363
Teodor MARUŞCA – coordonator T R A T A T DE RECONSTRUCŢIE ECOLOGICĂ A HABITATELOR DE PAJIŞTI ŞI TERENURI DEGRADATE MONTANE EDITURA UNIVERSITĂŢII TRANSILVANIA DIN BRAŞOV 2010

Teodor MARUŞCA – coordonator
T R A T A T
DE RECONSTRUCŢIE ECOLOGICĂ A HABITATELOR DE PAJIŞTI
ŞI TERENURI DEGRADATE MONTANE
EDITURA UNIVERSITĂŢII TRANSILVANIA DIN BRAŞOV 2010

Teodor MARUSCA – coordonator
Marius I. BĂRBOS, Vasile A. BLAJ, Vasile CARDAŞOL, Neculai DRAGOMIR, Vasile MOCANU, Ioan ROTAR, Mariana RUSU, Ion SECELEANU
T R A T A T
DE RECONSTRUCŢIE ECOLOGICĂ A HABITATELOR DE PAJIŞTI
ŞI TERENURI DEGRADATE MONTANE
EDITURA UNIVERSITĂŢII TRANSILVANIA DIN BRAŞOV 2010


3
CUPRINS
Pag. Cuprins ………………………………………………………………….. 3 Introducere ……………………………………………………………… 11
I. CERCETĂRI PRIVIND VEGETAŢIA PAJIŞTILOR MONTANE ŞI ALE STAŢIUNILOR ……………………………………………….. 13
1.1. Scurt istoric ………………………………………………………………. 131.2. Scheme de clasificare a pajiştilor şi tipuri de habitate de pajişti montane 171.3. Condiţii staţionale ale câmpurilor experimentale de cercetare pentru
îmbunătăţirea şi folosirea pajiştilor ……………………………………… 28II. DESCRIEREA PAJIŞTILOR DEGRADATE DE Nardus stricta DIN
JUDEŢUL BRAŞOV …………………………………………………… 322.1. Condiţii naturale ……………………………………………………….. 32
2.1.1. Cadrul geografic şi relieful ………………………………………………. 32 2.1.2. Geologia …………………………………………………………………. 33
2.1.3. Clima …………………………………………………………………….. 352.1.4. Solul ……………………………………………………………………… 382.1.5. Aspecte privind flora şi vegetaţia ………………………………………... 41
2.2. Obiectivele cercetării şi metoda de lucru ……………………………… 432.2.1. Principii generale ………………………………………………………… 442.2.2. Studiul geobotanic ……………………………………………………….. 452.2.3. Studiul tipologic …………………………………………………………. 542.2.4. Bonitarea nardetelor ……………………………………………………… 56
2.3. Ecologia şi compoziţia floristică a nardetelor ………………………… 582.3.1. Ecologia speciei Nardus stricta L. ………………………………………. 592.3.2. Câteva date privind structura compoziţiei floristice a nardetelor ……….. 70
2.4. Descrierea asociaţiilor de Nardus stricta L. ………………………….. 722.4.1. Asociaţia Nardetum alpigenum …………………………………………. 762.4.2. Asociaţia Nardo – Campanuletum (abietinae) ………………………….. 812.4.3. Asociaţia Nardo – Juncetum (conglomerati) ……………………………. 922.4.4. Indicii sinecologici ai nardetelor ………………………………………… 111
2.5. Descrierea tipurilor de nardete ………………………………………... 1132.5.1. Nardete subalpine situate pe pante foarte puternic înclinate sau abrupte 1132.5.2 Nardete subalpine situate pe terenuri plane sau uşor până la mijlociu
înclinate …………………………………………………………………. 1142.5.3. Nardete cu diferite specii de Vaccinium ………………………………… 1142.5.4. Nardete montane cu Festuca rubra şi Agrostis capillaris situate pe pante
foarte puternic înclinate sau abrupte …………………………………….. 1152.5.5. Nardete montane cu Festuca rubra şi Agrostis capillaris situate pe pante
foarte slab până la mijlociu înclinate ……………………………………. 115

4
2.5.6. Nardete colinare, cu diferite specii mezofile şi xeromezofile situate pe pante mijlocii până la foarte puternic înclinate ………………………….. 116
2.5.7. Nardete din depresiuni, cu diferite specii mezohigrofile şi higrofile situate pe terenuri plane sau uşor înclinate ……………………………… 116
2.6. Alte descrieri ale nardetelor …………………………………………… 1172.6.1. Bonitarea nardetelor ……………………………………………………… 1172.6.2. Relaţia între fitocenoze, tipuri şi bonitare ……………………………….. 119
III. ASPECTE PRIVIND ECOLOGIA PAJIŞTILOR DIN ZONA MONTANĂ …………………………………………………………… 122
3.1. Principalii factori ecologici care influenţează structura şi dinamica habitatelor de pajişti ……………………………………………………. 122
3.1.1. Factorii topografici (relieful) …………………………………………….. 1233.1.2. Factorii edafici …………………………………………………………… 1253.1.3. Factorii climatici …………………………………………………………. 1353.1.4. Factorii biotici …………………………………………………………… 136
3.2. Fitodiversitatea habitatelor de pajişti din zona montană ……………. 1403.3. Ameninţări potenţiale ………………………………………………….. 145IV. ÎNVELIŞUL DE SOLURI ALE PAJIŞTILOR MONTANE ………... 1514.1. Solurile din etajul pajiştilor alpine …………………………………….. 1524.2. Soluri din etajul jnepenişurilor (subalpin) ……………………………. 1564.3. Soluri din etajul molidişurilor (boreal) ……………………………….. 1584.4. Soluri din etajul foioaselor (nemoral) …………………………………. 166
4.5. Caracteristici generale ale solurilor …………………………………… 1874.5.1. Învelişul de soluri ale Carpaţilor Orientali ………………………………. 1874.5.2. Învelişul de soluri ale Carpaţilor Meridionali ……………………………. 1914.5.3. Învelişul de soluri ale Carpaţilor Occidentali ……………………………. 193
4.6. Rezultate de cercetare din punctele cheie de observaţii ……………… 195V. PRINCIPALELE TIPURI DE HABITATE DE PAJIŞTI DIN ZONA
MONTANĂ .............................................................................................. 2165.1. Argument pentru conservarea biodiversităţii ........................................ 2165.2. Consideraţii metodologice ……………………………………………… 2175.3. Descrierea principalelor tipuri de habitate de pajişti din zona
montană a României ……………………………………………………. 2185.3.1. Pajişti alpine şi subalpine ………………………………………………. 218
5.3.1.1. Habitatul R3601 - Pajişti sud-est carpatice de rogoz pitic (Kobresia myosuroides) şi Oxytropis carpatica …………………………………….. 218
5.3.1.2. Habitatul R3602 - Pajişti sud-est carpatice de coarnă (Carex curvula) şi Primula minima …………………………………………………………. 220
5.3.1.3. Habitatul R3603 - Pajişti sud-est carpatice de părul porcului (Juncus trifidus) şi Oreochloa disticha …………………………………………… 222

5
5.3.1.4. Habitatul R3604 - Pajişti sud-est carpatice de păruşcă (Festuca airoides) şi Potentilla aurea ssp. chrysocraspeda ………………………….. 224
5.3.1.5. Habitatul R3605 - Pajişti sud-est carpatice de păiuş de colţi (Festuca versicolor) şi Sesleria rigida ssp. haynaldiana …………………………. 226
5.3.1.6. Habitatul R3606 – Pajişti sud-est carpatice de păiuş de stânci (Festuca saxatilis) …………………………………………………………………. 228
5.3.1.7. Habitatul R3607 - Pajişti sud-est carpatice de Festuca amethystina şi Dianthus tenuifolius ……………………………………………………… 230
5.3.1.8. Habitat R3608 - Pajişti sud-est carpatice de Scorzonera purpurea ssp. rosea şi Festuca nigrescens ……………………………………………… 232
5.3.1.9. Habitatul R3609 - Pajişti sud-est carpatice de ţăpoşică (Nardus stricta) şi Viola declinata …………………………………………………………… 234
5.3.1.10 Habitatul R3610 - Pajişti sud-est carpatice de Poa media ……………... 2365.3.1.11 Habitatul R3611 – Pajişti sud-est carpatice de coada iepurelui (Sesleria
rigida ssp haynaldiana) şi rogoz (Carex sempervirens) ………………… 2385.3.1.12 Habitatul R3612 – Pajişti sud-est carpatice de rogoz (Carex
sempervirens) şi coarnă mare (Sesleria bielzii) ………………………… 2405.3.1.13 Habitatul R3613 – Pajişti sud-est carpatice de Festuca carpatica,
Carduus kerneri şi Trisetum fuscum …………………………………….. 2425.3.1.14 Habitatul R3614 - Pajişti sud-est carpatice de Festuca xanthina ……… 244
5.3.2. Pajişti umede şi comunităţi de ierburi înalte (buruienişuri) ………… 2465.3.2.1. Habitatul R3708 – Comunităţi daco-getice cu Angelica sylvestris, Crepis
paludosa şi Scirpus sylvaticus …………………………………………… 2465.3.2.2. Habitatul R3710 – Pajişti dacice de Molinia coerulea …………………... 2475.3.3.3. Habitat R3712 – Comunităţi dacice cu Deschampsia caespitosa şi
Agrostis stolonifera ………………………………………………………. 2485.3.3. Pajişti mezofile …
5
5.3.1.4. Habitatul R3604 - Pajişti sud-est carpatice de păruşcă (Festuca airoides) şi Potentilla aurea ssp. chrysocraspeda ………………………….. 224
5.3.1.5. Habitatul R3605 - Pajişti sud-est carpatice de păiuş de colţi (Festuca versicolor) şi Sesleria rigida ssp. haynaldiana …………………………. 226
5.3.1.6. Habitatul R3606 – Pajişti sud-est carpatice de păiuş de stânci (Festuca saxatilis) …………………………………………………………………. 228
5.3.1.7. Habitatul R3607 - Pajişti sud-est carpatice de Festuca amethystina şi Dianthus tenuifolius ……………………………………………………… 230
5.3.1.8. Habitat R3608 - Pajişti sud-est carpatice de Scorzonera purpurea ssp. rosea şi Festuca nigrescens ……………………………………………… 232
5.3.1.9. Habitatul R3609 - Pajişti sud-est carpatice de ţăpoşică (Nardus stricta) şi Viola declinata …………………………………………………………… 234
5.3.1.10 Habitatul R3610 - Pajişti sud-est carpatice de Poa media ……………... 2365.3.1.11 Habitatul R3611 – Pajişti sud-est carpatice de coada iepurelui (Sesleria
rigida ssp haynaldiana) şi rogoz (Carex sempervirens) ………………… 2385.3.1.12 Habitatul R3612 – Pajişti sud-est carpatice de rogoz (Carex
sempervirens) şi coarnă mare (Sesleria bielzii) ………………………… 2405.3.1.13 Habitatul R3613 – Pajişti sud-est carpatice de Festuca carpatica,
Carduus kerneri şi Trisetum fuscum …………………………………….. 2425.3.1.14 Habitatul R3614 - Pajişti sud-est carpatice de Festuca xanthina ……… 244
5.3.2. Pajişti umede şi comunităţi de ierburi înalte (buruienişuri) ………… 2465.3.2.1. Habitatul R3708 – Comunităţi daco-getice cu Angelica sylvestris, Crepis
paludosa şi Scirpus sylvaticus …………………………………………… 2465.3.2.2. Habitatul R3710 – Pajişti dacice de Molinia coerulea …………………... 2475.3.3.3. Habitat R3712 – Comunităţi dacice cu Deschampsia caespitosa şi
Agrostis stolonifera ………………………………………………………. 2485.3.3. Pajişti mezofile …………………………………………………………. 250
5.3.3.1. Habitatul R3801 Pajişti sud-est carpatice de Trisetum flavescens şi Alchemilla vulgaris …………………………………………………... 250
5.3.3.2. Habitatul R3802 - Pajişti daco-getice de Arrhenatherum elatius ……... 2525.3.3.3. Habitatul R3803 – Pajişti sud-est carpatice de Agrostis capillaris şi
Festuca rubra ……………………………………………………………. 2535.3.3.4. Habitatul R3804 – Pajişti daco-getice de Agrostis capillaris şi
Anthoxanthum odoratum ………………………………………………… 2545.3.4. Pajişti xerice ……………………………………………………………. 256
5.3.4.1. Habitatul R3401 - Pajişti sud-est carpatice de Asperula capitata şi Sesleria rigida ……………………………………………………….. 256
5.3.4.2. Habitatul R3402 – Pajişti sud-est carpatice de Helictotrichon decorum 2585.3.4.3. Habitatul R3403 Paj. daco-getice de Festuca pallens şi Melica ciliata 260

6
VI. GOSPODĂRIREA HABITATELOR DE PAJIŞTI ………………….. 2626.1. Gospodărirea habitatelor cu valoare furajeră şi conservativă ………. 262
6.1.1. Pajişti alpine şi subalpine ………………………………………………... 2636.1.2. Pajişti umede ……………………………………………………………. 2656.1.3. Pajişti mezofile …………………………………………………………... 272
6.2. Gospodărirea habitatelor degradate de Nardus stricta ………………. 2806.2.1. Lucrări biopedoameliorative generale …………………………………… 2806.2.2. Lucrări de îmbunătăţire, întreţinere şi valorificare a producţiei …………. 2816.2.3. Producţia de lapte pe nardetele îmbunătăţite ……………………………. 287
VII. GRADIENŢI DE PRODUCTIVITATE A PAJIŞTILOR PE ALTITUDINE ………………………………………………………….. 292
7.1. Gradienţi climatici ……………………………………………………… 2927.1.1. Temperatura aerului şi solului …………………………………………… 2937.1.2. Precipitaţii atmosferice şi alţi factori climatici ………………………….. 299
7.2. Distribuţia altitudinală a ecosistemelor forestiere şi tipuri de pajişti 3007.3. Productivitatea primară a ecosistemelor montane pe niveluri
altitudinale ……………………………………………………………… 3027.4. Gradienţi de productivitate a pajiştilor montane îmbunătăţite ……... 306
7.4.1. Munţii Cindrel …………………………………………………………… 3067.4.2. Munţii Făgăraş …………………………………………………………… 3167.4.3. Munţii Bucegi ……………………………………………………………. 323
7.5. Prognoza productivităţii pajiştilor montane în contextul încălzirii globale a climei …………………………………………………………. 328
7.5.1. Evoluţia climatică şi consecinţele ei …………………………………….. 3287.5.2. Prognoza modificărilor bioclimatice şi ale solului montan ……………… 3307.5.3. Prognoza productivităţii pajiştilor montane ……………………………… 332
BIBLIOGRAFIE ……………………………………………………….. 334

7
CONTENT Pag. Content ...................................................................................................... 3 Introduction .............................................................................................. 11
I. RESEARCH ON THE MOUNTAIN GRASSLAND VEGETATION AND PASTURE STATIONS .................................................................. 13
1.1. Brief History .............................................................................................. 131.2. Classification scheme of pastures and mountain grassland habitat types 171.3. Stationary conditions of experimental fields for improving and using
the pastures .............................................................................................. 28II. DESCRIPTION OF DEGRADED Nardus stricta GRASSLANDS
OF BRASOV COUNTY ......................................................................... 322.1. Natural conditions .................................................................................... 32
2.1.1. The geographical and terrain conditions .................................................... 32 2.1.2. Geology ...................................................................................................... 33
2.1.3. Climate ....................................................................................................... 352.1.4. Soil .............................................................................................................. 382.1.5. Aspects of the flora and vegetation ............................................................ 41
2.2. Research objectives and working methods ............................................ 432.2.1. General principles ...................................................................................... 442.2.2. Geo botany study ....................................................................................... 452.2.3. Typological study ....................................................................................... 542.2.4. Conditional evaluation of Nardus stricta areas .......................................... 56
2.3. Ecology and floristic composition of Nardus stricta areas .................... 582.3.1. Ecology of Nardus stricta L. Species ........................................................ 592.3.2. Some data on the structure of the floristic composition of Nardus stricta
areas ............................................................................................................ 702.4. Description of Nardus stricta L. associations .......................................... 72
2.4.1. Alpigenum Nardetum association ............................................................... 762.4.2. Nardo - Campanuletum (abietinae) association ......................................... 812.4.3. Nardo - Juncetum (conglomerates) association ......................................... 922.4.4. Sinecological indices of Nardus stricta areas ........................................... 111
2.5. Description of Nardus stricta area types ................................................ 1132.5.1. Subalpine Nardus stricta area situated on great slopes or steep conditions 1132.5.2 Subalpine Nardus stricta area situated on flat or slightly to the middle
slope ........................................................................................................... 1142.5.3. Nardus stricta area with various species of Vaccinium ............................. 1142.5.4. Mountain Nardus stricta area with Festuca rubra and Agrostis capillaris
species situated on very strong slopes or steep slopes ............................... 1152.5.5. Mountain Nardus stricta area with Festuca rubra and Agrostis capillaris
species situated on very low to medium slopes ........................................ 115

8
2.5.6. Hilly Nardus stricta area with different mesophilic and xeromezophilic
species situated on medium to very strongly inclined slopes ................... 1162.5.7. Nardus stricta area of depression, with different species and hydrophilic
mezohygrophile located on flat or gently sloping land .............................. 1162.6. Other descriptions of Nardus stricta area ............................................... 117
2.6.1. Nardus stricta area conditional evaluation ................................................. 1172.6.2. Relationship between phytocoenosis, types and conditional evaluation ... 119
III. ASPECTS OF ECOLOGY PASTURE FROM MOUNTAIN AREA 1223.1. The main ecological factors that influence the structure and
dynamics of grassland habitats ............................................................... 1223.1.1. Topographic factors .................................................................................... 1233.1.2. Edaphic factors ........................................................................................... 1253.1.3. Climatic factors .......................................................................................... 1353.1.4. Biotic factors .............................................................................................. 136
3.2. Phyto diversity of grassland habitats from the mountain area .......... 1403.3. Potential Threats ...................................................................................... 145IV. THE SOILS OF THE MOUNTAIN GRASSLANDS ........................... 1514.1. Soils in the alpine grassland zone ............................................................... 1524.2. Soils in the mountain pine zone(sub alpine) ............................................... 1564.3. Soils in the spruce zone (boreal) ................................................................ 1584.4. Soils in the hardwoods zone (nemoral) ...................................................... 166
4.5. General characteristics of soils ................................................................ 1874.5.1. Soils covering of the Eastern Carpathians .................................................. 1874.5.2. Soil covering of the Southern Carpathians ............................................... 1914.5.3. Soil covering of the Western Carpathians .................................................. 193
4.6. Research results of the observation key locations ................................. 195V. MAIN TYPES OF GRASSLAND HABITATS IN THE MOUNTAIN
AREA ....................................................................................................... 2165.1. Arguments for biodiversity conservation ............................................... 2165.2. Methodological Considerations ............................................................... 2175.3. Description of main types of grassland habitats in the mountain area
of Romania ............................................................................................... 2185.3.1. Alpine and subalpine grasslands ................................................................ 218
5.3.1.1. R3601 habitat - south-east Carpathian grasslands of dwarf sedge (Kobresia myosuroides) and Oxytropis carpatica ...................................... 218
5.3.1.2. R3602 habitat - south-east
8
2.5.6. Hilly Nardus stricta area with different mesophilic and xeromezophilic
species situated on medium to very strongly inclined slopes ................... 1162.5.7. Nardus stricta area of depression, with different species and hydrophilic
mezohygrophile located on flat or gently sloping land .............................. 1162.6. Other descriptions of Nardus stricta area ............................................... 117
2.6.1. Nardus stricta area conditional evaluation ................................................. 1172.6.2. Relationship between phytocoenosis, types and conditional evaluation ... 119
III. ASPECTS OF ECOLOGY PASTURE FROM MOUNTAIN AREA 1223.1. The main ecological factors that influence the structure and
dynamics of grassland habitats ............................................................... 1223.1.1. Topographic factors .................................................................................... 1233.1.2. Edaphic factors ........................................................................................... 1253.1.3. Climatic factors .......................................................................................... 1353.1.4. Biotic factors .............................................................................................. 136
3.2. Phyto diversity of grassland habitats from the mountain area .......... 1403.3. Potential Threats ...................................................................................... 145IV. THE SOILS OF THE MOUNTAIN GRASSLANDS ........................... 1514.1. Soils in the alpine grassland zone ............................................................... 1524.2. Soils in the mountain pine zone(sub alpine) ............................................... 1564.3. Soils in the spruce zone (boreal) ................................................................ 1584.4. Soils in the hardwoods zone (nemoral) ...................................................... 166
4.5. General characteristics of soils ................................................................ 1874.5.1. Soils covering of the Eastern Carpathians .................................................. 1874.5.2. Soil covering of the Southern Carpathians ............................................... 1914.5.3. Soil covering of the Western Carpathians .................................................. 193
4.6. Research results of the observation key locations ................................. 195V. MAIN TYPES OF GRASSLAND HABITATS IN THE MOUNTAIN
AREA ....................................................................................................... 2165.1. Arguments for biodiversity conservation ............................................... 2165.2. Methodological Considerations ............................................................... 2175.3. Description of main types of grassland habitats in the mountain area
of Romania ............................................................................................... 2185.3.1. Alpine and subalpine grasslands ................................................................ 218
5.3.1.1. R3601 habitat - south-east Carpathian grasslands of dwarf sedge (Kobresia myosuroides) and Oxytropis carpatica ...................................... 218
5.3.1.2. R3602 habitat - south-east Carpathian grasslands of Carex curvula and Primula minima ......................................................................................... 220
5.3.1.3. R3603 habitat - south-east Carpathian grasslands of pig hair (Juncus trifidus) and Oreochloa disticha ............................................................... 222
5.3.1.4. R3604 habitat - south-east Carpathian grasslands of Festuca airoides and

9
Potentilla aurea ssp chrysocraspeda .........................................................
224
5.3.1.5. R3605 habitat–south east Carpathian grasslands corner fescue (Festuca versicolor), and Sesleria rigida ssp haynaldiana ....................................... 226
5.3.1.6. R3606 habitat – south east Carpathian grasslands of rocks fescue (Festuca saxatilis) ....................................................................................... 228
5.3.1.7. R3607 habitat - south-east Carpathian grasslands of Festuca amethystina and Dianthus tenuifolius ............................................................................. 230
5.3.1.8. R3608 habitat - Carpathian southeast grasslands of Scorzonera purpurea ssp rosea and Festuca nigrescens ............................................................. 232
5.3.1.9. R3609 habitat - south-east Carpathian grasslands of Nardus stricta and Viola declinata ............................................................................................ 234
5.3.1.10 R3610 habitat - south-east Carpathian grasslands of Poa media ................ 2365.3.1.11 R3611 habitat - south-east Carpathian grasslands of tail rabbit (Sesleria
rigida subsp haynaldiana) and sedge (Carex sempervirens) ..................... 2385.3.1.12 R3612 habitat - south-eastern Carpathian grasslands of sedge (Carex
sempervirens) and large horns (Sesleria bielzii) ........................................ 2405.3.1.13 R3613 habitat - south-east Carpathian grasslands of Festuca carpatica ,
Carduus Kerneri and Trisetum fuscum ..................................................... 2425.3.1.14 R3614 habitat - southeastern Carpathian grasslands of Festuca xanthina 244
5.3.2. Wet grasslands and tall herb communities (weedings) ......................... 2465.3.2.1. Habitat R3708 - Communities Daco-getics with Angelica sylvestris,
Crepis paludosa and Scirpus sylvaticus ..................................................... 2465.3.2.2. R3710 habitat - Dacian grasslands of Molinia coerulea ............................ 2475.3.3.3. Habitat R3712 - Dacian communities with Deschampsia caespitosa and
Agrostis stolonifera .................................................................................... 2485.3.3. Mesophile grasslands ................................................................................ 250
5.3.3.1. R3801 habitat- south-east Carpathian grassland of Trisetum flavescens and Alchemilla vulgaris .............................................................................. 250
5.3.3.2. R3802 habitat - Daco-getics grasslands of Arrhenatherum elatius ........... 2525.3.3.3. R3803 habitat - south-east Carpathian grasslands of Agrostis capillaris
and Festuca rubra ...................................................................................... 2535.3.3.4. R3804 habitat - Daco-getic grasslands of Agrostis capillaris and
Anthoxanthum odoratum ........................................................................... 2545.3.4. Xerice pastures ........................................................................................... 256
5.3.4.1. R3401 habitat - south-east Carpathian grasslands of Asperula capitata and Sesleria rigida ..................................................................................... 256
5.3.4.2. R3402 habitat - south-east Carpathian grasslands of Helictotrichon decorum ..................................................................................................... 258
5.3.4.3. R3403 habitat- Daco-getic grassland of Festuca pallens and Melica ciliata .......................................................................................................... 260

10
VI. MANAGEMENT OF GRASSLAND HABITATS ................................ 2626.1. Management of habitats with forage and conservation value ............ 262
6.1.1. Alpine and subalpine grasslands ................................................................. 2636.1.2. Wet pastures ............................................................................................... 2656.1.3. Mesophile grasslands .................................................................................. 272
6.2. Management of degraded Nardus stricta habitats ................................. 2806.2.1. General bio pedo ameliorative Works ........................................................ 2806.2.2. Improvement works, maintenance and utilization of production ............... 2816.2.3. Milk production on improved Nardus stricta species area ........................ 287
VII. GRADIENTS OF GRASSLAND PRODUCTIVITY RELATED TO ALTITUDINAL LEVEL ......................................................................... 292
7.1. Climatic gradients ..................................................................................... 2927.1.1. Air and soil temperature ............................................................................. 2937.1.2. Rainfall and other climatic factors .............................................................. 299
7.2. Altitudinal distribution of forest types and grassland ecosystems ....... 3007.3. Primary productivity of mountainous ecosystems related to the
altitude levels ............................................................................................. 3027.4. Productivity gradients of mountain improved grasslands .......................... 306
7.4.1. The Cindrel Mountains ............................................................................... 3067.4.2. The Fagaras Mountains ............................................................................... 3167.4.3. The Bucegi Mountains ............................................................................... 323
7.5. Productivity prediction of mountain pastures in the context of global climate warming ....................................................................................... 328
7.5.1. The evolution of climate and its consequences .......................................... 3287.5.2. Prediction of changes in mountain soil and bioclimatic ............................ 3307.5.3. Prediction of the mountain pastures productivity ...................................... 332
BIBLIOGRAPHY ..................................................................................... 334

11
INTRODUCERE
Pajiştile montane din România, situate între 600 – 800 m altitudine până la 2.544m pe Vf. Moldoveanu din Munţii Făgăraş, au o suprafaţă de cca. 2,8 milioane hectare, reprezentând aproximativ 56 % din patrimoniul pastoral al ţării.
Cu excepţia pajiştilor din etajul alpin, al stâncăriilor şi mlaştinilor, vegetaţia pajiştilor montane din Carpaţi este de origine secundară, fiind un produs al intervenţiei omului şi animalelor sale, după defrişarea pădurii. Astfel, influenţa antropică este determinantă în evoluţia vegetaţiei ierboase şi a productivităţii ei.
Stadiul actual al păşunilor şi fâneţelor montane din ţara noastră este departe de situaţia normală din ţările europene dezvoltate, atât din punct de vedere al cunoaşterii lor, cât şi al gospodăririi (managementului) pentru a realiza în mod eficient produse animaliere încadrate în normele de siguranţă alimentară.
Alături de rolul decisiv al pajiştilor pentru hrana animalelor ierbivore, trebuie să se ţină cont şi de importanţa acestor suprafeţe pentru protecţia mediului şi dezvoltării rurale, prin îmbunătăţirea fertilităţii solului şi prevenirea eroziunii, alunecărilor şi inundaţiilor, conservarea biodiversităţii şi a frumuseţii peisajelor şi asigurarea unui potenţial îmbunătăţit de sechestrare a carbonului în sol.
Suprapăşunatul, subîncărcarea cu animale până la abandon, împreună cu lipsa de îmbunătăţire şi întreţinere permanentă sunt extrem de dăunătoare din toate punctele de vedere, îndeosebi în zona montană, cu ecosisteme mai fragile.
Pentru îndreptarea acestei situaţii sunt necesare studii preliminare asupra covorului ierbos şi cercetări ştiinţifice riguroase asupra posibilităţilor de îmbunătăţire şi folosire raţională a pajiştilor în toate componentele ecosistemului praticol, climă – sol – plantă – animal – om – mediu.
Cercetările din acest domeniu au fost focalizate pe stabilirea unor soluţii tehnice care să permită o interacţiune favorabilă între sistemele de cultură a plantelor şi creşterea animalelor cu resursele naturale din zona respectivă.
Fără a epuiza subiectul, în tratatul de faţă se încearcă abordarea ştiinţifică a problemei pajiştilor pe mai multe planuri.
După ce se prezintă studiile asupra vegetaţiei pajiştilor din ultimele opt decenii şi ale condiţiilor staţionale din câmpurile experimentale din ultima jumătate de secol, se tratează în mod unitar, ca exemplificare, pajiştile degradate de Nardus stricta din judeţul Braşov.
În continuare, se redau câteva aspecte privind ecologia pajiştilor din zona montană, având la bază ultimele noutăţi în acest domeniu pe relaţia sol – plantă.
Un interes aparte s-a acordat caracteristicilor solurilor din pajiştile montane, împreună cu unele precizări privind denumirile folosite în ultima perioadă şi o schemă – cheie de îmbunătăţire a factorilor de degradare a terenurilor.

12
În literatura noastră de specialitate a lipsit până acum o descriere mai amplă a habitatelor de pajişti după modelul ţărilor din UE şi al răspândirii lor în Carpaţii noştri. Acest neajuns este acum completat împreună cu câteva principii de gospodărire a habitatelor de pajişti.
Problemele legate de îmbunătăţirea şi folosirea raţională a pajiştilor sunt prezentate mai larg în „Ghidul de producere ecologică a furajelor de pajişti montane” apărut simultan cu prezentul tratat în aceeaşi generoasă editură.
În ultima parte sunt prezentaţi gradienţii altitudinali de productivitate a pajiştilor cu exemplificări din Munţii Cindrel, Făgăraş şi Bucegi, unde s-au acumulat mai multe date de cercetare ştiinţifică de pratologie şi pratotehnie.
Înainte de final se prezintă o predicţie a dinamicii vegetaţiei şi productivităţii pajiştilor la nivelul anilor 2070, în cazul încălzirii globale a climei în medie cu 3 0C în următorii ani.
Din enumerarea conţinutului tratatului se constată o oarecare lipsă de coeziune între componente de care suntem conştienţi şi noi.
Într-o oarecare măsură la această situaţie a contribuit şi reducerea fondurilor de cercetare din ultimii doi ani ai proiectului care nu ne-au permis întregirea cercetărilor şi atingerea obiectivelor conform planului iniţial.
Cu toate aceste neajunsuri, autorii speră ca acest tratat să aducă o contribuţie importantă la cunoaşterea covorului ierbos al pajiştilor montane şi la gospodărirea judicioasă a lui.
Folosim acest prilej pentru a mulţumi specialiştilor din MADR care au propus şi finanţat acest proiect, specialiştilor din cercetarea ştiinţifică din reţeaua ASAS, Academia Română, universităţile agricole şi nu în ultimul rând Universităţii Transilvania din Braşov, care a asigurat editarea cărţii.
Sperăm ca într-o ediţie viitoare să lărgim şi să întregim problematica acestui tratat de pajişti montane, fiind convinşi de importanţa acestei iniţiative.
Orice sugestie în această direcţie va fi bine primită de autori cu mulţumiri anticipate.
Autorii

13
I. CERCETĂRI PRIVIND VEGETAŢIA PAJIŞTILOR MONTANE ŞI ALE STAŢIUNILOR
1.1. SCURT ISTORIC
Primele scrieri asupra plantelor şi vegetaţiei de pe teritoriul ţării noastre au fost
făcute în anul 1578 de către MELIUS, un botanist care a publicat la Cluj o Botanică medicală după care au urmat numeroşi alţii până în zilele noastre. După BORZA şi BOŞCAIU (1965) completaţi de ŢUCRA şi NEACŞU (1988) în studiul vegetaţiei României s-au derulat următoarele etape:
I. etapa floristică descriptivă 1578 – 1863; II. etapa fitogeografică 1863 – 1918; III. etapa fitocenologică 1918 – 1936; IV. etapa fitocenologică – ecologică 1936 – 1960; V. etapa ecologică după 1960. Începutul fiecărei etape poartă pecetea unei lucrări de referinţă care se
individualizează prin idei şi metode noi admise că au adus un progres evident în studiul vegetaţiei. S-a amintit începutul etapei I-a în anul 1578, urmat de etapa a II-a marcată de lucrarea lui KERNER VON MARILAUN “Das Phlanzenleben der Donauländer” apărută în 1863 care leagă pentru prima dată vegetaţia de mediul ei.
Începând cu etapa a II-a fitocenologică se începe propriu zis studiul vegetaţiei după definirea asociaţiei vegetale ca unitate fundamentală la al 3-lea Congres de Botanică de la Bruxelles (1910). Principiile fitosociologiei elaborate de BRAUN – BLANQUET, au fost adoptate şi dezvoltate de BORZA în “Vegetaţia şi flora României” (1926) unde în Carpaţi se diferenţiază 4 etaje de vegetaţie (etajul fagului, molidului, subalpin şi alpin) la care se adaugă “Flora şi vegetaţia Munţilor Retezat” (1934), “Munţii Semenic” (1946), “Munţii Sebeşului” (1951) şi altele.
Etapa a IV-a fitocenologică – ecologică este prima etapă în care vegetaţia pajiştilor sunt studiate şi din punct de vedere agronomic, deschizător de drum fiind de această dată prof. SAFTA (1936) cu lucrarea “Contribuţii la studiul fitosocial agricol al fâneţelor din judeţul Cluj” în care introduce termenul “tip de fâneaţă” şi mai târziu în alte lucrări şi cel de “tip de păşune”. În această etapă se fac primele studii în relaţia staţiune – vegetaţie şi de apreciere a valorii economice a pajiştilor.
Etapa a V-a, ecologică debutează în 1960 cu lucrarea “Geobotanica aplicată la studiul pajiştilor şi al terenurilor cultivate” întocmită de prof. ANGHEL care a punctat în linii generale obiectivele şi a jalonat calea de urmat în această nouă etapă pe care le cităm în continuare:
pentru cunoaşterea temeinică a vegetaţiei pajiştilor sunt necesare, în primul rând, cercetări staţionale de lungă durată;
studiul vegetaţiei trebuie făcut în strânsă legătură cu condiţiile staţionale, în care scop geobotaniştii vor colabora cu alţi specialişti precum pedologi, climatologi, pratologi, silvicultori, zootehnişti, etc.;

14
pentru clasificarea pajiştilor şi pentru stabilirea măsurilor de îmbunătăţire sunt necesare în primul rând hărţi de vegetaţie la scară mare;
trebuie făcută o diferenţiere între asociaţia vegetală şi tipul de pajişte, prin tipul de pajişte netrebuind să se înţeleagă numai vegetaţia ci şi condiţiile staţionale;
clasificarea vegetaţiei pajiştilor numai după criterii floristice nu este satisfăcătoare pentru nevoile producţiei;
nomenclatura unităţilor în clasificările tipologice trebuie să cuprindă atât aspectul fitocenologic cât şi cel staţional.
Între etapele III şi V respectiv fitocenologică până la cea ecologică, nu se pot face distincţii clare întrucât multe din aceste studii continuă şi astăzi având ca obiective şi metodologie de lucru asemănătoare cu iniţiatorii menţionaţi mai înainte.
În plus pentru studiul vegetaţiei pajiştilor se remarcă o privire mai largă asupra rolului multifuncţional al acestora în ecosistemele terestre.
Prin apariţia lucrării “Habitatele din România” de DONIŢĂ şi colab. (2005) se poate considera că a început o nouă etapă a VI-a în studiul vegetaţiei ca parte componentă determinantă a unui habitat. Denumirea habitatelor s-a făcut într-o concepţie unitară, prin utilizarea a trei elemente definitorii pentru habitate naturale şi anume:
unitatea fitocenologică mare (păduri, tufărişuri, pajişti, comunităţi de mlaştini şi terenuri înmlăştinate, de stâncării, de dune de nisip, de ape etc.);
localizarea în domeniul geographic (sud-est carpatic, dacic, getic, moldav, panonic, danubian, vest-pontic, ponto-sarmatic, balcanic);
speciile de plante definitorii pentru tipul de habitat. Habitatele au fost descrise după următoarea schemă:
Cod şi denumire; Corespondenţa cu NATURA 2000, EMERALD, CORINE,
PALAEARCTIC HABITATS, EUNIS; Corespondenţa cu asociaţiile vegetale iar pentru păduri şi cu tipurile
de ecosisteme forestiere; Răspândirea în România; Suprafeţe aproximative; Staţiuni: altitudinea, clima, relief, roci, soluri; Structura biocenozei; Compoziţia floristică; Valoarea de conservare; Literatura utilizată; Redactorul habitatului.
La această nouă etapă şi metoda de lucru utilizată urmează să se studieze mai mult şi vegetaţia pajiştilor montane, pentru a ne conecta la sistemul de clasificare a habitatelor din Uniunea Europeană.

15
Cele mai importante sinteze privind pajiştile montane în ordinea apariţiei au fost următoarele: PUŞCARU, D. şi colab (1956) Păşunile alpine din Munţii Bucegi, Ed.
Academiei Române, Bucureşti, în care în prima parte se face un studiu geobotanic şi agrozootehnic şi în partea a II-a experienţe agrozootehnice în vederea ridicării productivităţii păşunilor alpine, fiind primul tratat românesc despre pajiştile montane.
BUIA, AL., SAFTA, I., PAVEL, C. şi colab (1962), Pajiştile din Masivul Parâng şi îmbunătăţirea lor, Ed. Agro-Silvică, Bucureşti, în care sunt prezentate la fel ca în precedenta lucrare o descriere amănunţită a condiţiilor naturale (geologie, geomorfologie, hidrologie, climă, soluri, vegetaţie primară, etc.) clasificarea vegetaţiei pajiştilor şi posibilităţi de îmbunătăţire a lor, fiind unul din cele mai complexe şi complete lucrări de acest gen din literatura noastră ştiinţifică din acest domeniu.
PUŞCARU-SOROCEANU Evdochia şi colab.(1963) Păşunile şi fâneţele din R.P.Română, Studiu geobotanic şi agroproductiv, Ed. Academiei Române, Bucureşti, în care se face pentru prima dată o prezentare mai cuprinzătoare a vegetaţiei pajiştilor montane din Carpaţii României încadrate la formaţiunile “Păşuni şi fâneţe de munte (800-1600-1800 m altitudine)” şi “Păşuni alpine” (1600-1800 – 2543 m altitudine), la care sunt prezentate suprafeţele şi repartiţia lor, aprecierea economică după producţia şi valoarea nutritivă precum şi căile şi perspective de sporire a productivităţii acestora cu recomandări practice.
RESMERIŢĂ, I.(1970) Flora, vegetaţia şi potenţialul productive pe Masivul Vlădeasa, Ed. Academiei Române, Bucureşti, a duce o contribuţie importantă la cunoaşterea vegetaţiei pajiştilor şi dinamica ei după despădurire, completată cu măsuri de îmbunătăţire a productivităţii lor.
PAVEL, C. şi colab. (1973), Pajiştile din zona subcarpatică a Olteniei, Ed. Scrisul Românesc, Craiova, în care se face o descriere complexă staţiune-vegetaţie cu posibilităţi concrete de îmbunătăţire a lor în această parte a ţării.
ANGHEL, GH., BĂRBULESCU, C., BĂRSAN A. şi colab. (1977) Pajiştile din Platforma Bran, Casa Agronomului Braşov, lucrare de referinţă pentru modul cum trebuiesc întocmite studiile monografice asupra vegetaţiei pajiştilor dintr-un areal clar delimitat între Masivele Bucegi şi Piatra Craiului, cu hărţile geologice, geomorfologice, hidrografice, izoterme, izohiete, harta solurilor şi profile geobotanice, după care sunt descrie tipurile de pajişti cu valoarea lor pastorală şi metoda de sporire a producţiei.
SAMOILĂ, Z. şi colab (1979) Pajiştile din Banat, Sporirea producţiei şi îmbunătăţirea calităţii lor, Redacţia de Propagandă Tehnică Agricolă, Bucureşti, unde se prezintă pentru Munţii Banatului, o hartă a raionării pedoclimatice, tipurile de pajişti montane şi tehnologii de sporire a producţiei şi calităţii pajiştilor pe baza unor experimentări riguroase în staţionarele din Munţii Semenicului şi Ţarcu-Godeanu.

