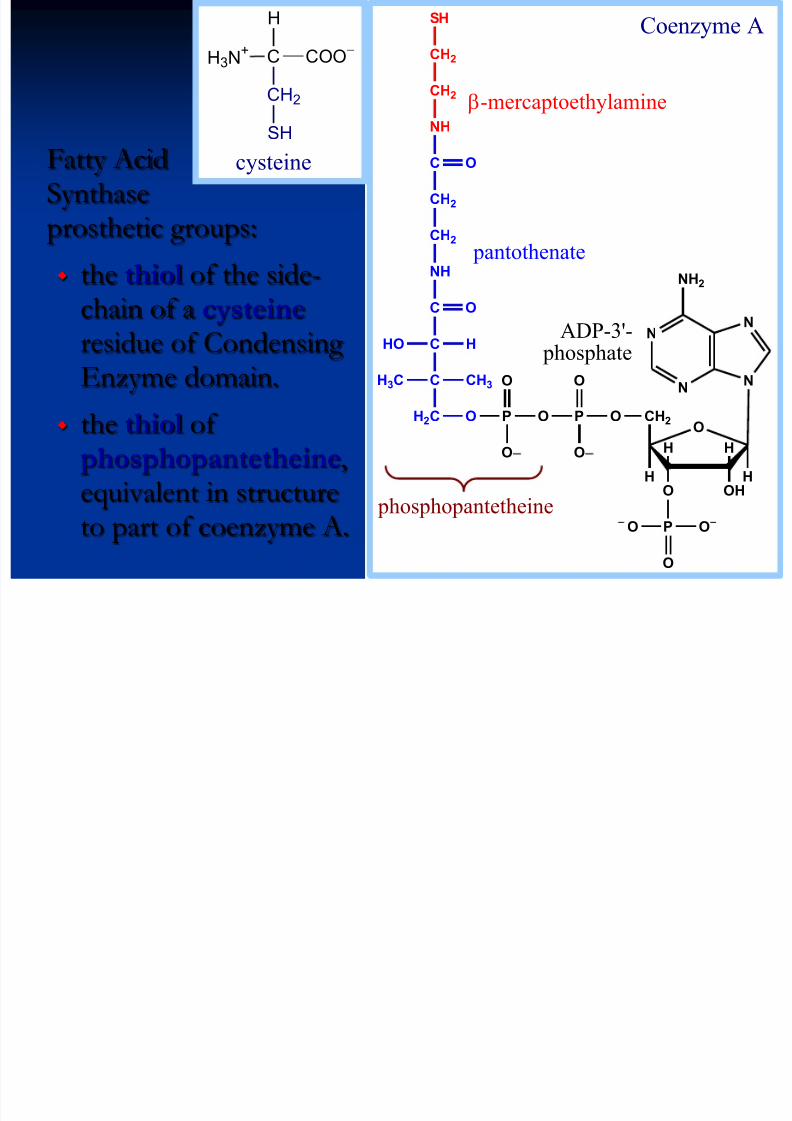
Fatty AcidSynthaseprosthetic groups:
the thiol of the side-chain of a cysteine residue of CondensingEnzyme domain.
the thiol of phosphopantetheine,equivalent in structureto part of coenzyme A.