16
MARUŞCA, T. (1982) Studiul geobotanic şi tipologic al nardetelor din judeţul Braşov, IANB Bucureşti, în care se prezintă fitocenozele şi tipurile de pajişti degradate dominate de Nardus stricta, de la 480 m altitudine din Depresiunea Făgăraşului până la 2210 m în Gaura de sub Vf. Omu din Masivul Bucegi, cu posibilităţi de îmbunătăţire a lor în staţionarele de la Vlădeni-Braşov şi Blana-Bucegi.
BĂRBULESCU, C., MOTCĂ, Gh. (1983) Păşunile munţilor înalţi, Ed. Ceres, Bucureşti, care se prezintă ca un veritabil ghid de recunoaştere a plantelor din păşunile montane, urmate de principalele tipuri de păşuni şi metode de ameliorare şi folosire a lor, bazaţi pe o vastă experienţă dobăndită în Carpaţii Meridionali (Iezer-Păpuşa, Leaota, Bucegi, etc.).
CIUCĂ Maria (1984) Flora şi vegetaţia pajiştilor din Munţii Ciucaş, Ed. Academiei Române, Bucureşti, este un exemplu de urmat pentru studiul covorului ierbos al unui munte, cu flora şi asociaţiile de pajişti, urmată de o hartă sugestivă a lor.
ŢUCRA, I. şi colab (1987) Principalele tipuri de pajişti din România, Redacţia de Propagandă Tehnică Agricolă, Bucureşti, lucrare de referinţă în care enunţă şi se dezvoltă principiile de bază în tipizarea pajiştilor cu sistematica şi criteriile de stabilire a unităţilor tipologice, nomenclatura şi elemente de caracterizare a tipurilor de pajişti, conspectul principalelor tipuri de pajişti din România, inclusiv cele montane, categorii de staţiuni cu pajişti şi măsuri tehnologice de îmbunătăţire.
MOTCĂ,Gh., OANCEA, I., GEAMĂNU Lidia Ivona (1994) Pajiştile României, Tipologie şi tehnologie, Ed. Tehnică Agricolă, Bucureşti, este una din cele mai complexe lucrări din domeniu care cuprinde principalele tipuri de pajişti, inclusiv cele montane, cu metode de îmbunătăţire şi folosire a lor, urmate de tehnologii diferenţiate pe tipuri de pajişti.
POPOVICI, D. şi colab (1996) Pajiştile din Bucovina, Ed. “Helios” prezintă condiţiile naturale şi răspândirea pajiştilor în această provincie în care zona montană este bine reprezentată, flora şi vegetaţia pajiştilor urmate de tipurile de pajişti la care sunt prezentate pentru fiecare tip rezultate experimentale privind măsurile de creştere a producţiei.
IONESCU, I. şi colab.(2001) Pajiştile permanente din nordul Olteniei, Ed. “Universitaria” Craiova, aduce ultimele noutăţi asupra caracterizării geografice a spaţiului subcarpatic şi carpatic din această provincie istorică cu principalele soluri, flora şi vegetaţia pajiştilor montane cu îmbunătăţirea lor prin fertilizare cu îngrăşăminte chimice şi organice cât şi valorificarea lor prin păşunat, fiind un model de urmat pentru cercetarea complexă a pajiştilor dintr-o regiune muntoasă mai întinsă.
SÂRBU Anca, VEEN, P. şi colab.(2004) Grasslands of Romania, Final report on National Grasslands Inventary 2000-2003, Proiect PIN-MATRA, Bucureşti, face pentru prima oară la noi o estimare a biodiversităţii pajiştilor din ţara noastră

17
cu identificarea alianţelor fitocenologice şi a speciilor ameninţate la nivel global, european şi naţional, la care coordonatorul prezentului tratat a participat direct ca responsabil al grupului de lucru de la Braşov pentru centrul ţării, unde s-au studiat pajiştile montane din Bucegi, Făgăraş şi Perşani din Carpaţii Meridionali.
Acestea au fost principalele 15 lucrări de sinteză privind vegetaţia pajiştilor din care
5 pe ţară, 5 pe zone fizico-geografice (Oltenia – 2, Bucovina, Banat şi Jud. Braşov) şi 5 pe masivii muntoşi (Bucegi, Parâng, Vlădeasa, Platforma Bran şi Ciucaş) la care se adaugă alte sute de lucrări cu titluri bibliografice din acest domeniu.
1.2. SCHEME DE CLASIFICARE A PAJIŞTILOR ŞI TIPURILE DE HABITATE DE PAJIŞTI MONTANE Începând cu primele clasificări a pajiştilor montane (SAFTA, 1943) până mai recent
(ŢUCRA, NEACŞU, 1984) au fost elaborate mai multe scheme din ce în ce mai perfecţionate, care au ţinut cont mai mult la altitudine, etajarea vegetaţiei forestiere , condiţii staţionale şi alte criterii.
Astfel, în tabelul 1.1. este prezentată o primă clasificare după distribuţia geografică şi tipurile de păşuni după SAFTA (1943) cu o conturare exactă a păşunilor alpine din Carpaţii Meridionali.
Tabelul 1.1.
Clasificarea şi distribuţia geografică a păşunilor montane din Transilvania Classification and geographic distribution of mountainous pastures from
Transylvania ( SAFTA , 1943)
Categorii Tipuri de păşuni
Păşunile etajului forestier şi montan
1. Tipul Agrostis tenuis-Cynosurus cristatus 2. Tipul Festuca rubra
Păşunile degradate ale etajului forestier şi montan
1. Tipul Nardus stricta ( climax) 2. Tinoavele
Păşunile alpine 1. Tipul Festuca supina-Agrostis rupestris f. frecvent între 1800 şi 2100 m ( zona de jnepeniş)
2. Tipul Carex curvula > 2100m
Cercetările efectuate de BURDUJA (1956) în Carpaţii Orientali ajung la o etajare (zonare) apropiată cu cea precedentă, în plus face o echivalenţă între tipul de pajişte şi asociaţiile de bază cu cele secundare (Tabelul 1.2).
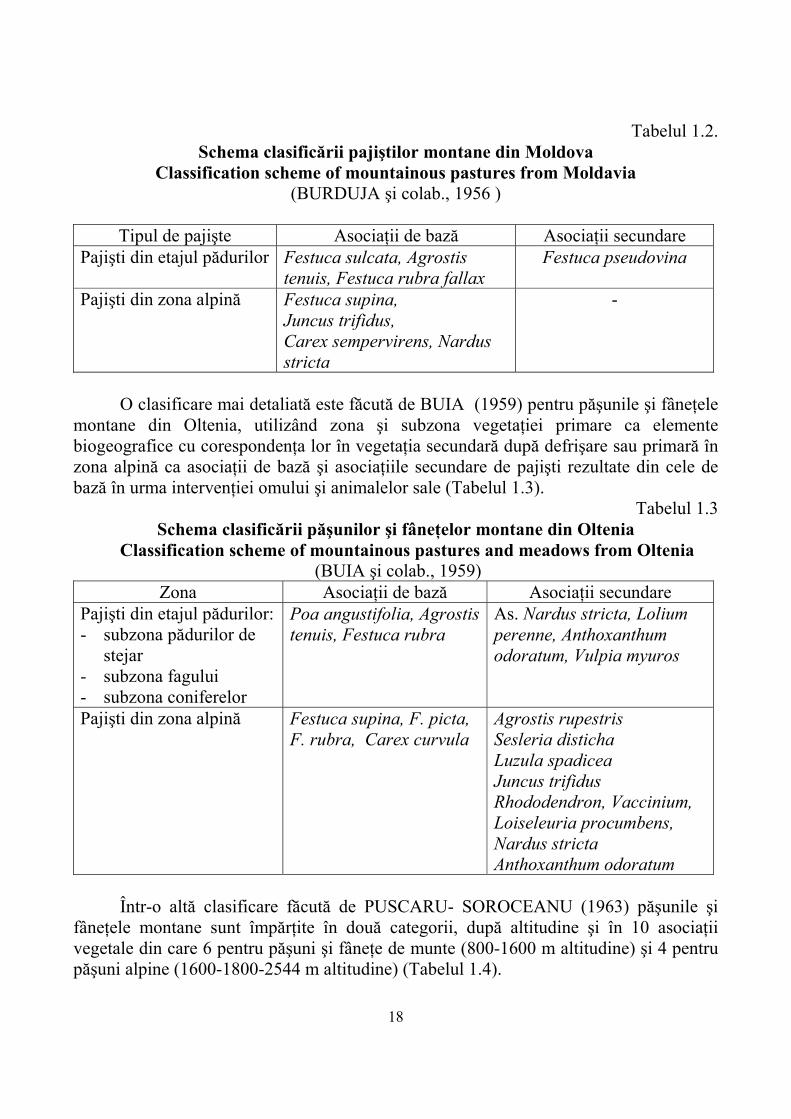
18
Tabelul 1.2. Schema clasificării pajiştilor montane din Moldova
Classification scheme of mountainous pastures from Moldavia (BURDUJA şi colab., 1956 )
Tipul de pajişte Asociaţii de bază Asociaţii secundare
Pajişti din etajul pădurilor Festuca sulcata, Agrostis tenuis, Festuca rubra fallax
Festuca pseudovina
Pajişti din zona alpină Festuca supina, Juncus trifidus, Carex sempervirens, Nardus stricta
-
O clasificare mai detaliată este făcută de BUIA (1959) pentru păşunile şi fâneţele montane din Oltenia, utilizând zona şi subzona vegetaţiei primare ca elemente biogeografice cu corespondenţa lor în vegetaţia secundară după defrişare sau primară în zona alpină ca asociaţii de bază şi asociaţiile secundare de pajişti rezultate din cele de bază în urma intervenţiei omului şi animalelor sale (Tabelul 1.3).
Tabelul 1.3 Schema clasificării păşunilor şi fâneţelor montane din Oltenia
Classification scheme of mountainous pastures and meadows from Oltenia (BUIA şi colab., 1959)
Zona Asociaţii de bază Asociaţii secundare Pajişti din etajul pădurilor: - subzona pădurilor de
stejar - subzona fagului - subzona coniferelor
Poa angustifolia, Agrostis tenuis, Festuca rubra
As. Nardus stricta, Lolium perenne, Anthoxanthum odoratum, Vulpia myuros
Pajişti din zona alpină Festuca supina, F. picta, F. rubra, Carex curvula
Agrostis rupestris Sesleria disticha Luzula spadicea Juncus trifidus Rhododendron, Vaccinium, Loiseleuria procumbens, Nardus stricta Anthoxanthum odoratum
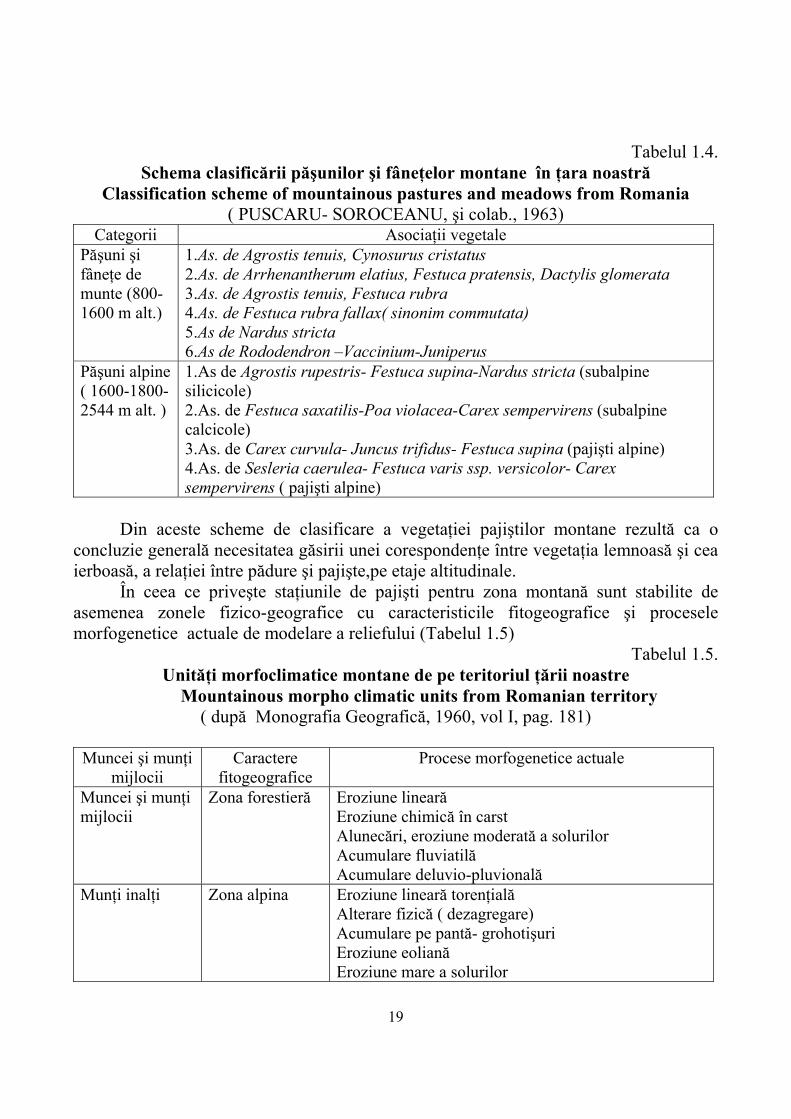
Într-o altă clasificare făcută de PUSCARU- SOROCEANU (1963) păşunile şi fâneţele montane sunt împărţite în două categorii, după altitudine şi în 10 asociaţii vegetale din care 6 pentru păşuni şi fâneţe de munte (800-1600 m altitudine) şi 4 pentru păşuni alpine (1600-1800-2544 m altitudine) (Tabelul 1.4).
19
Tabelul 1.4.
Schema clasificării păşunilor şi fâneţelor montane în ţara noastră Classification scheme of mountainous pastures and meadows from Romania
( PUSCARU- SOROCEANU, şi colab., 1963) Categorii Asociaţii vegetale
Păşuni şi fâneţe de munte (800-1600 m alt.)
1.As. de Agrostis tenuis, Cynosurus cristatus 2.As. de Arrhenantherum elatius, Festuca pratensis, Dactylis glomerata 3.As. de Agrostis tenuis, Festuca rubra 4.As. de Festuca rubra fallax( sinonim commutata) 5.As de Nardus stricta 6.As de Rododendron –Vaccinium-Juniperus
Păşuni alpine ( 1600-1800-2544 m alt. )
1.As de Agrostis rupestris- Festuca supina-Nardus stricta (subalpine silicicole) 2.As. de Festuca saxatilis-Poa violacea-Carex sempervirens (subalpine calcicole) 3.As. de Carex curvula- Juncus trifidus- Festuca supina (pajişti alpine) 4.As. de Sesleria caerulea- Festuca varis ssp. versicolor- Carex sempervirens ( pajişti alpine)
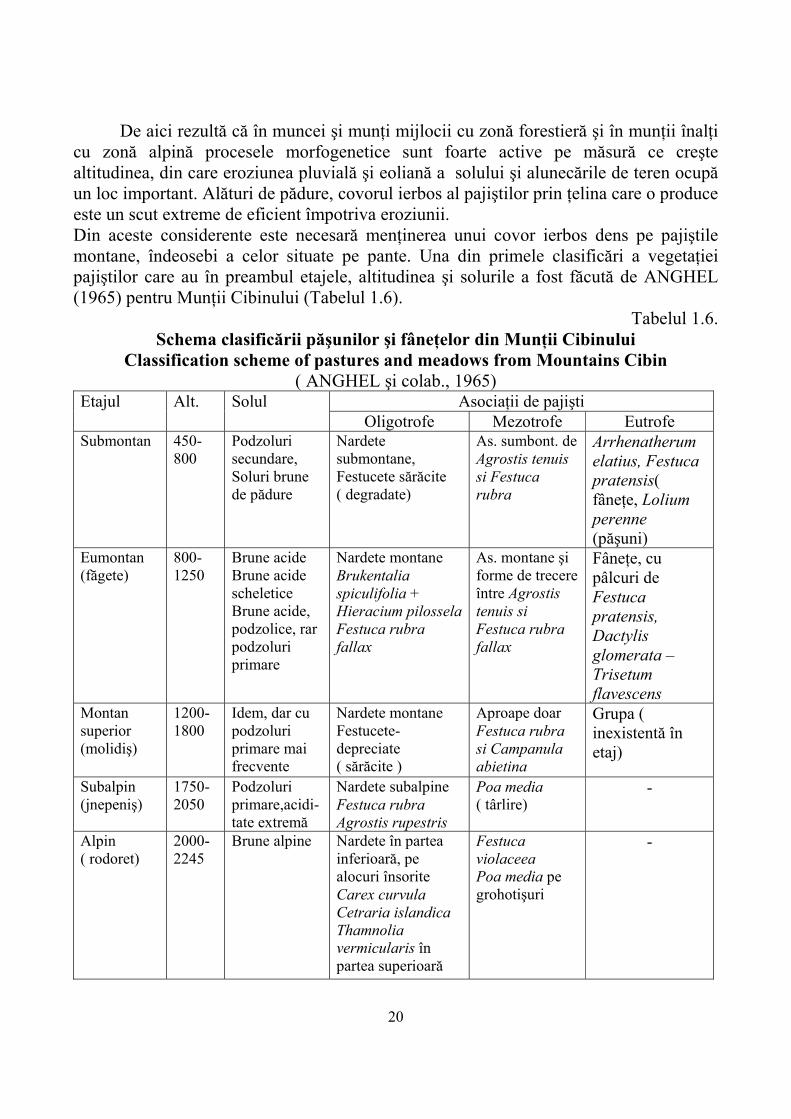
Din aceste scheme de clasificare a vegetaţiei pajiştilor montane rezultă ca o concluzie generală necesitatea găsirii unei corespondenţe între vegetaţia lemnoasă şi cea ierboasă, a relaţiei între pădure şi pajişte,pe etaje altitudinale. În ceea ce priveşte staţiunile de pajişti pentru zona montană sunt stabilite de asemenea zonele fizico-geografice cu caracteristicile fitogeografice şi procesele morfogenetice actuale de modelare a reliefului (Tabelul 1.5)
Tabelul 1.5. Unităţi morfoclimatice montane de pe teritoriul ţării noastre
Mountainous morpho climatic units from Romanian territory ( după Monografia Geografică, 1960, vol I, pag. 181)
Muncei şi munţi
mijlocii Caractere
fitogeografice Procese morfogenetice actuale
Muncei şi munţi mijlocii
Zona forestieră Eroziune lineară Eroziune chimică în carst Alunecări, eroziune moderată a solurilor Acumulare fluviatilă Acumulare deluvio-pluvională
Munţi inalţi Zona alpina Eroziune lineară torenţială Alterare fizică ( dezagregare) Acumulare pe pantă- grohotişuri Eroziune eoliană Eroziune mare a solurilor
20
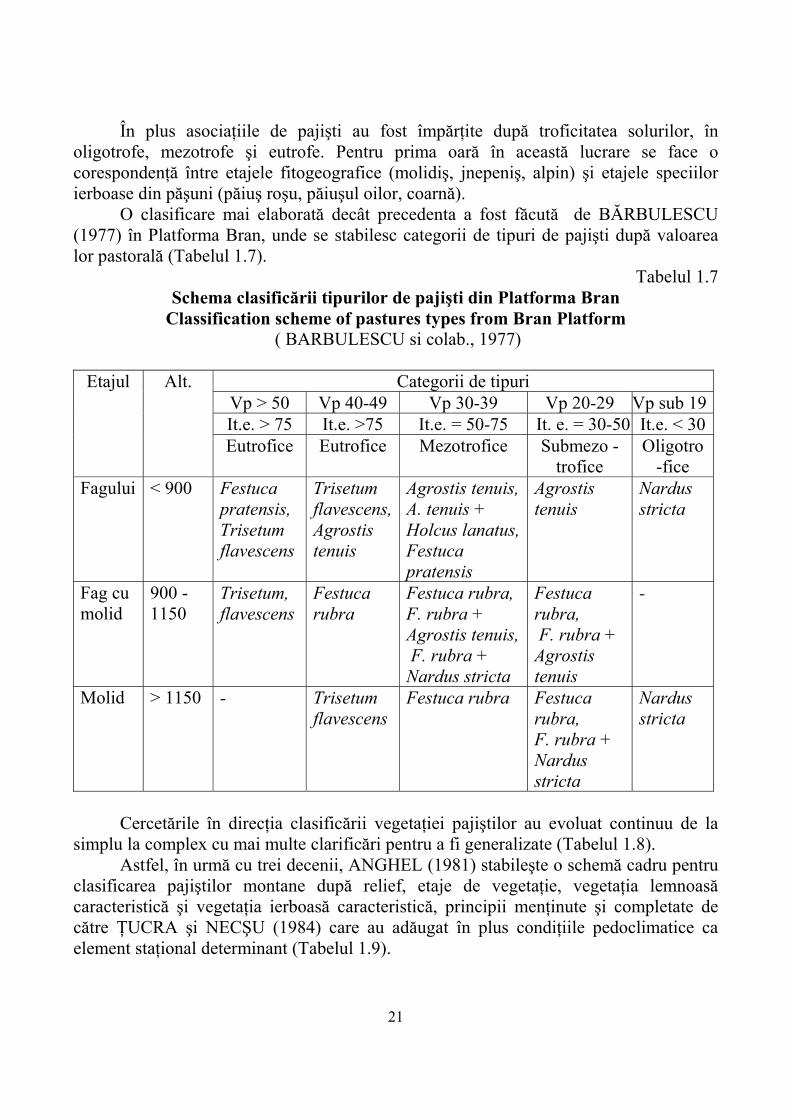
De aici rezultă că în muncei şi munţi mijlocii cu zonă forestieră şi în munţii înalţi cu zonă alpină procesele morfogenetice sunt foarte active pe măsură ce creşte altitudinea, din care eroziunea pluvială şi eoliană a solului şi alunecările de teren ocupă un loc important. Alături de pădure, covorul ierbos al pajiştilor prin ţelina care o produce este un scut extreme de eficient împotriva eroziunii. Din aceste considerente este necesară menţinerea unui covor ierbos dens pe pajiştile montane, îndeosebi a celor situate pe pante. Una din primele clasificări a vegetaţiei pajiştilor care au în preambul etajele, altitudinea şi solurile a fost făcută de ANGHEL (1965) pentru Munţii Cibinului (Tabelul 1.6).
Tabelul 1.6. Schema clasificării păşunilor şi fâneţelor din Munţii Cibinului
Classification scheme of pastures and meadows from Mountains Cibin ( ANGHEL şi colab., 1965)
Etajul Alt. Solul Asociaţii de pajişti Oligotrofe Mezotrofe Eutrofe
Submontan 450-800
Podzoluri secundare, Soluri brune de pădure
Nardete submontane, Festucete sărăcite ( degradate)
As. sumbont. de Agrostis tenuis si Festuca rubra
Arrhenatherum elatius, Festuca pratensis( fâneţe, Lolium perenne (păşuni)
Eumontan (făgete)
800-1250
Brune acide Brune acide scheletice Brune acide, podzolice, rar podzoluri primare
Nardete montane Brukentalia spiculifolia + Hieracium pilosselaFestuca rubra fallax
As. montane şi forme de trecere între Agrostis tenuis si Festuca rubra fallax
Fâneţe, cu pâlcuri de Festuca pratensis, Dactylis glomerata – Trisetum flavescens
Montan superior (molidiş)
1200-1800
Idem, dar cu podzoluri primare mai frecvente
Nardete montane Festucete- depreciate ( sărăcite )
Aproape doar Festuca rubra si Campanula abietina
Grupa ( inexistentă în etaj)
Subalpin (jnepeniş)
1750-2050
Podzoluri primare,acidi- tate extremă
Nardete subalpine Festuca rubra Agrostis rupestris
Poa media ( târlire)
-
Alpin ( rodoret)
2000-2245
Brune alpine Nardete în partea inferioară, pe alocuri însorite Carex curvula Cetraria islandica Thamnolia vermicularis în partea superioară
Festuca violaceea Poa media pe grohotişuri
-
21
În plus asociaţiile de pajişti au fost împărţite după troficitatea solurilor, în oligotrofe, mezotrofe şi eutrofe. Pentru prima oară în această lucrare se face o corespondenţă între etajele fitogeografice (molidiş, jnepeniş, alpin) şi etajele speciilor ierboase din păşuni (păiuş roşu, păiuşul oilor, coarnă).
O clasificare mai elaborată decât precedenta a fost făcută de BĂRBULESCU (1977) în Platforma Bran, unde se stabilesc categorii de tipuri de pajişti după valoarea lor pastorală (Tabelul 1.7).
Tabelul 1.7 Schema clasificării tipurilor de pajişti din Platforma Bran
Classification scheme of pastures types from Bran Platform ( BARBULESCU si colab., 1977)
Etajul Alt. Categorii de tipuri
Vp > 50 Vp 40-49 Vp 30-39 Vp 20-29 Vp sub 19It.e. > 75 It.e. >75 It.e. = 50-75 It. e. = 30-50 It.e. < 30Eutrofice Eutrofice Mezotrofice Submezo -
trofice Oligotro
-fice Fagului < 900 Festuca
pratensis, Trisetum flavescens
Trisetum flavescens, Agrostis tenuis
Agrostis tenuis, A. tenuis + Holcus lanatus, Festuca pratensis
Agrostis tenuis
Nardus stricta
Fag cu molid
900 -1150
Trisetum, flavescens
Festuca rubra
Festuca rubra, F. rubra + Agrostis tenuis, F. rubra + Nardus stricta
Festuca rubra, F. rubra + Agrostis tenuis
-
Molid > 1150 - Trisetum flavescens
Festuca rubra Festuca rubra, F. rubra + Nardus stricta
Nardus stricta
Cercetările în direcţia clasificării vegetaţiei pajiştilor au evoluat continuu de la
simplu la complex cu mai multe clarificări pentru a fi generalizate (Tabelul 1.8). Astfel, în urmă cu trei decenii, ANGHEL (1981) stabileşte o schemă cadru pentru
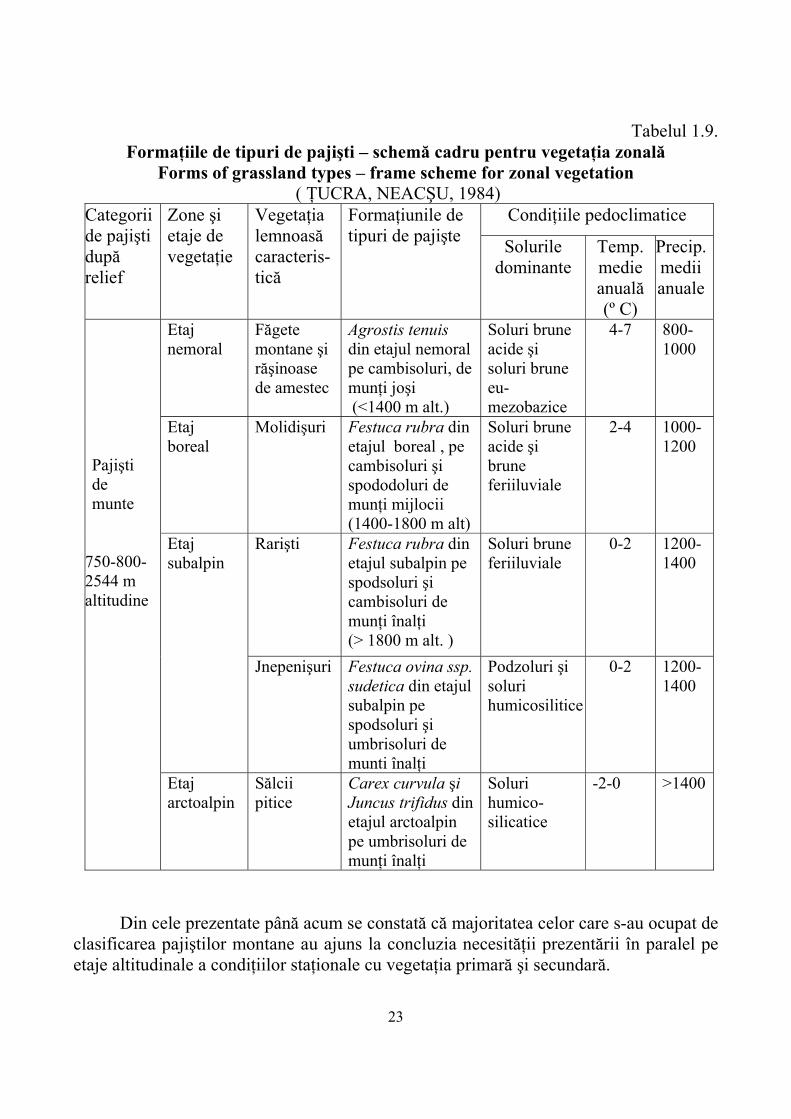
clasificarea pajiştilor montane după relief, etaje de vegetaţie, vegetaţia lemnoasă caracteristică şi vegetaţia ierboasă caracteristică, principii menţinute şi completate de către ŢUCRA şi NECŞU (1984) care au adăugat în plus condiţiile pedoclimatice ca element staţional determinant (Tabelul 1.9).

22
Tabelul 1.8. Schemă cadru pentru clasificarea vegetaţiei zonale montane a pajiştilor
permanente( naturale, seminaturale) din România Frame scheme of classification of zonal mountainous vegetation from grasslands
( ANGHEL 1981) Categorii de pajişti după
relief
Zone şi etaje de
vegetaţie
Vegetaţia lemnoasă caracteristică
Vegetaţia ierboasă caracteristică
Pajişti de munte ( 750-800-2544 m altitudine )
Etaj nemoral
Făgete montane Pajişti de Agrostis tenuis, A. tenuis cu Festuca rubra
Etaj boreal Molidişuri Pajişti de Festuca rubra ssp. fallax, Festuca rubra ssp. commutata
Etaj subalpin
Rarişti cu molid şi jnepenişuri
Pajişti de Festuca rubra ssp. commutata şi pajişti de Festuca ovina ssp. sudetica sin. Festuca supina
Etaj arctoalpin
Sălcii pitice (Salix herbacea etc.)
Pajişti de Carex curvula şi pajişti de Juncus trifidus
23
Tabelul 1.9. Formaţiile de tipuri de pajişti – schemă cadru pentru vegetaţia zonală
Forms of grassland types – frame scheme for zonal vegetation ( ŢUCRA, NEACŞU, 1984)
Categorii de pajişti după relief
Zone şi etaje de vegetaţie
Vegetaţia lemnoasă caracteris-tică
Formaţiunile de tipuri de pajişte
Condiţiile pedoclimatice
Solurile dominante
Temp. medie anuală (º C)
Precip.medii anuale
Pajişti de munte
750-800-2544 m altitudine
Etaj nemoral
Făgete montane şi răşinoase de amestec
Agrostis tenuis din etajul nemoral pe cambisoluri, de munţi joşi (<1400 m alt.)
Soluri brune acide şi soluri brune eu- mezobazice
4-7 800- 1000
Etaj boreal
Molidişuri Festuca rubra din etajul boreal , pe cambisoluri şi spododoluri de munţi mijlocii (1400-1800 m alt)
Soluri brune acide şi brune feriiluviale
2-4 1000- 1200
Etaj subalpin
Rarişti
Festuca rubra din etajul subalpin pe spodsoluri şi cambisoluri de munţi înalţi (> 1800 m alt. )
Soluri brune feriiluviale
0-2 1200- 1400
Jnepenişuri Festuca ovina ssp. sudetica din etajul subalpin pe spodsoluri şi umbrisoluri de munti înalţi
Podzoluri şi soluri humicosilitice
0-2 1200- 1400
Etaj arctoalpin
Sălcii pitice
Carex curvula şi Juncus trifidus din etajul arctoalpin pe umbrisoluri de munţi înalţi
Soluri humico-silicatice
-2-0 >1400
Din cele prezentate până acum se constată că majoritatea celor care s-au ocupat de
clasificarea pajiştilor montane au ajuns la concluzia necesităţii prezentării în paralel pe etaje altitudinale a condiţiilor staţionale cu vegetaţia primară şi secundară.

24
Ca urmare a necesităţii aducerii la un numitor comun al tuturor comunităţilor vegetale şi condiţiilor staţionale la nivel global şi implicit în Uniunea Europeană s-au căutat şi găsit soluţii de clasificare comune, numerotate pentru toate tipurile de habitate, inclusiv pentru pajiştile naturale.
Din analiza lucrării HABITATELE DIN ROMÂNIA de N. DONIŢĂ şi colaboratorii, rezultă că au fost descrise într-o primă aproximaţie 367 tipuri de habitate încadrate la 7 clase şi 24 subclase după sistemul de clasificare PALEARCTIC HABITATS (PAL. HAB.) .Dintre aceste 3 clase, 10 subclase şi 96 tipuri de habitate (26%) aparţin parţial sau în totalitate vegetaţiei pajiştilor naturale şi seminaturale.
Această nouă clasificare la care va trebui să se ralieze şi ţara noastră, complică destul de mult lucrurile, fiind necesare eforturi suplimentare de asimilare şi reaşezare a tuturor datelor privind vegetaţia pajiştilor şi staţiunile acestora acumulate până în prezent.
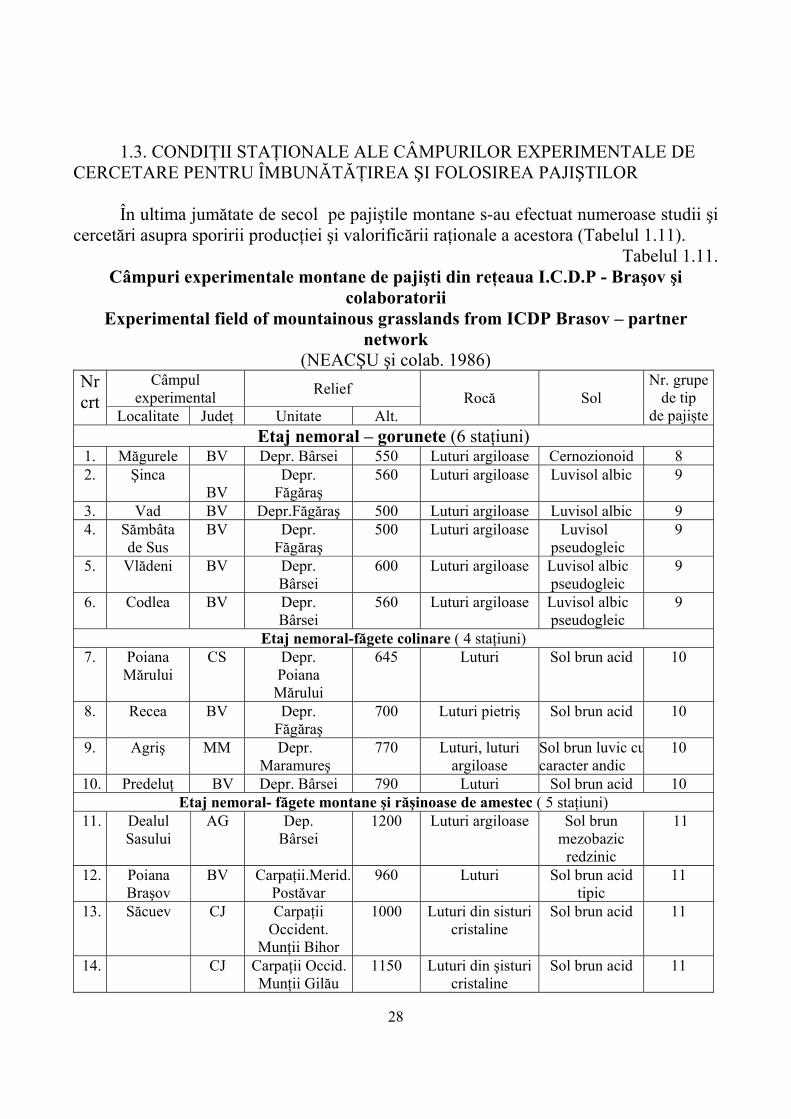
Într-o primă aproximaţie a încercării de clasificare a tipurilor de habitate de pajişti au rezultat un număr de 47 habitate în cele mai diverse condiţii naturale (Tabelul 1.10).
După acest sistem se constată că pajiştile aparţin la următoarele clase şi subclase: 3. Tufărişuri şi pajişti 36 pajişti alpine şi subalpine cu 19 tipuri de habitate;
37 Pajişti umede şi comunităţi de ierburi înalte cu 16 tipuri de habitate; 38 Pajişti mezofile cu 4 tipuri de habitate; 6. Grohotişuri, stâncării şi nisipuri continentale 63 Vegetaţie chinofilă cu 6 tipuri de habitate; 8. Terenuri agricole şi peisaje artificiale 87 Comunităţi ruderale cu 2 tipuri de habitate de pajişti. Rezultă în final 47 tipuri de habitate de pajişti montane cu o repartizare extrem de
heterogenă atât sub aspectul condiţiilor staţionale, al valorii conservative cât şi al valorii economice exprimate prin indicatorii de valoare furajeră. Cele mai răspândite ca suprafaţă şi valoroase pajişti din punct de vedere calitativ şi productiv sunt cele mezofile la care din păcate sunt menţionate doar 4 tipuri !?
În schimb pentru pajiştile cu vegetaţie chinofilă, mult mai neînsemnate decât primele se descriu 6 tipuri de habitate. Această constatare face ca să elaborăm pe viitor o clasificare a habitatelor de pajişti mult mai laborioasă care să corespundă atât nevoilor de conservare a biodiversităţii cât şi a celor economice.

25
Tabelul 1.10. Principalele tipuri de habitate cu pajişti montane, din România
Principal types of mountainous grassland habitats from Romania (DONIŢĂ şi colab, 2005)
Cod Tip Habitat CV(x) VP(xx) CP(xxx)3 Tufărişuri şi pajişti 36 Pajisti alpine si subalpine (R-FM) (1-35) 0,1-0,7R 3601 Pajişti sud-est carpatice de rogoz pitic
(Kobresia myosuroides ) şi Oxytropis carpaticaM 1-5 0,1
R 3602 Pajişti sud-est carpatice de coarna (Carex curvula ) şi Primula minima
R-M 15-20 0,3-0,4
R 3603 Pajişti sud-est carpatice de părul porcului ( Juncus trifidus) şi Potentilla minima
R 5-10 0,1-0,2
R 3604 Pajişti sud-est carpatice de păruşcă ( Festuca supina) şi Potentilla ternata
M 5-15 0,1-0,3
R 3605 Pajişti sud-est carpatice de păiuş de colţi (Festuca versicolor) şi Sesleria rigida ssp. haynaldiana
M 5-10 0,1-0,2
R 3606 Pajişti sud-est carpatice de păiuş de stanci ( Festuca saxatilis)
M 5-10 0,1-0,2
R 3607 Pajişti sud-est carpatice de Festuca amenthystina şi Dianthus tenuifolius
M 5-10
0,1-0,2
R 3608 Pajişti sud-est carpatice de Scorzonera rosea şi Festuca nigrescens (sin. F. rubra commutata)
m 25-35 0,5-0,7
R 3609 Pajişti sud-est carpatice de ţăpoşică( Nardus stricta) şi Viola declinata
m 10-15 0,2-0,3
R 3610 Pajişti sud-est carpatice de Poa media m 15-20 0,3-04 R 3611 Pajişti sud-est carpatice de coada iepurelui
( Sesleria rigida ssp. haynaldiana) şi rogoz ( Carex sempervirens)
M 5-10 0,1-0,2
R 3612 Pajişti sud-est carpatice de rogoz ( Carex sempervirens) şi coarnă mare ( Sesleria bielzii)
M 5-10 0,1-0,2
R 3613 Pajişti sud-est carpatice de Carduus kerneri, Festuca carpatica şi Trisetum fuscum
M 15-25 0,3-0,5
(x) C.V. – apreciere valoare conservativă: R – redusă m – moderată
M – mare FM – foarte mare
(xx) V.P. - indice pentru valoarea pastorală: < 5 - pajişte degradată
5-15 – foarte slabă; 15- 25 – slabă; 25-50 – mijlocie;
50-75 – bună; 75-100 – pajişte foarte bună
(xxx) - C.P.- capacitate păşunat = VP x 0,02 (UVM/ha)

26
Cod Tip Habitat C.V. V.P. C.P. R 3614 Pajişti sud-est carpatice de Festuca xanthina M 5-10 0,1-0,2R 3615 Tufărişuri pitice sud-est carpatice de sălcii
alpine ( Salix herbacea) M 1-5
0,1
R 3616 Tufărişuri pitice sud-est carpatice de sălcii alpine ( Salix retusa, S. reticulata)
M 1-5 0,1
R 3617 Tufărişuri pitice de arginţică ( Dryas octopetala) M 1-5 0,1 R 3618 Tufărişuri pitice sud-est carpatice de sălcii
endemice ( Salix kitaibeliana) cu degetăruţ ( Soldanella hungarica)
FM 1-5
0,1
R 3619 Tufărişuri pitice sud-estice de sălcii endemice (Salix kitaibeliana) cu degetăruţ alpin ( Soldanella pusilla)
FM 1-5 0,1
37 Pajişti umede şi comunităţi de ierburi inalte ( buruienişuri)
(R-M)
(1-70) (0,1-1,4)
R 3701 Comunităţi sud-est carpatice de buruienişuri înalte cu Aconitum tauricum
R 1-5
0,1
R 3702 Comunităţi sud-est carpatice de buruienişuri înalte de Adenostyles alliaria şi Doronicum austriacum
R 1-5 0,1
R 3703 Comunităţi sud-est carpatice de buruienişuri înalte de Cirsium waldsteinii şi Heracleum sphondylium ssp. transilvanicum
R 1-5
0,1
R 3704 Comunităţi sud-est carpatice de buruienişuri înalte de Senecio subalpinus şi ştevia stanelor ( Rumex alpinus)
R 1-5
0,1
R 3705 Comunităţi sud-est carpatice de buruienişuri înalte de Rumex obtusifolia şi Urtica dioica
R 1-5 0,1
R 3706 Comunităţi sud-est carpatice de buruienişuri înalte de Petasites kablikianus
R 1-5
0,1
R 3707 Comunităţi sud-est carpatice de buruienişuri înalte de Telekia speciosa şi Petasites hybridus
R 1-5
0,1
R 3708 Comunităţi daco-getice cu Angelica syslvestris, Crepis paludosa şi Scirpus sylvaticus
R-M 1-5
0,1
R 3709 Comunităţi danubiene de Juncus effusus, J. inflexus şi Agrostis canina
R 5-15 0,1-0,3
R 3710 Pajişti dacice de Molinia coerulea m 1-5 0,1R 3711 Pajişti dacice de Nardus stricta şi Molinia
coerulea m 5-15 0,1-0,3
R 3712 Comunităţi dacice cu Deschampsia caespitosa şi Agrostis stolonifera
R 10-30 0,2-0,6

27
Cod Tip Habitat C.V. V.P. C.P. R 3713 Pajişti antropice de Juncus tenuis şi Trifolium
repens R 10-20 0,2-0,4
R 3714 Comunităţi daco-getice cu Filipendula ulmaria, Geranium palustre şi Alopecurus pratensis
R 1-5 0,1
R 3715 Pajişti danubian-panonice de Agrostis stolonifera
R-M 30-60 0,6-1,2
R 3716 Pajişti danubian-pontice de Poa pratensis, Festuca pratensis şi Alopecurus pratensis
m 40-70 0,8-1,4
38 Pajişti mezofile (R-m) (40-80) (0,8-1,6)
R 3801 Pajişti sud-est carpatice de Trisetum flavescens şi Alchemilla vulgaris
R 50-70 1,0-1,4
R 3802 Pajişti daco-getice de Arrhenatherum elatius m 60-80 1,2-1,6R 3803 Pajişti sud-est carpatice de Agrostis capillaris
(sin Agrostis tenuis)şi Festuca rubra R 40-60 0,8-1,2
R 3804 Pajişti daco-getice de Agrostis capillaris şi Anthoxanthum odoratum
m 40-60 0,8-1,2
6 Grohotişuri, stâncării şi nisipuri continentale 63 Vegetaţie chinofilă (R-m) (1-5) (0,1) R6301 Comunităţi sud-est carpatice chinofile cu
Arenaria biflora R 1-5
0,1
R 6302 Comunităţi sud est carpatice chinofile cu Polytrichum sexangularis
R 1-5 0,1
R 6303 Comunităţi sud est carpatice chinofile cu Luzula alpino-pilosa
R 1-5
0,1
R 6304 Comunităţi sud est carpatice chinofile cu Ranunculus crenatus şi Soldanella pusilla
R 1-5
0,1
R6305 Comunităţi sud est carpatice chinofile cu Gnaphalium supinum şi Nardus stricta
R 1-5 0,1
R 6306 Comunităţi sud est carpatice chinofile cu Poa supina şi Cerastium cerastioides
m 1-5
0,1
8 Terenuri agricole şi peisaje artificiale 87 Comunităţi ruderale (R-M) (1-15) (0,1-
03) R8706 Comunităţi sud-est carpatice cu Calamagrostis
arundinacea, Epilobium angustifolium şi Atropa bella-donna
R 1-5 0,1
R8707 Comunităţi sud-est carpatice cu Poa supina R 5-10 0,1-0,2
28
1.3. CONDIŢII STAŢIONALE ALE CÂMPURILOR EXPERIMENTALE DE
CERCETARE PENTRU ÎMBUNĂTĂŢIREA ŞI FOLOSIREA PAJIŞTILOR
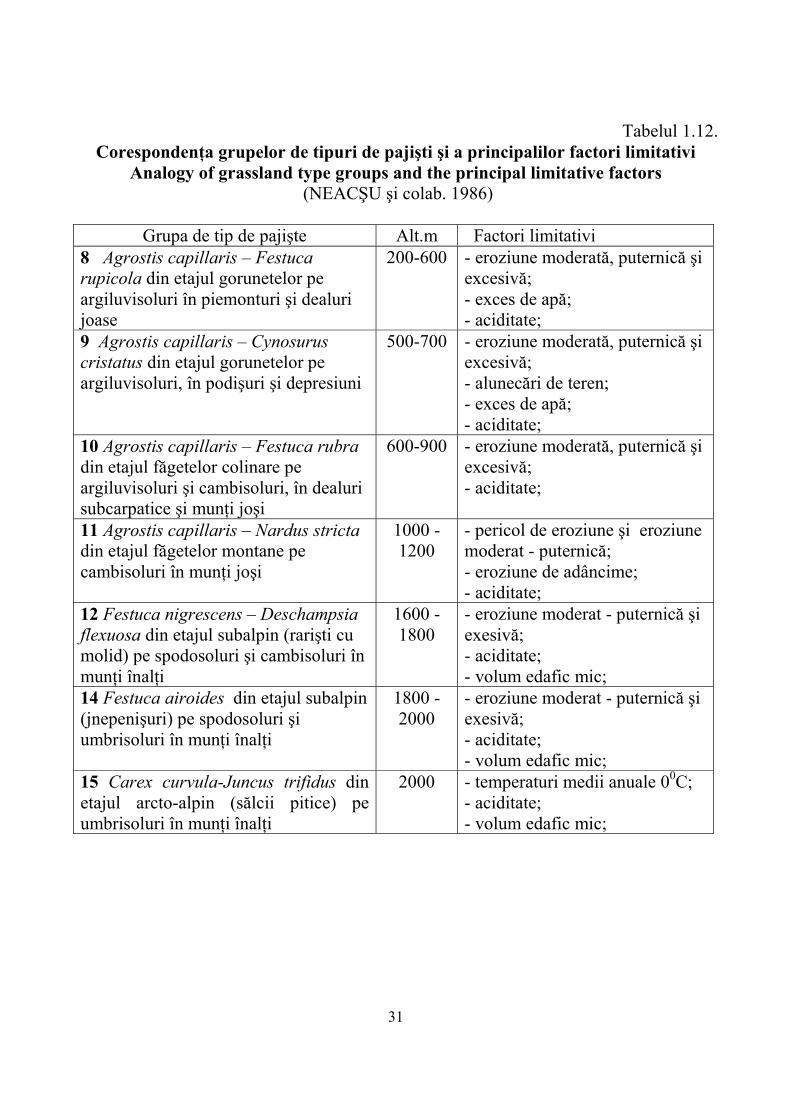
În ultima jumătate de secol pe pajiştile montane s-au efectuat numeroase studii şi cercetări asupra sporirii producţiei şi valorificării raţionale a acestora (Tabelul 1.11).
Tabelul 1.11. Câmpuri experimentale montane de pajişti din reţeaua I.C.D.P - Braşov şi
colaboratorii Experimental field of mountainous grasslands from ICDP Brasov – partner
network (NEACŞU şi colab. 1986)
Nr crt
Câmpul experimental
Relief Rocă Sol
Nr. grupede tip
de pajişteLocalitate Judeţ Unitate Alt. Etaj nemoral – gorunete (6 staţiuni)
1. Măgurele BV Depr. Bârsei 550 Luturi argiloase Cernozionoid 8 2. Şinca
BV Depr.
Făgăraş 560 Luturi argiloase Luvisol albic 9
3. Vad BV Depr.Făgăraş 500 Luturi argiloase Luvisol albic 9 4. Sămbâta
de Sus BV Depr.
Făgăraş 500 Luturi argiloase Luvisol
pseudogleic 9
5. Vlădeni BV Depr. Bârsei
600 Luturi argiloase Luvisol albic pseudogleic
9
6. Codlea BV Depr. Bârsei
560 Luturi argiloase Luvisol albic pseudogleic
9
Etaj nemoral-făgete colinare ( 4 staţiuni) 7. Poiana
Mărului CS Depr.
Poiana Mărului
645 Luturi Sol brun acid 10
8. Recea BV Depr. Făgăraş
700 Luturi pietriş Sol brun acid 10
9. Agriş MM Depr. Maramureş
770 Luturi, luturi argiloase
Sol brun luvic cucaracter andic
10
10. Predeluţ BV Depr. Bârsei 790 Luturi Sol brun acid 10 Etaj nemoral- făgete montane şi răşinoase de amestec ( 5 staţiuni)
11. Dealul Sasului
AG Dep. Bârsei
1200 Luturi argiloase Sol brun mezobazic
redzinic
11
12. Poiana Braşov
BV Carpaţii.Merid.Postăvar
960 Luturi Sol brun acid tipic
11
13. Săcuev CJ Carpaţii Occident.
Munţii Bihor
1000 Luturi din sisturi cristaline
Sol brun acid 11
14. CJ Carpaţii Occid.Munţii Gilău
1150 Luturi din şisturi cristaline
Sol brun acid 11

29
Nr crt
Câmpul experimental
Relief
Rocă
Sol
Nr. grupede tip
de pajişteLocalitate Judeţ Unitate Alt. 15. Padiş BH Carpatii
Occident. Munţii Bihor
1280 Luturi din gresii cuarţitice
Sol brun acid criptospodic
11
Etaj boreal- molidişuri (8 staţiuni) 16. Vlădeasa CJ Carpaţii
Occident. Munţii Bihor
1480 Luturi din riolite Sol brun acid criptozolic
12
17. Semenic CS Carpaţii Occident.
Grup.M-ţilor Banatului
1420 Luturi din sisturi cristaline
Sol brun acid criptozolic
12
18. Semenic CS Carpaţii Occident. Grupa M-ţilor
Banatului
1430 Luturi din şisturi cristaline
Sol brun feriiluvial litic
12
19. Crint SB Carpaţii Merid.
Grupa Parâng
1200 Luturi din sisturi cristaline
Sol brun feriiluvial litic
12
20. Păltiniş SB Carpaţii Merid.
Grupa Parâng
1350 Luturi din sisturi cristaline
Sol podzolic brun
12
21. Ciucaş-Brătei
PH Carpaţii Orientali,
grupa Sudică
1400 Luturi din conglomerate
Sol brun acid criptospodic
12
22. Gârbova ( Munţii Baiului)
PH Carpaţii Merid.
1675 Deluvii, din strate de Sinaia
Sol brun Acid
criptospodic
12
23. Rarău ( Munţii Rarau)
SV Carpaţii Orientali,
Grup. Nordică
1500 Luturi Sol brun acid criptospodic
12
Etajul subalpin-rarişti ( 3 staţiuni) 24. Leaota DB Carpaţii
Meridionali grupa Fagaraş
1750 Luturi din sisturi cristaline
Sol brun feriiluvial
13
25. Rânca ( Munţii Parâng)
GJ Carpatii Meridionali
grupa Parang
1680 Luturi din sisturi cristaline
Podzol tipic 13
26. Rânca GJ Carpaţii Meridionali
grupa Parâng
1650 Luturi din sisturi cristaline
Sol podzolic brun
13
Etaj subalpin-jnepenişuri ( 6 staţiuni) 27. Vlădeasa
(Munţii Bihor)
CJ Carpaţii Occident
grupa M-ţilor Apuseni
1830 Luturi din porfirite andezite
Sol brun acid criptospodic
14

30
Nr crt
Câmpul experimental
Relief
Rocă
Sol
Nr. grupede tip
de pajişteLocalitate Judeţ Unitate Alt. 28. Transfăgă-
răşan ( Munţii Făgăraş)
SB Carpaţii Merid. grupa
Fagaraş
1700 Luturi din sisturi cristaline
Podzol tipic 14
29. Iezer-Papuşa
AG Carpatii Merid. grupa
Fagaras
1800 Luturi din sisturi cristaline
Podzol tipic 14
30 Muntele Mic
(Ţarcu)
CS Carpatii Occident.
Munţii Banatului
1800 Luturi din roci eruptive
Sol brun feriiluvial
14
31. Muntele Roşu
( Munţii Ciucaş)
PH Carpaţii Orientali,
grupa sudică
1710 Luturi din gonglomerate
Podzol tipic 14
32. Blana (Munţii Bucegi)
DB Carpaţii Merid.
1820 Luturi din conglomerate
Sol brun feriiluvial
14
Etaj arcto-alpin- sălcii pitice ( 1 staţiune)
33. Cabana Bâlea
( Munţii Făgăraş)
SB Carpaţii Meridionali
Muntii Fagaraş
2240 Luturi din şisturi cloritoseriticioase
Sol humicosililatic
15
Pe cele trei mari grupe de munţi din lanţul carpatic românesc, staţiunile în care s-
au efectuat studii şi cercetări privind îmbunătăţirea pajiştilor se prezintă astfel : Carpaţii Orientali 4 staţiuni; C-ţii Meridionali 20 staţiuni şi Carpaţii Occidentali 9 staţiuni.
Din cele prezentate se constată un număr mai mare de staţiuni în Carpaţii Meridionali( 20) faţă de cei Occidentali ( 9) şi semnificativ mai mare faţă de Orientali (4) în ambele zone cercetările trebuind intensificate pe viitor
31
Tabelul 1.12. Corespondenţa grupelor de tipuri de pajişti şi a principalilor factori limitativi
Analogy of grassland type groups and the principal limitative factors (NEACŞU şi colab. 1986)
Grupa de tip de pajişte Alt.m Factori limitativi
8 Agrostis capillaris – Festuca rupicola din etajul gorunetelor pe argiluvisoluri în piemonturi şi dealuri joase
200-600 - eroziune moderată, puternică şi excesivă; - exces de apă; - aciditate;
9 Agrostis capillaris – Cynosurus cristatus din etajul gorunetelor pe argiluvisoluri, în podişuri şi depresiuni
500-700 - eroziune moderată, puternică şi excesivă; - alunecări de teren; - exces de apă; - aciditate;
10 Agrostis capillaris – Festuca rubra din etajul făgetelor colinare pe argiluvisoluri şi cambisoluri, în dealuri subcarpatice şi munţi joşi
600-900 - eroziune moderată, puternică şi excesivă; - aciditate;
11 Agrostis capillaris – Nardus stricta din etajul făgetelor montane pe cambisoluri în munţi joşi
1000 -1200
- pericol de eroziune şi eroziune moderat - puternică; - eroziune de adâncime; - aciditate;
12 Festuca nigrescens – Deschampsia flexuosa din etajul subalpin (rarişti cu molid) pe spodosoluri şi cambisoluri în munţi înalţi
1600 -1800
- eroziune moderat - puternică şi exesivă; - aciditate; - volum edafic mic;
14 Festuca airoides din etajul subalpin (jnepenişuri) pe spodosoluri şi umbrisoluri în munţi înalţi
1800 -2000
- eroziune moderat - puternică şi exesivă; - aciditate; - volum edafic mic;
15 Carex curvula-Juncus trifidus din etajul arcto-alpin (sălcii pitice) pe umbrisoluri în munţi înalţi
2000 - temperaturi medii anuale 00C; - aciditate; - volum edafic mic;

32
II. DESCRIEREA PAJIŞTILOR DEGRADATE DE Nardus stricta L. DIN JUDEŢUL BRAŞOV
2.1 CONDIŢII NATURALE 2.1.1. Cadrul geografic şi relieful Judeţul Braşov este situat în sud-estul Depresiunii intracarpatice a
Transilvaniei, fiind încadrat la est, sud-est şi sud de Carpaţii Orientali, la sud şi sud-vest de Carpaţii Meridionali şi la nord de interfluviul Târnava Mare-Olt.
Judeţul Braşov este învecinat cu 8 din judeţele ţării şi anume: la nord judeţele Mureş şi Harghita, la est judeţele Covasna şi Buzău, la sud judeţele Argeş, Dâmboviţa şi Prahova, la vest judeţul Sibiu.
În ansamblu, judeţul Braşov se prezintă ca o unitate bine conturată. Suprafaţa judeţului se desfăşoară pe 5351 km2 şi cuprinde unităţi şi forme de relief foarte variate.
Relieful judeţului Braşov prezintă în amănunt o mare complexitate morfologică şi morfogenetică, fiind rezultatul unei îndelungate evoluţii pe un fundament de geosinclinal carpatic şi pe variate structuri geologice. Analiza unor indici morfometrici şi a unor aspecte morfografice dă posibilitatea identificării unor complexe naturale bine definite. Urmărite de la sud spre nord, în cuprinsul judeţului se disting trei trepte majore de relief (Fig. 2.1.)
- treapta munţilor înalţi, situaţi la limita sudică, cu altitudini absolute depăşind 1700 m, în Făgăraş, Piatra Craiului, Leaota, Bucegi, Postăvarul, Piatra Mare şi Ciucaş; aceste grupe muntoase, deşi diferenţiate structural, au avut totuşi o dezvoltare relativ asemănătoare pe flancul nordic, condiţionată de nivelul de bază local, reprezentat prin râul Olt;
- treapta munţilor scunzi, între cca. 800 şi 1700 m, situaţi la limita estică, sud-estică şi parţial sudică, în care se încadrează Întorsura Buzăului, Dârstelor, Tâmpa, Poiana Braşovului, Măgura Codlei şi Perşani;
- treapta depresiunilor şi a dealurilor de podiş, realizate pe unitate de geosinclinal, situată între 450 şi 700 m altitudine, în partea centrală şi nordică a judeţului, în care câmpiile apar desfăşurate în cele mai tipice depresiuni intramontane şi submontane interne. În configuraţia generală a acestei trepte se disting: Câmpia Bârsei şi golurile depresionare Zărneşti -Tohan, Vlădeni şi culoarul Măieruş, Câmpia Făgăraşului, depresiunea Homoroadelor şi culmile deluroase ale laturii sud-estice a Podişului Transilvaniei (colinele Podişului Târnavelor).
Cea mai mare parte din suprafaţa judeţului revine câmpiilor şi dealurilor de 450-700 m, cca. 60%, munţii scunzi (800-1700 m) ocupă 22% iar munţii înalţi (peste 1700 m) ocupă 18% (IANCU şi col. 1971).
Aceste elemente de relief au influenţă determinantă în repartizarea unităţilor de vegetaţie primară şi a celor secundare, din care un loc important îl au pajiştile.

33
Fig. 2.1. Unităţile de relief din judeţul Braşov Typography units from Brasov County
2.1.2. Geologia Sub raport geologic, judeţul Braşov apare ca un adevărat mozaic: cristalin, calcare, conglomerate, gresii, marne etc., a căror răspândire este rezultatul frământărilor scoarţei terestre şi acţiunii agenţilor externi. În alcătuirea munţilor înalţi predomină:
- în m. Făgăraşului: şisturi cristaline, predominant acide; - în m.Bucegi şi Piatra Craiului: calcare jurasice şi conglomerate cretacice; - în m. Piatra Mare şi Postăvarul: calcare jurasice şi conglomerate
cretacice; - în m. Ciucaş: flişil cretacic format din marne, gresii, şisturi marno-
grezoase şi conglomerate calcaroase; - în m. Leaota şisturi cristaline (filite, şisturi sericitocloritoase); În alcătuirea munţilor scunzi predomină: - în m.Perşani: roci cristaline, conglomerate calcaroase, gresii şi marne; - în m. Buzăului: fliş cretacic şistos-grezos; - în m. Dârstei, Tâmpa, Poiana Braşovului: conglomerate calcaroase,
calcare, marne şi gresii; - în Măgura Codlei: conglomerate, gresii, calcar şi marne.

34
Depresiunea Bârsei este instalată pe un fundament de fliş cretacic şi a reprezentat un lac intramontan care a rămas închis până la sfârşitul cuaternarului inferior. În dacian depresiunea a fost colmatată cu nisipuri şi marne, iar în levantin cu pietrişuri şi nisipuri. În pleistocen şi holocen colmatarea depresiunii a continuat prin depunerea de bolovănişuri, nisipuri, pietrişuri, argile, toate cu caracter torenţial.
Dealurile învecinate sunt constituite din depozite deluviale alcătuite din rocile montane din apropiere.
Câmpia piemontană este alcătuită din materiale heterogene proluviale (pietriş, nisip, argile). Pe terase ultimele depuneri sunt mai fine (argiloase, lutoase) decât restul câmpiei, datorită transportului eolian al materialului fin şi unei sedimentări mai liniştite. Ele acoperă, într-un strat gros de 1-3 m aceste terase.
În luncile din depresiune se întâlnesc depozite aluviale stratificate (nisip, pietriş) în general carbonatice. Pe văile torenţiale din zone de deal se găsesc pietrişuri şi bolovănişuri de conglomerate şi gresii.
Depresiunea Făgăraşului este colmatată cu depozite miocene şi pliocene inferioare, peste care se suprapun depozitele pliocenului superior (nisipuri, pietrişuri, marne şi argile marnoase).
În cuaternar, sub influenţa schimbărilor climatice (glaciare-interglaciare) au loc suprapuneri şi juxtapuneri pe conuri de dejecţie alcătuite din depozite torenţiale, aluvio-proluviale.
În zona de deal predomină formaţiuni miocene: conglomerate, marne etc., acoperite adesea de un depozit mai fin lutos.
Câmpia piemontană este alcătuită din depozite cu stratificaţie încrucişată, groase (40-50 m grosime), formate din pietrişuri, nisipuri şi luturi argiloase, cu reacţie acidă. Aceste depozite sunt acoperite cu material fin de natură proluvială şi eoliană, cu grosime de 0,5-3 m, pe relieful vechi.
Luncile sunt formate din depozite aluviale stratificate, fine şi grosiere, carbonatice în lunca Oltului şi acide în luncile râurilor ce coboară din munţii Făgăraş.
Podişul Târnavelor este alcătuit din materiale sedimentare ce aparţin sarmaţianului. Ridicarea în masă a Carpaţilor, la sfârşitul pliocenului, a determinat cutarea depozitelor sedimentare existente. Ele sunt formate din alternanţe de marne şi nisipuri cu intercalaţii de gresii şi tufuri. Aceste roci sunt slab consolidate, bazice şi intermediare. Alternanţa rocilor sarmatice, impermeabilitatea unora dintre ele şi panta terenului sunt factori determinanţi ai alunecărilor de teren destul de frecvente în această zonă. De-a lungul văilor din podiş sunt depozite aluviale şi aluvio-coluviale ce aparţin cuaternarului (Monografia geografică a RPR 1960).
Substratul geologic al judeţului Braşov, precum şi factorii climatici şi biotici au avut o influenţă determinantă în formarea şi predominarea unor soluri acide în zona de munte şi depresiunea Făgăraşului, slab acide în depresiunea Bârsei şi neutre spre bazice în podişul Târnavelor.
În linii generale răspândirea nardetelor în judeţ este legată de prezenţa solurilor acide, formate pe roci acide sau alte categorii de roci puternic debazificate.

35
2.1.3. Clima Unităţile şi formele de relief amintite, cu înălţimi absolute de la 400-500 m
până la peste 2500 m altitudine exercită influenţe deosebite asupra condiţiilor climatice. Acestea determină variaţii specifice ale temperaturilor şi precipitaţiilor, care se reflectă prin zonalitatea pe verticală a învelişului de sol şi vegetaţie.
2.1.3.1. Temperatura aerului Variaţiile suprafeţei subiacente-active (alternarea culmilor muntoase cu
văile adânci, a suprafeţelor împădurite cu cele lipsite de păduri, a luncilor umede cu suprafeţe mai uscate), au repercusiune directă asupra temperaturii aerului, ca urmare a repartizării neuniforme a radiaţiei solare.
De la marile înălţimi montane din sudul judeţului până la dealurile joase de la periferia sa nordică, temperatura medie anuală a aerului creşte de la - 2,6 0C (Vârful Omul), pe vârfurile de peste 2500 m altitudine, la 4,9 0C (Predeal) pe munţii de înălţime mijlocie, pentru ca jos în depresiuni să ajungă la peste 8 0C (Făgăraş 8,2 0C) (Fig.2.2.)
Luna cea mai rece a anului este, pe culmile alpine, februarie, când temperatura medie scade la – 11,1 0C, iar în munţii mijlocii şi depresiuni, ianuarie, cu - 5,1 0C şi respectiv - 5,3 0C temperatură medie. Temperatura minimă absolută în ultimele şapte decenii a coborât pe teritoriul judeţului Braşov la – 38,5 0C (înregistrată la Bod la data de 25 ianuarie 1942), aceasta fiind şi cea mai scăzută temperatură de pe teritoriul ţării noastre.
Vara, temperatura aerului este pozitivă pe întreg teritoriul judeţului, luna cea mai caldă fiind august pe vârfurile munţilor (5,7 0C) şi iulie în depresiuni, unde încălzirea aerului este mai intensă şi temperatura depăşeşte 18 0C.
În anotimpurile de tranziţie, primăvara şi toamna, temperatura se menţine sub 0 0C pe înălţimile muntoase, dar creşte la 10-12 0C în depresiuni.
Ca urmare a caracteristicilor de relief, în judeţul Braşov se produc frecvente inversiuni de temperatură, ce duc în depresiuni la scăderea accentuată a temperaturii aerului, la apariţia îngheţurilor târzii de primăvară şi a celor timpurii de toamnă, la creşterea numărului de zile ceţoase etc., fenomene ce au repercusiuni negative asupra vegetaţiei pajiştilor şi a celorlalte culturi agricole.
Durata medie a intervalului fără îngheţ este de 140-160 zile anual, la altitudini mai joase, corespunzând în linii mari şi cu perioada de păşunat a pajiştilor.
2.1.3.2. Precipitaţiile atmosferice Datorită aceloraşi cauze şi precipitaţiile atmosferice au o repartiţie
neuniformă ca şi temperatura aerului, altitudinea imprimând şi cantităţilor de precipitaţii o zonalitate verticală.
Vârfurile înalte ale Bucegilor şi Făgăraşului primesc anual peste 1300 mm, în timp ce pe şesurile depresionare cantitatea de precipitaţii scade la mai puţin de jumătate (600 mm). Deschiderea largă spre V-NV înlesneşte advecţia maselor de aer umed, astfel încât pe câmpul depresionar al Făgăraşului cantitatea de precipitaţii este cu peste 100 mm mai ridicată decât pe cel al Bârsei.
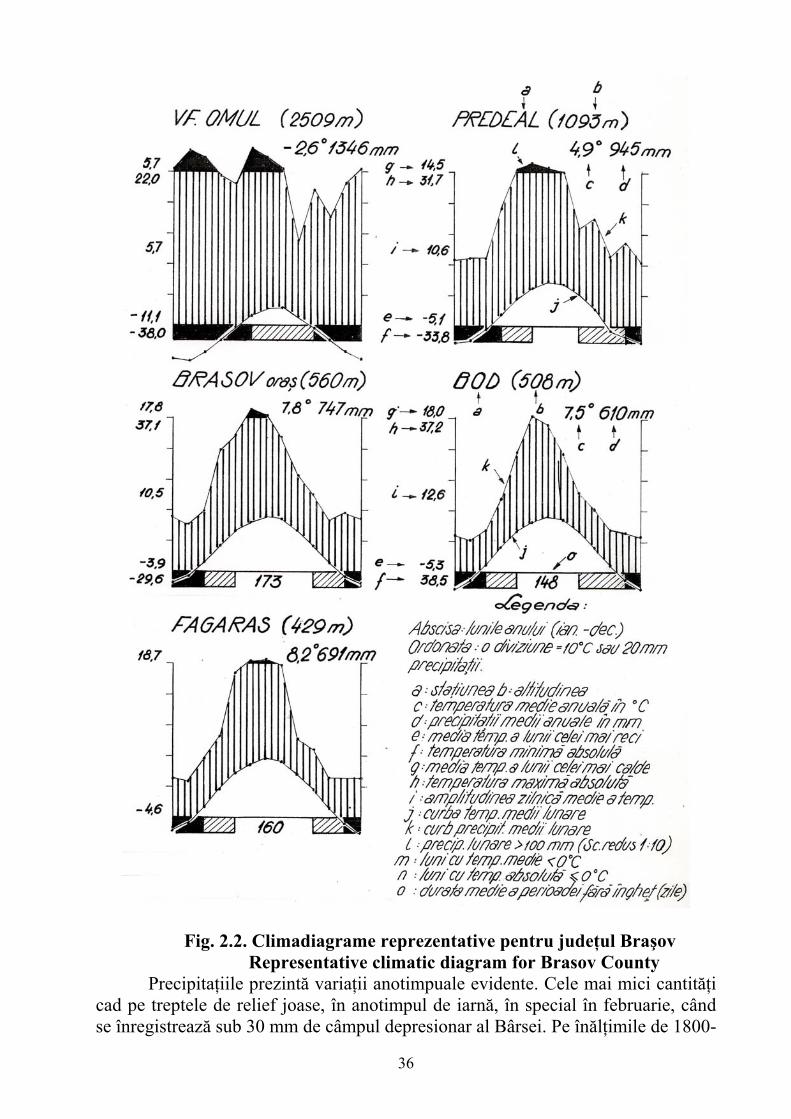
36
Fig. 2.2. Climadiagrame reprezentative pentru judeţul Braşov Representative climatic diagram for Brasov County
Precipitaţiile prezintă variaţii anotimpuale evidente. Cele mai mici cantităţi cad pe treptele de relief joase, în anotimpul de iarnă, în special în februarie, când se înregistrează sub 30 mm de câmpul depresionar al Bârsei. Pe înălţimile de 1800-

37
2000 m, luna cea mai săracă în ploi este septembrie cu valori sub 55 mm. În luna iunie cade cea mai mare cantitate de precipitaţii, depăşind 100 mm pe întreg teritoriul judeţului, înregistrându-se astfel maximumul pluviometric anual.
În anii cu activitate ciclonică deosebit de frecventă şi intensă, cantitatea de precipitaţii a fost de peste 1000 mm (Predeal 1321 mm; Braşov 1225 mm), iar pe munţii înalţi a depăşit 2000 mm (Vârful Omul 2402 mm).
În anii când activitatea anticiclonică este persistentă, sumele anuale de precipitaţii nu au depăşit 400-500 mm pe şesul depresionar al Bârsei (Bod 382 mm; Braşov 479 mm) şi 900 mm pe vârfurile munţilor (Vârful Omul 850 mm).
Aceste abateri de la un an la altul, se înregistrează şi pe parcursul fiecărui an. Astfel, cele mai intense precipitaţii pe teritoriul judeţului au fost înregistrate la Bod cu 220,5 mm în luna iulie 1898, iar în perioada 1-12 mai 1970 la Braşov şi Făgăraş au căzut peste 160 mm. Au fost înregistrate şi luni în care nu au căzut precipitaţii (Bod, septembrie 1903).
Constatările au dus la aprecierea că pe teritoriul judeţului Braşov, cantitatea de precipitaţii medii multianuale cade în timp de peste 130 zile pe an.
La sfârşitul toamnei şi începutul iernii, odată cu scăderea temperaturii aerului sub 0 0C, precipitaţiile cad mai mult sub formă de ninsoare. Dacă pe culmile muntoase zăpada se aşterne în strat continuu începând uneori cu luna septembrie, în depresiuni îşi face apariţia abia în luna noiembrie. Data medie a ultimei ninsori este mai timpurie în depresiuni, în martie şi mai târzie cu trei luni pe înălţimile munţilor, mai precis în luna iunie.
Stratul de zăpadă se depune discontinuu, având grosimi şi durate diferenţiate în funcţie de suprafaţa subiacentă respectivă. Pe înălţimile muntoase numărul mediu de zile cu strat de zăpadă depăşeşte 215 zile, în timp ce în zona joasă este în medie de 55 zile (IANCU şi colab. 1971).
Alături de temperatură şi ceilalţi factori climatici, precipitaţiile au un rol determinant în repartizarea vegetaţiei. Astfel, în judeţul Braşov, nardetele se întâlnesc numai în zonele unde precipitaţiile medii anuale depăşesc 600 mm.
2.1.3.3. Regimul vântului Vârfurile cele mai înalte ale Făgăraşului, Bucegilor şi Piatra Craiului, cu
peste 2000 m altitudine absolută, sunt supuse permanent vânturilor de nord-vest şi vest, a căror frecvenţă anuală depăşeşte 25 respectiv 20 %. Variaţiile de relief determină devierea şi canalizarea curenţilor de aer pe anumite direcţii. În sectorul estic al judeţului, pe câmpul depresionar al Bârsei, direcţia dominantă este de NE-SV (orientare generală a văii Oltului). Depresiunea Făgăraşului fiind larg deschisă spre NV-V, este supusă influenţei vânturilor dominante ce pendulează pe aceste direcţii.
Pe înălţimile munţilor, vânturile dominante bat aproape permanent (cca. 90 % din an) cu o viteză medie de peste 7-8 m/sec, în unele cazuri în lunile mai şi iunie ea depăşeşte 25-20 m/sec. În depresiuni, vânturile bat aproximativ 50-75 % din an, cu viteze medii de 2,5-3,5 m/sec.
Pe versantul nordic al Făgăraşului şi cel estic al Perşanilor, primăvara apar mişcări cu caracter föhnic, care determină şi alte fenomene meteorologice. Astfel,

38
în depresiunea Făgăraşului, "Vântul Mare" topeşte zăpada în câteva zile (M. Iancu şi colab. 1971).
2.1.3.4. Caracterizarea generală a climei După Monografia geografică a României (1960), pe teritoriul luat în studiu
în prezenta lucrare, se pot deosebi următoarele sectoare: Podişul Târnavelor şi cele două mari depresiuni a Bârselor şi Făgăraşului, situate la altitudini de 450 m până la cca.800 m, se caracterizează ca sectoare cu climă continentală moderată, cu subdiviziunile: IB-de dealuri, IG-de depresiuni şi IP-de pădure. Zonei de munte (IV) îi este specifică clima de munte în care se deosebesc subdiviziunile: IV-C-de munţi mijlocii, IV-D-clima munţilor înalţi, ambele încorporând subţinuturile cu climă de versanţi expuşi vânturilor vestice (E). în munţii Făgăraşului se mai adaugă districtul climatic cu efecte de föhn (f).
Indicii de ariditate de Martonne sunt cuprinşi între 38 la Făgăraş, 63 la Predeal şi 174 la Vârful Omul.
Nardetele în judeţul Braşov, din punct de vedere al indicilor de ariditate sunt cuprinse în limitele 40-120, prezentând deci o amplitudine largă în cadrul unui climat mai umed spre foarte umed.
2.1.4. Solul Diversitatea mare a condiţiilor de mediu din judeţul Braşov, determină
procese variate de solificare, care duc la formarea numeroaselor tipuri, subtipuri şi varietăţi de soluri.
Judeţul Braşov este situat în zona forestieră umedă (Iar > 35) şi cuprinde două subzone pedogeografice (PĂUNESCU, 1975).
- Subzona inferioară, ocupată de păduri de foioase şi amestec cu condiţii bioclimatice care determină un circuit biologic activ şi o acidifiere moderată a solurilor.
Aici predomină solurile brune, slab sau moderat acide şi solurile argiloiluviale evoluate din acestea (solurile brune podzolite şi solurile podzolice argiloiluviale).
- Subzona superioară, ocupată de molidişuri şi jnepenişuri, cu condiţii bioclimatice care determină un circuit biologic moderat acid sau lent şi o acidifiere puternică sau foarte puternică a solurilor. Sunt caracteristice acestei subzone solurile brune acide şi solurile evoluate din acestea prin procese de acidifiere puternică şi podzolire humicoferiiluvială (podzolurile şi solurile brune podzolice).
Sub influenţa predominantă a condiţiilor de substrat de relief sau umezire, se formează solurile intrazonale (limitrofe, hidromorfe, neevoluate).
Solurile formate în condiţii fizico-geografice ale judeţului Braşov sunt clasificate şi caracterizate pe baza criteriilor stabilite de ICPA (1976).
Soluri din seria podzolirii argiloiluviale. 1. Soluri brune tipice, mollice, vertice, cumulice, rendzinice,
pseudorendzinice, regosolice, litice, gleizate, pseudogleizate se întâlnesc între 500-800 (1000) m altitudine în depresiunile Braşov şi Făgăraş (pe terase, piemonturi,

39
conuri de dejecţie, pe versanţii şi culmile zonei colinare şi montane inferioare, pe podişul Târnavelor (pe culmi, versanţi, terase). S-au format pe materiale parentale (depozite) foarte variate ca origine, compoziţie mineralogică, textură şi vârstă (luturi şi argile, marne, gresii, calcare, conglomerate, şisturi cristaline, nisipuri şi pietrişuri9. În general sunt bogate în componente bazice, carbonatice sau necarbonatice.
Apa freatică se găseşte în general mai jos de 5 m, cu excepţia teraselor inferioare, la extrema din aval a conurilor de dejecţie la baza unor versanţi.
Vegetaţia forestieră: păduri de Quercus petraea, Quercus robur şu uneori amestec de Fagus silvatica şi Quercus petraea.
Vegetaţia ierboasă: pajişti de Arrhenatherum elatius, Festuca pratensis, Agrostis capillaris şi altele.
Solurile brune se caracterizează prin textură grosieră (nisipo-lutoasă), mijlocie (luto-nisipoasă, lutoasă) şi fină (luto-argiloasă), nediferenţiată textural pe profil: sunt slab moderat acide (pH 5,4-6,8), oligomezobazice şi mezobazice (V 55-90 %), slab-bogat humifere (2-10 %), mai slab humifere pe terenurile cultivate. Sunt soluri mijlociu fertile.
2. Soluri brune podzolite, tipice, vertice, planice, pseudogleizate. Se întâlnesc în arealul solurilor brune dar pe relief mai vechi (terase înalte, relief piemontan vechi, culmi şi versanţi slab înclinaţi).
Roca şi materialul parental este constituit din depozite mai fine, relativ groase, uneori stratificate (soluri policiclice).
Soluri cu textură mijlocie şi fină (lutoase, luto-argiloase), evident diferenţiate textural pe profil: sunt slab puternic acide (pH 4,5-6,5) mezobazice (V 55-75 %), slab-mijlociu humifere (1-3 %). Valorile pH, V şi humus sunt mai scăzute în orizontul eluvial (El) comparativ cu orizontul A. fertilitatea este slabă-mijlocie, în funcţie de caracterul factorilor ecopedologici limitativi (textură fină, drenaj slab, reacţie acidă etc.).
3. Solurile podzolice argiloiluviale, tipice, glosice, holoacide, vertice, planice, pseudogleizate, pseudogleice.
Se întâlnesc în arealul solurilor brune podzolite, frecvent asociate cu acestea şi cu solurile pseudogleice podzolite, pe forme vechi de relief din cadrul piemonturilor, pe terase înalte, pe culmi în general pe terenuri cu pante mici şi zone depresionare.
S-au format pe materiale (depozite) fine, uneori polistratificate (cu urme ale proceselor periglaciare), cu grosimi de peste 2 m.
Sunt soluri cu textură mijlocie (luto-nisipoasă-lutoasă) în orizontul A şi Ba şi fină (luto-argiloasă-argiloasă) în orizontul Bt, diferenţierea texturală fiind puternică, sunt moderat-puternic acide (pH 4,3-5,8), oligobazice-oligomezobazice (V 25-75 %)pe terenurile cultivate, slab-mijlociu humifere (1-3 %), conţin aluminiu şi mangan uneori în doze nocive plantelor, însuşirile fizice nefavorabile (porozitate redusă, tasare ridicată, aproape impermeabil, capacitate redusă de structurare). Sunt soluri cu fertilitate scăzută.
Pe aceste soluri factorii ecopedologici cu caractere negative pentru vegetaţie, determină pe pajişti ca specia Nardus stricta să invadeze şi să

40
înlocuiască speciile mai valoroase de Festuca rubra şi Agrostis tenuis. Procesul este mai accentuat acolo unde şi drenajul solului este mai slab (zone plane, depresionare cu stagnări de apă din precipitaţii). Cele mai mari suprafeţe de nardete din zona depresionară şi de podiş a judeţului se găsesc pe aceste soluri de regulă asociate cu solurile pseudogleice. Soluri din seria podzolirii humico-feriiluviale 1. Soluri brune acide, tipice, litice, gleizate, se întâlnesc în zona montană, în zona colinară şi pe conurile de dejecţie ale văilor ce coboară din munţii Făgăraş spre depresiunea cu acelaşi nume. Altitudinal sunt frecvente între 700 şi 1400 m, pe culmi şi versanţi de regulă nordici.
S-au format pe roci acide: şisturi cristaline (munţii Făgăraş, Perşanii sudici, Piatra Craiului, Perşani); gresii şi conglomerate silicioase (munţii Ciucaş, Bârsei, Bucegi); pe depozite de dezagregare-alterare ale unor roci carbonatice, mai puţin strate Sinaia (pe platformele Bran şi Poiana Mărului).
Vegetaţia caracteristică arealului acestor soluri: molidişurile, făgete pure şi în amestec cu răşinoase, însoţite de o floră acidofilă, de muşchi verzi, mai puţin de Vaccinium myrtillus.
Sunt soluri cu textură grosieră (nisipo-lutoasă), mijlocie (luto-nisipoasă), argila fiind sub 30%; moderat puternic acide (pH4,3-5,5), oligobazice-oligomezobazice (V 11-55 %), conţinutul în humus variabil (5-25 %) în general legate în materie organică slab descompusă spre limita superioară a arealului, conţin Fe2O3 şi Al2O3 liber dar migrarea lor spre orizontul B este slabă, cu cât ea se accentuează solul are caractere sporadice evidente. Fertilitatea acestor soluri este mijlocie şi scăzută în limita superioară a arealului. În depresiunea Făgăraş, solurile brune acide sunt ocupate mai ales de terenurile arabile.
2. Solurile brune podzolice, tipice, umbrice, criptopodzolice, litice, gleizate, turboase.
Se întâlnesc în limita superioară a zonei forestiere montane în molidişuri şi în etajul alpin inferior sub jnepenişuri şi ienuperete, pe roci acide şi substrate sărace în minerale calcice şi feromagnetice în climat rece şi umed.
Sunt soluri cu textură grosieră (nisipo-lutoasă) şi mijlocie (luto-nisipoasă), puternic-foarte puternic acide (pH < 5), oligobazice (V < 40 %), conţinutul în humus ridicate (10-25 %) ca şi de materie organică slab descompusă. Fertilitatea este mai scăzută ca cea a solurilor brune acide, factorii ecopedologici limitativi fiind clima şi însuşirile chimice ale solului.
3. Podzoluri tipice, friiluviale, brune, litice, gleizate, turboase. Se întâlnesc spre limita superioară a subetajului molidului şi în etajul alpin
al jnepenişurilor, la altitudini mai mari de 1700 m, pe culmi largi, platforme, versanţi slab înclinaţi, pe roci acide silicioase (şisturi cristaline, gresii, conglomerate) pe depozite eluvio-deluviale debazificate.
Sunt soluri cu textură grosieră (nisipo-lutoasă), puternic şi foarte puternic acide (pH < 4,5), oligobazice (V < 30 %), conţinut ridicat de materie organică slab descompusă, săracă în azot, calciu şi magneziu, cu activitate biologică scăzută. Fertilitatea naturală este foarte scăzută.

41
Condiţiile pedoclimatice din arealul solurilor din seria podzolirii humico-feriiluviale sunt favorabile dezvoltării şi extinderii nardetelor în pajişti şi pe terenurile defrişate.
În cuprinsul judeţului Braşov, în afara solurilor caracterizate mai sus, se întâlnesc cernoziomuri tipice, levigate şi argilice (în depresiunea Braşov şi podişul Târnavelor) cultivate aproape în totalitate datorită fertilităţii ridicate, rendzine (pe piemontul Săcele şi în zona montană pe calcare), pseudorendzine (pe podişul Târnavelor).
Apariţia solurilor azonale este determinată de un factor pedoecologic predominant: umiditate crescută (soluri hidromorfe-lăcovişti, rendzine) relief şi rocă (regosoluri, litosoluri, soluri aluviale).
Dată fiind importanţa mai mică pe care o au aceste soluri pentru nardete, ele nu se mai descriu în lucrarea de faţă.
2.1.5. Aspecte privind flora şi vegetaţia Poziţia geografică, particularităţile formelor de relief şi condiţiile
pedoclimatice au determinat, în decursul timpului, dezvoltarea unei vegetaţii deosebit de variate pe teritoriul judeţului Braşov.
Amplitudinea mare de relief, de peste 2000 m, a constituit factorul hotărâtor al etajării vegetaţiei actuale. Printre alte particularităţi, expoziţia nordică şi înclinarea mare a versanţilor Bucegilor, Pietrei Craiului şi Făgăraşului, au determinat unele particularităţi neîntâlnite în alte masive muntoase.
Vegetaţia alpină ocupă culmile munţilor înalţi (Bucegi, Făgăraş) cu climat vitreg şi perioadă de vegetaţie scurtă (3-4 luni). În astfel de staţiuni apar caracteristicile tufişurilor scunde de plante lemnoase ca: Loiseleuria procumbens, Salix reticulata, Salix retusa, Salix herbacea. Speciile ierboase adeseori pajişti întinse (Festuca airoides, Agrostis rupestris, Carex curvula şi altele) ori perniţe (Primula minima, Silene acualis, Minuartia sedoides etc).
Vegetaţia subalpină se situează deasupra pădurilor de molid, mai sus de 1700-1800 m, în masivele muntoase ce constituie graniţa sudică a judeţului Braşov. Speciile lemnoase sunt bine reprezentate de Pinus mugo, Juniperus sibirica, Rhododendron myrtifolium, Vaccinum myrtillus ş.a. pajiştile care apar ca vegetaţie secundară sunt dominate de Nardus stricta, Festuca airoides, Agrostis rupestris şi uneori de Festuca rubra.
Pădurile sunt bine reprezentate, fiind mult mai restrânse decât în trecut, când acopereau aproape în întregime teritoriul judeţului, excepţie făcând doar crestele alpine şi abrupturile.
Molidişurile (păduri de Picea excelsa) ocupă partea superioară a zonei forestiere, între 1200-1700 m, adeseori coborând însă până la 700-800 m pe versanţii nordici. Spre limita inferioară molidul intră adeseori în amestec cu fagul.
Pajiştile bine întreţinute din etajul molidului sunt în cea mai mare parte ocupate de Festuca rubra, care prin degradare se transformă în nardete.
Făgetele ( păduri de Fagus silvatica) se desfăşoară normal între 800-1200 m, dar ele urcă uneori pe versanţii favorabili până la 1350-1500 m (Bucegi, Făgăraş) sau coboară alteori la 600 m pe versanţii nordici ai Postăvarului sau ai

42
Perşanilor. Pe versanţii vestici ai Postăvarului spre Poiana Braşov pe creste domină fagul, iar mai jos până la 600 m coboară bradul.
Gorunetele (păduri de Quercus petraea) se amestecă la altitudini mici (500-600 m), cu stejarul şi uneori cu carpenul, iar spre limita superioară (600-800 m) cu fagul. Pe versanţii însoriţi ai Perşanilor, gorunul urcă până la 1000 m, în timp ce fagul coboară pe versanţii umbriţi şi pe văi, sub gorun. Pe Măgura Codlei se întâlnesc adeseori exemplare izolate, vârstnice de gorun la cca. 1000 m în plină pădure de fag (MORARIU şi colab. 1970). Au fost semnalate situaţii în care gorunul este substituit de brad (STĂNESCU 1958).
Pajiştile care s-au instalat după defrişarea pădurilor de fag şi gorun sunt în marea lor majoritate compuse din Festuca rubra şi Agrostis capillaris, graminee care domină în covorul ierbos. Prin degradarea acestor pajişti se ajunge la nardete de altitudine mai joasă, forte bine reprezentate în judeţul Braşov (depresiunea Făgăraş şi Vlădeni).
Stejeretele (păduri de Quercus robur), care aveau răspândirea naturală în depresiuni, au fost în cea mai mare parte defrişate în vederea lărgirii terenurilor agricole. Resturi ale acestor păduri, întinse odinioară, se găsesc în apropierea Prejmerului, pădurea dintre Crizbav şi Satu Nou, vestita pădure de la Poiana Narciselor. Stejerete mai apar insular şi în sud-estul Podişului Transilvaniei, în luncile râurilor din dealurile dintre Rupea şi Făgăraş.
În locul stejeretelor, după defrişare, s-au instalat cele mai valoroase pajişti din judeţul Braşov, care cuprind în componenţa lor speciile: Arrhenatherum elatius, Trisetum flavescens, Poa pratensis, Poa trivialis, Festuca pratensis şi altele la care adăugăm şi Agrostis capillaris, deseori dominantă în covorul ierbos. În puţine cazuri se întâlnesc şi nardete (Poiana Narciselor, Lunca Oltului).
Pe versanţii însoriţi, pe dealurile aparţinând îndeosebi Podişului Transilvaniei apar fitocenoze relativ restrânse cu vegetaţie xerofilă, în care apar Festuca valesiaca, Festuca rupicola, Botriochloa ischaemum, Stipa capillata, Carex humilis ş.a.
Depresiunile din lunca Oltului (Ţara Bârsei şi Făgăraşului), pe lângă faptul că sunt zone cu altitudinea cea mai mică din cuprinsul judeţului, prezintă şi unele particularităţi sub raport floristic şi de vegetaţie, prin faptul că au conservat în decursul mileniilor câteva relicte glaciale valoroase.
Posibilităţile menţinerii în mlaştinile eutrofe a unor elemente rare, a fost asigurată de particularităţile apei izvoarelor (reci şi cu puţin CaCo3), ce brăzdează pajiştile pe zeci de hectare.
Menţionăm dintre rarităţile floristice ale mlaştinilor următoarele specii: Armeria alpina ssp. barcensis - endemism al Tării Bârsei (Hărman-Prejmer), Pedicularis sceptrum carolinum, Primula farinosa, Lysimachia thyrsiflora, Cnidium dubium, Cladium mariscus, Sweertia perennis, Lingularia sibirica, Carex davalliana, Comarum palustre, Valeriana simplicifolia, Drosera anglica, etc.
În judeţul Braşov se mai întâlnesc numeroase alte specii de plante rare şi endemisme cum sunt: Dianthus callizonus, Linaria alpina, Saxifraga demissa, Daphne cneorum în Piatra Craiului; Hepatica transsilvanica, Bromus barcensis, Coronilla coronata pe Tâmpa Amygdalus nana, Stipa pulcherrima, Carex humilis

43
pe Lempeş Sâmpetru, Narcissus stellaris la Dumbrava Vadului şi multe alte specii care sunt ocrotite în cele 13 rezervaţii floristice, fiind declarate monumente ale naturii (MORARIU şi colab. 1971).
În raionarea floristică, conform concepţiei BORZA şi BOŞCAIU, 1965, judeţul Braşov aparţine regiunii eurosiberiene, provinciei central-europeană est-carpatică ce cuprinde circumscripţiile: masivul Bucegilor şi Munţii Bârsei; Harghita - Perşani; Carpaţii Meridionali cristalini (Munţii Făgăraşului) şi Podişul Transilvaniei (Podişul Târnavelor). Această clasificare are în vedere în mod deosebit speciile relicte şi endemice care dezvăluie istoria vegetaţiei, fără a se lua în considerare etajele de vegetaţie sau substratul geologic, după cum afirmă, de altfel şi autorii ei.
2. 2. OBIECTIVELE CERCETĂRII ŞI METODA DE LUCRU
Răspândirea nardetelor pe suprafeţe întinse în numeroase ţări cu relief
muntos din Europa, ASIA, Africa de Nord şi alte areale ale globului, au atras atenţia a numeroşi oameni de ştiinţă care de peste un secol au adus o contribuţie însemnată la cunoaşterea şi îmbunătăţirea acestor pajişti degradate.
Întreg lanţul Munţilor Carpaţi la care se adaugă dealurile şi depresiunile ce îi însoţesc pe teritoriul ţării noastre, sunt prin excelenţă acoperiţi de întinse suprafeţe de pajişti, în care nardeteele sunt bine reprezentate. De aceea, sunt puţine descrieri ale vegetaţiei munţilor noştri şi a dealurilor cu climat mai umed în care să nu fie remarcate nardetele.
Studiile geobotanice asupra nardetelor, deşi numeroase şi temeinic efectuate în majoritatea lucrărilor, sunt destul de heterogene datorită metodelor diferite de caracterizare şi clasificare adaptate de majoritatea autorilor. S-a ivit astfel problema sistematizării acestor rezultate după o metodă unitară, obiectiv luat în studiu şi în lucrarea de faţă.
În ceea ce priveşte caracterizarea tipurilor de nardete care să servească nevoilor imediate ale producţiei se constată că cercetările sunt abia la început. Continuarea şi adâncirea studiilor de tipologie care să înglobeze şi elemente de îmbunătăţire a nardetelor a stat în atenţia acestei lucrări. De asemenea, sunt mai puţin cunoscute relaţiile şi raporturile dintre fitocenoze şi tipurile de pajişti în scopul asimilării studiilor geobotanice efectuate până în prezent pentru a fi mai accesibile în condiţii de producţie.
Îmbunătăţirea şi folosirea nardetelor a fost destul de bine cercetată, îndeosebi în zona de munte. În această direcţie dispunem de date mai puţine pentru nardetele din zona de deal şi depresionară. Câteva probleme privind îmbunătăţirea nardetelor situate la altitudini mai joase şi-a propus să abordeze şi această lucrare. În final stabilirea randamentului în producţie animală a nardetelor îmbunătăţite se impune ca o necesitate în concluzionarea mai precisă a celor mai eficiente mijloace de îmbunătăţire.
Aceste câteva considerente au stat la baza iniţierii unui studiu unitar, geobotanic şi tipologic cu implicaţii de ordin practic asupra nardetelor din judeţul