N
N N
N
NH2
O
OHO
HH
H
CH2
H
OPOPOH2C
O
O O
O
P
O
O
O
C
C
C
NH
CH2
CH2
C
NH
CH3H3C
HHO
O
CH2
CH2
O
-mercaptoethylamine
pantothenate
ADP-3'- phosphate
y
phosphopantetheine
H3N+ C COO
-
CH2
SH
cysteine
SHh h t th i
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 51/71
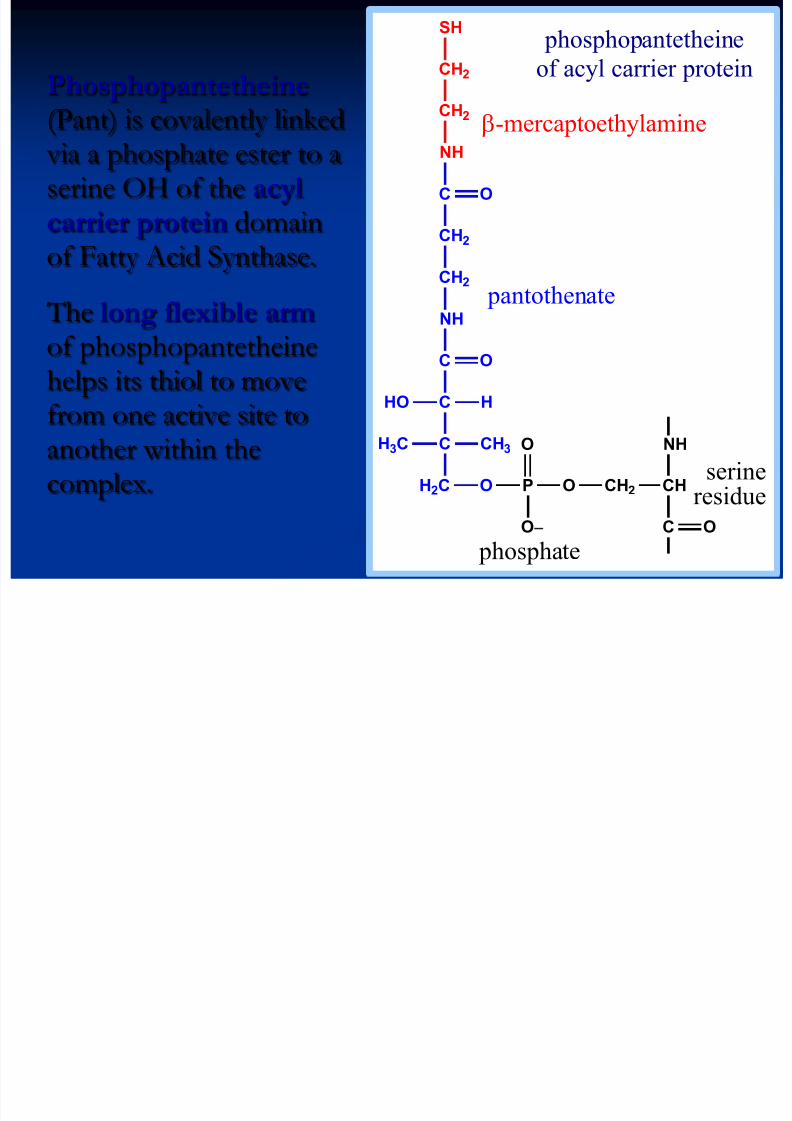
Phosphopantetheine (Pant) is covalently linked
via a phosphate ester to aserine OH of the acylcarrier protein domainof Fatty Acid Synthase.
The long flexible arm of phosphopantetheinehelps its thiol to move
from one active site toanother within thecomplex. OPOH2C
O
OC
C
C
NH
CH2
CH2
C
NH
CH3H3C
HHO
O
CH2
CH2
O
CH2 CH
NH
C O
-mercaptoethylamine
pantothenate
serineresidue
phosphopantetheine
of acyl carrier protein
phosphate
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 52/71
As each of the substrates acetyl-CoA & malonyl-CoA bindto the complex, the initial attacking group is the oxygen ofa serine hydroxyl group of the Malonyl/acetyl-CoA
Transacylase enzyme domain.
Each acetyl or malonyl moiety is transiently in ester linkageto this serine hydroxyl, before being transferred into
thioester linkage with the phosphopantetheine thiol ofthe acyl carrier protein (ACP) domain.
Acetate is subsequently transferred to a cysteine thiol ofthe Condensing Enzyme domain.