44
Braşov, unde pe o suprafaţă relativ restrânsă sunt cuprinse principalele fitocenoze şi tipuri edificate de specia Nardus stricta în ţara noastră.
2.2.1. Principii generale Principiile şi metodele de cercetare a covorului ierbos edificat de specia
Nardus stricta au evoluat de-a lungul timpului. Autorii acestor studii au adoptat adeseori principii diferite de caracterizare şi clasificare a nardetelor, ceea ce creează unele dificultăţi în sintetizarea şi generalizarea rezultatelor. în studiile geobotanice, punctele de vedere sunt controversate datorită faptului că nardetele nu au specii caracteristice proprii care să satisfacă pe deplin sistemul de clasificare al asociaţiilor vegetale după principiile şcolii floristice (BRAUN-BLANQUET, 1964; BOŞCAIU, 1971).
Pe de altă parte absolutizarea dominaţiei ca unic criteriu de apreciere şi clasificare a fitocenozelor de Nardus stricta s-a dovedit în cele din urmă deficitar, fiind adeseori lipsit de posibilitatea conturării unor însuşiri ecologice proprii de diferenţiere faţă de alte fitocenoze (BELDIE, 1967).
În ultima vreme, dezvoltarea şi adâncirea cercetărilor de ecologie a speciilor şi a influenţei factorilor staţionali asupra vegetaţiei, au creat numeroase premise de ieşire din acest impas. Caracterizarea fitocenozelor este tot mai mult legată de condiţiile staţionale şi a influenţelor exercitate de diferiţi factori ecologici, care sunt conturaţi şi determinaţi adesea prin observaţii şi măsurători riguroase.
Pe aceste baze noi, în lucrarea de faţă se încearcă un studiu geobotanic al nardetelor judeţului Braşov.
În ceea ce priveşte tipologia nardetelor, există un consens unanim în încercările făcute până în prezent şi anume acelea de a stabili principii şi metode de investigaţia care să servească nevoilor imediate a producţiei.
Studiile privind tipologia nardetelor sunt puţine la noi în ţară, preocupările în acesată direcţie s-au intensificat abia în ultimii ani, după principii relativ unitare în comparaţie cu studiile geobotanice.
Recent a apărut o direcţie nouă în ceeace priveşte studiul pajiştilor şi anume acela al bonitării, care încearcă să sintetizeze toate datele acumulate asupra structurii ecosistemului (fitocenoză+habitat), să evalueze ecologic şi agronomic factorii determinanţi ai bioproductivităţii, în vederea gospodăririi raţionale a acestor resurse naturale.
De remarcat, că între aceste trei direcţii de studiere a vegetaţiei pajiştilor, geobotanic, tipologic şi al bonitării, există o strânsă interdependenţă întrucât se bazează pe metode de lucru comune asupra fitocenozelor şi factorilor staţionali. Fără studiul geobotanic nu poate fi continuat cel tipologic. Împreună aceste metode de lucru dar mai ales studiul tipologic este elementul de bază al bonitării pajiştilor.
Diferenţa dintre aceste direcţii mari de studiu al vegetaţiei şi staţiunii constă în diversificarea şi aprofundarea unor laturi şi elemente care să servească în final obiectivelor dinainte stabilite.
În această lucrare pe lângă studiul geobotanic şi tipologic se face bonitarea nardetelor cu datele din teren, adunate înainte de apariţia "Instrucţiunilor de bonitare a pajiştilor - Braşov, 1978" elaborate de SCCP Braşov, IANB şi ICPA

45
Bucureşti. Pentru acest considerent o parte din elementele staţionale ale solurilor necesare bonitării nardetelor sunt extrase din diferite lucrări de sinteză şi literatura de specialitate.
2.2.2. Studiul geobotanic Unitatea de bază în studiul geobotanic o constituie "Asociaţia vegetală"
stabilită la al III-lea Congres de botanică de la Bruxelles în 1910, unde FLAHAULT şi SCHRŐTER o definesc ca "o întovărăşire de plante cu o anumită compoziţie floristică, prezentând o fizionomie uniformă şi vegetând în condiţii staţionale uniforme" la care BRAUN - BLANQUET 1913, precizează că "asociaţia posedă una sau mai multe specii caracteristice" (citat după BORZA şi BOSCAIU, 1965).
Întemeietorul şcolii fitosociologice - floristice BRAUN - BLANQUET, 1928, 1964) descrie pentru prima dată asociaţia Nardetum strictae alpinum pe care o încadrează în alianţa Nardion Br-Bl 26, Ordinul Caricetalia curvulae Br-Bl 26, clasa Caricetea curvulae Br-Bl 48, cu răspândirea în Alpi, Pirinei, Carpaţi, Apenini, Balcani, etc.
PREISING 1949 şi OBERDORFER 1957,1959 completaţi de HUNDT 1964 descriu mai multe asociaţii de Nardus stricta în Europa după principiile şcolii floristice la care adaugă şi aspecte ecologice cum sunt distribuţia altitudinală a vegetaţiei primare şi zona geografică care influenţează repartizarea nardetelor pe continentul nostru. Astfel, nardetele din ţara noastră sunt încadrate în alianţa păşunilor şi părul porcului alpine, subalianţa Carpaţilor estici.
OBERDORFER 1957, reuneşte asociaţia de nardete în cinci grupe de asociaţii conturate după altitudine şi regimul de umiditate al staţiunilor, pe care le încadrează în cl. Nardo-Callunetea Prag.49, Ord. Nardetalia Oberd.49, Al Eu- Nardion br.Bl. 26.
ELLENBERG 1963 şi KLAPP 1965 descriu la rândul lor asociaţiile de Nardus stricta după principiile floristico - ecologice elaborate de OBERDORFER 1957 la care adaugă unele detalii care să servească scopurilor practice de îmbunătăţire a acestor pajişti.
Şi la noi în ţară principiile şi metodele de cercetare a covorului ierbos au evoluat de-a lungul timpului de asemenea şi cele referitoare la nardete.
O primă direcţie de studiu a nardetelor a fost în concepţia şcolii Zűrich-Montpellier ale cărui principii au fost adoptate prima oară la noi de către BORZA 1934, continuate până în prezent de BOŞCAIU 1971, DIHORU 1975 şi alţii. Adepţii şcolii floristice analizează şi clasifică vegetaţia după speciile caracteristice, fidele asociaţiei, alianţei, ordinului şi clasei, precum şi a diferenţelor ecologice pentru subasociaţii, a dominaţiei pentru facies-uri şi alte criterii. În concepţia acestor şcoli, obiectivul de bază îl constituie clasificarea şi încadrarea comunităţilor vegetale după principii fitosociologice.
A doua orientare majoră în studiul vegetaţiei, are la bază criteriul dominanţei după modelul şcolii sovietice, adoptat de către Secţia de geobotanică a Institutului de Geologie. Ideile principale ale acestei orientări au fost prezentate de către SERBĂNESCU 1960, la prima consfătuire naţională de geobotanică. După

46
acest principiu, dominanţa unei specii sau a unui grup de 2-3 specii pot fi considerate ca edificatoare pentru o asociaţie, grup de asociaţii (alianţă) şi formaţiuni vegetale (ordin-clasă). La aceste principii au aderat numeroşi pratologici (ANGHEL şi col. 1965, 1977; BARBULESCU,1961; BURCEA, 1965; GRINEANU, 1967; NIEDERMAIER, 1968; ŞTEFĂNESCU, 1970; MOTCĂ, 1972; MARUŞCA, 1982 şi alţii), precum şi geobotaniştii (PUSCARU-SOROCEANU şi col. 1963; PAUCA, 1960; CIUCA, 1963 şi alţii), care au adus şi unele îmbunătăţiri, prezentând lista speciilor pe grupe economice după principii mai mult tipologice (graminee, leguminoase, ciperacee + juncaceae, alte familii, muşchi + licheni). Diagnoza fitocenozei se face prin alăturarea speciilor dominante şi codominante, de exemplu: Nardus stricta + Agrostis tenuis - Paucă 60, în scopul accesibilităţii pentru practica agricolă.
În altă direcţie de studiu a vegetaţiei şi implicit a nardetelor, s-a preluat criteriul fitocenologic al speciilor dominante edificatoare pentru identificarea asociaţiilor, orânduind la fel speciile şi releveuri pe grupe economice, asemănător tipurilor, la care în final se foloseşte nomenclatura fitosociologică a şcolii floristice atât în denumirea asociaţiilor cât şi al celorlalţi cetaxoni de rang superior şi inferior asociaţiei (PUŞCARU şi col. 1956; BUIA şi col. 1962; BUIA 1963; PUIA şi col. 1973 şi alţii).
Ţinând seama de aceste trei orientări, fitosociologic, fitocenologic şi combinaţia dintre ele, în care s-au utilizat principii şi metode diferite pentru studiul nardetelor, din punct de vedere geobotanic în sens larg, se pun următoarele probleme:
au nardetele specii caracteristice-fidele în sensul şcolii floristice ? care este optimum ecologic al speciei Nardus stricta şi al asociaţiilor care
le edifică în condiţiile răspândirilor pe un areal altitudinal atât de vast (230-2200 m altitudine) la noi în ţară ?
în ce unităţi superioară (alianţă, ordin, clasă) se integrează asociaţiile de Nardus stricta ?
ce recomandări utile pentru transformarea nardetelor în pajişti mai productive, cantitativ şi calitativi, se pot desprinde din studiile geobotanice efectuate până în prezent şi altele.
În ce priveşte existenţa speciilor caracteristice fidele, părerile fitosociologilor de orientare floristică sunt împărţite. Unii autori susţin existenţa speciilor caracteristice (BRAUN-BLANQUET et j. 1926; BARTSCH et j. 1940; OBERDORFER 1957; KLAPP 1965; BORZA 1959; BOSCAIU 1971; DIHORU 1975 şi alţii), mai ales pentru nardetele situate la altitudini din etajul subalpin şi uneori pentru cel montan.
Alţii contestă existenţa unor specii caracteristice - fidele, remarcând existenţa unor specii transgresive, rămase din alte asociaţii pe care le-a invadat specia Nardus stricta (PUSCARU şi col.1956; BELDIE 1967).
Sunt autori care au contestat caracterul independent al asociaţiilor de Nardus stricta considerându-le facies-uri din lipsă de specii caracteristice (BUIA 1943), ca apoi să revină asupra acestui punct de vedere, confirmând existenţa asociaţiilor edificate de Nardus stricta (BUIA şi col. 1962; BUIA 1963).

47
Au existat deci şi există păreri diferite în ceea ce priveşte existenţa speciilor caracteristice-fidele pentru nardete, o apropiere din acest punct de vedere se remarcă numai la nardetele situate în etajele alpin-subalpin, parţial pentru cele din etajele molidului şi fagului şi puţine precizări deocamdată în etajul gorunului şi zona stejarului.
Condiţiile staţionale au un rol determinant în răspândirea nardetelor. Optimum ecologic al speciei Nardus stricta este reprezentat de soluri acide şi foarte acide, puternic debazeificate, sărace în elemente fertilizante, situate în etajul alpin-subalpin până la zona colinară-planarp. Din punct de vedere al temperaturilor nardetele sunt mai puţin exigente fiind întâlnite în zone cu 0-8 0C media anuală, pe pante preponderent însorite la altitudini ridicate şi pe toate expoziţiile la altitudini mai joase. La fel şi umiditatea joacă un rol mai puţin important fiind remarcate xeronardetele şi hygronardetele. Totuşi regimul precipitaţiilor medii anuale depăşesc 600 mm, iar solurile sunt temporar sau permanent umede.
Persistenţa nardetelor este favorizată şi de păşunatul moderat cu animalele îndeosebi cu ovinele. Aproape fără excepţie autorii străini şi autohtoni, stabilesc că răspândirea speciei Nardus stricta depinde în primul rând de condiţiile edafice şi apoi de cele climatice şi antropogene, factori ce se găsesc în strânsă interdeoendenţă.
Aceste observaţii asupra ecologiei nardetelor cu cortegiul de specii caracteristice, aduc multe clarificări în direcţia clasificării lor.
Referitor la încadrarea asociaţiilor de Nardus stricta în unităţi superioare (alianţă, ordin, clasă) pe baza lucrărilor elaborate până în prezent se conturează câteva concluzi.
La nivelul alianţei, primele nardete descrise în etajele alpine - subalpine, respectiv Nardetum alpinum Br-Bl 26 şi Nardetum alpigenum Br-Bl 49 au fost încadrate în alianţe Nardion strictae Br-Bl 26 sau Eu-Nardion Br-Bl 26, ordinul Caricetalia curvulae Br-Bl 48, clasa Caricetea curvulae Br-Bl 48.
Ulterior, pentru nardetele submontane din Germania s-a conturat alianţa Nardo-Galion (saxatilis) PREISG 49, adoptată de OBERDORFER 1957; KLAPP 1965; ROTHMALER 1970 şi alţii. Această alianţă Nardo-Galion împreună cu alianţa Eu-Nardion au fost încadrate în ordinul Nardetalia Preisg 49, clasa Nardo-Calunetea Preisg 49, (Pajişti europene de părul porcului şi tufărişuri pitice).
Este pentru prima dată când nardetele alpine, subalpine, montane, submontane şi planare din vestul şi centrul continentului nostru, sunt âncadrate în acelaşi ordin şi clasă de vegetaţie.
La noi în ţară primele nardete descrise de BORZA 1934, în Retezat au fost încadrate în alianţa Nardion strictae Br-Bl 26, ordinul Caricetalia curvulae Br-Bl 26, clasa Caricetalia curvulae Br-Bl 26, punct de vedere menţinut şi în descrierea nardetelor din Valea Sebeşului (BORZA 1959).
Pentru nardetele din masivul Bucegi, PUŞCARU şi col 1956 contestă apartenenţa lor la ordinul Caricetalia curvulae Br-Bl 26, considerându-le că aparţinând unei formaţiuni (ordin) distincte, Nardatalia strictae în care pot fi deosebite mai multe grupe de asociaţii (alianţe) şi anume: Nardion strictae alpinum, Nardion strictae montanum şi Nardion strictae campestre.

48
După aceleaşi principii au fost studiate şi clasificate nardetele din Masivul Parâng de către BUIA, PĂUN şi PAVEL 1962, unde se afirmă apartenenţa nardetelor la ordinul Nardetalia cu două alianţe Nardion strictae alpinum şi Nardion strictae montanum existente în zona luată în studiu.
BOSCAIU 1971, apreciază încă utilă menţinerea denumirilor de Nardetum strictae alpinum şi Nardetum strictae montanum, (PUŞCARU et al.56, BUIA et al. 62 non SILLINGER 33) pentru sublinierea repartiţiei altitudinale a acestor pajişti şi mai ales a unor indici distincţi de productivitate a biomasei. Din punct de vedere cenotaxonomic această nomenclatură este puţin satisfăcătoare. Ridicările ambelor tabele publicate de BUIA şi col. 1962, susţine BOSCAIU 1971, aparţin în realitate la alianţa Potentillo(ternatae) – Nardion, SIMON 57 şi este dificil ca în cadrul lor să se recunoască existenţa a două asociaţii şi cu atât mai puţin a două alianţe distincte (Nardion strictae montanum - PUŞCARU et al. 56 şi Nardion strictae alpinum PUŞCARU et al. 56).
Astfel, BOSCAIU 1971 încadrează nardetele din Carpaţii sud-estici, în alianţa Potentillo-Nardion, dar le menţine în ordinul Caricetalia curvulae Br-Bl 26 şi le introduce în clasa Juncetae trifidi - HADAČ et KLIKA 1944 (Pajişti din tufărişuri scunde de pe soluri alpine, oligotrofe, intens acide).
Nardetele din Muntele Siriu, descrise de DIHORU 1975 sunt de asemenea încadrate în alianţa Potentillo(ternatae)-Nardion, dar spre deosebire de BOSCAIU 1971, apreciază mai justă apartenenţa lor la ordinul Nardetalia, clasa Nardo-Callunetea. În aceeaşi alianţă, ordin, clasă, adoptate de DIHORU 1975, sunt încadrate şi nardetele din Bazinul superior al Motrului (MALOS 1977).
Cu toate observaţiile şi precizările făcute de către BOSCAIU 1971 şi DIHORU 1975, cu privire la încadrarea nardetelor montane în alianţa Potentillo-Nardion, în unele lucrări mai recente (DUMITRU 1980; STEFAN 1980), se continuă încadrarea lor după principii mai mult tipologice în alianţa Nardion strictae montanum propusă de PUŞCARU şi col. 1956.
O problemă aparte o prezintă încadrarea nardetelor din etajele de vegetaţie situate la altitudini mai joase, mai ales la nivelul de alianţă. Până în prezent la noi în ţară aceste nardete au fost încadrate adesea în alianţa Nardion strictae collinum de către numeroşi autori (TURCU 1968; PĂUN şi col 1973; DUMITRU 1980, şi alţii). Dacă se consideră nepotrivită din punct de vedere cenotaxonomic alianţa Nardion strictae campestre (collinum) Puşcaru et col. 56, Buia 56, Buia et col. 62, având mai mult un caracter tipologic pentru departajarea nardetelor de joasă altitudine de cele montane şi alpine, atunci este oportună a se stabili alte formule de încadrare conform cu principiile şcolii floristice. Acest aspect este relevat şi de MORARU şi DANCIU 1977 care precizează că în conformitate cu articolul 34 din Codul de nomenclatură fitosociologică trebuiesc îndepărtate din denumirea cenotaxonilor unele epitete ce indică o particularitate geografică şi bineînţeles una altitudinală. Astfel, este descrisă asociaţia Thymo (comosi)-Caricetum humilis, în Ţara Bârsei care înlocuieşte pe cea descrisă mai înainte şi anume: Caricetum humilis transylvanicum -Zolyomi 34. În cazul acesta specia Thymus comosus este caracteristică pentru asociaţie, înlocuind numele mai vag de transylvanicum.

49
În cazul nardetelor, mai multe denumiri care au apărut şi în literatura noastră de specialitate ca de exemplu: xeronardetum, hygronardetum, alpinum, subalpinum, montanum, submontanum, campestre, collinum, austro-carpaticum, biharicum, siculum, marmarosicum, nudum, etc., după noile îndreptări nomenclaturale trebuiesc reformulate alăturând denumirii de Nardion sau Nardetum o specie caracteristică care să indice şi să înglobeze particularităţile geografice altitudinale, staţionale şi altele.
Pentru încadrarea nardetelor submontane şi planare, aşa cum s-a mai amintit, OBERDORFER E. 1957 şi alţii descriu pentru Germania alianţa Nardo - Galion (saxatilis) Preisg 49 pe care o consideră sinonimă cu Violion caninae Schwick 44. În Cehoslovacia MORAVEC 1965, include nardetele colinare în alianţa Violion caninae pe care o asimilează cu Violon(caninae)-Nardion Oberd 49.
De asemenea, ELENBERG 1978, încadrează nardetele joase din Europa Centrală în alianţa Violo-Nardion.
BOSCAIU 1971, citează că pentru nardetele colinare s-a propus delimitarea alianţei Nardo-Agrostion (tenuis) Sillinger 33 pe care KRAJNA 1933 a relatat-o la ordinul Arrhenatheretalia iar ulterior SOŐ 1964 la Nardetalia. Această apartenenţă este reconfirmată de SOŐ 1973 care include nardetele de joasă altitudine din Ungaria în alianţa Nardo-Agrostion şi Violion caninae, încadrând-o la Nardetalia, clasa Nardo-Callunetea.
Şi la noi se descriu nardetele premontane din Munţii Perşani, jud. Braşov (ULARU 1972), ca aparţinând alianţei Violion caninae.
Urmărind o transectă din vestul spre centrul şi estul continentului european, se constată ca nardetele colinare şi planare sunt încadrate de majoritatea autorilor în alianţa Violo (caninae)-Nardion Oberd.49 sau altre alianţe considerate sinonime cu acestea.
Din această enumerare de puncte de vedere privind apartenenţa asociaţiilor de Nardus stricta, în urma unei analize amănunţite, apreciem că nardetele din judeţul Braşov, reprezentative pentru întreg lanţul carpatic românesc se pot încadra în alianţele:
Potentillo(ternatae) – Nardion, Simion 57 (pentru nardetele subalpine –montane); Violo (caninae) – Nardion Oberd 49 (pentru nardetele colinare planare). Ambele alianţe le includem în ordinul Nardetalia Preisg 49, care aşa cum s-
a arătat este aproape unanim acceptat în ultimul timp, dată fiind răspândirea nardetelor pe o amplitudine altitudinală atât de vastă la noi şi în alte ţări.
În ceeace priveşte clasa de vegetaţie din aceleaşi considerente de răspândire a nardetelor, acceptăm clasa Nardo-Callunetea Preisg 49, în locul aceleia de Caricetea curvulaie Br-Bl 49 sau Juncetea trifidi Hadac et Klika 44. Această opţiune pentru încadrarea nardetelor în ordinul Nardetalia şi clasa Nardo-Callunetea este susţinută în numeroase teze de doctorat care au avut ca temă studiul vegetaţiei în diferite zone ale ţării noastre, prezentate în centrele universitare din Bucureşti, Cluj, Iaşi şi Timişoara (RESMERIŢĂ 1969; PANAIT 1969; MOLDOVAN 1970; TURCU 1970; SUTEU 1970; ZANOVSCHI 1971;

50
ULARU 1972; PASCAL 1973; MALOS 1977; STEFAN 1980; DUMITRU 1980 şi alţii).
Problema încadrării asociaţiilor edificate de Nardus stricta va crea în continuare unele controverse, deşi în linii generale cercetările din ultimul timp au ajuns la concluzii asemănătoare sau apropiate. Pe măsura acumulării de material faptic asupra nardetelor mai ales a celor situate în etajele de vegetaţie mai joase, colinare şi depresionare, aceste puncte de vedere privind încadrarea acestor asociaţii în unităţi superioare, se vor apropia şi mai mult.
În legătură cu posibilităţile de transformare a nardetelor considerate pe drept ca pajişti degradate, în pajişti mai productive, din studiile fitosociologice după şcoala Zűrich-Montpellier sau fotosociologice după şcoala sovietică şi a combinaţiei dintre acestea, se desprind concluzii pe care le prezentăm în continuare.
Sunt utile pentru cunoaşterea vegetaţiei, toate releveurile floristice indiferent după care metodă au fost întocmite. Din studiile efectuate după şcoala floristică unde se clasifică cenotaxonii după speciile caracteristrice-fidele, importanţă prezintă unele dintre ele, care conturează un etaj de vegetaţie primară, un stadiu de umiditate a solului şi alte lemente staţionale. De asemenea, facies-urile caracterizate după speciile dominante pot fi cu mai multă uşurinţă folosite pentru studiile de îmbunătăţire a nardetelor.
Cercetările asupra vegetaţiei după criteriul dominanţei speciilor sunt mai utile producţiei, întrucât de la prima vedere şi în realitate speciile dominante au ponderea în realizarea fitomasei asupra căreia se acţionează în procesul de îmbunătăţire a nardetelor. Aşezarea speciilor în releveuri, pe grupe economice, după criterii mai mult tipologice, uşurează şi mai mult munca celui care este chemat să intervină pe aceste pajişti, având de la bun început dispoziţie o ordonare a speciilor valoroase furajere (graminee, leguminoase) pe care de fapt caută să le stimuleze, pe unele dintre ele prin diferite procedee, urmate de cele mai puţin valoroase şi buruieni încadrate la ciperacee, juncacee şi alte familii de plante.
În alte ordine de idei, sistemul de notare a abundenţei şi dominanţei speciilor efectuate după scara BRAUN-BLANQUET prin cifre de la 1 la 5, folosită frecvent de adepţii şcolii floristice este mai greu de înţeles la prima vedere, necesitând transformarea lor în procente, mai uşor de interpretat din punct de vedere cantitativ.
Pentru a evita acest inconvenient, se utilizează tot mai mult metoda preconizată de KLAPP-ELLENBERG de apreciere directă în procente de participare a speciilor, care se pretează la interpretări statistice, ecologice, şi mai ales, este uşor accesibilă pentru practicieni.
Aşa cum susţine şi KLAPP 1965, este dificil de realizat un sistem de clasificare a pajiştilor care să corespundă în întregime concepţiei fitosociologice, floristice şi totodată să aibă şi aplicabilitatea agricolă.
Au fost făcute şi alte încercări de adaptare la nevoile producţiei, studii asupra vegetaţiei pajiştilor. Cele mai importante pot fi considerate lărgirea numărului de date cu privire la staţiune (altitudine, expoziţie, înclinaţie, sol, etaj vegetaţie primară, mod de folosinţă, fitomasă utilă, etc.), ca preambul la releveuri,

51
ordonarea speciilor pe grupe economice, notarea în procente a participării speciilor, toate fiind mai mult tipologice, alături de care pentru fiecare specie se indică elementul floristic, bioforma şi apartenenţa cenotică (caracteristica asociaţiei, alianţei, ordinului, clasei, diferenţiată pentru subasociaţie sau fără valoare indicatoare) prezentate după principii fitosociologice.
Aceste îmbinări de metode ale aşezării speciilor în releveuri pe grupe economice, cu posibilităţi de menţionare a elementelor staţionale biocenotice, nu se abat decât formal de la principiile riguroase ale fitocenologiei sau fitosociologiei clasice. În schimb releveul cu fotocenoza prazentată astfel, îşi măreşte considerabil gradul de accesibilitate pentru practica agricolă având în acelaşi timp suficiente elemente de caracterizare fitosociologică şi cenotaxonomică.
În acest sens, cităm lucrările mai vechi elaborate de PUSCARU şi col. 1965, şi BUIA şi col. 1962, unde sunt prezentate releveuri sintetice după principii combinate, fitocenologice-tipologice cât şi lucrări mai noi (PĂUN 1973) ce prezintă releveuri după principii fitosociologice-tipologice. De altfel, un releveu sintetic "fitosociologic-tipologic" complet şi bine prezentat, poate fi cu uşurinţă transformat într-un releveu după principii riguroase fitosociologice, prin renunţarea la ordonarea speciilor pe grupe economice şi reaşezarea lor după caracteristicile asociaţiei, alianţei, ordinului, clasei, etc., prezentate în faţa fiecărei specii pe coloana apartenenţei cenotice.
Procedeul de transformare inversă, este uneori deficitar, întrucât multe din releveele strict fitosociologice nu conţin suficiente date staţionale şi de altă natură care să satisfacă reordonarea releveelor pentru solicitările cât de cât acceptabile ale pratotehnicii.
În lucrarea de faţă, se prezintă releveuri după modelul folosit de PĂUN şi col. 1973 pentru Subcarpaţii Olteniei, cu unele modificări. S-a considerat necesar a se prezenta în paralel pentru comparaţie şi asimilare, participarea speciilor după scara BRAUN-BLANQUET şi în procente după KLAPP, se indică un număr sporit de condiţii staţionale.
Cu toate acestea, studiile asupra nardetelor care se rezumă aproape exclusiv la vegetaţie, considerăm că sunt necesare, dar nu suficiente pentru a lua decizii corecte privind metodele de îmbunătăţire cantitativă şi calitativă a acestor fitocenoze. Este necesar a se ţine seama, pe lângă studiul vegetaţiei, de factorii staţionali (relief, sol, climă, etc.) şi mai ales de posibilităţile de îmbunătăţire, care fac, de altfel, obiectul unei alte direcţii de studiu al vegetaţiei şi anume, tipologia.
În această controversă asupra oportunităţii studiilor despre vegetaţia pajiştilor după principii fitosociologice sau fitocenologice, orientarea mai nouă, include pajiştea într-un agroecosistem în care condiţiile staţionale şi vegetaţia caracterizată după speciile dominante au un rol esenţial în caracterizarea lanţurilor trofice şi productivităţii ecosistemelor (PUIA, SORAN 1979; PÎRVU şi col. 1980).
Nardetele din judeţul Braşov au fost cercetate după metoda geobotanică pe itinerariu, efectuându-se 205 releveuri, pe întreaga perioadă de vegetaţie pentru a cuprinde cât mai complet şi aspectele sezoniere ale compoziţiei floristice.

52
Pe teren având la dispoziţie o hartă la scara 1: 25.000, după parcurgerea unei suprafeţe de pajişti edificate de specia Nardus stricta s-a ales cu cele mai reprezentative porţiuni de câte 100 mp, unde s-a întocmit lista integrală a speciilor, notările asupra vegetaţiei primare din apropiere, expoziţia şi panta terenului, modul de folosinţă actual al pajiştii, lucrările de întreţinere efectuate, constatate în discuţii cu localnicii şi cadrele tehnice a unităţilor agricole din teritoriu şi alte date. După analiza floristică de pe aceeaşi suprafaţă de teren (100 mp) pentru 99 releveuri, s-au luat probe de sol de la adâncimea de 0 – 10 şi 10 – 20 cm cu sonda agrochimică din 25 – 30 locuri dispersate cât mai uniform.
Pentru notările asupra vegetaţiei pe suprafeţe de probă din cele câteva metode de lucru cunoscute care sunt utilizate în prezent pentru descrierea vegetaţiei s-a ales metoda BRAUN-BLANQUET (1928,1964) în ceea ce priveşte aprecierea abundenţei + dominanţei, frecvenţei, etc., cât şi elaborată de KLAPP 1949, cu privire la aprecierea procentuală a participării în biomasă, aranjarea speciilor pe grupe economice etc.
În punctele reprezentative pentru asociaţii, notările asupra vegetaţiei s-au făcut după metoda BRAUN-BLANQUET, pentru o mai uşoară comparaţie cu studiile asupra nardetelor de până acum. Cu toate că cele şase valori ale scării convenţionale BRAUN-BLANQUET pentru aprecierea combinată a abundenţei şi dominanţei au corespondent în procente, totuşi ea nu satisface pe deplin interpretările statistice mai riguroase pentru factorii ecologici, datorită intervalelor prea mari dintre o valoare şi alta, care ajung uneori la 10-15 % aproximaţie. Astfel, că s-a folosit în toate cazurile metoda KLAPP de apreciere directă în procente a participării fiecărei specii în biomasă având posibilităţi ulterioare mai bune de interpretare matematică a factorilor ecologici şi calitativi ai covorului ierbos.
După prelucrarea datelor din teren, respectiv notările asupra vegetaţiei, condiţiile staţionale, probe de sol, s-a trecut la faza de laborator unde au fost făcute determinările chimice, releveuri sintetice, analize comparative, calcule statistice, etc.
Determinarea caracteristicilor chimice ale solului s-au efectuat în laboratoarele Institutului de cercetări pentru pedologie şi agrochimie din Bucureşti şi anume:
- valoarea pH după metoda potenţiometrică cu electrozi de sticlă; - humus (%) după metoda Schollemberger; - azotul total (%) după metoda Kjeldhal; - P2O5 total (mg/ 100 g sol) după metoda nefelometrică; - K2O schimbabil (mg/ 100 g sol) după metoda debazificării cu acid acetic; - Al mobil (mg/100 g sol) după metoda Socolov; - Ca (%) după metoda percolării cu acid acetic. Celelalte însuşiri ale solului cum sunt gradul de saturaţie în baze (V %) şi
capacitatea de schimb cationic (T) au fost determinate prin analize premergătoare (SB; Ah; etc.) şi calculate după diferite formule uzuale.
Caracterizarea însuşirilor chimice ale solurilor s-au făcut după clasele stabilite de diferiţi autori: OBREJANU 1961, pentru pH; CERNESCU 1959 pentru

53
V%; STÎNGĂ pentru Al mobil; DANILIUC pentru Ca % din T; TEACI 1954 pentru humus %; DAVIDESCU 1960 pentru N total; P2O5 total şi K2O schimbabil.
Interpretarea şi valorificarea rezultatelor privind autecologia principalelor specii din nerdatele judeţului Braşov s-a făcut după metoda descrisă în lucrarea lui HUNDT 1966, îmbunătăţită de LAUER 1974. La început s-au întocmit şiruri de variaţie pentru fiecare caracteristică chimică a solului. După aceasta s-a trecut la întocmirea şirurilor de variaţie pentru 17 specii mai răspândite în nardete, atât din punct de vedere al participării cât şi al prezenţei în funcţie de caracteristicile solului. Valorile maxime ale participării şi prezenţei unei specii au fost egalate cu 100 %, faţă de care s-au recalculat celelalte valori mai mici obţinându-se astfel dependenţa relativă.
Pe baza valorilor astfel obţinute, au fost întocmite ecodiagrame în care se reprezintă participarea şi prezenţa relativă a speciilor luate în studiu, faţă de caracteristicile solului nardetelor.
Prin această metodă s-a determinat valoarea indicatoare a câtorva specii care diferenţiază condiţiile ecologice în care se întâlnesc nardetele.
Individualizarea asociaţiilor vegetale s-a făcut după criteriul floristic-ecologic, stabilindu-se pentru fiecare asociaţie o grupă de specii de recunoaştere, care exprimă prin participarea în biomasă şi dependenţa de anumite condiţii staţionale cele mai sigure indicii de diferenţiere.
Pentru conturarea subasociaţiilor s-au ţinut seamă de modificările locale ale factorilor staţionali stabilindu-se prin determinări agrochimice ale solului şi a altor indici ecologici (umiditate, temperatură, etc.) care sunt speciile diferenţiale ecologice cele mai reprezentative. S-au determinat astfel de specii indicatoare de condiţii staţionale pentru fiecare subasociaţie în parte.
Prin stabilirea spectrelor biologice dar mai ales a celor fitogeografice s-au adus completări la criteriile de diferenţiere a fitocenozelor edificate de specia Nardus stricta. Însuşirile biologice, ecologice şi economice ale speciilor au fost adoptate după KOVACS 1979 şi cele fitogeografice după BELDIE 1978, 1979. Aceşti indicatori au fost sintetizaţi pentru fiecare asociaţie edificată de Nardus stricta utilizând diferite metode.
Astfel spectrul bioformelor şi cel fitogeografic a fost întocmit după sistemul adoptat de PANAIT 1969, în care pe lângă raportul numeric şi prezenţă este redată şi acoperirea procentuală, element important pentru fitocenozele dominate de o specie cum sunt nardetele.
Reprezentarea geografică a răspândirii nardetelor în funcţie de expoziţie şi panta terenului (înclinaţie în grade) a fost după metoda utilizată de CIUBOTARU 1974.
În final, indicele de valoare furajeră a fitocenozelor a fost determinat cu ajutorul metodei botanice după formula: Vf = FB + 3/4 B +1/2 M + 1/4R + CF-2T descrisă de NIEDERMAIER 1967.
Datele analitice privind compoziţia floristică, condiţiile staţionale şi elementele economice determinate pentru fiecare releveu în parte, au fost analizate, interpretate şi ulterior prezentate sub forma unor releveuri de sinteză care cuprind indicatoru medii şi limite de variaţie minimă şi maximă.

54
Prin această condensare a datelor s-a căutat o exprimare cât mai concisă a principalelor caracteristici care conturează fitocenozele descrise. Alături de releveurile sintetice se prezintă o listă a staţiunilor unde au fost descrise fitocenozele şi în final o hartă la scara 1 : 250.000 cu răspândirea nardetelor în judeţul Braşov.
2.2.3. Studiul tipologic În studiul vegetaţiei pajiştilor, noţiunea de tip fâneaţă (STEBLER şi
SCHRŐTER, 1893) a precedat definirea noţiunii de asociaţie vegetală (Congresul de botanică de la Bruxelles din 1910), astfel că cele două unităţi de bază au fost utilizate în paralel de-alungul timpului până azi.
Întemeietorii metodei de cercetare tipologică în domeniul pajiştilor (STEBLER şi SCHRŐTER) definesc tipul de fâneaţă ca fiind acele subunităţi locale care se identifică prin una sau mai multe dominanţe, sau printr-o specie foarte caracteristică pentru specia dominantă.
Definiţia astfel formulată pune în discuţie existenţa unor suprapuneri între noţiunea de tip de pajişte şi asociaţie vegetală în cazul în care aceasta din urmă se caracterizează după speciile dominante edificatoare, aşa cum procedează unele şcoli geobotanice. Aceste tipuri determinate după speciile dominante şi alte indicii fitocenologice sunt corespondente cu asociaţiile sau sociaţiile iar uneori facies-ul (ELLENBERG 1963).
Desigur în tipologie se face apel la cunoştinţele privind descrierea vegetaţiei după principiile geobotanicii, dar aceasta nu este suficient. Spre deosebire de asociaţie, noţiunea de tip înglobează şi alte detalii care servesc cu deosebire practicii imediate cum sunt productivitatea, calitatea, mijloacele de îmbunătăţire etc., KLAPP 1965, menţionează că este greu de realizat un sistem de clasificare a asociaţiilor de pajişte care să corespundă în întregime concepţiei fitosociologice floristice şi tot odată să aibă şi aplicabilitatea agricolă.
Din aceste considerente, acest autor preconizează o clasificare în care asociaţiile sunt denumite după nomenclatura şcolii floristice, fiind apoi grupate după modul de folosinţă, potenţialul de producţie, fertilitatea solului.
Un punct de vedere apropiat este susţinut şi de MORAVEC 1965, care stabileşte asociaţiile de pajişti după principiile şcolii floristice pe care le împarte apoi în două grupe mari: pajişti fertilizate şi nefertilizate. Fiecare din aceste grupe se subîmpart după regimul de umiditate al solului şi vegetaţia primară (asociaţiile de pădure), respectiv stadiul de climax după CLEMENTS 1938.după cum era de aşteptat tipologia ca ştiinţă şi-a găsit un larg câmp de aplicabilitate în silvicultură.
Criteriile principale de diferenţiere a tipurilor de pădure rămân cele recomandate de SUKACEV, cu anumite nuanţări originale. Tipul se diferenţiază în funcţie de: compoziţie, productivitate,aspectul arborilor şi calitatea produselor, regenerarea naturală, subarboretul şi pătura ierbacee, caracterele edafice, caracterele climatice şi măsurile silvotehnice (STĂNESCU 1973).
În spiritul acestor principii s-au iniţiat în ţara noastră studii riguroase, concretizate în sinteza "Tipuri de pădure din RPR" (PASCOVSCHI şi LEANDRU 1958). După această primă etapă în silvicultura noastră cercetările s-au amplificat,

55
remarcându-se o tipologie a pădurilor, o tipologie a staţiunilor forestiere şi mai nou se conturează tipologiile terenurilor degradate şi al exploatărilor forestiere (CHIRITA şi col. 1964, 1974; DONITA şi col. 1970; STĂNESCU 1973; PĂUNESCU 1975).
S-a făcut această scurtă digresiune întrucât multe din principiile care stau la baza diferenţierii şi clasificării tipurilor de pădure au fost adoptate pentru tipologia pajiştilor. Dacă avem în vedere că pajiştile cu puţine excepţii (etajul alpin, stepe, lunci, mlaştini, etc) în ţara noastră reprezintă vegetaţia secundară care s-a instalat după defrişarea vegetaţiei lemnoase de către om, este cu atât mai evidentă legătura care există între tipul de pădure şi tipul de pajişte. Un grad şi mai înalt de apropiere există între tipurile de staţiuni ale pădurilor şi pajiştilor care au foarte multe elemente comune.
În ţara noastră, pentru prima dată, noţiunea de tip de pajişte şi caracterizarea vegetaţiei pajiştilor pe baze tipologice a fost făcută de SAFTA 1936. În descrierea tipurilor de fâneţe din judeţul Cluj, întemeietorul tipologiei praticole româneşti, aşează pe prim plan compoziţia floristică determinată după dominanţă+abundenţă şi frecvenţă. La acestea se adaugă aprecierile privind productivitatea şi indicele calitativ, după o clasificare originală. Pentru fiecare tip de fâneaţă se anunţă câteva caracteristici staţionale cum sunt:umiditatea, relieful, uneori expoziţia şi vegetaţia primară. La sfârşitul fiecărei descrieri se indică şi mijloacele de îmbunătăţire.
Pentru clasificarea tipurilor de păşuni din judeţul Timiş, în descrierea condiţiilor staţionale (SAFTA 1938), pe lângă starea umidităţii solului se menţionează şi sărăturarea acestuia. Este demn de remarcat importanţa ştiinţifică şi practică a raporturilor stabilite de SAFTA I. între tipul de păşune, climă şi sol.
În studiul asupra păşunilor din Transilvania (SAFTA, 1943) se arată că "valoarea păşunilor depinde în primul rând de compoziţia floristică care la rândul ei este expresia factorilor climatologici şi ecologici activi în fiecare pajişte".
Ulterior însemnate contribuţii la definitivarea noţiunilor de bază pentru identificarea şi clasificarea tipurilor de pajişti din ţara noastră au fost aduse de ANGHEL 1960, 1967; ANGHEL şi col. 1965, 1971,1975; VASIU şi col.1961; SAMOILĂ 1971; MOTCĂ 1972; LAUER 1974; CERNELEA 1974; NYARADY şi col. 1974; GUS 1976; KOVACS şi col. 1978; SAFTA, GRIGORE şi col. 1979 MARUŞCA 1982 şi alţii.
În aceste lucrări, apărute mai recent sunt respectate în general principiile elaborate de SAFTA 1943, cu câteva completări care adâncesc latura pretabilităţii tipurilor de pajişti la intensificarea şi valorificarea superioară a producţiei pajiştilor.
ANGHEL şi MOTCĂ 1975 definesc tipul de pajişte "ca o unitate de vegetaţie care se caracterizează prin uniformitatea compoziţiei floristice, a condiţiilor staţionale, a productivităţii şi calităţii furajului, a măsurilor de îmbunătăţire şi direcţiilor de evoluţie în funcţie de aceste măsuri ". Această definiţie cuprinde de fapt şi o succintă prezentare a criteriilor care se propun a fi luate în considerare pentru identificarea tipurilor de pajişti.
Pe baza acestor criterii au fost identificate şi tipurile de nardete din lucrarea de faţă. La acesta adăugăm câteva particularităţi impuse de nardete, cum sunt

56
condiţiile staţionale extrem de diversificate şi stadiul de degradare a covorului ierbos care necesită cu precădere intervenţii pentru îmbunătăţirea însuşirilor productive şi de calitate.
A fost introdusă noţiunea de subtip care conturează unele faze ale stadiului de degradare pe baza căruia se aleg variantele de îmbunătăţire.
În cazul tipurilor de nardete se disting subtipuri determinate de volumul edafic al solului şi anume:
a. pe soluri profunde; b. pe soluri scheletice. La acestea adăugăm subtipurile determinate de alţi factori, în principal
biotici cum sunt: c. cu muşuroaie (marghile); d. cu vegetaţie lemnoasă (arborescentă). Pe teren, în practică se întâlnesc subtipuri de nardete determinate de un
singur factor vizibil de degradare cât şi combinaţii ale acestor factori. Stabilirea subtipurilor de nardete, aduc elemente suplimentare în elaborarea
unor tehnologii care urmează a fi aplicate în vederea îmbunătăţirii compoziţiei floristice, a ridicării nivelului productiv şi calitativ al fitomasei utile pe ansamblul unor condiţii staţionale date.
2.2.4. Bonitarea nardetelor În procesul de adâncire a studiilor asupra vegetaţiei şi condiţiilor staţionale,
bonitarea pajiştilor reprezintă o etapă superioară de evaluare a potenţialului actual şi de perspectivă a acestui mod de folosinţă a terenurilor agricole. Obiectivele bonitării şi metoda de lucru sunt clar reprezentate în "Instrucţiunile pentru elaborarea lucrărilor de bonitare a pajiştilor" editată de SCCP Măgurele - Braşov, IANB şi ICPA Bucureşti 1978. Numeroase elemente privind bonitarea terenurilor agricole au fost elaborate pentru prima dată la noi în ţară de TEACI 1970. Pentru pajişti primele cercetări au fost efectuate de BARBULESCU 1970; BĂRBULESCU Viorica 1975; ANGHEL şi col. 1977, cu privire mai ales la exprimarea valorii pastorale după metoda DAGET-POISSONET (1969, 1971).
După stabilirea unei metode unitare de bonitare a pajiştilor pentru exemplificare, recent au apărut primele lucrări cu acest obiectiv nou de caracterizare a tipurilor şi staţiunilor de pajişti (KOVACS şi col. 1978; NEACŞU, KOVACS 1979; ŢUCRA şi col. 1980).
Bonitarea cuprinde două componente principale şi anume: vegetaţia şi staţiunea, indisolubil legate între ele.
Elementele folosite pentru bonitarea vegetaţiei sunt: valoarea pastorală, indicele de umiditate şi producţia actuală.
Pentru determinarea indicelui sintetic de valoare pastorală (Vp) s-au folosit releveurile floristice în care participarea speciilor a fost apreciată după acoperire (metoda procentuală elaborată de KLAPP), în majoritatea cazurilor. De asemenea, pentru comparaţie s-au efectuat 5 releveuri în care participarea speciilor a fost stabilită după frecvenţă (metoda dublului metru după DAGET-POISSONET 1969,

57
în acest din urmă caz valoarea pastorală, ca indice sintetic de calitate a fost calculată după formula:
Cs x Is Vp = x 100
500 în care: Vp = valoarea pastorală;Cs = contribuţia specifică; Is = indicele specific de calitate; 500 = nota maximă care se poate da unei pajişti (indice 5 x 100 prezenţa). Această metodă de determinare a valorii pastorale folosită pentru prima dată de DAGET-POISSONET, este foarte riguroasă solicitând un timp mai îndelungat de lucru pe teren şi calcule mai complicate, fapt ce o recomandă în cazul unor teritorii mai restrânse sau cercetări comparative mai aprofundate. Pentru aceste considerente, determinarea valorii pastorale s-a făcut având ca bază de plecare acoperirea procentuală a speciilor de nardete. A fost utilizată formula:
P1 x Is + P2 x Is +……….Pn x Is Vp =
100 în care: P1, P2 ….Pn = procentul de participare a fiecărei specii din releveu;
Is = indicele specific de calitate pentru fiecare specie în parte. Aprecierea valorii pastorale după acoperirea procentuală are marele avantaj
că este expeditivă şi totodată permite folosirea datelor vechi existente în literatura noastră (Instrucţiuni SCCP, IANB, ICPA, 1978).
Al doilea element component al bonitării vegetaţiei şi anume, indicele de umiditate (U) se calculează numai pentru speciile care au exigenţe specifice faţă de acest factor, astfel că speciile cu mare variabilitate ecologică notate cu X nu au fost incluse în calcul.
S-a utilizat formula: 1 x PU1 + 2 x PU2 +……….9 x PU9 U = PU1 + PU2 +……….PU9
în care: 1; 2; 3;…..9 = gradaţii indicii de umiditate pentru fiecare specie; PU1……. PU9 = suma procentelor de participare a speciilor ce corespund unei gradaţii de umiditate. Ultimul element pentru bonitarea vegetaţiei şi anume producţie actuală a fost determinat prin cosirea suprafeţelor de probă în staţionarele Vlădeni şi Platoul Bucegi cât şi prin estimări pe teren. Producţia actuală (Pa) a fost exprimată în tone masă verde la hectar. În ceea ce priveşte bonitarea staţiunii s-au folosit datele proprii asupra reliefului terenului (pantă, expoziţie, etc.), altitudine, etaj vegetaţie primară, hidrologie şi unele însuşiri ale solului. Pentru determinarea resurselor climatice ale teritoriului, respectiv media anuală a temperaturilor şi precipitaţiilor au fost folosite hărţile la scara 1 : 500.000 elaborate de IMH 1971, pentru stabilirea teritoriilor ecologice omogene.

58
În urma determinării şi calculării unui număr destul de mare de date s-au stabilit notele de bonitare pentru vegetaţie şi pentru staţiune şi s-a estimat în final potenţialul de producţie al nardetelor. Din cele prezentate rezultă că bonitarea pajiştilor este o sinteză a metodei geobotanice (ANGHEL şi col. 1971) a celei pratologice (DAGET, POISSONET 1971; KLAPP 1971) şi a bonitării terenurilor agricole (TEACI 1970). În lucrarea de faţă se încearcă a se stabili corelaţiile care există între fitocenoză (asociaţie, subasociaţie), tip de pajişte şi bonitare (vegetaţie + staţiune), corespunzând unor etape ale procesului de cunoaştere a nardetelor sub aspect geobotanic, tipologic, pratologic şi pratotehnic. 2.3. ECOLOGIA ŞI COMPOZIŢIA FLORISTICĂ A NARDETELOR
Nardetele din judeţul Braşov au o arie largă de răspândire datorită condiţiilor fizico – geografice, pedoclimatice cât şi a celor economice legate de activitatea omului, care au influenţat şi determinat instalarea nardetelor atât pe culmile şi platourile munţilor înalţi, Făgăraş, Bucegi, Piatra Mare cât şi în staţiuni relativ joase, planare şi colinare a depresiunilor Făgăraş, Vlădeni şi altele.
Între aceste două extreme, nardetele prezintă o continuitate remarcabilă, ceea ce face ca pe un spaţiu relativ restrâns să întâlnim principalele asociaţii şi tipuri edificate de specia Nardus stricta în ţara noastră. Factorii ecologici acţionează permanent într-o strânsă interdependenţă şi dinamică. Determinările efectuate asupra constituenţilor chimici ai solului, legaţi de participarea şi prezenţa speciilor din nardete, aduc câteva elemente noi de referinţă asupra ecologiei nardetelor. Aceste determinări împreună cu ceilalţi factori ecologici prezentaţi la metodică, au permis să se contureze valoarea fitocenotică a unor specii mai des întâlnite în nardete, pe baza cărora au fost stabilite asociaţiile, subasociaţiile şi în final tipurile edificate de specia Nardus stricta. Este cunoscut faptul că nardetele sunt prin excelenţă medii mai stabile în raport cu alte comunităţi vegetale ierboase. Specia dominată Nardus stricta are la rândul ei o influenţă determinată asupra constituenţilor chimiei ai solului. Aceştia influenţează în acelaşi timp existenţa altor specii care de cele mai multe ori aparţin fitocenozelor invadate de Nardus stricta . În această situaţie, o analiză a ecologiei speciilor mai răspândite în nardete, privite din punct de vedere al caracteristicilor chimice ale solurilor, conturează în primul rând amplitudinea ecologică a speciei Nardus stricta şi valoarea indicatoare a unor specii cu tendinţe de comportare faţă de factorii din sol. Diagramele ecologice din prezenta lucrare, exprimă cerinţele speciilor faţă de factorii chimici ai solului, în condiţiile în care specia Nardus stricta are un caracter dominant. Compoziţia floristică a nardetelor determinată în cele 205 releveuri, cuprinde cum era de aşteptat, un număr relativ restrâns de specii în comparaţie cu alte fitocenoze. Structura speciilor este foarte heterogenă, date fiind condiţiile staţionale extreme în care se întâlnesc nardetele.

59
Stabilirea aspectelor biologice şi ale celor fitogeografice a completat caracteristicile nardetelor din zona luată în studiu.
2.3.1. Ecologia speciei Nardus stricta L., cu referiri şi asupra nardetelor Răspândirea speciei Nardus stricta de la altitudini de 200 m până la peste
2200m la noi în ţară, ne indică o plasticitate ecologică deosebită. Această însuşire egalată de puţine specii de ierburi, face ca Nardus stricta să fie întâlnită în staţiuni cu condiţii extreme şi să domine în numeroase pajişti.
În Europa nardetele sunt răspândite în toate regiunile muntoase din Pirinei, Alpi, Tatra, Carpaţi, Apenini, Balcani, etc., coborând în văi până la 700 - 800 m alt. (BRAUN – BLANQUET 1964). Nardete întinse se întâlnesc în Africa de Nord în Atlasul Mare la 2700 - 3500 m alt. (JAGER şi col., 1965), cât şi în etajul alpin şi subalpin al Caucazului şi munţilor din Asia Centrală (TAROSENKO 1952). Specia Nardus stricta se întâlneşte şi în alte părţi de pe glob cum sunt India, Noua Zeelandă, Insulele Azore, Groenlanda şi America de Nord, unde probabil este sinantropă (HULTEN 1958).
De ecologia speciei Nardus stricta şi a asociaţiilor care le edifică, s-au ocupat numeroşi autori (COULON 1923; OBREJANU 1941, 1957; PUSCARU şi col., 1956; CHADWICK 1960; RESMERITA şi col., 1963; KING şi NICHOLSON 1964; BARBULESCU VIORICA 1975;MONTARD şi GACHON 1976) şi alţii care au stabilit factorii care contribuie la răspândirea şi dominanţa acestei specii.
Fără excepţie aceşti autori stabilesc că răspândirea speciei Nardus stricta depinde în primul rând de condiţiile edafice şi apoi de cele climatice şi antropo-zoogene, factori ce se regăsesc într-o strânsă legătură de interdependenţă.
În studiile privind ecologia speciei Nardus stricta efectuate în judeţul Braşov, se încearcă o primă conturare a optimului ecologic legat de condiţiile edafice şi anume constituenţii chimici ai solului.
Dependenţa participării relative a speciei Nardus stricta faţă de valoarea pH, care se extinde de la 4,3 - 5,8 se prezintă sub forma unei curbe care atinge un maximum la valorile de 4,5 - 4,6 care delimitează domeniul foarte puternic acid de cel puternic acid (Fig. 2.3.).
Această curbă are o scădere uşoară spre domeniul acid până la 5,5 - 5,6 după care se înregistrează o scădere bruscă a participării speciei Nardus stricta spre valorile pH 5,7 - 5,8. Faţă de aciditatea solului optim ecologic al speciei Nardus stricta din zona luată în studiu, este cuprinsă în limitele pH 4,5 - 4,8 unde se înregistrează şi cel mai înalt grad de participare în covorul ierbos.
Exigenţa speciei Nardus stricta faţă de solurile acide a fost constatată pentru prima oară de COULON 1923, care stabileşte ca limită superioară de creştere şi dezvoltare pH 6,1. Mai recent LARIN 1951 specifică că în URSS nardetele se întâlnesc cel mai frecvent pe solurile cu un pH cuprins între 4,0 -5,0 CHADWICK 1960 pentru nardetele din Insulele Britanice determină un pH în apă cuprins între 4,4 - 6,0 şi KLAPP 1965 pentru Germania un pH în KCl de 3,8 - 4,5 date foarte apropiate de cele obţinute în judeţul Braşov.
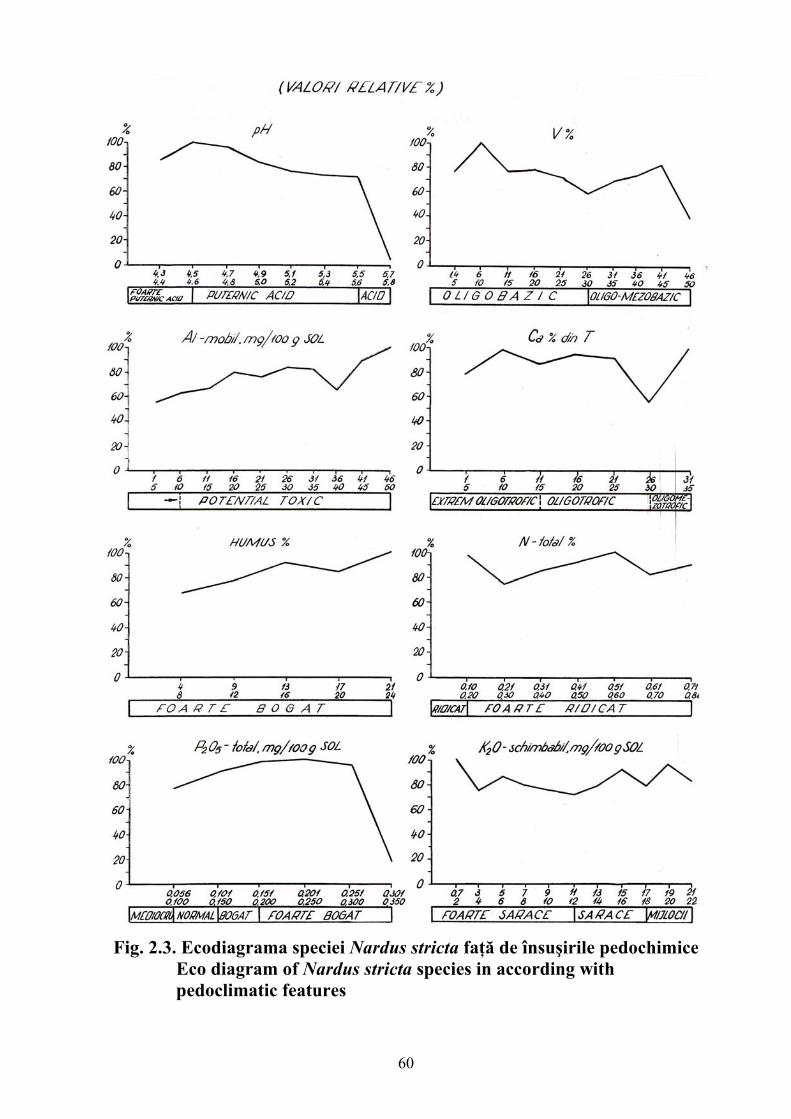
60
Fig. 2.3. Ecodiagrama speciei Nardus stricta faţă de însuşirile pedochimice
Eco diagram of Nardus stricta species in according with pedoclimatic features

61
În ceea ce priveşte dependenţa participării relative a speciei Nardus stricta faţă de gradul de saturaţie în baze (V%) putem deduce că optimum ecologic se găseşte pe solurile oligobazice la valoarea de 6 – 10%, spre cele oligo-mezobazice.
Faţă de conţinutul în Al mobil, humus % şi N total %, participarea relativă a speciei Nardus stricta urmează un curs ascendent odată cu creşterea valorilor pe ecodiagramă, conturând un optim ecologic pe solurile puternic humifere (24 %) cu Al mobil ce atinge 60 mg/100 g sol cu mult peste potenţialul considerat ca toxic.
În raport cu conţinutul solurilor în P2O5 total, se înregistrează un optim în limitele a 0,15 - 0,30 mg/100 g sol, considerat ca foarte bogat în acest element, dar inaccesibil plantelor datorită acidităţii pronunţate şi al procesului intens de debazificare al acestor soluri.
Faţă de K2O schimbabil şi Ca % din T al solului, care se găsesc în limitele foarte sărace spre mijlocii în primul caz dependenţa participării speciei Nardus stricta este indiferentă.
Această comportare a participării speciei Nardus stricta faţă de caracteristicile solului o desemnează ca plantă puternic acidofilă, care găseşte optimul de dezvoltare a fitomasei supraterane pe solurile oligobazice, puternic humifere, cu un grad ridicat de Al mobil, precum şi azot, fosfor şi alte elemente greu accesibile nutriţiei plantelor.
La aceeaşi concluzie ajung şi HUNDT 1964, MORAVEC 1965 şi KLAPP 1965 pentru nardetele din Europa Centrală.
Majoritatea solurilor pe care vegetează specia Nardus stricta au un regim aerohidric deficitar fiind slab aerate din cauza excesului temporar de umiditate şi al proceselor de gleizare care au loc. În aceste condiţii procesele de oxido-reducere sunt încetinite în materia organică nedescompusă care se acumulează în cantităţi apreciabile.
Faţă de aceste condiţii mai puţin propice pentru plante, specia Nardus stricta prezintă câteva adaptări cum sunt modul de înfrăţire, sistemul radicular şi nutriţia. Astfel mugurii de înfrăţire dispuşi intravaginal, sunt situaţi deasupra solului fiind mai puţin afectaţi de compactitatea şi slaba aerisire a acestuia. Lăstarii care pornesc din rizomi sunt foarte strâns alipiţi unul de altul, formând tufe dense în care alte plante nu pot supravieţui. Numărul lăstarilor ajunge la 78 pe o plantă (tufă) de Nardus stricta având o longevitate de 2-3 ani uneori mai mult, faţă de numai un an la specia Phleum pratense (PERKINS 1965). Rădăcinile sunt de două tipuri: unele de suprafaţă, fine şi scurte (7-14 cm) şi altele mai groase şi lungi (35-50-80 cm) cu ţesut aerifer (aerenchim) bine dezvoltat, prezentând totodată şi micoriză endotrofă. Nutriţia micotrofă oferă plantei posibilitatea să crească pe soluri mai sărace folosind substanţele nutritive mobilizate de ciupercă din humusul brut acid, acumulat adeseori în exces.
Toate aceste caracteristici morfologice şi adaptări explică într-o mare măsură robusteţea acestei specii şi posibilităţile care le are de a domina în condiţii staţionale dintre cele mai diferite, edificând fitocenoze stabile de lungă durată.
PERSIKOVA 1959 stabileşte durata medie de viaţă a unei plante de Nardus stricta la aproximativ 40 ani. Stadiul vegetativ durează 6-8 ani după care începe cel generativ ce atinge un maxim la 11-15 ani. Sfârşitul perioadei

62
generative survine la 25-30 ani, când ritmul de lăstărire se diminuează, planta devine bătrână şi eventual moare în următorii 5-7 ani.
După PUSCARU, 1956, nardetele din Bucegi ar avea o vârstă de 300-400 ani, iar SIFFERS 1952, citat de aceeaşi autor apreciază vârsta nardetelor din Caucaz la aproape 1000 de ani.
Un rol important în răspândirea nardetelor îl au factorii climatici cum sunt umiditatea şi temperatura. În ceea ce priveşte umiditatea solului, specia Nardus stricta suportă atât terenuri mai uscate formând xeronardete cât şi terenuri mai umede formând higronardete. Nu rezistă însă la secetă excesivă şi nici la inundare îndelungată (PUSCARU, 1956; BUIA 1963).
În judeţul Braşov specia Nardus stricta se întâlneşte mai abundent în zonele cu precipitaţii medii anuale mai mari de 600 mm până la 1200 mm şi temperaturi sub 80 C până la 0 grade, media anuală înregistrată în etajul alpin.
Se poate afirma că acestea sunt limite existenţei nardetelor în ţara noastră, exprimat sintetic în indicele de ariditate de MARTONNE, cu valori cuprinse între 30-120. Optimum de dezvoltare se întâlneşte în staţiunile cu indice de ariditate cuprins între 40-55, corespunzând (zonei) etajului pădurilor de fag şi molid.
Referitor la ceilalţi factori ecologici se constată o corelaţie între nivelul altitudinii şi expoziţia terenului pe care o preferă Nardus stricta (Fig. 2.4.).
Pe măsură ce altitudinea creşte nardetele se instalează cu precădere de expoziţiile însorite ale terenurilor. Peste 2000 m atitudine nardetele se întâlnesc numai pe pante însorite, fiind absente pe cele umbrite.
Pe expoziţii umbrite se întâlnesc nardetele de la altitudini joase în deosebi cele cuprinse între 800-1200 m suprafeţe mari de nardete se întâlnesc pe suprafeţele relativ plane din depresiunile Făgăraş şi Vlădeni la altitudinea de 500-700 m unde deţin aproape 2/3 din total.

63
Fig. 2.4. Exigenţele nardetelor faţă de expoziţia terenului pe altitudine
Requirements of Nardus stricta species compared with altitudinal land exposure

64
Privite în ansamblu (de la 480-2210 m alt.) nardetele din judeţul Braşov sunt repartizate destul de uniform pe toate expoziţiile terenului (Fig. 2.5.).
Fig. 2.5. Repartizarea generală a nardetelor în funcţie de expoziţia terenului General distribution of Nardus stricta areas depending on land exposure
În funcţie de înclinaţia terenului nardetele din judeţ au cea mai mare
răspândire pe locuri relativ plane (39,0 %), urmate de cele cu înclinaţie 5-100 (22,9 %); 11-200 (11,7 %); 21-300 (10,7 %); 31-400 (10,3 %) şi 41-450 ce reprezintă numai 5,4 % din cele 205 staţiuni studiate (Fig. 2.6.).
Fig. 2.6. Repartizarea nardetelor în funcţie de panta terenului, exprimată în grade
Distribution of Nardus stricta areas depending on slope of the land
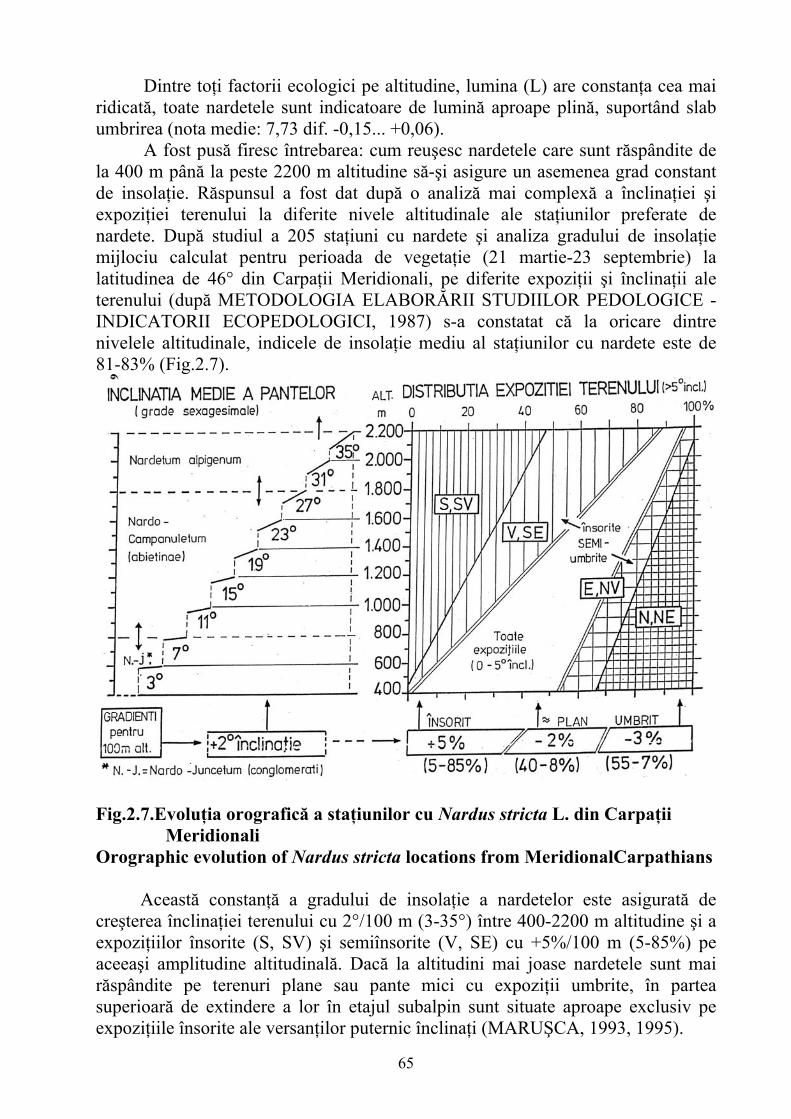
65
Dintre toţi factorii ecologici pe altitudine, lumina (L) are constanţa cea mai ridicată, toate nardetele sunt indicatoare de lumină aproape plină, suportând slab umbrirea (nota medie: 7,73 dif. -0,15... +0,06). A fost pusă firesc întrebarea: cum reuşesc nardetele care sunt răspândite de la 400 m până la peste 2200 m altitudine să-şi asigure un asemenea grad constant de insolaţie. Răspunsul a fost dat după o analiză mai complexă a înclinaţiei şi expoziţiei terenului la diferite nivele altitudinale ale staţiunilor preferate de nardete. După studiul a 205 staţiuni cu nardete şi analiza gradului de insolaţie mijlociu calculat pentru perioada de vegetaţie (21 martie-23 septembrie) la latitudinea de 46° din Carpaţii Meridionali, pe diferite expoziţii şi înclinaţii ale terenului (după METODOLOGIA ELABORĂRII STUDIILOR PEDOLOGICE - INDICATORII ECOPEDOLOGICI, 1987) s-a constatat că la oricare dintre nivelele altitudinale, indicele de insolaţie mediu al staţiunilor cu nardete este de 81-83% (Fig.2.7).
Fig.2.7.Evoluţia orografică a staţiunilor cu Nardus stricta L. din Carpaţii Meridionali Orographic evolution of Nardus stricta locations from MeridionalCarpathians
Această constanţă a gradului de insolaţie a nardetelor este asigurată de creşterea înclinaţiei terenului cu 2°/100 m (3-35°) între 400-2200 m altitudine şi a expoziţiilor însorite (S, SV) şi semiînsorite (V, SE) cu +5%/100 m (5-85%) pe aceeaşi amplitudine altitudinală. Dacă la altitudini mai joase nardetele sunt mai răspândite pe terenuri plane sau pante mici cu expoziţii umbrite, în partea superioară de extindere a lor în etajul subalpin sunt situate aproape exclusiv pe expoziţiile însorite ale versanţilor puternic înclinaţi (MARUŞCA, 1993, 1995).
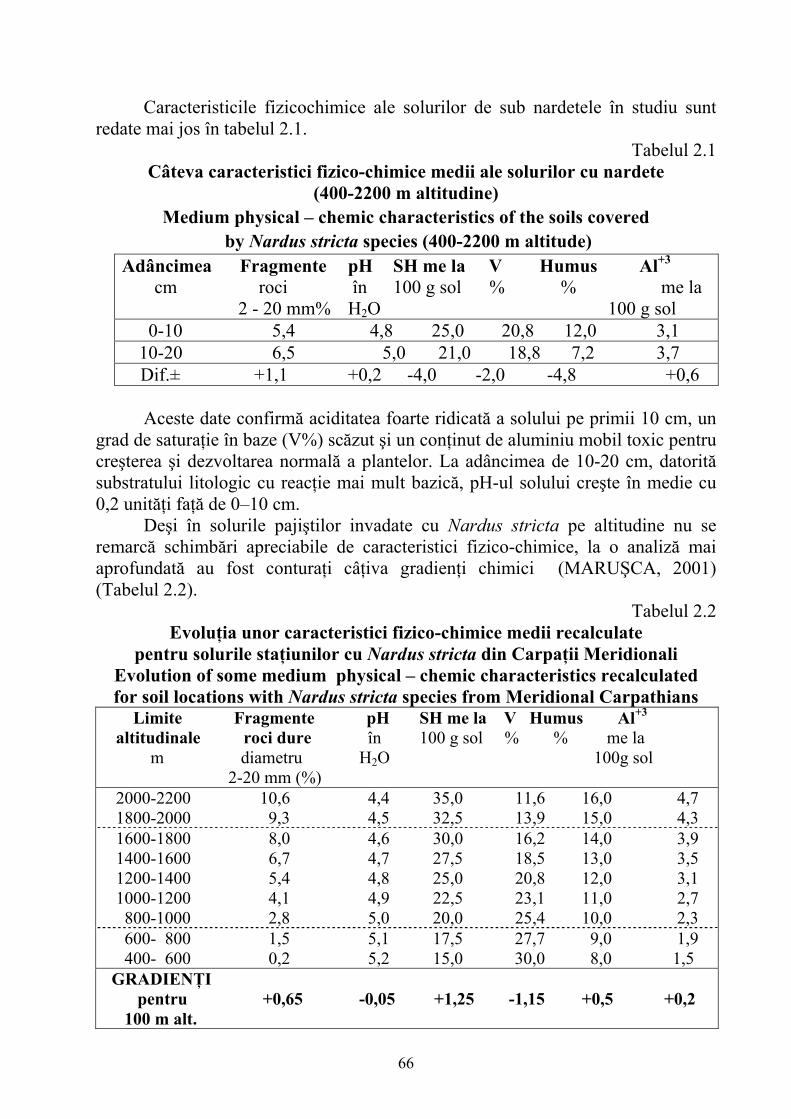
66
Caracteristicile fizicochimice ale solurilor de sub nardetele în studiu sunt redate mai jos în tabelul 2.1.
Tabelul 2.1 Câteva caracteristici fizico-chimice medii ale solurilor cu nardete
(400-2200 m altitudine) Medium physical – chemic characteristics of the soils covered
by Nardus stricta species (400-2200 m altitude) Adâncimea Fragmente pH SH me la V Humus Al+3
cm roci în 100 g sol % % me la 2 - 20 mm% H2O 100 g sol 0-10 5,4 4,8 25,0 20,8 12,0 3,1 10-20 6,5 5,0 21,0 18,8 7,2 3,7 Dif.± +1,1 +0,2 -4,0 -2,0 -4,8 +0,6
Aceste date confirmă aciditatea foarte ridicată a solului pe primii 10 cm, un grad de saturaţie în baze (V%) scăzut şi un conţinut de aluminiu mobil toxic pentru creşterea şi dezvoltarea normală a plantelor. La adâncimea de 10-20 cm, datorită substratului litologic cu reacţie mai mult bazică, pH-ul solului creşte în medie cu 0,2 unităţi faţă de 0–10 cm. Deşi în solurile pajiştilor invadate cu Nardus stricta pe altitudine nu se remarcă schimbări apreciabile de caracteristici fizico-chimice, la o analiză mai aprofundată au fost conturaţi câţiva gradienţi chimici (MARUŞCA, 2001) (Tabelul 2.2).
Tabelul 2.2 Evoluţia unor caracteristici fizico-chimice medii recalculate
pentru solurile staţiunilor cu Nardus stricta din Carpaţii Meridionali Evolution of some medium physical – chemic characteristics recalculated for soil locations with Nardus stricta species from Meridional Carpathians
Limite Fragmente pH SH me la V Humus Al+3 altitudinale roci dure în 100 g sol % % me la m diametru H2O 100g sol 2-20 mm (%) 2000-2200 10,6 4,4 35,0 11,6 16,0 4,7 1800-2000 9,3 4,5 32,5 13,9 15,0 4,3 1600-1800 8,0 4,6 30,0 16,2 14,0 3,9 1400-1600 6,7 4,7 27,5 18,5 13,0 3,5 1200-1400 5,4 4,8 25,0 20,8 12,0 3,1 1000-1200 4,1 4,9 22,5 23,1 11,0 2,7 800-1000 2,8 5,0 20,0 25,4 10,0 2,3 600- 800 1,5 5,1 17,5 27,7 9,0 1,9 400- 600 0,2 5,2 15,0 30,0 8,0 1,5 GRADIENŢI pentru +0,65 -0,05 +1,25 -1,15 +0,5 +0,2 100 m alt.
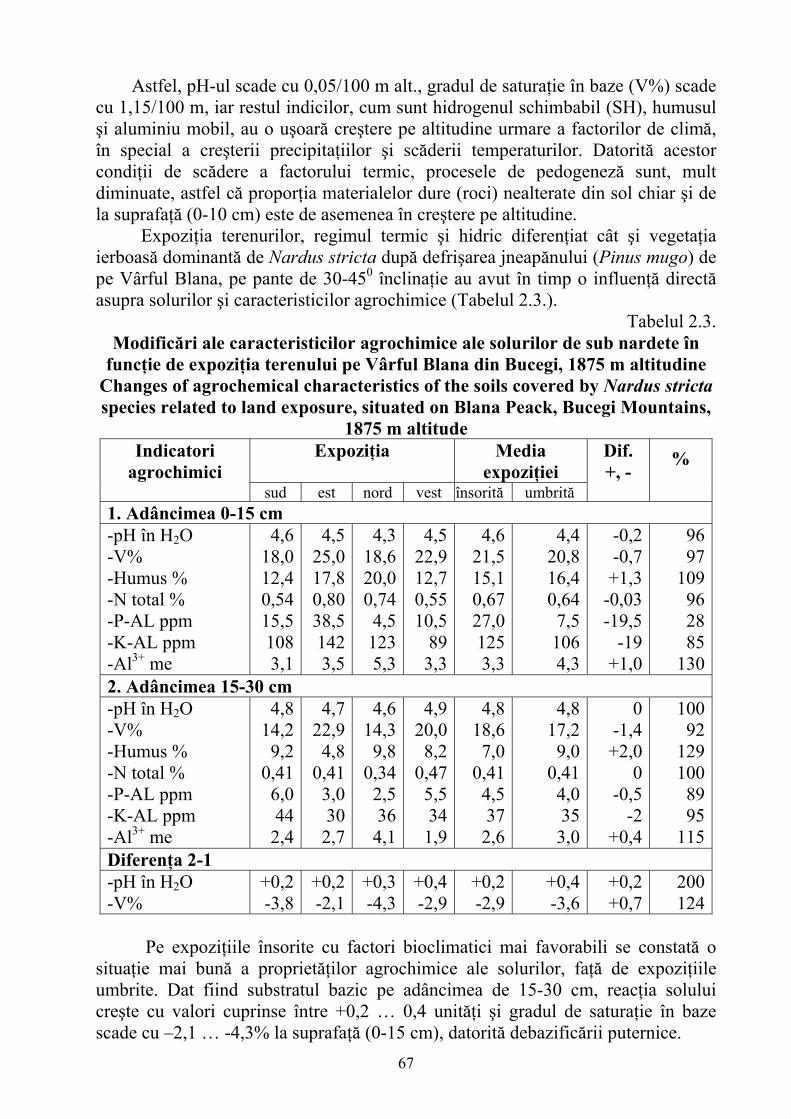
67
Astfel, pH-ul scade cu 0,05/100 m alt., gradul de saturaţie în baze (V%) scade cu 1,15/100 m, iar restul indicilor, cum sunt hidrogenul schimbabil (SH), humusul şi aluminiu mobil, au o uşoară creştere pe altitudine urmare a factorilor de climă, în special a creşterii precipitaţiilor şi scăderii temperaturilor. Datorită acestor condiţii de scădere a factorului termic, procesele de pedogeneză sunt, mult diminuate, astfel că proporţia materialelor dure (roci) nealterate din sol chiar şi de la suprafaţă (0-10 cm) este de asemenea în creştere pe altitudine. Expoziţia terenurilor, regimul termic şi hidric diferenţiat cât şi vegetaţia ierboasă dominantă de Nardus stricta după defrişarea jneapănului (Pinus mugo) de pe Vârful Blana, pe pante de 30-450 înclinaţie au avut în timp o influenţă directă asupra solurilor şi caracteristicilor agrochimice (Tabelul 2.3.).
Tabelul 2.3. Modificări ale caracteristicilor agrochimice ale solurilor de sub nardete în
funcţie de expoziţia terenului pe Vârful Blana din Bucegi, 1875 m altitudine Changes of agrochemical characteristics of the soils covered by Nardus stricta species related to land exposure, situated on Blana Peack, Bucegi Mountains,
1875 m altitude Indicatori
agrochimici Expoziţia Media
expoziţiei Dif. +, -
%
sud est nord vest însorită umbrită 1. Adâncimea 0-15 cm -pH în H2O -V% -Humus % -N total % -P-AL ppm -K-AL ppm -Al3+ me
4,6 18,0 12,4 0,54 15,5 108 3,1
4,525,017,80,8038,51423,5
4,318,620,00,744,51235,3
4,522,912,70,5510,5
893,3
4,621,515,10,6727,01253,3
4,4 20,8 16,4 0,64 7,5 106 4,3
-0,2 -0,7 +1,3
-0,03 -19,5
-19 +1,0
9697
109962885
1302. Adâncimea 15-30 cm -pH în H2O -V% -Humus % -N total % -P-AL ppm -K-AL ppm -Al3+ me
4,8 14,2 9,2
0,41 6,0 44
2,4
4,722,94,8
0,413,030
2,7
4,614,39,8
0,342,536
4,1
4,920,08,2
0,475,534
1,9
4,818,67,0
0,414,537
2,6
4,8 17,2 9,0
0,41 4,0 35
3,0
0 -1,4 +2,0
0 -0,5
-2 +0,4
10092
129100
8995
115Diferenţa 2-1 -pH în H2O -V%
+0,2 -3,8
+0,2-2,1
+0,3-4,3
+0,4-2,9
+0,2-2,9
+0,4 -3,6
+0,2 +0,7
200124
Pe expoziţiile însorite cu factori bioclimatici mai favorabili se constată o situaţie mai bună a proprietăţilor agrochimice ale solurilor, faţă de expoziţiile umbrite. Dat fiind substratul bazic pe adâncimea de 15-30 cm, reacţia solului creşte cu valori cuprinse între +0,2 … 0,4 unităţi şi gradul de saturaţie în baze scade cu –2,1 … -4,3% la suprafaţă (0-15 cm), datorită debazificării puternice.

68
Pe expoziţiile umbrite mai reci şi mai umede, este de aşteptat ca şi productivitatea pajiştilor să fie mai scăzută, asemănătoare cu a celor situate cu peste 200 m altitudine mai sus, întâlnite pe expoziţii însorite. Din acelaşi considerente pajiştile de pe pantele însorite se regăsesc cu cca 200 m mai jos pe expoziţiile umbrite ale aceluiaşi masiv montan.
Dintre factorii biotici mai ales cel antropogen are o influenţă hotărâtoare în crearea unor condiţii propice instalării şi proliferării speciei Nardus stricta. PERKINS 1965, pe baza unor documente istorice şi lucrări ştiinţifice constată că pajiştile din Britania până în secolul al XVI-lea au fost păşunate cu taurine, fiind înlocuite treptat de ovine care până la mijlocul secolului al XIX-lea au păşunat intensiv atât vara cât şi iarna. În această perioadă specia Nardus stricta a avut o răspândire redusă. Odată cu scurtarea perioadei păşunatului de iarnă cu oile şi utilizarea cu precădere numai a celui de vară la începutul secolului nostru s-a mărit cantitatea (resturile) de Nardus stricta nepăşunată, ceea ce a dus la proliferarea şi dominarea acestei specii în pajişti care altădată erau mai valoroase.
La aceiaşi concluzie ajunge LARIN 1952; BĂRBULESCU, 1978 şi alţii care consideră că păşunatul moderat mai ales cu oile înlesneşte dezvoltarea şi răspândirea nardetelor în timp ce păşunatul intens cu vitele mari – bovine şi cabaline – stânjeneşte sau chiar distrug plantele de Nardus stricta.
Suspendarea păşunatului pe suprafeţe împrejmuite în acest scop, a dus la scăderea numărului şi a dominaţiei speciei Nardus stricta. HEDDLE şi OGG 1936, menţionaţi de PERKINS 1965, au găsit că numărul lăstarilor de Nardus stricta descreşte de la 44 la 22 % după mai mulţi ani de îndepărtare a animalelor de la păşunat BUS 1937, citat de LARIN 1952, constată că pe o suprafaţă unde păşunatul a fost suspendat 5 ani, participarea speciei Nardus stricta a căzut de la 87 la 64 % iar Agrostis tenuis a crescut de la 4 la 17 %.
Precizări importante au fost aduse de KLAPP 1965, care consideră că prin păşunatul permanent şi selectiv cu ovinele, speciile valoroase sunt consumate până la distrugerea lor totală în timp ce specia Nardus stricta rămâne practic neconsumată, având posibilităţi mari de extindere. Acelaşi autor, stabileşte pentru prima dată o relaţie între distanţa unde păşunează animalele, locul de odihnă a animalelor şi gradul de răspândire a speciei Nardus stricta. Astfel, pe o păşune extensivă unde animalele se întorc zilnic acasă la fermă, o parte din dejecţii sunt îndepărtate de pe teren, având loc o scădere permanentă de substanţe nutritive, situaţie în care specia Nardus stricta se extinde în detrimentul altor specii mai exigente faţă trofismul solului. Asupra influenţei mişcării şi staţionării animalelor pe nardete sunt cunoscute mai recent şi cercetările efectuate de SPATZ 1970; MONTARD şi GACHON 1976, prin care se stabileşte gradul de participare a speciei Nardus stricta în covorul ierbos în funcţie de distanţa adăposturilor montane de vară.
În judeţul Braşov este uşor de observat influenţa care o are mişcarea şi staţionarea turmelor de oi asupra nardetelor montane şi alpine. Astfel, în imediata apropiere a adăposturilor de vară pe locurile de odihnă peste noapte intens târlite, nardetele sunt înlocuite de o vegetaţie nitrofilă specifică. La o distanţă variabilă de 100-300 m (-500 m), întâlnim o vegetaţie mai valoroasă cu un grad mai redus de

69
participare a speciei Nardus stricta unde animalele pasc mai intens şi staţionează adeseori pe timpul zilei (Fig. 2.8.).
Fig. 2.8. Repartizarea nardetelor montane în funcţie de intensitatea
păşunatului şi depărtarea de locul de odihnă al oilor Distribution of mountainous Nardus stricta areas depending on grazing
intensity and the distance from resting place of sheep Pe măsură ce ne îndepărtăm de adăpostul de vară şi împrejurimile mai
apropiate, specia Nardus stricta are o participare din ce în ce mai mare în covorul ierbos, ceea ce corespunde zonei cu păşunat moderat şi permanent cu aport minim de dejecţii animale. La marginile zonei de păşunat acolo unde turmele de oi ajung mai rar, participarea speciei Nardus stricta începe să scadă, datorită slăbirii intensităţii de păşunat sau al unui păşunat sporadic care duce la invazia ericaceelor şi a unor specii lemnoase. Privită de sus această distribuire a participării speciilor Nardus stricta ne apare sub forma unor cercuri concentrice sau segmentate în cerc în funcţie de locul amplasării adăpostului, forma suprafeţei păşunii şi condiţiile de relief.

70
Asemănător zonei de munte în depresiunile Făgăraşului, Nardus stricta are cea mai mare participare în covorul ierbos la o oarecare distanţă de localităţi, adeseori pe limită de hotar.
Aici cirezile de vite şi turmele de oi în drumul lor pasc numai la mijlocul zilei, iar dimineaţa şi seara uneori noaptea se găsesc în apropierea localităţilor unde păşunatul este mai intens şi dejecţiile au un aport mai ridicat în îmbunătăţirea păşunii.
În prezent prin aplicarea pe suprafeţe mari a îngrăşămintelor şi al construirii unor tabere de vară, aceste delimitări ale răspândirii speciei Nardus stricta în raport cu distanţa faţă de localităţi au început să se şteargă.
Desigur, aceste câteva observaţii privind răspândirea speciei Nardus stricta în funcţie de sistemele tradiţionale de păşunat din diferite ţări, merită a fi continuate şi aprofundate. Alături de tradiţie, mai intervin şi alţi factori cum sunt prezenţa animalelor sălbatice răpitoare, posibilităţile de adăpostire a animalelor domestice împotriva condiţiilor nefavorabile de mediu, îndeosebi a vânturilor. Aceste cauze de multe ori împiedică îmbunătăţirea structurală a sistemelor de păşunat practicate, care să ducă în final la reducerea şi înlăturarea speciei Nardus stricta din pajişti şi prin repartizarea mai uniformă a intensităţii de păşunat, evitarea târlirii excesive, etc.
Din cele prezentate se constată influenţa determinantă care o are omul şi animalele sale în răspândirea speciei Nardus stricta. Nardetele se prezintă astfel ca un ecosistem complex şi durabil pe suprafeţe întinse, indicând un stadiu avansat de degradare a unor pajişti mai valoroase pe care le-au înlocuit treptat.
2.3.2. Câteva date privind structura compoziţiei floristice a nardetelor Datorită condiţiilor ecologice în care se întâlnesc nardetele, numărul de
specii care le alcătuiesc este destul de redus. Fizionomia nardetelor este monotonă şi numai răspândirea lor pe un areal altitudinal atât de vast ca în judeţul Braşov face ca diversitatea florei să fie totuşi remarcată.
În cele 205 releveuri analizate pe acest teritoriu au fost determinate 202 specii aparţinând la 124 genuri şi 41 familii.
Numărul de specii întâlnite în nardetele judeţului Braşov se consideră a fi destul de ridicat, întrucât în întreaga ţară au fost determinate până acum aproximativ 290 specii.
Din totalul de 205 releveuri, 30 aparţin etajului subalpin, 72 zonei montane şi 103 în zona de deal şi depresionară a judeţului Braşov. Repartizarea speciilor pe zone şi grupe economice este prezentat în Tabelul 2.4.
Astfel în etajul subalpin au fost întâlnite 50 specii, în zona montană 88 specii şi în cea colinară - depresionară 138 specii.
Dintre acestea doar 6 specii sunt comune întregului areal de răspândire a nardetelor din zona luată în studiu şi anume: Nardus stricta, Festuca rubra, Agrostis capillaris (A.tenuis), Anthoxantum odoratum, Trifolium repens şi Bruckethalia spiculifolia.

71
Tabelul 2.4. Repartizarea pe zone geografice a speciilor de plante determinate în
nardetele jud. Braşov Distribution of plants species determinate in Nardus stricta areas
from Brasov County Grupe de
specii Repartizarea pe zone (etaje) Total
Sub alpine
Subalpmont.
Mon - tane
Mont. Colin.
Coli-nare
Subalp. Mont. Colin.
Nr. de specii
%
Gramineae 3 4 3 6 7 4 27 13,3 Cyperacee şi Juncaceae
3 1 2 4 12 - 22 10,9
Leguminosae - 1 - 5 8 1 15 7,4 Alte familii 19 11 14 28 51 1 124 61,4
Sp. lemnoase 2 - 1 2 9 - 14 7,0 Total nr. specii 27 17 20 45 87 6 202 100,0% participare 13,3 8,4 9,9 22,3 43,1 3,0 100,0 X
Alte detalii asupra compoziţiei floristice a nardetelor pe cele trei zone mari
(subalpine – montane – colinare) vor fi prezentate în descrierea asociaţiilor edificate de Nardus stricta care sunt conturate în linii generale în funcţie de aceste zone altitudinal geografice cu speciile lor caracteristice.
Spectrul bioformelor este dominat de hemicriptofite. Sub raport numeric din cele 202 specii, 72,1 % sunt hemicriptofite, care sunt prezente într-o proporţie medie de 86,8 % în cele 205 releveuri analizate.Proporţia foarte ridicată a hemicriptofitelor în nardetele luate în studiu ne indică un grad înalt de stabilitate a acestor fitocenoze.
Spectrul fitogeografic atât sub raport numeric cât şi al prezenţei este dominat de elemente eurasiatice (35,6%) urmat de circumpolare (15,3%), europene (15,3%), alpine (13,8%), central europene (6,5%), cosmopolite (5,0%), dacobalcanice (3%), continentale (3%) şi alte elemente cu participare mai redusă din care cele mai importante sunt cele endemice cu 0,5%.