Order of domains in primary structure of mammalian Fatty Acid Synthase
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 53/71
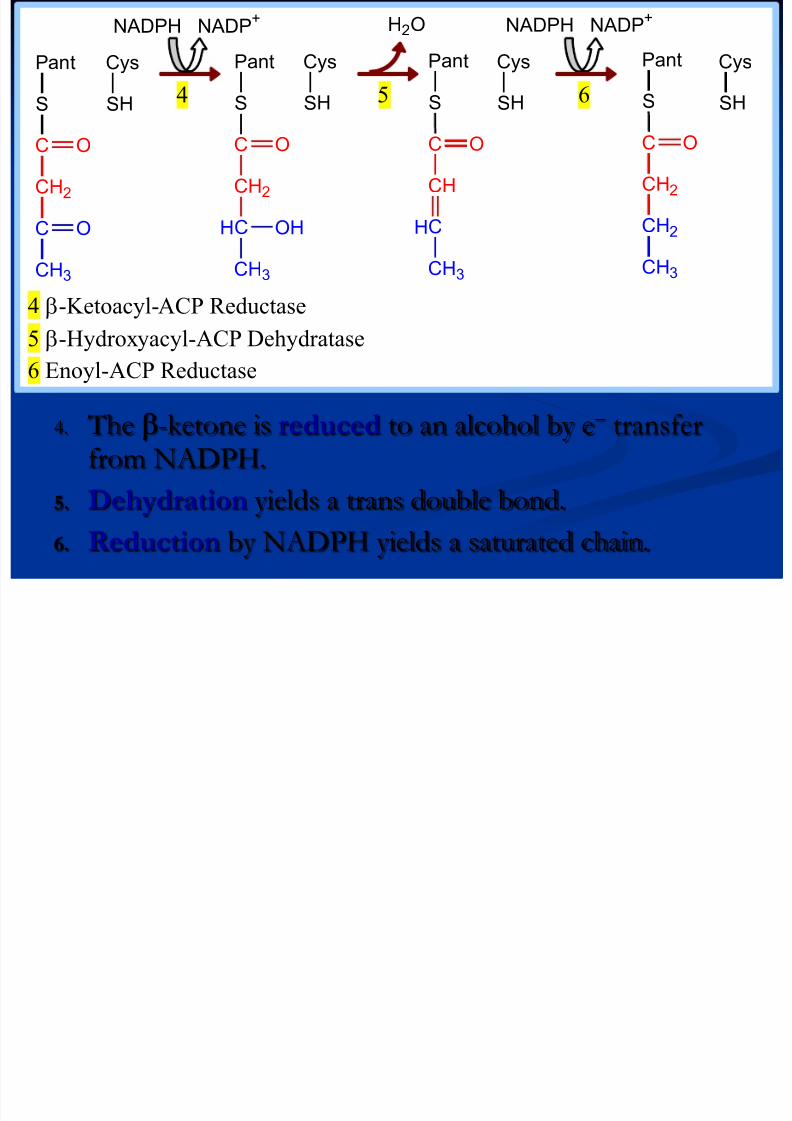
NADPH NADP+NADPH NADP
+ H2O
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 54/71
4. The -ketone is reduced to an alcohol by e- transferfrom NADPH.
5. Dehydration yields a trans double bond.
6. Reduction by NADPH yields a saturated chain.
Pant
S
Cys
SH
C
CH2
C
O
CH3
O
Pant
S
Cys
SH
C
CH2
HC
O
CH3
Pant
S
Cys
SH
Pant
S
Cys
SH
C
CH
HC
O
CH3
C
CH2
CH2
O
CH3
OH
4 5 6
4 -Ketoacyl-ACP Reductase
5 -Hydroxyacyl-ACP Dehydratase
6 Enoyl-ACP Reductase
Malonyl S CoA HS CoA
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 55/71
Following transfer of the growing fatty acid fromphosphopantetheine to the Condensing Enzyme's cysteinesulfhydryl, the cycle begins again, with another malonyl-
CoA.
Pant
S
Cys
SH
C
CH2
CH2
O
CH3
Pant
SH
Cys
S
C
CH2
O
CH2
CH3
Pant
S
Cys
S
C
CH2
O
CH2
CH3
C
CH2
COO-
O
Malonyl-S-CoA HS-CoA
7 2
7 Condensing Enzyme
2 Malonyl/acetyl-CoA-ACP Transacylase (repeat).
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 56/71
Product release:
When the fatty acid is 16 carbon atoms long, a Thioesterase domain catalyzes hydrolysis of the thioesterlinking the fatty acid to phosphopantetheine.
The 16-C saturated fatty acid palmitate is the finalproduct of the Fatty Acid Synthase complex.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 57/71
Condensing Malonyl/acetyl-CoA Dehydratase Enoyl -Ketoacyl ACP Thioesterase(C ) l (S ) d d ( )
N- -C
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 58/71
The solved structure does not resolve the position of ACP & Thioesterase domains, predicted from primary structureto be near -Ketoacyl Reductase (KR) domains of lateral"arms" of the complex.