72
2. 4. DESCRIEREA ASOCIAŢIILOR DE Nardus stricta L. Nardetele judeţului Braşov a căror răspândire prezentată pe hartă (Fig. 2.9.)
sunt alcătuite din trei asociaţii şi şase subasociaţii care au fost încadrate în două alianţe, un ordin şi o clasă după cum urmează:
CLASA NARDO - CALLUNETEA, Preisg 49. Ordinul Nardetalia, Preisg 49. Alianţa Potentillo (ternatae) - Nardion, Simon 57. 1. As. Nardetum alpigenum (Br.Bl. 49), Borza 59. 2. As. Nardo - Campanuletum (abietinae), Boşcaiu 71. nom.corr.
- nardetosum, Buia 63. - festucetosum rubrae, Buia 63.
Alianţa Violo (caninae) - Nardion, Oberd 49. 3. As. Nardo - Juncetum (conglomerati), Resmeriţă 69. nom.corr.as.nov.?
- agrostidetosum tenui, Buia 63., Turcu 68. - agrostidetosum canini Turcu 68. - molinietosum Gergely 56., nom.nud - eriphoretosum subas.nov.
Principalele specii caracteristice ale clasei Nardo - Callunetea sunt: Potentilla erecta, Sieglingia decumbens, Thymus pulegioides, Luzula campestris, Veronica officinalis, Deschampsia flexuosa, Luzula multiflora, Hieracium pilosella şi Antenaria dioica.
Ecodiagrama speciei Sieglingia decumbens una din cele mai reprezentative din această clasă indică soluri puternic acide cu un pH în apă cuprins între 4,5 – 5,5, oligobazice, cu un conţinut în Al mobil situat la limita inferioară a potenţialului toxic, fiind puţin influenţată de conţinutul în cationi de Ca din sol şi K2O schimbabil (Fig. 2.10.)
Caracteristicile ordinului Nardetalia sunt: Nardus stricta, Carex pallescens, Carex leporina, Hypericum maculatum,Coeloglossum viride, Alchemilla hybrida şi Euphrasia stricta.
Ecodiagrama speciei Carex pallescens una din cele mai răspândite plante din nardetele colinare şi montane indică soluri puternic acide cu un pH cuprins între 4,5 – 5,6, cu un optim la pH 5,5, soluri oligobazice spre oligomezobazice, oligomezotrofice cu un conţinut scăzut în Al mobil (Fig. 2.11).
Faţă de conţinutul în humus al solului se remarcă o preferinţă spre valori mai scăzute, nedepăşind 16%. Celelalte elemente din sol nu influenţează evident participarea şi prezenţa acestei specii în covorul ierbos.
Aceste date privind optimum ecologic al speciilor caracteristice clasei Nardo - Callunetea şi ordinului Nardetalia confirmă rezultatele şi concluziile la care au ajuns HUNDT 1964 şi KLAPP 1965, asupra valorii indicatoare pe care le au Nardus stricta, Sieglingia decumbens, Carex pallescens şi altele, pentru unii constituenţi ai solurilor unde întâlnim nardete.
În ceea ce priveşte speciile caracteristice ale alianţei Potentillo - Nardion, le amintim pe următoarele: Potentilla ternata, Viola declinata, Campanula serrata, Euphrasia tatrae, Cerastium fontanum şi Carlina acaulis.
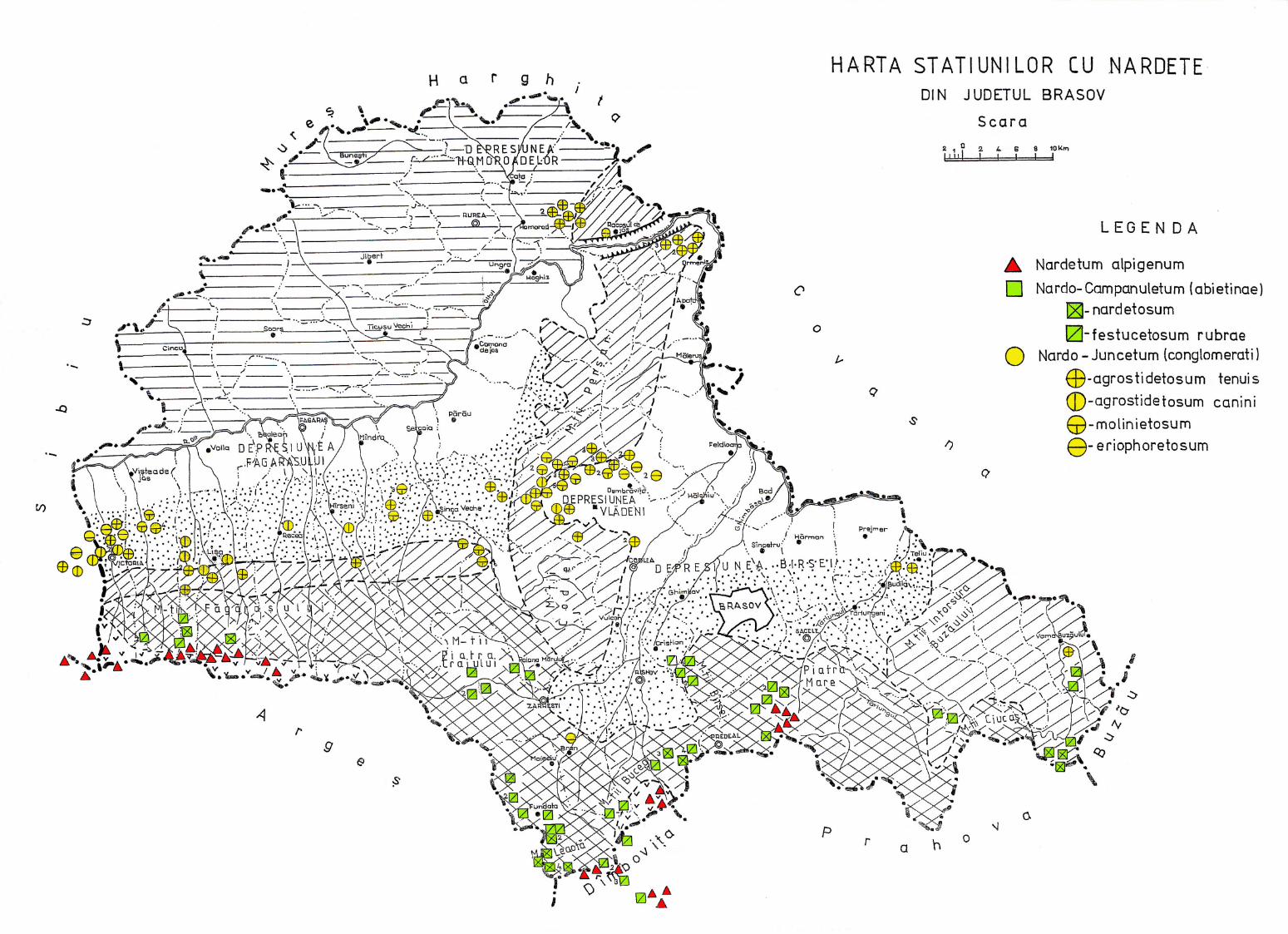

73
Pajişti subalpine din Platoul Bucegilor invadate de Nardus stricta
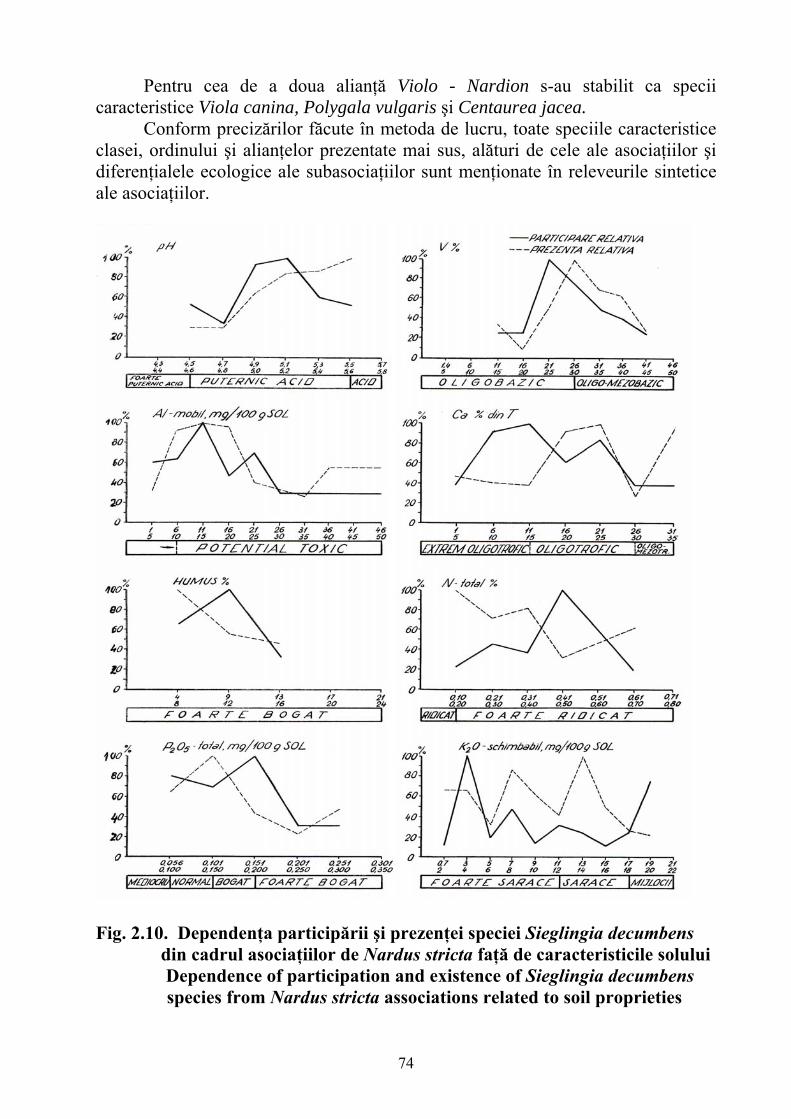
74
Pentru cea de a doua alianţă Violo - Nardion s-au stabilit ca specii caracteristice Viola canina, Polygala vulgaris şi Centaurea jacea.
Conform precizărilor făcute în metoda de lucru, toate speciile caracteristice clasei, ordinului şi alianţelor prezentate mai sus, alături de cele ale asociaţiilor şi diferenţialele ecologice ale subasociaţiilor sunt menţionate în releveurile sintetice ale asociaţiilor.
Fig. 2.10. Dependenţa participării şi prezenţei speciei Sieglingia decumbens din cadrul asociaţiilor de Nardus stricta faţă de caracteristicile solului
Dependence of participation and existence of Sieglingia decumbens species from Nardus stricta associations related to soil proprieties
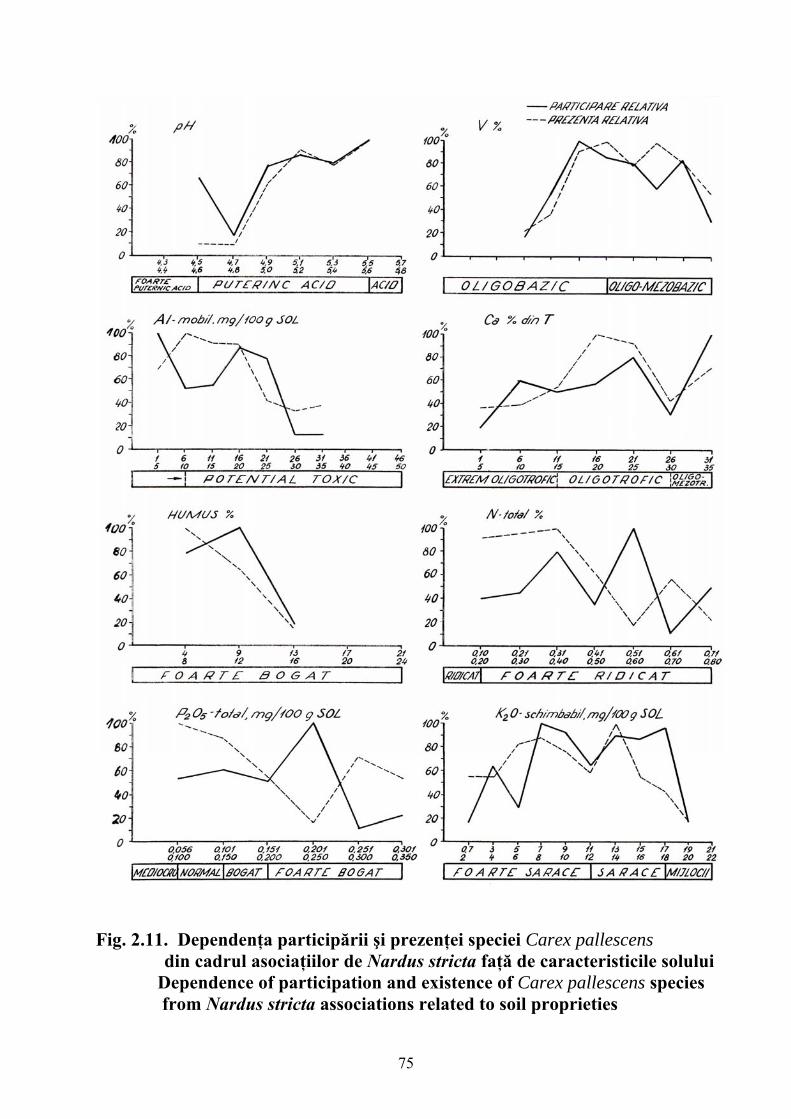
75
Fig. 2.11. Dependenţa participării şi prezenţei speciei Carex pallescens din cadrul asociaţiilor de Nardus stricta faţă de caracteristicile solului Dependence of participation and existence of Carex pallescens species from Nardus stricta associations related to soil proprieties

76
2.4.1. As. Nardetum alpigenum Br. Bl. 49, Borza 59. (Syn.Nardetum, Domin. 33; Nardetum strictae Soo 28; Nardetum alpigenum
austro - carpaticum, Borza 59; Nardetum strictae alpinum, Borza 34 p.p.; Ghişa 40 p.p.; Puşcaru et al. 56 p.p.; Buia 63 p.p.; Beldie 67 p.p., As Nardus stricta alpină, Puşcaru et al. 1963).
Această asociaţie vegetală aşa cum indică numele se întâlneşte în etajul subalpin al rariştilor de molid până la jnepenişurile din M-ţii Făgăraş, Masivul Bucegi şi M-tele Piatra Mare, la o altitudine de 1780 - 2210 m, cu caracteristici staţionale şi productive prezentate în tabelul 2.5. şi 2.6.
Tabelul 2.5. As. Nardetum alpigenum (Br.Bl. 49) Borza 59
Association of Nardetum alpigenum
Caracteristici staţionale şi productive Nardetum alpigenum
Număr staţiuni 30 Altitudine (m) 1945 (1780 - 2210) Înclinaţie (garde) 250 (0 - 450) Substrat geologic conglom., şisturi cristaline Temperatura medie anuală ( 0 C) 0 - 20 Precipitaţii medii anuale (mm) peste 1200 Etajul de vegetaţie primară jneapăn, rarişte molid Tipuri de soluri Al, P, AB pH 4,7 (4,5 - 4.9) V % 14,9 (12,5 - 18.2) Humus % 14,5 (8,7 - 20.2) P2O5 total mg / 100 g sol 0,20 (0,16 - 0,27) K2O schimbabil mg / 100 g sol 9,7 (7,0 - 10.9)
Fragmente rocă Ø 2-20 mm (%) 7,6 (2,3 - 14.4)
Adâncime fragmente rocă cu Ø>20 mm (m) 0,0 -0,4 Modul de folosire actual al pajiştii păşune ovine Acoperirea solului cu vegetaţie (%) 80 - 90 Producţia de iarbă (t/ha) 2,1 (1,5 - 2,5) Valoarea pastorală 14 (9 - 27) Bio for ma
Element floristic
Aparte nenţa
cenotică
SPECII
AD
P %
K
Total % Cl
Gramineae (87) - - - H. Eua Car. ord. Nardus stricta 3 - 5 58 30 100 V H. Alp Rec. as Festuca airoides
(ovina sudetica) 1 - 2 8 27 90 V
H. Alp Rec. as Agrostis rupestris 1 - 2 10 25 83 V H. Eua Anthoxantum odoratum + - 1 1 22 73 IV H. Cp Poa alpina + - 2 5 20 67 IV H. Cp Festuca rubra 1 - 2 2 13 43 III H. Alp Phleum alpinum + - 1 + 8 27 II
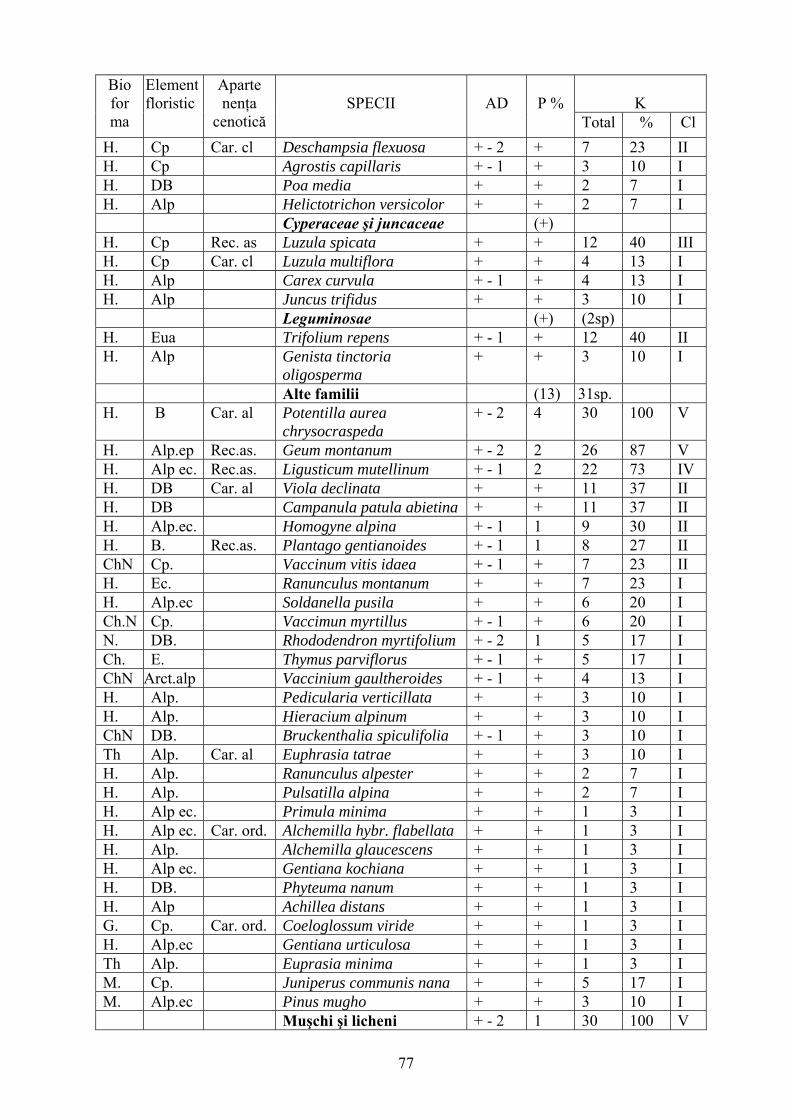
77
Bio for ma
Element floristic
Aparte nenţa
cenotică
SPECII
AD
P %
K
Total % Cl
H. Cp Car. cl Deschampsia flexuosa + - 2 + 7 23 II H. Cp Agrostis capillaris + - 1 + 3 10 I H. DB Poa media + + 2 7 I H. Alp Helictotrichon versicolor + + 2 7 I Cyperaceae şi juncaceae (+) H. Cp Rec. as Luzula spicata + + 12 40 III H. Cp Car. cl Luzula multiflora + + 4 13 I H. Alp Carex curvula + - 1 + 4 13 I H. Alp Juncus trifidus + + 3 10 I Leguminosae (+) (2sp) H. Eua Trifolium repens + - 1 + 12 40 II H. Alp Genista tinctoria
oligosperma + + 3 10 I
Alte familii (13) 31sp. H. B Car. al Potentilla aurea
chrysocraspeda + - 2 4 30 100 V
H. Alp.ep Rec.as. Geum montanum + - 2 2 26 87 V H. Alp ec. Rec.as. Ligusticum mutellinum + - 1 2 22 73 IV H. DB Car. al Viola declinata + + 11 37 II H. DB Campanula patula abietina + + 11 37 II H. Alp.ec. Homogyne alpina + - 1 1 9 30 II H. B. Rec.as. Plantago gentianoides + - 1 1 8 27 II ChN Cp. Vaccinum vitis idaea + - 1 + 7 23 II H. Ec. Ranunculus montanum + + 7 23 I H. Alp.ec Soldanella pusila + + 6 20 I Ch.N Cp. Vaccimun myrtillus + - 1 + 6 20 I N. DB. Rhododendron myrtifolium + - 2 1 5 17 I Ch. E. Thymus parviflorus + - 1 + 5 17 I ChN Arct.alp Vaccinium gaultheroides + - 1 + 4 13 I H. Alp. Pedicularia verticillata + + 3 10 I H. Alp. Hieracium alpinum + + 3 10 I ChN DB. Bruckenthalia spiculifolia + - 1 + 3 10 I Th Alp. Car. al Euphrasia tatrae + + 3 10 I H. Alp. Ranunculus alpester + + 2 7 I H. Alp. Pulsatilla alpina + + 2 7 I H. Alp ec. Primula minima + + 1 3 I H. Alp ec. Car. ord. Alchemilla hybr. flabellata + + 1 3 I H. Alp. Alchemilla glaucescens + + 1 3 I H. Alp ec. Gentiana kochiana + + 1 3 I H. DB. Phyteuma nanum + + 1 3 I H. Alp Achillea distans + + 1 3 I G. Cp. Car. ord. Coeloglossum viride + + 1 3 I H. Alp.ec Gentiana urticulosa + + 1 3 I Th Alp. Euprasia minima + + 1 3 I M. Cp. Juniperus communis nana + + 5 17 I M. Alp.ec Pinus mugho + + 3 10 I Muşchi şi licheni + - 2 1 30 100 V
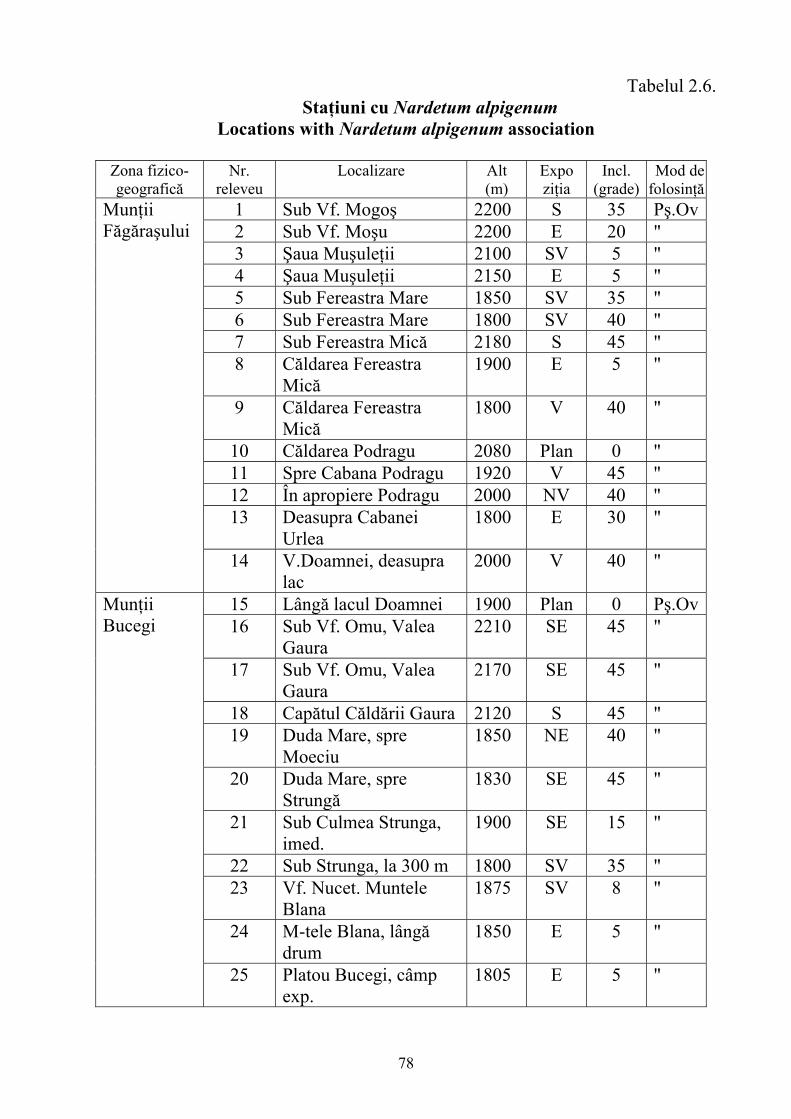
78
Tabelul 2.6. Staţiuni cu Nardetum alpigenum
Locations with Nardetum alpigenum association
Zona fizico-geografică
Nr. releveu
Localizare Alt (m)
Expo ziţia
Incl. (grade)
Mod defolosinţă
Munţii Făgăraşului
1 Sub Vf. Mogoş 2200 S 35 Pş.Ov2 Sub Vf. Moşu 2200 E 20 " 3 Şaua Muşuleţii 2100 SV 5 " 4 Şaua Muşuleţii 2150 E 5 " 5 Sub Fereastra Mare 1850 SV 35 " 6 Sub Fereastra Mare 1800 SV 40 " 7 Sub Fereastra Mică 2180 S 45 " 8 Căldarea Fereastra
Mică 1900 E 5 "
9 Căldarea Fereastra Mică
1800 V 40 "
10 Căldarea Podragu 2080 Plan 0 " 11 Spre Cabana Podragu 1920 V 45 " 12 În apropiere Podragu 2000 NV 40 " 13 Deasupra Cabanei
Urlea 1800 E 30 "
14 V.Doamnei, deasupra lac
2000 V 40 "
Munţii Bucegi
15 Lângă lacul Doamnei 1900 Plan 0 Pş.Ov16 Sub Vf. Omu, Valea
Gaura 2210 SE 45 "
17 Sub Vf. Omu, Valea Gaura
2170 SE 45 "
18 Capătul Căldării Gaura 2120 S 45 " 19 Duda Mare, spre
Moeciu 1850 NE 40 "
20 Duda Mare, spre Strungă
1830 SE 45 "
21 Sub Culmea Strunga, imed.
1900 SE 15 "
22 Sub Strunga, la 300 m 1800 SV 35 " 23 Vf. Nucet. Muntele
Blana 1875 SV 8 "
24 M-tele Blana, lângă drum
1850 E 5 "
25 Platou Bucegi, câmp exp.
1805 E 5 "

79
Zona fizico-geografică
Nr. releveu
Localizare Alt (m)
Expo ziţia
Incl. (grade)
Mod de folosinţă
Munţii Bârsei (Piatra Mare)
26 Lângă Vf. Piatra Mare 1840 Plan 0 " 27 Sub Vf. Piatra Mare 1830 S 30 " 28 Platou Piatra Mare 1800 E 5 " 29 Baza Platou Piatra
Mare 1780 V 25 "
30 Baza Platou spre Tamina
1780 NV 20 "
Asociaţia cantonează îndeosebi pe expoziţii însorite (SV; S; SE) cu pante
preponderent repezi 25-450 (Fig. 2.12.).
Fig.2.12. Răspândirea asociaţiei Nardetum alpigenum în funcţie de expoziţie(a)
şi panta terenului (b) Distribution of Nardetum alpigenum association related to
exposure (a) and slope of the land (b)
Temperatura medie anuală foarte scăzută 0-2 0C, precipitaţiile abundente 1200 mm pe an şi mai ales regimul eolian foarte activ, caracteristic bioclimatului subalpin, au o influenţă determinantă asupra vegetaţiei primare şi a solurilor.
Vegetaţia primară a fost de jnepenişuri care prin defrişare şi păşunat a fost succedată de o vegetaţie de rodorete şi vaccinete de Agrostis rupestris, Festuca airoides şi Nardus stricta care ajunge în cele din urmă să edifice asociaţii larg răspândite. În partea inferioară a nardetelor subalpine actuale, vegetaţia primară a fost constituită din rarişti de molid iar în cea superioară nardetele ajung până la limita de extindere a vegetaţiei lemnoase, respectiv a jneapănului, uneori depăşind-o invadând pe alocuri vegetaţia ierboasă primară caracteristică etajului alpin.

80
S-a pus problema existenţei unor fitocenoze primare edificate de Nardus stricta în etajul alpin. Acest fapt nu s-a semnalat încă, toţi autorii care s-au ocupat de geneza nardetelor sunt de acord că aceste fitocenoze sunt de origine secundară.
Principalele tipuri de soluri pe care s-au dezvoltat nardetele subalpine sunt: humicosilicatice (Al), podzoluri humicoferiiluviale (Sp) şi brune acide (BO, AB).
Aceste soluri prezintă cea mai pronunţată aciditate şi cel mai scăzut conţinut de baze schimbabile faţă de alte tipuri de sol pe care se întâlnesc nardetele în judeţul Braşov.
Analiza ecologică a unor specii mai răspândite în nardetele subalpine, aduc câteva lămuriri suplimentare asupra condiţiilor de sol.
Ecodiagrama speciei Agrostis rupestris ne indică o dependenţă faţă de aciditatea pronunţată a solului (pH 4,3-5,2) cu gradul de saturaţie în baze de până la 30 %, atinge optimul ecologic la un conţinut de cca. 20 % humus şi preferă solurile bogate în N şi P2O5 total şi aprovizionate mijlociu în K2O schimbabil.
Prezenţa speciei Agrostis rupestris în nardetele subalpine ne mai indică un conţinut foarte scăzut în calciu a solurilor respective care sunt considerate din acest punct de vedere ca extrem oligotrofice, situaţie în care şi celelalte elemente de nutriţie sunt mai greu accesibile plantelor. În aceste condiţii staţionale se descrie o singură asociaţie Nardetum alpigenum având ca specii de recunoaştere: Festuca ovina sudetica, Agrostis rupestris, Luzula spicata, Geum montanum,Ligusticum mutellium, Plantago gentianoides şi alte specii caracteristice etajului subalpin.
În cele 30 releveuri descrise, asociaţia cuprinde 50 specii din care 11 Graminee, 4 Cyperaceae şi Juncaceae, 2 Leguminoase, 31 alte familii şi 2 specii lemnoase.
Bioformele cele mai răspândite sunt hemicriptofitele care ating 78 % ca raport numeric, 98,6 % participare în biomasă şi 90,4 % prezenţă (Fig. 2.13.).
Fig. 2.13. Spectrul bioformelor asociaţiei Nardetum alpigenum
Bioforma spectrum of Nardetum alpigenum association

81
Această distribuţie a bioformelor conferă asociaţiei un grad înalt de stabilitate în timp, consemnat şi în paragraful precedent. Spectrul fitogeografic sub raport numeric şi al prezenţei este dominat de elemente alpine - subalpine şi circumpolare. În schimb participarea în biomasă este dominată de elemente eurasiatice urmată de cele alpin - subalpin, circumpolare şi altele. Analizând distribuţia spectrelor fitogeografice, constatăm predominarea elementelor alpine şi circumpolare ce conturează mai clar asociaţia Nardetum alpigenum faţă de celelalte asociaţii edificate de specia Nardus stricta. Sub raport economic asociaţia Nardetum alpigenum are o însemnătate mai redusă. Acest fapt se datoreşte producţiei de fitomasă de 2,1 t/ha şi indicele de valoare pastorală de numai 14 în medie, respectiv foarte săracă. La această apreciere trebuie să se aibă în vedere şi condiţiile climatice deosebit de aspre pentru om şi animalele care valorifică producţia, accesibilitatea acestor teritorii, posibilităţile reduse de îmbunătăţire a acestor asociaţii, aspecte care se vor prezenta pe larg în descrierea tipurilor de nardete. Valorificarea actualî a producţiei de masă verde se face în general prin păşunat cu oi, tineret ovin şi sporadic cu tineret taurin, vaci şi cai în locuri mai accesibile din Masivul Bucegi.
2.4.2. As. Nardo - Campanuletum (abietinae) Boşcaiu 71. nom.corr. (Syn. Nardetum strictae montanum Puşcaru et al.56; Buia et al. 62; As de Nardus stricta, montană, Puşcaru şi col. 63; As Nardus stricta + Thymus dacicus Anghel şi colab. 65). Reprezintă una din asociaţiile de bază a vegetaţiei pajiştilor din judeţul Braşov, care s-a identificat în 72 staţiuni. Cuprinsă între 850-1710 m altitudine, asociaţia Nardo - Campanuletum beneficiază de condiţii bioclimatice mai favorabile faţă de nardetele subalpine (Tabelele 2.7. şi 2.8.). Asociaţia se întâlneşte pe expoziţii diferite, mai ales însorite dar şi pe cele semiînsorite, semiumbrite şi chiar umbrite la altitudini mai joase. Înclinaţia pantelor este mai domoală, predominând cu valori de 200. Temperatura medie anuală de până la 60C şi precipitaţii de 900-1200 mm, sunt evident mai propice vieţuirii plantelor faţă de condiţii bioclimatice ale etajului subalpin. Vegetaţia primară este bine reprezentată de întinse păduri de molid în partea superioară, fag în partea inferioară şi de amestec (molid + fag; fag - molid), la confluenţa acestora. De asemenea, condiţiile edafice sunt mai bune, predominând tipurile de sol ca: brune acide (BO), brune podzolice (BP), brune podzolice feriiluviale (Bfe), podzolice feriiluviale (P), rendzine (R) şi altele.
După defrişarea pădurilor de molid şi fag prin succesiunea unor stadii ale vegetaţiei ierboase de tăietură în prezenţa păşunatului se instalează fitocenoze edificate în principal de speciile Festuca rubra şi Agrostis tenuis.

82
Tabelul 2.7. As. Nardo - Campanuletum (abietinae) Boşcaiu 71.
Association of Nardo - Campanuletum (abietinae) species
Caracteristici staţionale şi productive
Nardo - Campanuletum
1. nardetosum
2. festucetosum rubrae
Număr staţiuni 72 Altitudine (m) 1300 (850-1750) 1570 (850-1750) 1190(850-1750) Înclinaţie (grade) 21 (2 - 40) 25 (4 - 40) 20 (4 - 40) Temperatura medie anuală ( 0 C) 2 - 6 0 2 - 6 0 2 - 6 0 Precipitaţii medii anuale (mm) 900 - 1200 900 - 1200 900 - 1200 Etajul de vegetaţie primară Molid, molid+fag, fag Molid, Molid, molid+fag,
fag Tipuri de soluri BO,Bfe,P,BM,R Bfe, P BO,BP,Bfe,BM,R pH 4,9 (4,3 - 5,3) 4,3 (4,3 - 4,8) 5,0 (4,6 - 5,3) V % 20,3 (5,7 - 34,4) 14,3 (5,7 - 21,8) 22,2 (11,3-34,4) Humus % 11,8 (4,6 - 19,8) 14,3 (9,6 - 19,8) 10,9 (4,6-13,8) P2O5 total mg / 100 g sol 0,18 (0,13- 0,25) 0,20 (0,18 - 0,22) 0,17(0,13-0,25) K2O schimbabil mg / 100 g sol 13,8 (7,8 - 23,0) 16,7 (8,2 - 22,2) 12,7 (7,8 - 23,0)
Fragmente rocă Ø 2-20 mm (%) 5,4 (0,0 -27,9) 8,1 (0,8 - 27,9) 4,1 (0,0 - 14,3)
Adâncime fragmente rocă cu Ø>20 mm (m) 0,0 - 1,2 0,0 - 0,5 0,0 - 1,2 Modul de folosire actual al pajiştii Ovine, bovine, mixt, fâneaţă Ovine, bovine Ovine, bovine, mixt Acoperirea solului cu vegetaţie (%) 85 - 100 85 - 100 85 - 100 Producţia de iarbă (t/ha) 3,4 (2,5 - 4,5) 3,2 (2,5 - 3,5) 3,6 (3,0 - 4,5) Valoarea pastorală 20 (2 - 46) 8 (2 - 15) 27 (17 - 46)
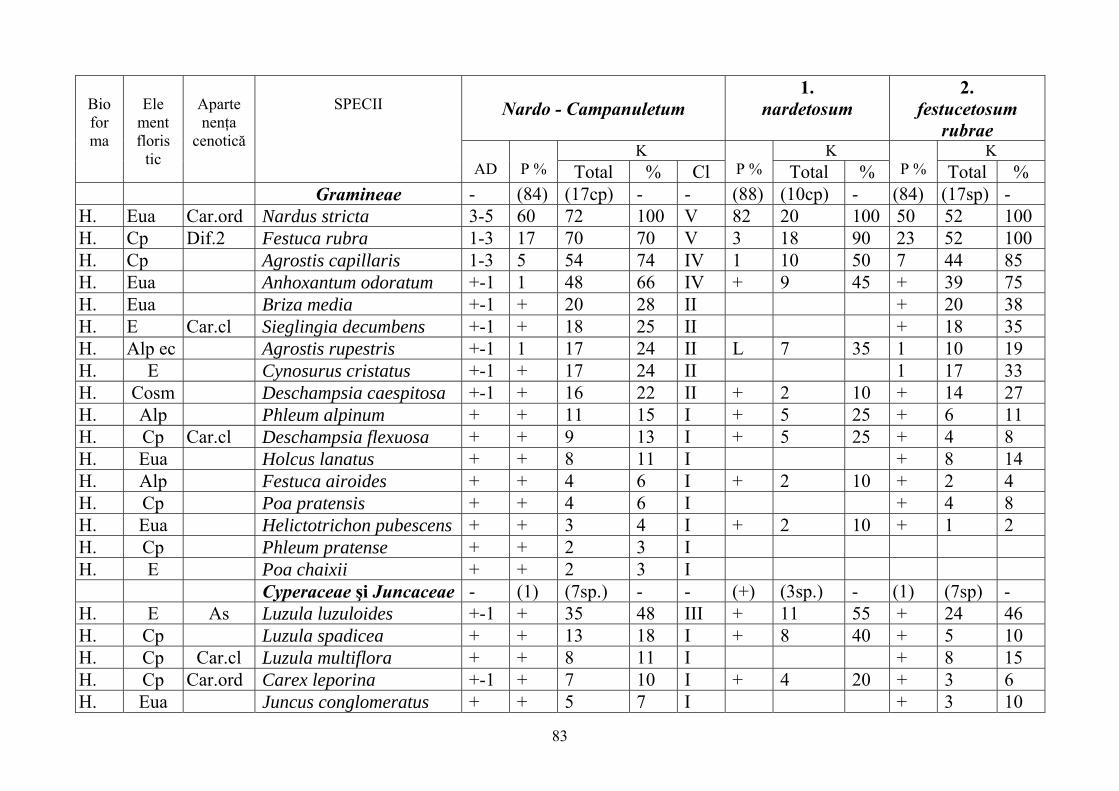
83
Bio for ma
Ele
ment floris
tic
Aparte nenţa
cenotică
SPECII
Nardo - Campanuletum
1. nardetosum
2. festucetosum
rubrae
AD
P % K
P % K
P % K
Total % Cl Total % Total % Gramineae - (84) (17cp) - - (88) (10cp) - (84) (17sp) - H. Eua Car.ord Nardus stricta 3-5 60 72 100 V 82 20 100 50 52 100 H. Cp Dif.2 Festuca rubra 1-3 17 70 70 V 3 18 90 23 52 100 H. Cp Agrostis capillaris 1-3 5 54 74 IV 1 10 50 7 44 85 H. Eua Anhoxantum odoratum +-1 1 48 66 IV + 9 45 + 39 75 H. Eua Briza media +-1 + 20 28 II + 20 38 H. E Car.cl Sieglingia decumbens +-1 + 18 25 II + 18 35 H. Alp ec Agrostis rupestris +-1 1 17 24 II L 7 35 1 10 19 H. E Cynosurus cristatus +-1 + 17 24 II 1 17 33 H. Cosm Deschampsia caespitosa +-1 + 16 22 II + 2 10 + 14 27 H. Alp Phleum alpinum + + 11 15 I + 5 25 + 6 11 H. Cp Car.cl Deschampsia flexuosa + + 9 13 I + 5 25 + 4 8 H. Eua Holcus lanatus + + 8 11 I + 8 14 H. Alp Festuca airoides + + 4 6 I + 2 10 + 2 4 H. Cp Poa pratensis + + 4 6 I + 4 8 H. Eua Helictotrichon pubescens + + 3 4 I + 2 10 + 1 2 H. Cp Phleum pratense + + 2 3 I H. E Poa chaixii + + 2 3 I Cyperaceae şi Juncaceae - (1) (7sp.) - - (+) (3sp.) - (1) (7sp) - H. E As Luzula luzuloides +-1 + 35 48 III + 11 55 + 24 46 H. Cp Luzula spadicea + + 13 18 I + 8 40 + 5 10 H. Cp Car.cl Luzula multiflora + + 8 11 I + 8 15 H. Cp Car.ord Carex leporina +-1 + 7 10 I + 4 20 + 3 6 H. Eua Juncus conglomeratus + + 5 7 I + 3 10
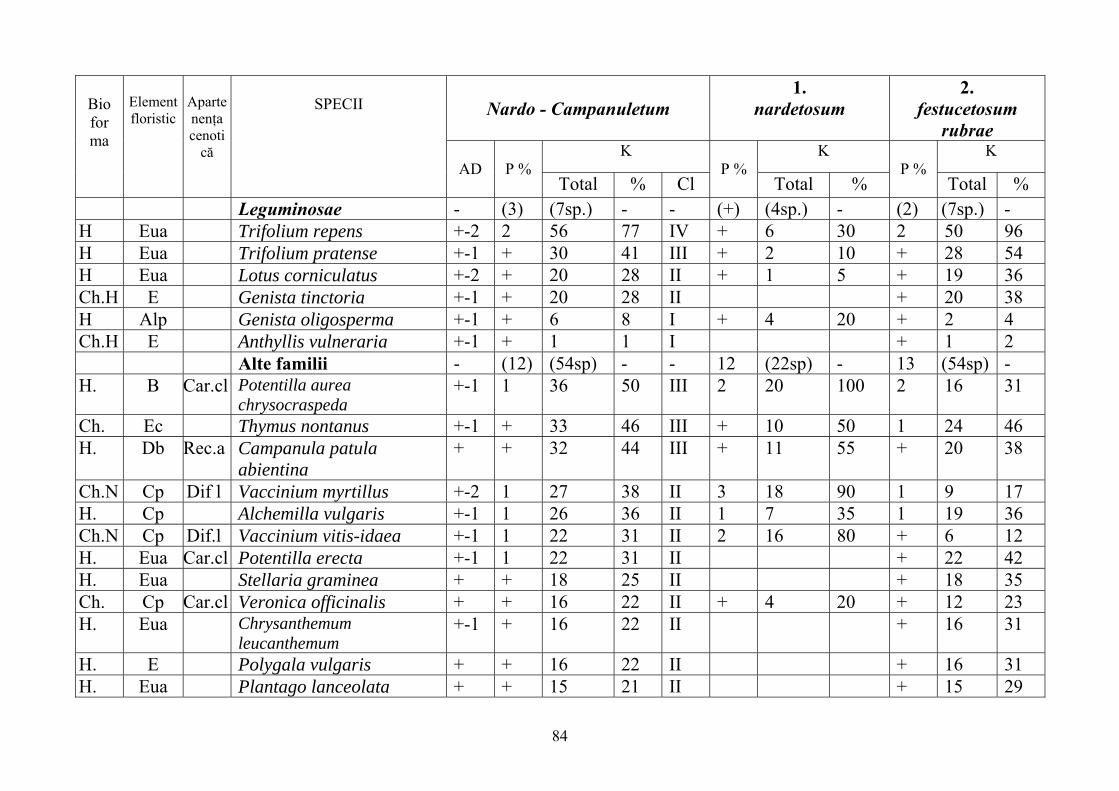
84
Bio for ma
Element floristic
Apartenenţa cenoti
că
SPECII
Nardo - Campanuletum
1. nardetosum
2. festucetosum
rubrae
AD
P % K
P % K
P % K
Total % Cl Total % Total % Leguminosae - (3) (7sp.) - - (+) (4sp.) - (2) (7sp.) - H Eua Trifolium repens +-2 2 56 77 IV + 6 30 2 50 96 H Eua Trifolium pratense +-1 + 30 41 III + 2 10 + 28 54 H Eua Lotus corniculatus +-2 + 20 28 II + 1 5 + 19 36 Ch.H E Genista tinctoria +-1 + 20 28 II + 20 38 H Alp Genista oligosperma +-1 + 6 8 I + 4 20 + 2 4 Ch.H E Anthyllis vulneraria +-1 + 1 1 I + 1 2 Alte familii - (12) (54sp) - - 12 (22sp) - 13 (54sp) - H. B Car.cl Potentilla aurea
chrysocraspeda +-1 1 36 50 III 2 20 100 2 16 31
Ch. Ec Thymus nontanus +-1 + 33 46 III + 10 50 1 24 46 H. Db Rec.a Campanula patula
abientina + + 32 44 III + 11 55 + 20 38
Ch.N Cp Dif l Vaccinium myrtillus +-2 1 27 38 II 3 18 90 1 9 17 H. Cp Alchemilla vulgaris +-1 1 26 36 II 1 7 35 1 19 36 Ch.N Cp Dif.l Vaccinium vitis-idaea +-1 1 22 31 II 2 16 80 + 6 12 H. Eua Car.cl Potentilla erecta +-1 1 22 31 II + 22 42 H. Eua Stellaria graminea + + 18 25 II + 18 35 Ch. Cp Car.cl Veronica officinalis + + 16 22 II + 4 20 + 12 23 H. Eua Chrysanthemum
leucanthemum +-1 + 16 22 II + 16 31
H. E Polygala vulgaris + + 16 22 II + 16 31 H. Eua Plantago lanceolata + + 15 21 II + 15 29
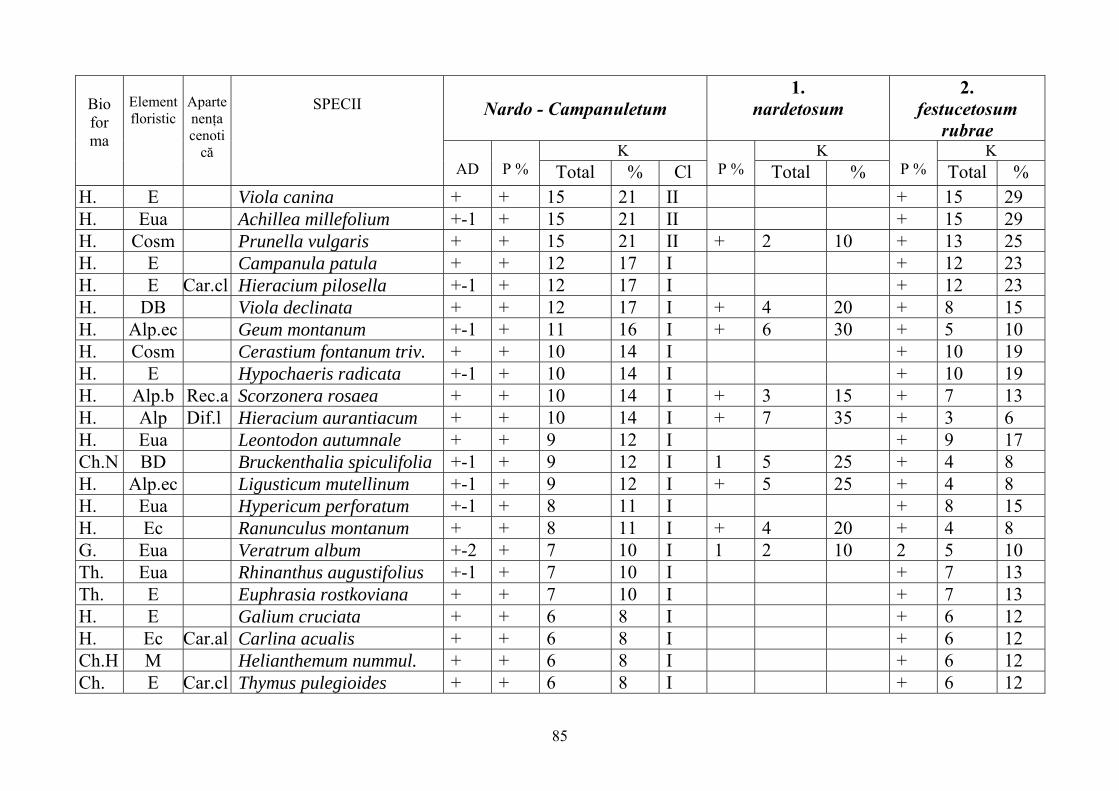
85
Bio for ma
Element floristic
Apartenenţa cenoti
că
SPECII
Nardo - Campanuletum
1. nardetosum
2. festucetosum
rubrae
AD
P % K
P % K
P % K
Total % Cl Total % Total % H. E Viola canina + + 15 21 II + 15 29 H. Eua Achillea millefolium +-1 + 15 21 II + 15 29 H. Cosm Prunella vulgaris + + 15 21 II + 2 10 + 13 25 H. E Campanula patula + + 12 17 I + 12 23 H. E Car.cl Hieracium pilosella +-1 + 12 17 I + 12 23 H. DB Viola declinata + + 12 17 I + 4 20 + 8 15 H. Alp.ec Geum montanum +-1 + 11 16 I + 6 30 + 5 10 H. Cosm Cerastium fontanum triv. + + 10 14 I + 10 19 H. E Hypochaeris radicata +-1 + 10 14 I + 10 19 H. Alp.b Rec.a Scorzonera rosaea + + 10 14 I + 3 15 + 7 13 H. Alp Dif.l Hieracium aurantiacum + + 10 14 I + 7 35 + 3 6 H. Eua Leontodon autumnale + + 9 12 I + 9 17 Ch.N BD Bruckenthalia spiculifolia +-1 + 9 12 I 1 5 25 + 4 8 H. Alp.ec Ligusticum mutellinum +-1 + 9 12 I + 5 25 + 4 8 H. Eua Hypericum perforatum +-1 + 8 11 I + 8 15 H. Ec Ranunculus montanum + + 8 11 I + 4 20 + 4 8 G. Eua Veratrum album +-2 + 7 10 I 1 2 10 2 5 10 Th. Eua Rhinanthus augustifolius +-1 + 7 10 I + 7 13 Th. E Euphrasia rostkoviana + + 7 10 I + 7 13 H. E Galium cruciata + + 6 8 I + 6 12 H. Ec Car.al Carlina acualis + + 6 8 I + 6 12 Ch.H M Helianthemum nummul. + + 6 8 I + 6 12 Ch. E Car.cl Thymus pulegioides + + 6 8 I + 6 12

86
Bio
forma
Element floristic
Apartenenţa cenoti
că
SPECII
Nardo - Campanuletum
1. nardetosum
2. festucetosum
rubrae
AD
P % K
P % K
P % K
Total % Cl Total % Total % G. B Crocus heufelianus + + 6 8 I + 4 20 + 2 4 H. End Car.al Campanula serrata + + 5 7 I + 2 10 + 3 6 H. Eua Hypericum maculatum + + 5 7 I + 2 10 + 3 6 Ch.H E Veronica chamaedrys + + 5 7 I + 5 10 Ch.N Cp Vaccinum uliginosum +-1 + 5 7 I + 4 15 + 1 2 H. Cp Polygonum bistorta + + 5 7 I + 2 10 + 3 6 TH.H Eua Carlina vulgaris +-1 + 5 7 I + 5 10 H. Eua Ranunculus polyanthemos + + 4 6 I + 4 8 H. Alp.b Gentiana praecox + + 4 6 I + 4 8 H. E Bellis perennis + + 3 4 I + 3 6 H. Eua Plantago media + + 3 4 I + 3 6 H. Alp Pedicularis verticillata + + 3 4 I + 1 5 + 2 4 Ch. Cp Car.cl Antenaria dioica + + 3 4 I + 3 6 H-G E Ranunculus bulbosus + + 2 3 I + 2 4 H. Ec Diarithus carthusianorum + + 1 1 I + 1 2 TH. Eua Carum carvi + + 1 1 I + 1 2 G. E Colchicum autumnale + + 1 1 I + 1 2 Th. Ec Car.ord Euphrasia stricta + + 1 1 I + 1 2 TH.H Eua Tragopogon orientalis + + 1 1 I + 1 2 Specii lemnoase (puieţi) - + (3sp) - - (1) (2sp) - + (3sp) - M. Cp Juniperus comunnis +-1 + 16 23 II 1 9 45 + 7 13 M.M. E Picea albies +-1 + 5 7 I + 3 15 + 2 4 M.M. Eua Betula verrucosa + + 1 1 I + 1 2
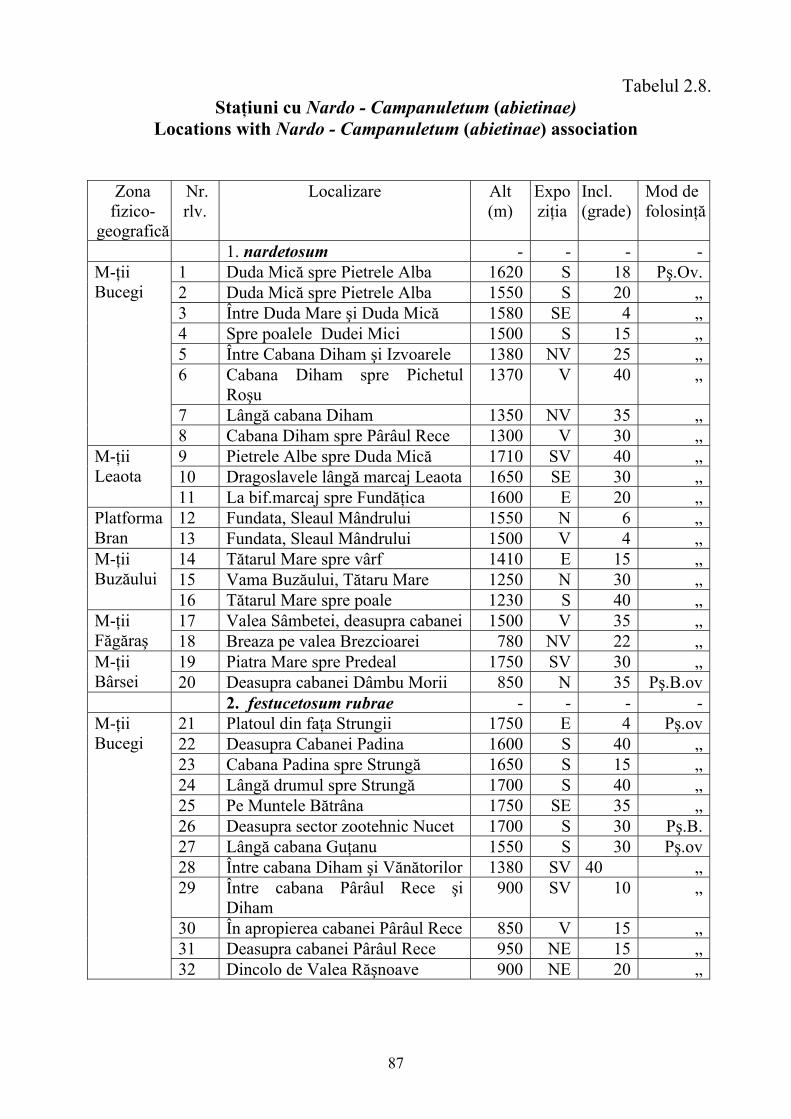
87
Tabelul 2.8. Staţiuni cu Nardo - Campanuletum (abietinae)
Locations with Nardo - Campanuletum (abietinae) association
Zona
fizico-geografică
Nr. rlv.
Localizare Alt (m)
Expo ziţia
Incl. (grade)
Mod de folosinţă
1. nardetosum - - - -M-ţii Bucegi
1 Duda Mică spre Pietrele Alba 1620 S 18 Pş.Ov.2 Duda Mică spre Pietrele Alba 1550 S 20 „3 Între Duda Mare şi Duda Mică 1580 SE 4 „4 Spre poalele Dudei Mici 1500 S 15 „5 Între Cabana Diham şi Izvoarele 1380 NV 25 „6 Cabana Diham spre Pichetul
Roşu 1370 V 40 „
7 Lângă cabana Diham 1350 NV 35 „8 Cabana Diham spre Pârâul Rece 1300 V 30 „
M-ţii Leaota
9 Pietrele Albe spre Duda Mică 1710 SV 40 „10 Dragoslavele lângă marcaj Leaota 1650 SE 30 „11 La bif.marcaj spre Fundăţica 1600 E 20 „
Platforma Bran
12 Fundata, Sleaul Mândrului 1550 N 6 „13 Fundata, Sleaul Mândrului 1500 V 4 „
M-ţii Buzăului
14 Tătarul Mare spre vârf 1410 E 15 „15 Vama Buzăului, Tătaru Mare 1250 N 30 „16 Tătarul Mare spre poale 1230 S 40 „
M-ţii Făgăraş
17 Valea Sâmbetei, deasupra cabanei 1500 V 35 „18 Breaza pe valea Brezcioarei 780 NV 22 „
M-ţii Bârsei
19 Piatra Mare spre Predeal 1750 SV 30 „20 Deasupra cabanei Dâmbu Morii 850 N 35 Pş.B.ov
2. festucetosum rubrae - - - -M-ţii Bucegi
21 Platoul din faţa Strungii 1750 E 4 Pş.ov22 Deasupra Cabanei Padina 1600 S 40 „23 Cabana Padina spre Strungă 1650 S 15 „24 Lângă drumul spre Strungă 1700 S 40 „25 Pe Muntele Bătrâna 1750 SE 35 „26 Deasupra sector zootehnic Nucet 1700 S 30 Pş.B.27 Lângă cabana Guţanu 1550 S 30 Pş.ov28 Între cabana Diham şi Vănătorilor 1380 SV 40 „29 Între cabana Pârâul Rece şi
Diham 900 SV 10 „
30 În apropierea cabanei Pârâul Rece 850 V 15 „31 Deasupra cabanei Pârâul Rece 950 NE 15 „32 Dincolo de Valea Răşnoave 900 NE 20 „

88
Zona fizico-geograf.
Nr. rlv.
Localizare Alt (m)
Expo ziţia
Incl. (grade)
Mod defolosinţă
Platforma Bran
33 Fundata spre Plaiul Mândrului 1600 NV 38 Pş.ov34 Fundata la cca. 3 km V de şcoală 1260 SV 6 „35 Sub Cabana Guţanu, Moeciu 1300 S 35 „36 Fundata, marcaj spre Padina 1230 S 22 „37 Şirnea la 2 km NE de centru 1220 E 15 Mixt38 Şirnea mai jos rel. 67 1190 E 25 „39 Şirnea deasupra re. 67 1250 NV 40 „
40 Fundata lângă şcoală 1240 NE 15 Pş.ovM-ţii Făgăraş
41 Valea Viştei Mari sub grohotiş 1620 SE 45 „42 Valea Viştei Mari în căldare 1600 E 40 „43 Valea Sâmbetei în căldarea gla. 1650 N 6 „44 Deasupra cabanei Sâmbăta 1500 NE 30 „
M-ţii Bârsei
45 Piatra Mare spre Tamina 1550 SE 35 Pş.ov46 Între Tamina şi Timişu de Sus 1120 NE 20 „47 Deasupra cabanei Dâmbu Morii 850 N 32 Pş.B.ov48 Pe traseul Dâmbu Morii, P. Mare 980 E 30 „
M-ţii Poiana Braşov
49 Poiana Mică lângă post „Trafo” 1000 E 8 Pş.B.ov50 Poiana Mică sub post „Trafo” 960 V 8 „51 Poiana Mică lângă cabana
Vănători 950 SE 6
52 P. Mică drumul spre loc.Cristian 940 NE 30 „53 Poiana Mică lângă câmp experim. 920 SV 6 „54 Poiana Mică deasupra câmp exp. 930 S 4 „55 Poiana Mică lângă cab.
Vănătorul 930 SV 5 „
56 Poiana Mică lângă rezerv.zimbri 940 SE 8 „57 Poiana Mică sub tăietură lângă
şos. 960 V 5 „
58 Poiana Mică lângă post „Trafo” 950 V 4 „59 Poiana Mică lângă adăpost
animale 950 E 2 „
60 Poiana Mică lângă staţia Meteo 950 SV 2 „61 Cristian spre Poiana Mică 970 SE 15 „
M-ţii Codlei
62 Poiana Mărului,Valea lui Paul 1050 NV 25 Pş.B.63 Poiana Mărului deasupra b.
Fierului 1020 V 5 Pş.B.ov
64 Zărneşti alea Bârsei Fierului 870 NV 35 Pş.ov.M-ţii Ciucaş
65 Sub Vf. Teslei spre Poiană 1430 E 15 Pş.ov66 Poiana Teslei 1420 NE 30 „
M-ţii Piatra Craiului
67 Zărneşti deasupra Văii Bârsei 1010 N 15 Mixt68 Zărneşti deasupra Văii Bârsei 920 NE 8 „69 Zărneşti deasupra Bârsei 910 SV 25 „
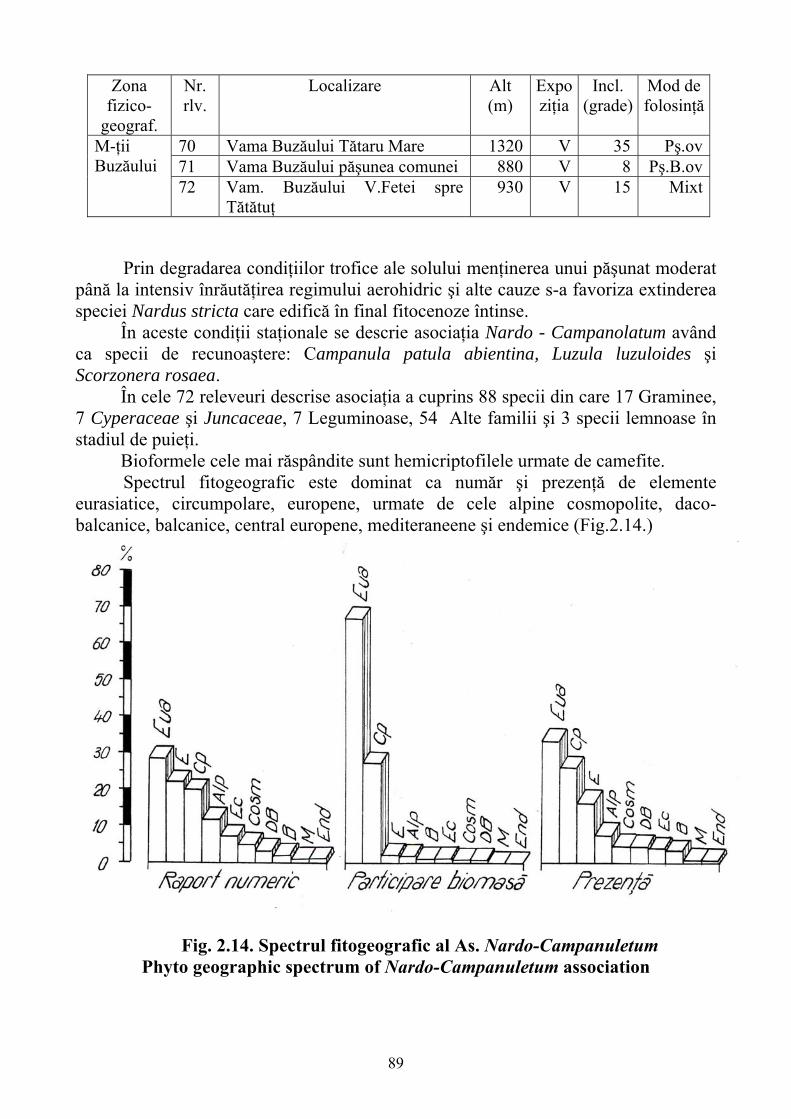
89
Zona fizico-
geograf.
Nr. rlv.
Localizare Alt (m)
Expo ziţia
Incl. (grade)
Mod de folosinţă
M-ţii Buzăului
70 Vama Buzăului Tătaru Mare 1320 V 35 Pş.ov71 Vama Buzăului păşunea comunei 880 V 8 Pş.B.ov72 Vam. Buzăului V.Fetei spre
Tătătuţ 930 V 15 Mixt
Prin degradarea condiţiilor trofice ale solului menţinerea unui păşunat moderat până la intensiv înrăutăţirea regimului aerohidric şi alte cauze s-a favoriza extinderea speciei Nardus stricta care edifică în final fitocenoze întinse. În aceste condiţii staţionale se descrie asociaţia Nardo - Campanolatum având ca specii de recunoaştere: Campanula patula abientina, Luzula luzuloides şi Scorzonera rosaea. În cele 72 releveuri descrise asociaţia a cuprins 88 specii din care 17 Graminee, 7 Cyperaceae şi Juncaceae, 7 Leguminoase, 54 Alte familii şi 3 specii lemnoase în stadiul de puieţi. Bioformele cele mai răspândite sunt hemicriptofilele urmate de camefite.
Spectrul fitogeografic este dominat ca număr şi prezenţă de elemente eurasiatice, circumpolare, europene, urmate de cele alpine cosmopolite, daco-balcanice, balcanice, central europene, mediteraneene şi endemice (Fig.2.14.)
Fig. 2.14. Spectrul fitogeografic al As. Nardo-Campanuletum Phyto geographic spectrum of Nardo-Campanuletum association

90
În biomasă participă în proporţie de 2/3 elementele eurasiatice urmate de cca. 27 % circumpolare şi altele în proporţie de sub 2 % fiecare.
Se poate observa că în asociaţia Nardo - Campanulatum speciile eurasiatice circumpolare şi europene sunt forte bine reprezentate iar speciile de origine alpine - subalpine sunt mult mai puţine decât în nardetele subalpine. În plus apar elementele cosmopolite şi mediterane care nu se întâlnesc în nardetele alpine. În cadrul asociaţiei Nardo - Campanulatum s-au conturat pe baza speciilor diferenţiale ecologice, un număr de două subasociaţii şi anume 1. Nardetum şi 2. Festucatosum rubrae.
2.4.2.1. Nardo - Campanuletum nardetosum (Syn Nardetum strictae montanum nardetosum (typicum) Buia, 63) Este identificată în 20 de staţiuni luate la altitudini cuprinse între 1230-1710 m, pe pante însorite cu înclinaţie de 15 - 400. Situată în etajul molidului pe soluri brune podzolice feriiluviale (Bfe) şi podzolice brune feriiluviale (P) această subasociaţie se întâlneşte pe solurile cele mai acide şi cu trofismul cel mai scăzut între fitocenozele edificate de Nardus stricta în zona montană. În aceste condiţii specia Nardus stricta participă în proporţie de 82 % în biomasă, cel mai ridicat procent în fitocenozele edificate de această specie în zona luată în studiu. Ca specii diferenţiale ecologice pentru această subasociaţie au fost desemnate: Vaccinum myrtillus,Vaccinum vitis idaea şi Hieracium aurantiacum. Ecodiagrama speciei Vaccinum myrtillus indică cele mai acide soluri pe care se întâlnesc nardetele în judeţul Braşov (Fig. 2.15.)
La acestea se adaugă caracterul lor oligobazic, extrem oligotrofic foarte puternic humifer cu un conţinut foarte ridicat în Al mobil şi mijlociu aprovizionate în potasiu. Producţia de masă verde este de 3,2 t/ha în medie şi este de o calitate foarte scăzută. Valoarea pastorală medie este de 8 fiind de asemenea foarte scăzută.
2.4.2.2. Nardo - Campanuletum festucetosum rubrae Buia, 63 (Syn Nardetum strictae montanum festucetosum rubrae, Buia, 63) A fost identificată în 52 staţiuni situate la altitudini cuprinse între 870 - 1750 m
pe diferite expoziţii şi înclinaţii de 5 – 35 0. Subasociaţia se întâlneşte în etajul pădurilor de molid şi la altitudini nai joase
în etajul pădurilor de amestec între molid şi fag, şi uneori şi a celor de fag din întreaga zonă montană luată în studiu.
Solurile sunt de tipul brune acide (BC), brune podzolite (BP) pe alocuri brune podzolice feriiluviale (Bfe) mai puţin acide şi cu un grad de troficitate ceva mai ridicat decât ale subasociaţiei tipice descrise mai înainte.
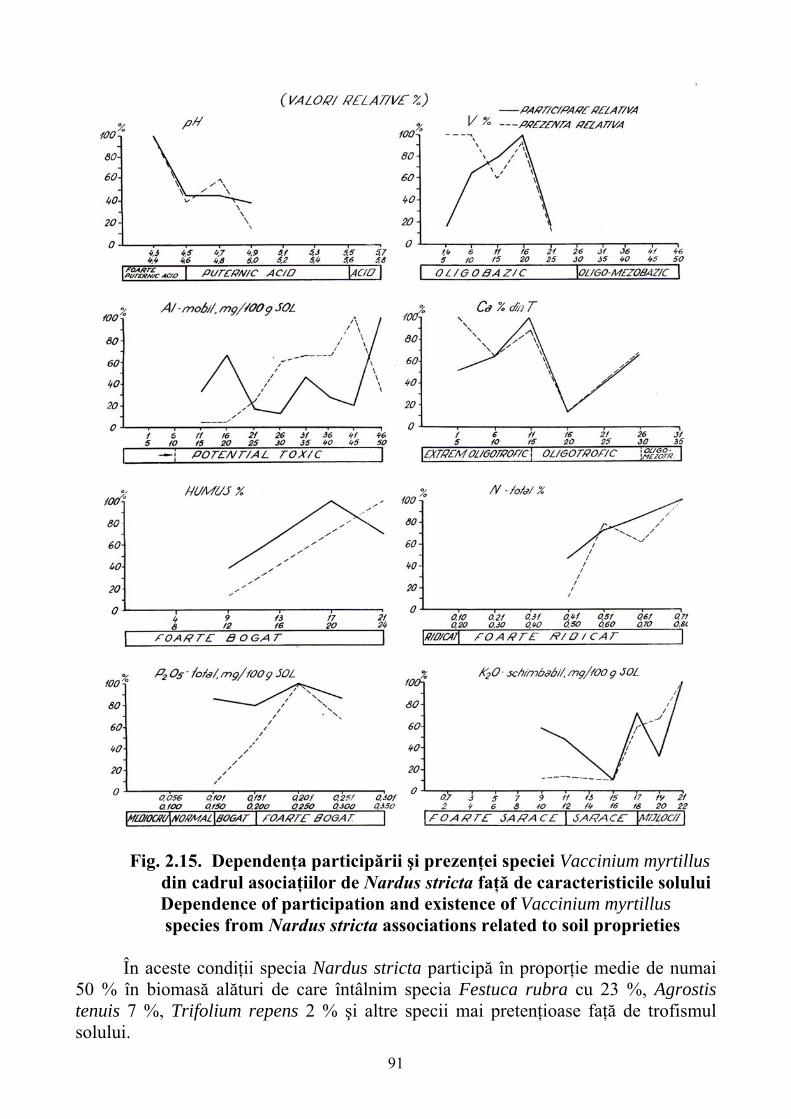
91
Fig. 2.15. Dependenţa participării şi prezenţei speciei Vaccinium myrtillus din cadrul asociaţiilor de Nardus stricta faţă de caracteristicile solului
Dependence of participation and existence of Vaccinium myrtillus species from Nardus stricta associations related to soil proprieties
În aceste condiţii specia Nardus stricta participă în proporţie medie de numai
50 % în biomasă alături de care întâlnim specia Festuca rubra cu 23 %, Agrostis tenuis 7 %, Trifolium repens 2 % şi altre specii mai pretenţioase faţă de trofismul solului.

92
Specia Festuca rubra este considerată diferenţială ecologică a subasociaţiei. La fel participarea neînsemnată a speciilor Vaccinium myrtillus şi Vaccinum vitis idaea este un indiciu de diferenţiere faţă de subasociaţia Nardo – Campanuletum nardetosum unde aceste specii sunt bine reprezentate.
Ecodiagrama speciei Festuca rubra indică optimum de participare şi prezenţă relativă la un pH 4,9-5,0 pe soliri oligobazice, humifere, cu un conţinut ridicat de Al mobil şi P2O5 total, relativ indiferentă faţă de K2O schimbabil.
Din punct de vedere al evoluţiei Nardo - Campanuletum festucetosum rubrae face legătura între fitocenozele de Festuca rubra şi nardetele tipice reprezentând un stadiu de degradare a acestor pajişti.
Producţia de masă verde se estimează la cca. 3,6 t/ha, fiind de o calitate mediocră spre slabă.
Valoarea pastorală este de 28 fiind de asemenea destul de scăzută. Productivitatea potenţială a subasociaţiei este mai ridicată datorită
posibilităţilor oferite de îmbunătăţirea condiţiilor de troficitate prin fertilizare, când specia Festuca rubra poate înlătura cu uşurinţă din covorul ierbos specia dominantă Nardus stricta.
2.4.3. As Nardo - Juncetum (conglomerati) Resmeriţă, 69 (Syn Nardetum strictae collinum, Buia 63; Nardetum strictae submontanum,
Resmeriţă et al. 63; As de Nardus stricta, Safta 43) Prezenţa cu indici de frecvenţă şi constanţă ridicaţi ai speciei Juncus
conglomeratus în nardetele colinare şi depresionare (atât în cele cu exces pronunţat de umiditate cât şi în cele relativ uscate) ne-a determinat să le încadrăm în asociaţia Nardo - Juncetum (conglomerati).
Putem confirma că între asociaţia Nardo - Juncetum (conglomerati) descrisă în această lucrare şi Nardo - Juncetum conglomerati – effusi există numai o similitudine nomenclatorală, asociaţia fiind dealtfel o subasociaţie (Nardo - Juncetum molinietosum sau Agrostidetosum canini).
Nardetele de Juncus conglomeratus din ţara noastră ar constitui o asociaţie paralelă cu Nardo - Juncetum squarrosi (Nordh 20 ) Bűk 42, care este descrisă în zonele planare din vestul şi centrul Europei. Este remarcabil faptul că paralelismul celor două asociaţii se continuă şi la nivelul subunităţilor pe care le cuprind. Astfel, în cadrul asociaţiei Nardo - Juncetum squarrosi sunt menţionate de OBERDORFER E. 1957, trei subasociaţii:
- agrostidetosum tenuis PREISG, 51, condiţii relativ uscate ce conduce spre Nardo - Galion.
- molinietosum, subas.nov.pe terenuri alternativ umede. - eriophoretosum angustifolii PREISG 51, pe terenuri permanent umede,
mijlocind spre Caricion canescenti - fuscae. Corespondenţa subunităţilor asociaţiei Nardo-Juncetum (conglomerati)
identificate în zona cercetată sunt: - agrostidetosum tenuis, care se individualizează prin speciile Agrostis
capillaris, Calamagrostis arundinacea, Euphorbia cyparissias, Dorychnium herbaceum şi altele, ce evidenţiează condiţii staţionale mai uscate.

93
- molinietosum, care are ca diferenţiele ecologice speciile Molinia coerulea, Gentiana pneumonanthe, Achillea ptarmica şi altele fiind legată de staţiuni cu regim de umiditate temporară spre permanentă.
- eriophoretosum latifolii, care grupează nardetele cu caracter de trecere spre unităţile clasei Scheuchzerio - Caricetae nigrae Nordh 36, şi se diferenţiează prin speciile Carex nigra (fusca), Eriophorum latifolium, Sphagnum sp., Polytrichum sp. şi alte specii legate de terenuri permanent umede, îmmlăştinate.
La acestea se mai adaugă şi subas. Nardo - Juncetum agrostidetosum canini care evidenţiează staţiuni cu umiditate temporară, având ca diferenţială principală, specia Agrostis canina.
Asociaţia Nardo - Juncetum (conglomerati) a fost întâlnită în 103 staţiuni cuprins între 480-820 m altitudine în Depresiunile Făgăraş, Vlădeni, Homoroadelor şi Bârsei, cât şi pe toate dealurile ce însoţesc munţii din judeţul Braşov (Tabel 2.9.-2.10.). Asociaţia este răspândită îndeosebi pe terenuri plane şi pante cu înclinaţie de până la 20 % şi expoziţii umbrite.
Temperatura medie anuală este cuprinsă între 4 - 8 0C şi precipitaţiile ating 700-900 mm anual.
Vegetaţia primară este reprezentată de păduri de fag în partea superioară, amestec de fag şi gorun urmată de cele de gorun şi stejar în zonele cele mai joase.
În aceste condiţii solurile predominante sunt: argiloiluviale, brune podzolite (Bpo), brune acide (BO), podzolice argiloiluviale (SPn), argiloiluviale pseudogleice (TP), aluviale frecvent gleizate (SA) şi altele.
După defrişarea pădurilor de fag şi gorun şi trecerea prin diferite stadii ierboase de tâietură în prezenţa păşunatului şi cosirii se instalează fitocenoze edificate în principal de speciile: Agrostis tenuis, Festuca rubra, Agrostis canina, Festuca pratensis şi Festuca valesiaca. Un loc aparte îl ocupă fitocenozele edificate de Molinia coerulea care se consideră a avea un caracter relictar RESMERIŢĂ I., 1969.
Înlăturarea vegetaţiei lemnoase a înrăutăţit regimul aerohidric al solului, suprafeţe mari fiind supuse excesului temporar de umiditate. La aceste fenomene se adaugă şi modul de folosire necorespunzător ce a dus la degradarea continuă a condiţiilor trofice care a favorizat extinderea speciei Nardus stricta în cele mai diferite fitocenoze.
În aceste condiţii staţionale se descrie asociaţia Nardo - Juncetum (conglomeraţi), având ca specii de recunoaştere: Juncus conglomeratus, ranunculus polyatemos,Succisa pratensis.
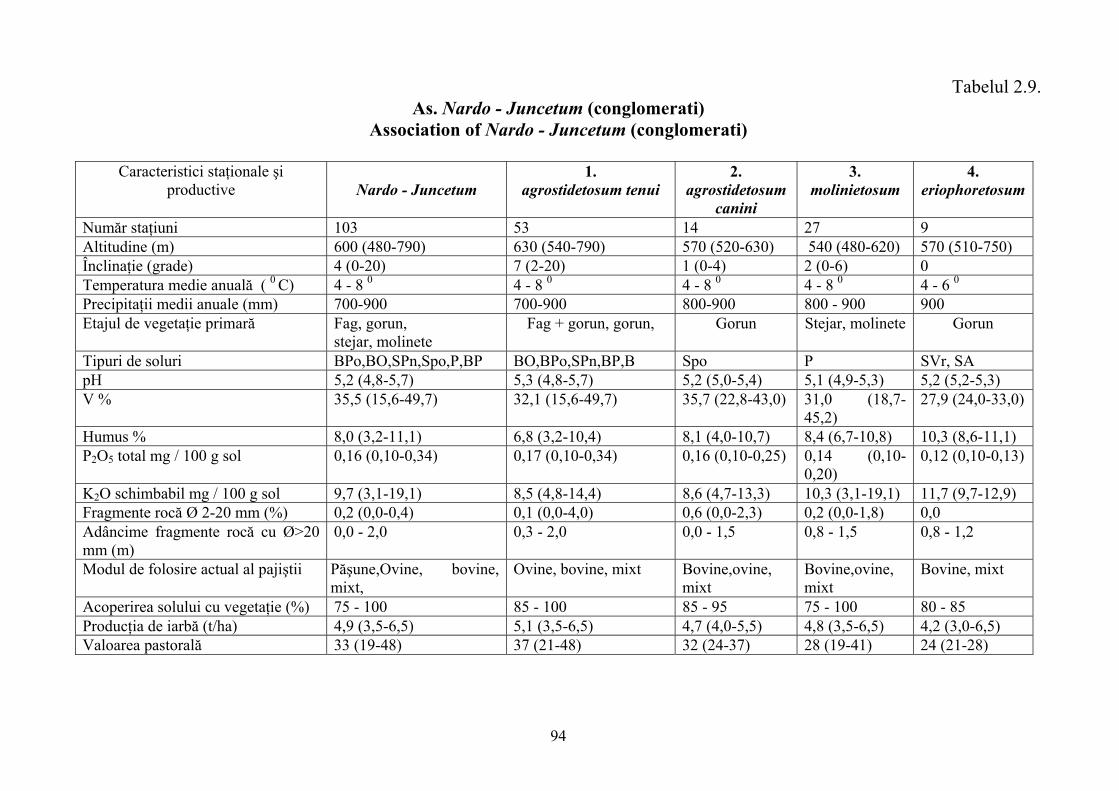
94
Tabelul 2.9. As. Nardo - Juncetum (conglomerati)
Association of Nardo - Juncetum (conglomerati)
Caracteristici staţionale şi productive
Nardo - Juncetum
1. agrostidetosum tenui
2. agrostidetosum
canini
3. molinietosum
4. eriophoretosum
Număr staţiuni 103 53 14 27 9 Altitudine (m) 600 (480-790) 630 (540-790) 570 (520-630) 540 (480-620) 570 (510-750) Înclinaţie (grade) 4 (0-20) 7 (2-20) 1 (0-4) 2 (0-6) 0 Temperatura medie anuală ( 0 C) 4 - 8 0 4 - 8 0 4 - 8 0 4 - 8 0 4 - 6 0 Precipitaţii medii anuale (mm) 700-900 700-900 800-900 800 - 900 900 Etajul de vegetaţie primară Fag, gorun,
stejar, molinete Fag + gorun, gorun, Gorun Stejar, molinete Gorun
Tipuri de soluri BPo,BO,SPn,Spo,P,BP BO,BPo,SPn,BP,B Spo P SVr, SA pH 5,2 (4,8-5,7) 5,3 (4,8-5,7) 5,2 (5,0-5,4) 5,1 (4,9-5,3) 5,2 (5,2-5,3) V % 35,5 (15,6-49,7) 32,1 (15,6-49,7) 35,7 (22,8-43,0) 31,0 (18,7-
45,2) 27,9 (24,0-33,0)
Humus % 8,0 (3,2-11,1) 6,8 (3,2-10,4) 8,1 (4,0-10,7) 8,4 (6,7-10,8) 10,3 (8,6-11,1) P2O5 total mg / 100 g sol 0,16 (0,10-0,34) 0,17 (0,10-0,34) 0,16 (0,10-0,25) 0,14 (0,10-
0,20) 0,12 (0,10-0,13)
K2O schimbabil mg / 100 g sol 9,7 (3,1-19,1) 8,5 (4,8-14,4) 8,6 (4,7-13,3) 10,3 (3,1-19,1) 11,7 (9,7-12,9) Fragmente rocă Ø 2-20 mm (%) 0,2 (0,0-0,4) 0,1 (0,0-4,0) 0,6 (0,0-2,3) 0,2 (0,0-1,8) 0,0 Adâncime fragmente rocă cu Ø>20 mm (m)
0,0 - 2,0 0,3 - 2,0 0,0 - 1,5 0,8 - 1,5 0,8 - 1,2
Modul de folosire actual al pajiştii Păşune,Ovine, bovine, mixt,
Ovine, bovine, mixt Bovine,ovine, mixt
Bovine,ovine, mixt
Bovine, mixt
Acoperirea solului cu vegetaţie (%) 75 - 100 85 - 100 85 - 95 75 - 100 80 - 85 Producţia de iarbă (t/ha) 4,9 (3,5-6,5) 5,1 (3,5-6,5) 4,7 (4,0-5,5) 4,8 (3,5-6,5) 4,2 (3,0-6,5) Valoarea pastorală 33 (19-48) 37 (21-48) 32 (24-37) 28 (19-41) 24 (21-28)
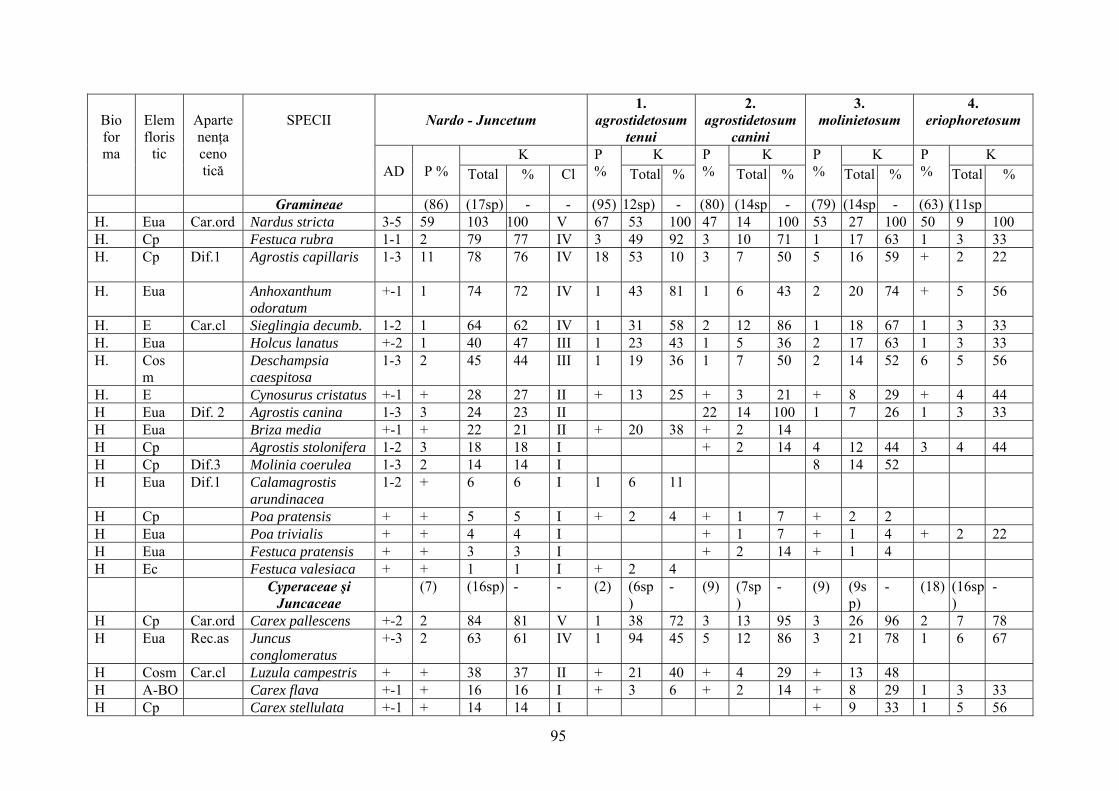
95
Bio for ma
Elem floris
tic
Aparte nenţa ceno tică
SPECII
Nardo - Juncetum
1. agrostidetosum
tenui
2. agrostidetosum
canini
3. molinietosum
4. eriophoretosum
AD
P %
K P %
K P %
K P %
K P %
K Total % Cl Total % Total % Total % Total %
Gramineae (86) (17sp) - - (95) (12sp) - (80) (14sp - (79) (14sp - (63) (11sp H. Eua Car.ord Nardus stricta 3-5 59 103 100 V 67 53 100 47 14 100 53 27 100 50 9 100 H. Cp Festuca rubra 1-1 2 79 77 IV 3 49 92 3 10 71 1 17 63 1 3 33 H. Cp Dif.1 Agrostis capillaris 1-3 11 78 76 IV 18 53 10 3 7 50 5 16 59 + 2 22
H. Eua Anhoxanthum odoratum
+-1 1 74 72 IV 1 43 81 1 6 43 2 20 74 + 5 56
H. E Car.cl Sieglingia decumb. 1-2 1 64 62 IV 1 31 58 2 12 86 1 18 67 1 3 33 H. Eua Holcus lanatus +-2 1 40 47 III 1 23 43 1 5 36 2 17 63 1 3 33 H. Cos
m Deschampsia
caespitosa 1-3 2 45 44 III 1 19 36 1 7 50 2 14 52 6 5 56
H. E Cynosurus cristatus +-1 + 28 27 II + 13 25 + 3 21 + 8 29 + 4 44 H Eua Dif. 2 Agrostis canina 1-3 3 24 23 II 22 14 100 1 7 26 1 3 33 H Eua Briza media +-1 + 22 21 II + 20 38 + 2 14 H Cp Agrostis stolonifera 1-2 3 18 18 I + 2 14 4 12 44 3 4 44 H Cp Dif.3 Molinia coerulea 1-3 2 14 14 I 8 14 52 H Eua Dif.1 Calamagrostis
arundinacea 1-2 + 6 6 I 1 6 11
H Cp Poa pratensis + + 5 5 I + 2 4 + 1 7 + 2 2 H Eua Poa trivialis + + 4 4 I + 1 7 + 1 4 + 2 22 H Eua Festuca pratensis + + 3 3 I + 2 14 + 1 4 H Ec Festuca valesiaca + + 1 1 I + 2 4 Cyperaceae şi
Juncaceae (7) (16sp) - - (2) (6sp
) - (9) (7sp
) - (9) (9s
p) - (18) (16sp
) -
H Cp Car.ord Carex pallescens +-2 2 84 81 V 1 38 72 3 13 95 3 26 96 2 7 78 H Eua Rec.as Juncus
conglomeratus +-3 2 63 61 IV 1 94 45 5 12 86 3 21 78 1 6 67
H Cosm Car.cl Luzula campestris + + 38 37 II + 21 40 + 4 29 + 13 48 H A-BO Carex flava +-1 + 16 16 I + 3 6 + 2 14 + 8 29 1 3 33 H Cp Carex stellulata +-1 + 14 14 I + 9 33 1 5 56
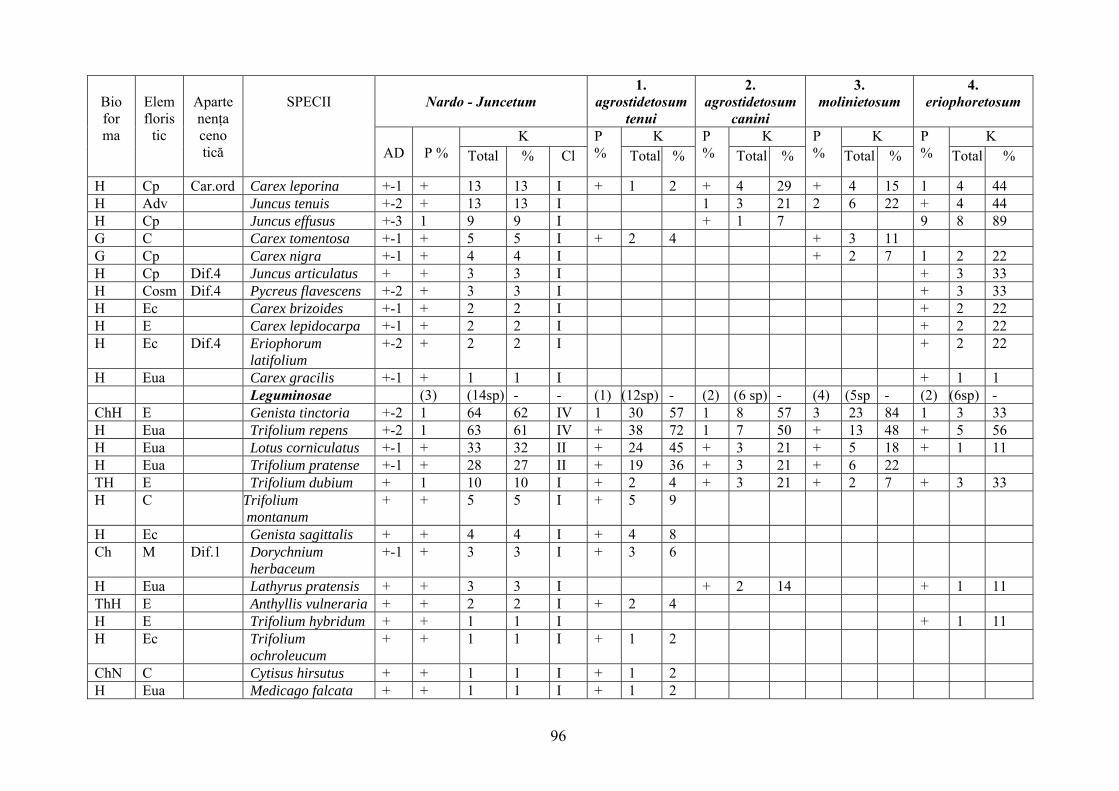
96
Bio for ma
Elem floris
tic
Aparte nenţa ceno tică
SPECII
Nardo - Juncetum
1. agrostidetosum
tenui
2. agrostidetosum
canini
3. molinietosum
4. eriophoretosum
AD
P %
K P %
K P %
K P %
K P %
K Total % Cl Total % Total % Total % Total %
H Cp Car.ord Carex leporina +-1 + 13 13 I + 1 2 + 4 29 + 4 15 1 4 44 H Adv Juncus tenuis +-2 + 13 13 I 1 3 21 2 6 22 + 4 44 H Cp Juncus effusus +-3 1 9 9 I + 1 7 9 8 89 G C Carex tomentosa +-1 + 5 5 I + 2 4 + 3 11 G Cp Carex nigra +-1 + 4 4 I + 2 7 1 2 22 H Cp Dif.4 Juncus articulatus + + 3 3 I + 3 33 H Cosm Dif.4 Pycreus flavescens +-2 + 3 3 I + 3 33 H Ec Carex brizoides +-1 + 2 2 I + 2 22 H E Carex lepidocarpa +-1 + 2 2 I + 2 22 H Ec Dif.4 Eriophorum
latifolium +-2 + 2 2 I + 2 22
H Eua Carex gracilis +-1 + 1 1 I + 1 1 Leguminosae (3) (14sp) - - (1) (12sp) - (2) (6 sp) - (4) (5sp - (2) (6sp) - ChH E Genista tinctoria +-2 1 64 62 IV 1 30 57 1 8 57 3 23 84 1 3 33 H Eua Trifolium repens +-2 1 63 61 IV + 38 72 1 7 50 + 13 48 + 5 56 H Eua Lotus corniculatus +-1 + 33 32 II + 24 45 + 3 21 + 5 18 + 1 11 H Eua Trifolium pratense +-1 + 28 27 II + 19 36 + 3 21 + 6 22 TH E Trifolium dubium + 1 10 10 I + 2 4 + 3 21 + 2 7 + 3 33 H C Trifolium
montanum + + 5 5 I + 5 9
H Ec Genista sagittalis + + 4 4 I + 4 8 Ch M Dif.1 Dorychnium
herbaceum +-1 + 3 3 I + 3 6
H Eua Lathyrus pratensis + + 3 3 I + 2 14 + 1 11 ThH E Anthyllis vulneraria + + 2 2 I + 2 4 H E Trifolium hybridum + + 1 1 I + 1 11 H Ec Trifolium
ochroleucum + + 1 1 I + 1 2
ChN C Cytisus hirsutus + + 1 1 I + 1 2 H Eua Medicago falcata + + 1 1 I + 1 2
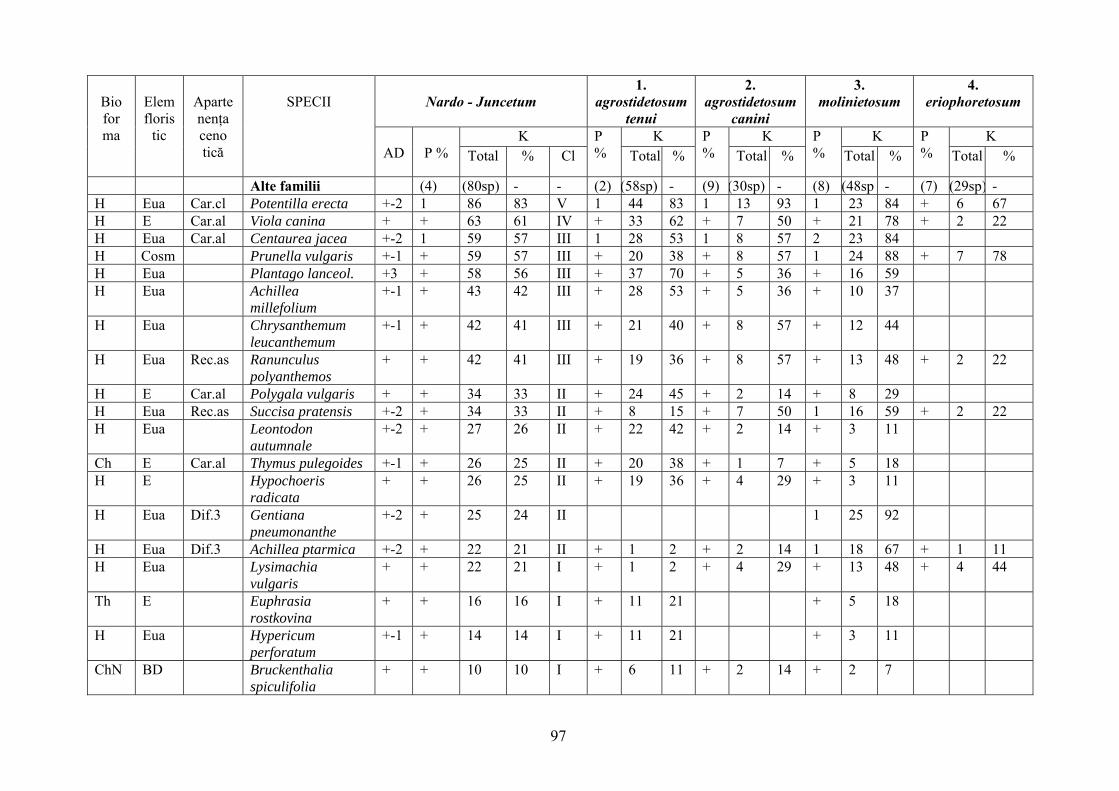
97
Bio for ma
Elem floris
tic
Aparte nenţa ceno tică
SPECII
Nardo - Juncetum
1. agrostidetosum
tenui
2. agrostidetosum
canini
3. molinietosum
4. eriophoretosum
AD
P %
K P %
K P %
K P %
K P %
K Total % Cl Total % Total % Total % Total %
Alte familii (4) (80sp) - - (2) (58sp) - (9) (30sp) - (8) (48sp - (7) (29sp) - H Eua Car.cl Potentilla erecta +-2 1 86 83 V 1 44 83 1 13 93 1 23 84 + 6 67 H E Car.al Viola canina + + 63 61 IV + 33 62 + 7 50 + 21 78 + 2 22 H Eua Car.al Centaurea jacea +-2 1 59 57 III 1 28 53 1 8 57 2 23 84 H Cosm Prunella vulgaris +-1 + 59 57 III + 20 38 + 8 57 1 24 88 + 7 78 H Eua Plantago lanceol. +3 + 58 56 III + 37 70 + 5 36 + 16 59 H Eua Achillea
millefolium +-1 + 43 42 III + 28 53 + 5 36 + 10 37
H Eua Chrysanthemum leucanthemum
+-1 + 42 41 III + 21 40 + 8 57 + 12 44
H Eua Rec.as Ranunculus polyanthemos
+ + 42 41 III + 19 36 + 8 57 + 13 48 + 2 22
H E Car.al Polygala vulgaris + + 34 33 II + 24 45 + 2 14 + 8 29 H Eua Rec.as Succisa pratensis +-2 + 34 33 II + 8 15 + 7 50 1 16 59 + 2 22 H Eua Leontodon
autumnale +-2 + 27 26 II + 22 42 + 2 14 + 3 11
Ch E Car.al Thymus pulegoides +-1 + 26 25 II + 20 38 + 1 7 + 5 18 H E Hypochoeris
radicata + + 26 25 II + 19 36 + 4 29 + 3 11
H Eua Dif.3 Gentiana pneumonanthe
+-2 + 25 24 II 1 25 92
H Eua Dif.3 Achillea ptarmica +-2 + 22 21 II + 1 2 + 2 14 1 18 67 + 1 11 H Eua Lysimachia
vulgaris + + 22 21 I + 1 2 + 4 29 + 13 48 + 4 44
Th E Euphrasia rostkovina
+ + 16 16 I + 11 21 + 5 18
H Eua Hypericum perforatum
+-1 + 14 14 I + 11 21 + 3 11
ChN BD Bruckenthalia spiculifolia
+ + 10 10 I + 6 11 + 2 14 + 2 7