Enzyme (Cys) Transacylase (Ser ) Reductase Reductase (Pant) N -C
Order of domains in primary structure of mammalian Fatty Acid Synthase
KR KR
DH DH
KS KS
MAT MAT
ER ER
Arrangement of domains
in Fatty Acid Synthase
“arm”
“leg”
These domains may betoo flexible to be resolved.KR = -Ketoacyl Reductase;ER = Enoyl Reductase;
Ketoacyl-ACP Synthase III, equivalent to thedomains of the mammalian Fatty Acid Synthasethat catalyze the initial acetylation andcondensation reactions.
Oxidation & Fatty Acid Synthesis
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 61/71
-Oxidation & Fatty Acid SynthesisCompared
Oxidation Pathway Fatty Acid Synthesis
pathway location mitochondrial matrix cytosol
acyl carriers(thiols)
Coenzyme-A phosphopantetheine(ACP) & cysteine
e
acceptors/donor FAD & NAD+ NADPH
-OH intermediate L D
2-C product/donor acetyl-CoAmalonyl-CoA
(& acetyl-CoA)
F A id S h i i i ll l d
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 62/71
Fatty Acid Synthase is transcriptionally regulated.In liver:
Insulin, a hormone produced when blood glucose ishigh, stimulates Fatty Acid Synthase expression. Thus excess glucose is stored as fat. Transcription factors that that mediate the stimulatory
effect of insulin include USFs (upstream stimulatoryfactors) and SREBP-1.SREBPs (sterol response element binding proteins)
were first identified for their regulation of cholesterol
synthesis. Polyunsaturated fatty acids diminish transcription of
the Fatty Acid Synthase gene in liver cells, bysuppressing production of SREBPs.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 63/71
In fat cells:
Expression of SREBP-1 and of Fatty Acid Synthase isinhibited by leptin, a hormone that has a role in regulatingfood intake and fat metabolism.
Leptin is produced by fat cells in response to excess fatstorage.
Leptin regulates body weight by decreasing food intake,
increasing energy expenditure, and inhibiting fatty acidsynthesis.
Elongation beyond the 16-C length of the palmitate product
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 64/71
Elongation beyond the 16-C length of the palmitate productof Fatty Acid Synthase occurs in mitochondria andendoplasmic reticulum (ER).
Fatty acid elongation within mitochondria involves the-oxidation pathway running in reverse, but NADPH
serves as electron donor for the final reduction step.
Polyunsaturated fatty acids esterified to CoA aresubstrates for the ER elongation machinery, which usesmalonyl-CoA as donor of 2-carbon units.
The reaction sequence is similar to Fatty Acid Synthase
but individual steps are catalyzed by separate proteins. A family of enzymes designated Fatty Acid Elongases catalyze the initial condensation step for elongation of
saturated or polyunsaturated fatty acids.
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 65/71
Desaturases introduce double bonds at specific
positions in a fatty acid chain.Mammalian cells are unable to produce double bonds atcertain locations, e.g., D12.
Thus some polyunsaturated fatty acids are dietaryessentials, e.g., linoleic acid, 18:2 cis D9,12 (18 C atomslong, with cis double bonds at carbons 9-10 & 12-13).
C
O
OH
910
oleate 18:1 cis D9
O
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 66/71
Formation of a double bond in a fatty acid involves thefollowing endoplasmic reticulum membrane proteins in
mammalian cells: NADH-cyt b5 Reductase, a flavoprotein with FAD
as prosthetic group.
Cytochrome b5, which may be a separate protein or adomain at one end of the desaturase.
Desaturase, with an active site that contains twoiron atoms complexed by histidine residues.
COH
910
oleate 18:1 cis D9
8/13/2019 Sinteza AG
http://slidepdf.com/reader/full/sinteza-ag 67/71
The desaturase catalyzes a mixed function oxidation reaction.
There is a 4-electron reduction of O2 2 H2O as a fattyacid is oxidized to form a double bond.
2e pass f rom NADH to the desaturase via theFAD-containing reductase & cytochrome b5, the
order of electron transfer being:NADH FAD cyt b5 desaturase
2e are extracted from the fatty acid as the double
bond is formed.E.g., the overall reaction for desaturation of stearate (18:0)to form oleate (18:1 cis D9 ) is:stearate + NADH + H+ + O2 oleate + NAD+ +