98
Bio for ma
Elem floris
tic
Aparte nenţa ceno tică
SPECII
Nardo - Juncetum
1. agrostidetosum
tenui
2. agrostidetosum
canini
3. molinietosum
4. eriophoretosum
AD
P %
K P %
K P %
K P %
K P %
K Total % Cl Total % Total % Total % Total %
Th Adv Erigeron annuus + + 10 10 I + 5 9 + 1 7 + 4 15 Th Eua Centaurium
erytraea + + 10 10 I + 7 13 + 3 11
Ch Cp Car.al Veronica officin. + + 10 9 I + 4 8 + 2 14 + 3 11 Th Eua Rhinanthus
augustifolius + + 9 9 I + 4 8 + 1 7 + 4 15
G Eua Veratrum album +-2 + 9 9 I + 2 4 2 2 14 + 5 18 H Eua Leontodon hispidus +-1 + 9 9 I + 4 8 + 3 21 + 4 15 H Eua Serratula tinctoria + + 8 8 I + 1 7 + 5 18 + 2 22 H Eua Ranunculus
flammula + + 8 8 I + 5 18 + 3 33
H Eua Dif.1 Euphorbia cyparissias
+-1 + 8 8 I + 8 15
H E Car.al Hieracium pilosella + + 7 7 I + 6 11 + 1 4 H E Betonica officinalis + + 7 7 I + 4 8 + 1 7 + 2 7 H E Ajuga reptans + + 7 7 I + 4 18 + 2 22 H Eua Ranunculus acer + + 7 7 I + 1 7 + 5 15 + 2 22 G Cos
m Pteridium
aquilinum +-2 + 6 6 I 1 6 11
H E Campanula patula + + 6 6 I + 6 11 H Eua Sanguisorba minor + + 6 6 I + 1 7 + 3 11 + 2 22 H Ec Salvia verticillata + + 6 6 I + 1 7 + 4 15 + 1 11 H M Mentha pulegium + + 6 6 I + 5 18 + 1 11 H Cos
m Rumex acetosa + + 5 5 I + 2 4 + 1 7 + 2 7
ChH M Helianthemum nummularium
+ + 4 4 I + 4 8
H Eua Stellaria graminaea + + 4 4 I + 2 4 + 2 7 H Cosm Plantago major + + 4 4 I + 3 11 + 1 11 Ch Eua Lysimachia
nummularia + + 3 3 I + 1 4 + 2 22
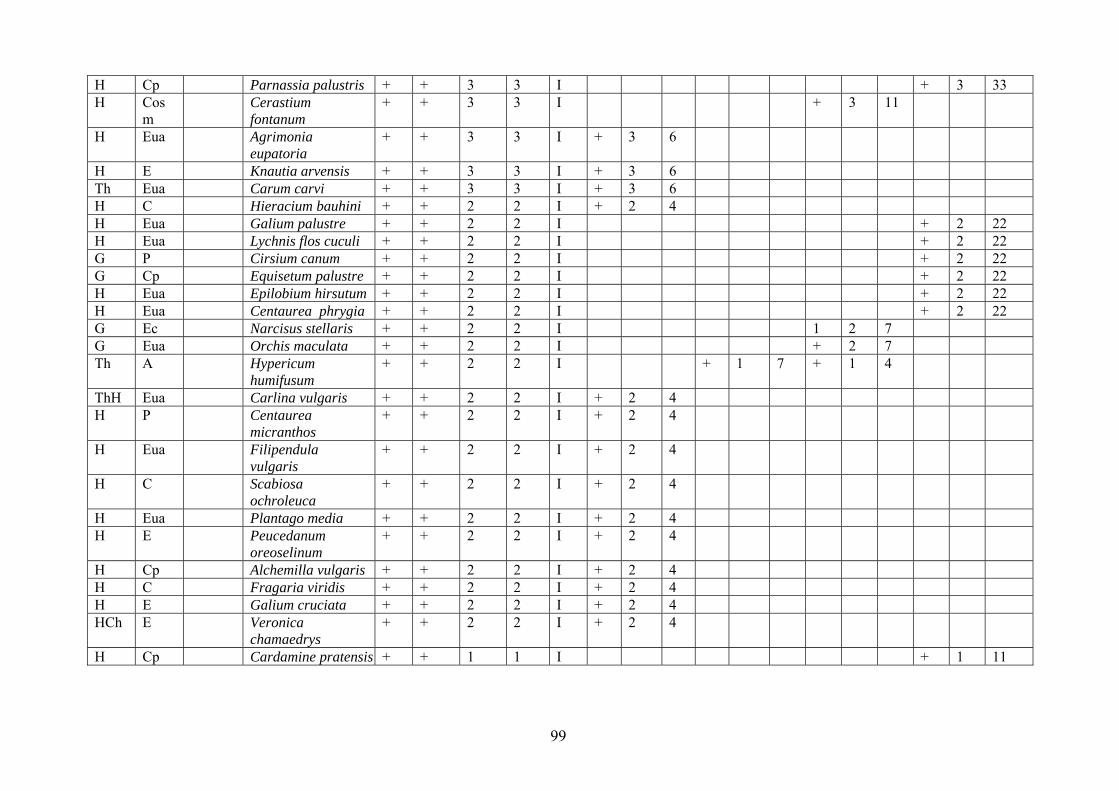
99
H Cp Parnassia palustris + + 3 3 I + 3 33 H Cos
m Cerastium
fontanum + + 3 3 I + 3 11
H Eua Agrimonia eupatoria
+ + 3 3 I + 3 6
H E Knautia arvensis + + 3 3 I + 3 6 Th Eua Carum carvi + + 3 3 I + 3 6 H C Hieracium bauhini + + 2 2 I + 2 4 H Eua Galium palustre + + 2 2 I + 2 22 H Eua Lychnis flos cuculi + + 2 2 I + 2 22 G P Cirsium canum + + 2 2 I + 2 22 G Cp Equisetum palustre + + 2 2 I + 2 22 H Eua Epilobium hirsutum + + 2 2 I + 2 22 H Eua Centaurea phrygia + + 2 2 I + 2 22 G Ec Narcisus stellaris + + 2 2 I 1 2 7 G Eua Orchis maculata + + 2 2 I + 2 7 Th A Hypericum
humifusum + + 2 2 I + 1 7 + 1 4
ThH Eua Carlina vulgaris + + 2 2 I + 2 4 H P Centaurea
micranthos + + 2 2 I + 2 4
H Eua Filipendula vulgaris
+ + 2 2 I + 2 4
H C Scabiosa ochroleuca
+ + 2 2 I + 2 4
H Eua Plantago media + + 2 2 I + 2 4 H E Peucedanum
oreoselinum + + 2 2 I + 2 4
H Cp Alchemilla vulgaris + + 2 2 I + 2 4 H C Fragaria viridis + + 2 2 I + 2 4 H E Galium cruciata + + 2 2 I + 2 4 HCh E Veronica
chamaedrys + + 2 2 I + 2 4
H Cp Cardamine pratensis + + 1 1 I + 1 11
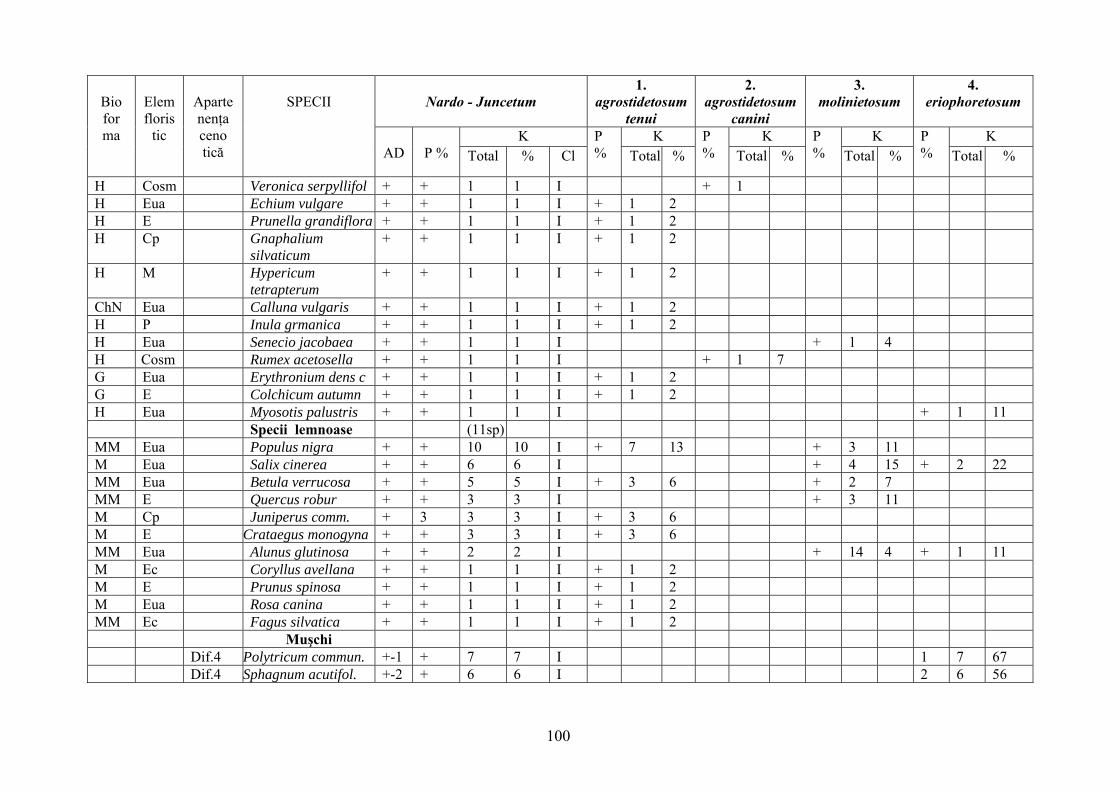
100
Bio for ma
Elem floris
tic
Aparte nenţa ceno tică
SPECII
Nardo - Juncetum
1. agrostidetosum
tenui
2. agrostidetosum
canini
3. molinietosum
4. eriophoretosum
AD
P %
K P %
K P %
K P %
K P %
K Total % Cl Total % Total % Total % Total %
H Cosm Veronica serpyllifol + + 1 1 I + 1 H Eua Echium vulgare + + 1 1 I + 1 2 H E Prunella grandiflora + + 1 1 I + 1 2 H Cp Gnaphalium
silvaticum + + 1 1 I + 1 2
H M Hypericum tetrapterum
+ + 1 1 I + 1 2
ChN Eua Calluna vulgaris + + 1 1 I + 1 2 H P Inula grmanica + + 1 1 I + 1 2 H Eua Senecio jacobaea + + 1 1 I + 1 4 H Cosm Rumex acetosella + + 1 1 I + 1 7 G Eua Erythronium dens c + + 1 1 I + 1 2 G E Colchicum autumn + + 1 1 I + 1 2 H Eua Myosotis palustris + + 1 1 I + 1 11 Specii lemnoase (11sp) MM Eua Populus nigra + + 10 10 I + 7 13 + 3 11 M Eua Salix cinerea + + 6 6 I + 4 15 + 2 22 MM Eua Betula verrucosa + + 5 5 I + 3 6 + 2 7 MM E Quercus robur + + 3 3 I + 3 11 M Cp Juniperus comm. + 3 3 3 I + 3 6 M E Crataegus monogyna + + 3 3 I + 3 6 MM Eua Alunus glutinosa + + 2 2 I + 14 4 + 1 11 M Ec Coryllus avellana + + 1 1 I + 1 2 M E Prunus spinosa + + 1 1 I + 1 2 M Eua Rosa canina + + 1 1 I + 1 2 MM Ec Fagus silvatica + + 1 1 I + 1 2 Muşchi Dif.4 Polytricum commun. +-1 + 7 7 I 1 7 67 Dif.4 Sphagnum acutifol. +-2 + 6 6 I 2 6 56
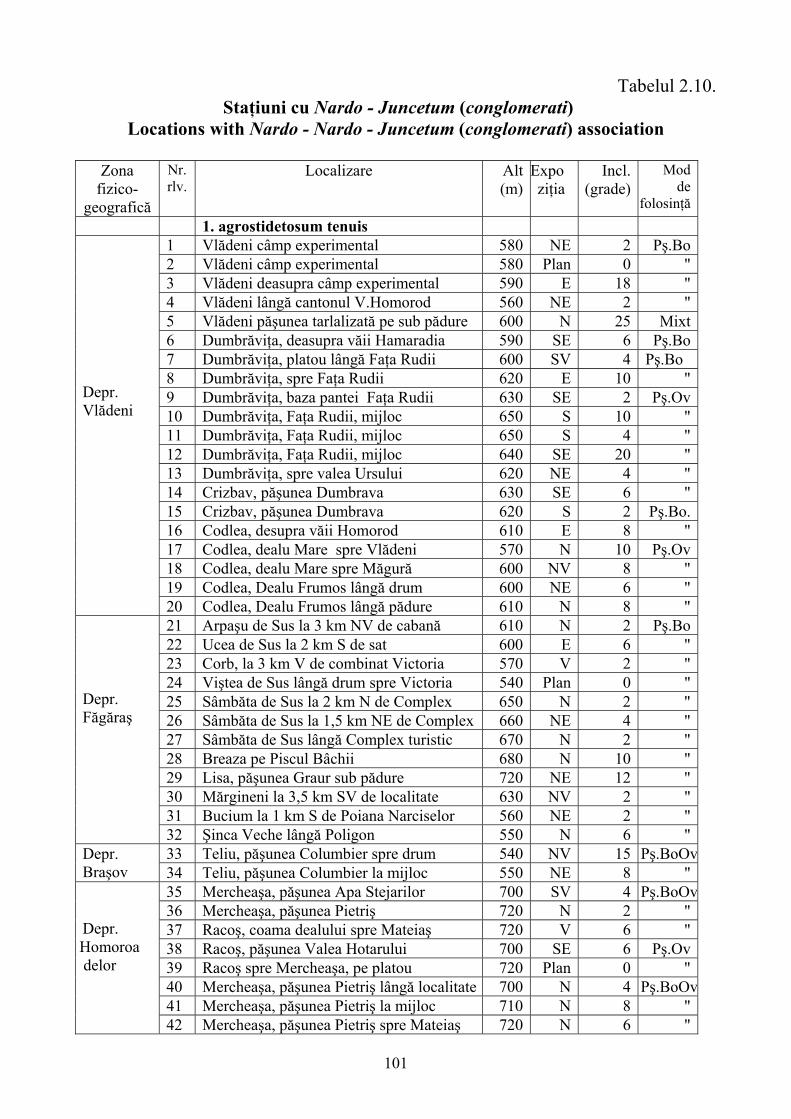
101
Tabelul 2.10. Staţiuni cu Nardo - Juncetum (conglomerati)
Locations with Nardo - Nardo - Juncetum (conglomerati) association
Zona fizico-
geografică
Nr. rlv.
Localizare Alt (m)
Expo ziţia
Incl. (grade)
Mod de
folosinţă
1. agrostidetosum tenuis Depr. Vlădeni
1 Vlădeni câmp experimental 580 NE 2 Pş.Bo 2 Vlădeni câmp experimental 580 Plan 0 "3 Vlădeni deasupra câmp experimental 590 E 18 "4 Vlădeni lângă cantonul V.Homorod 560 NE 2 "5 Vlădeni păşunea tarlalizată pe sub pădure 600 N 25 Mixt6 Dumbrăviţa, deasupra văii Hamaradia 590 SE 6 Pş.Bo7 Dumbrăviţa, platou lângă Faţa Rudii 600 SV 4 Pş.Bo 8 Dumbrăviţa, spre Faţa Rudii 620 E 10 "9 Dumbrăviţa, baza pantei Faţa Rudii 630 SE 2 Pş.Ov10 Dumbrăviţa, Faţa Rudii, mijloc 650 S 10 "11 Dumbrăviţa, Faţa Rudii, mijloc 650 S 4 "12 Dumbrăviţa, Faţa Rudii, mijloc 640 SE 20 "13 Dumbrăviţa, spre valea Ursului 620 NE 4 "14 Crizbav, păşunea Dumbrava 630 SE 6 "15 Crizbav, păşunea Dumbrava 620 S 2 Pş.Bo.16 Codlea, desupra văii Homorod 610 E 8 "17 Codlea, dealu Mare spre Vlădeni 570 N 10 Pş.Ov18 Codlea, dealu Mare spre Măgură 600 NV 8 "19 Codlea, Dealu Frumos lângă drum 600 NE 6 "20 Codlea, Dealu Frumos lângă pădure 610 N 8 "
Depr. Făgăraş
21 Arpaşu de Sus la 3 km NV de cabană 610 N 2 Pş.Bo22 Ucea de Sus la 2 km S de sat 600 E 6 "23 Corb, la 3 km V de combinat Victoria 570 V 2 "24 Viştea de Sus lângă drum spre Victoria 540 Plan 0 "25 Sâmbăta de Sus la 2 km N de Complex 650 N 2 "26 Sâmbăta de Sus la 1,5 km NE de Complex 660 NE 4 "27 Sâmbăta de Sus lângă Complex turistic 670 N 2 "28 Breaza pe Piscul Bâchii 680 N 10 "29 Lisa, păşunea Graur sub pădure 720 NE 12 "30 Mărgineni la 3,5 km SV de localitate 630 NV 2 "31 Bucium la 1 km S de Poiana Narciselor 560 NE 2 "32 Şinca Veche lângă Poligon 550 N 6 "
Depr. Braşov
33 Teliu, păşunea Columbier spre drum 540 NV 15 Pş.BoOv34 Teliu, păşunea Columbier la mijloc 550 NE 8 "
Depr. Homoroa delor
35 Mercheaşa, păşunea Apa Stejarilor 700 SV 4 Pş.BoOv36 Mercheaşa, păşunea Pietriş 720 N 2 "37 Racoş, coama dealului spre Mateiaş 720 V 6 "38 Racoş, păşunea Valea Hotarului 700 SE 6 Pş.Ov39 Racoş spre Mercheaşa, pe platou 720 Plan 0 "40 Mercheaşa, păşunea Pietriş lângă localitate 700 N 4 Pş.BoOv41 Mercheaşa, păşunea Pietriş la mijloc 710 N 8 "42 Mercheaşa, păşunea Pietriş spre Mateiaş 720 N 6 "
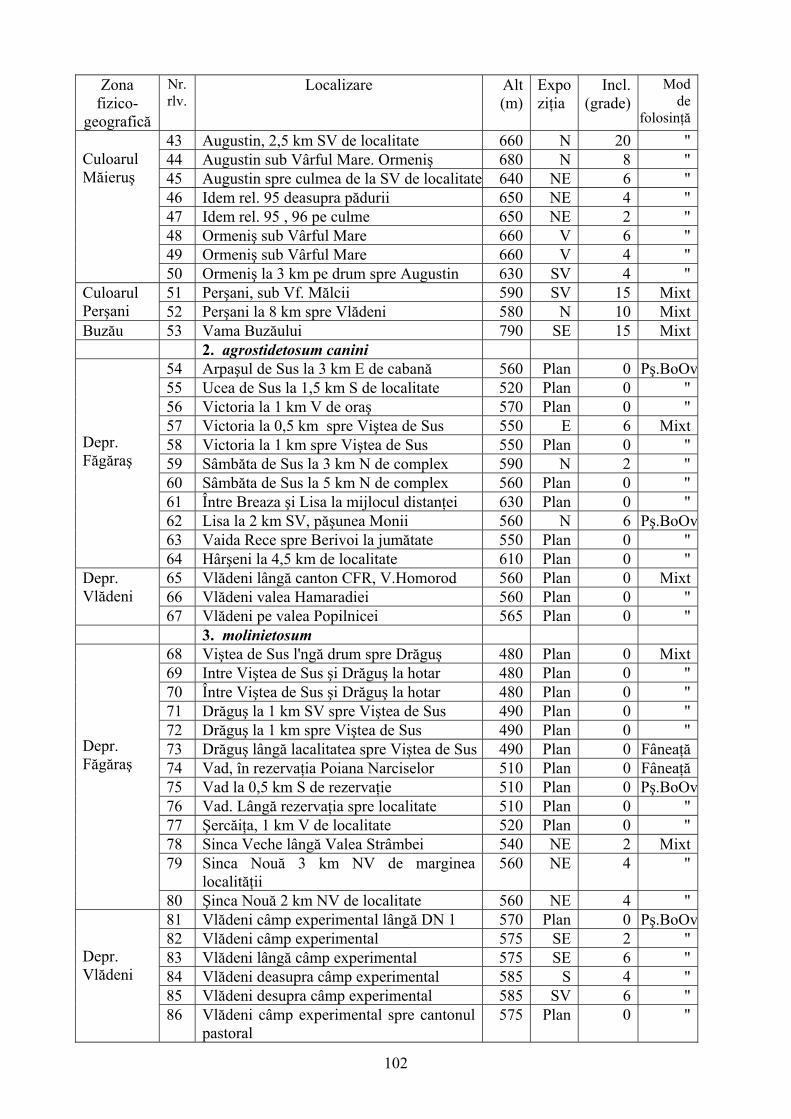
102
Zona fizico-
geografică
Nr. rlv.
Localizare Alt (m)
Expo ziţia
Incl. (grade)
Mod de
folosinţă
Culoarul Măieruş
43 Augustin, 2,5 km SV de localitate 660 N 20 "44 Augustin sub Vârful Mare. Ormeniş 680 N 8 "45 Augustin spre culmea de la SV de localitate 640 NE 6 "46 Idem rel. 95 deasupra pădurii 650 NE 4 "47 Idem rel. 95 , 96 pe culme 650 NE 2 "48 Ormeniş sub Vârful Mare 660 V 6 "49 Ormeniş sub Vârful Mare 660 V 4 "50 Ormeniş la 3 km pe drum spre Augustin 630 SV 4 "
Culoarul Perşani
51 Perşani, sub Vf. Mălcii 590 SV 15 Mixt52 Perşani la 8 km spre Vlădeni 580 N 10 Mixt
Buzău 53 Vama Buzăului 790 SE 15 Mixt 2. agrostidetosum canini Depr. Făgăraş
54 Arpaşul de Sus la 3 km E de cabană 560 Plan 0 Pş.BoOv55 Ucea de Sus la 1,5 km S de localitate 520 Plan 0 "56 Victoria la 1 km V de oraş 570 Plan 0 "57 Victoria la 0,5 km spre Viştea de Sus 550 E 6 Mixt58 Victoria la 1 km spre Viştea de Sus 550 Plan 0 "59 Sâmbăta de Sus la 3 km N de complex 590 N 2 "60 Sâmbăta de Sus la 5 km N de complex 560 Plan 0 "61 Între Breaza şi Lisa la mijlocul distanţei 630 Plan 0 "62 Lisa la 2 km SV, păşunea Monii 560 N 6 Pş.BoOv63 Vaida Rece spre Berivoi la jumătate 550 Plan 0 "64 Hârşeni la 4,5 km de localitate 610 Plan 0 "
Depr. Vlădeni
65 Vlădeni lângă canton CFR, V.Homorod 560 Plan 0 Mixt66 Vlădeni valea Hamaradiei 560 Plan 0 "67 Vlădeni pe valea Popilnicei 565 Plan 0 "
3. molinietosum Depr. Făgăraş
68 Viştea de Sus l'ngă drum spre Drăguş 480 Plan 0 Mixt69 Intre Viştea de Sus şi Drăguş la hotar 480 Plan 0 "70 Între Viştea de Sus şi Drăguş la hotar 480 Plan 0 "71 Drăguş la 1 km SV spre Viştea de Sus 490 Plan 0 "72 Drăguş la 1 km spre Viştea de Sus 490 Plan 0 "73 Drăguş lângă lacalitatea spre Viştea de Sus 490 Plan 0 Fâneaţă74 Vad, în rezervaţia Poiana Narciselor 510 Plan 0 Fâneaţă75 Vad la 0,5 km S de rezervaţie 510 Plan 0 Pş.BoOv76 Vad. Lângă rezervaţia spre localitate 510 Plan 0 "77 Şercăiţa, 1 km V de localitate 520 Plan 0 "78 Sinca Veche lângă Valea Strâmbei 540 NE 2 Mixt79 Sinca Nouă 3 km NV de marginea
localităţii 560 NE 4 "
80 Şinca Nouă 2 km NV de localitate 560 NE 4 " Depr. Vlădeni
81 Vlădeni câmp experimental lângă DN 1 570 Plan 0 Pş.BoOv82 Vlădeni câmp experimental 575 SE 2 "83 Vlădeni lângă câmp experimental 575 SE 6 "84 Vlădeni deasupra câmp experimental 585 S 4 "85 Vlădeni desupra câmp experimental 585 SV 6 "86 Vlădeni câmp experimental spre cantonul
pastoral 575 Plan 0 "
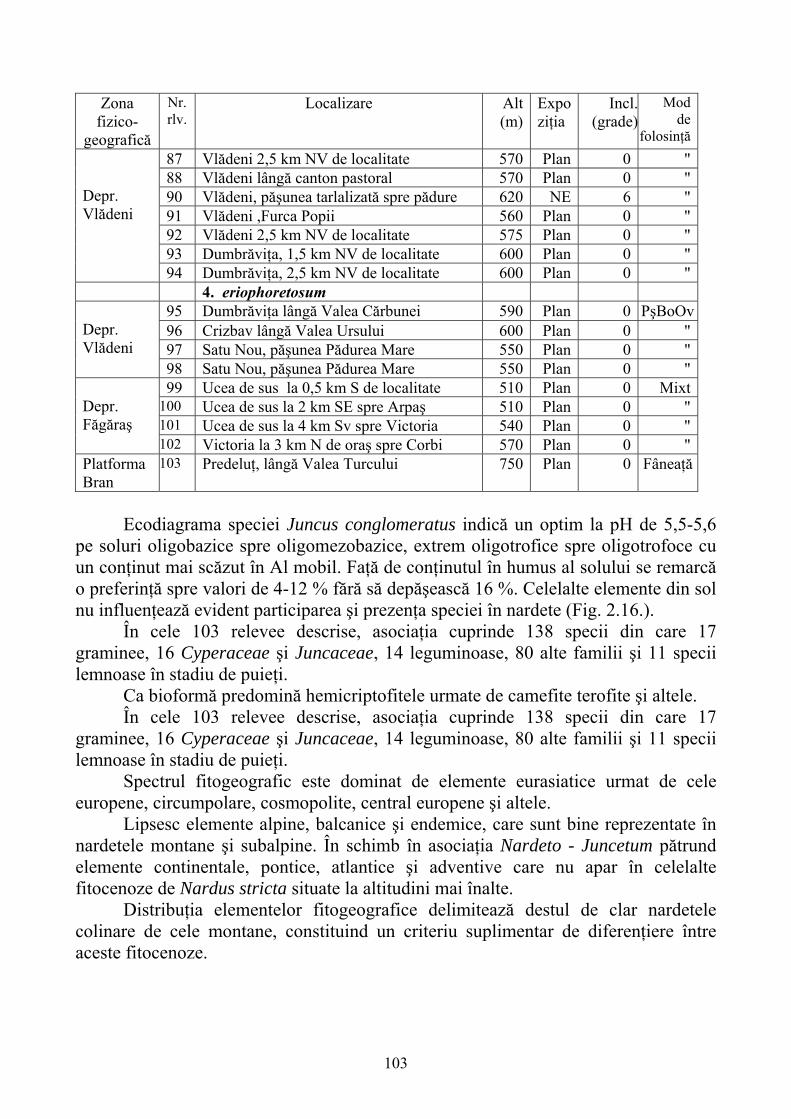
103
Zona
fizico-geografică
Nr. rlv.
Localizare Alt (m)
Expo ziţia
Incl.(grade)
Mod de
folosinţă
Depr. Vlădeni
87 Vlădeni 2,5 km NV de localitate 570 Plan 0 "88 Vlădeni lângă canton pastoral 570 Plan 0 "90 Vlădeni, păşunea tarlalizată spre pădure 620 NE 6 "91 Vlădeni ,Furca Popii 560 Plan 0 "92 Vlădeni 2,5 km NV de localitate 575 Plan 0 "93 Dumbrăviţa, 1,5 km NV de localitate 600 Plan 0 "94 Dumbrăviţa, 2,5 km NV de localitate 600 Plan 0 "
4. eriophoretosum Depr. Vlădeni
95 Dumbrăviţa lângă Valea Cărbunei 590 Plan 0 PşBoOv96 Crizbav lângă Valea Ursului 600 Plan 0 "97 Satu Nou, păşunea Pădurea Mare 550 Plan 0 "98 Satu Nou, păşunea Pădurea Mare 550 Plan 0 "
Depr. Făgăraş
99 Ucea de sus la 0,5 km S de localitate 510 Plan 0 Mixt100 Ucea de sus la 2 km SE spre Arpaş 510 Plan 0 "101 Ucea de sus la 4 km Sv spre Victoria 540 Plan 0 "102 Victoria la 3 km N de oraş spre Corbi 570 Plan 0 "
Platforma Bran
103 Predeluţ, lângă Valea Turcului 750 Plan 0 Fâneaţă
Ecodiagrama speciei Juncus conglomeratus indică un optim la pH de 5,5-5,6
pe soluri oligobazice spre oligomezobazice, extrem oligotrofice spre oligotrofoce cu un conţinut mai scăzut în Al mobil. Faţă de conţinutul în humus al solului se remarcă o preferinţă spre valori de 4-12 % fără să depăşească 16 %. Celelalte elemente din sol nu influenţează evident participarea şi prezenţa speciei în nardete (Fig. 2.16.).
În cele 103 relevee descrise, asociaţia cuprinde 138 specii din care 17 graminee, 16 Cyperaceae şi Juncaceae, 14 leguminoase, 80 alte familii şi 11 specii lemnoase în stadiu de puieţi.
Ca bioformă predomină hemicriptofitele urmate de camefite terofite şi altele. În cele 103 relevee descrise, asociaţia cuprinde 138 specii din care 17
graminee, 16 Cyperaceae şi Juncaceae, 14 leguminoase, 80 alte familii şi 11 specii lemnoase în stadiu de puieţi.
Spectrul fitogeografic este dominat de elemente eurasiatice urmat de cele europene, circumpolare, cosmopolite, central europene şi altele.
Lipsesc elemente alpine, balcanice şi endemice, care sunt bine reprezentate în nardetele montane şi subalpine. În schimb în asociaţia Nardeto - Juncetum pătrund elemente continentale, pontice, atlantice şi adventive care nu apar în celelalte fitocenoze de Nardus stricta situate la altitudini mai înalte.
Distribuţia elementelor fitogeografice delimitează destul de clar nardetele colinare de cele montane, constituind un criteriu suplimentar de diferenţiere între aceste fitocenoze.

104
Fig. 2.16. Dependenţa participării şi prezenţei speciei Juncus conglomeratus din cadrul asociaţiilor de Nardus stricta faţă de caracteristicile solului
Dependence of participation and existence of Juncus conglomeratus species from Nardus stricta associations related to soil proprieties

105
2.4.3.1. Nardo-Juncetum conglomerati agrostidetosum tenuis (Buia 63, Turcu 68)(Syn.Nardetum strictae collinumtypicum, Buia 63; Nardetum strictae collinum agrostidetosum tenuis, Turcu 68; As. de Nardus stricta + Agrostis tenuis, Pauca 60).
A fost identificată în 53 de staţiuni situate la altitudini de 540 - 790 m, pe pante umbrite cu înclinaţie de 2-200, în medie 70.
Subasociaţia este situată în etajul pădurilor de amestec între fag şi gorun la confluenţa cu nardetele montane şi a etajului pădurilor de gorun la altitudini mai loase.
Solurile sunt în marea majoritate brune acide (BO), brune podzolite (BPo), podzolice argiloiluviale (SP, BP, B).
Modul de folosinţă este mai ales prin păşunat cu oile şi mai puţin cu bovinele. În aceste condiţii specia Nardus stricta participă în proporţie de 67 % în
biomasă, fiind cel mai ridicat procent înregistrat în nardetele colinare. Această subasociaţie prezintă cel ,ai avansat proces de degradare a fitocenozelor mezofile situate în zona de deal şi depresionară a judeţului Braşov. Specia Nardus stricta a invadat fitocenozele edificate de Agrostis tenuis, fiind favorizată de acidifierea şi oligotrofismul solului determinate de modul de întreţinere şi folosire a covorului ierbos.
Subasociaţia are în componenţă aproape exclusiv elemente mezofile, fapt ce o diferenţiază ecologic de celelalte fitocenoze edificate de specia Nardus stricta în zona de deal şi depresionară. Astfel, lipsesc speciile mezohigrofile şi higrofile ca: Agrostis canina, Agrostis stolonifera, Molinia coerulea, Poa trivialis, Carex stellulata, Juncus effusus,Gentiana pneumonanthe, Ranunculus flammula, Ajuga reptans, etc. Apar în schimb specii xeromezofite ca: Festuca valesiaca, Dorychnium herbaceum, Euphorbia cyparissias, Agrimonia eupatoria, Plantago media şi altele.
Prezenţa destul de frecventă în subasociaţie a speciilor Dechampsia caespitosa şi Juncus conglomeratus, care după cum se ştie sunt specii care vieţuiesc în condiţii de umiditate mai ridicată a solului, creează unele confuzii privind caracterul pronunţat mezofil al acestei fitocenoze. În acest caz, vitalitatea celor două specii menţionate are un rol determinant în delimitarea fitocenozelor mezofile de cele mezohigrofile şi higrofile. Astfel, în subasociaţia Nardo-Juncetum agrostidetosum tenuis, speciile Dechampsia caespitosa şi Juncus conglomeratus se prezintă ca indivizi izolaţi subdezvoltaţi, nu formează tufe caracteristice fiind la limita vieţuirii lor în nardete. Celelalte fitocenoze edificate de Nardus stricta situate în condiţii cu umiditate mai pronunţată, speciile de Dechampsia caespitosa şi Juncus conglomeratus sunt cu mult mai viguroase şi formează tufe mari, dese, având o participare însemnată în covorul ierbos.
Ecodiagrama speciei Agrostis tenuis confirmă şi mai bine condiţiile ecologice diferenţiate în care se întâlneşte subasociaţia. În nardete specia Agrostis tenuis se întâlneşte la un pH cuprins între 4,3- 5,8 realizează optimum ecologic la un pH acid (5,7-5,8) pe soluri oligomezobazice spre mezobazice, oligomezotrofice, medii humifer (4-5 % humus) cu un conţinut mai ridicat în P2O5 şi variabil în K2O. Faţă de Al mobil se remarcă un optim ecologic la limita unde începe potenţialul toxic (6-10 %).

106
Analizând dependenţa speciei Agrostis tenuis faţă de caracteristicile solului în pajiştile din zona Caraşova a Carstului Bănăţean, LAUER C., 1974, stabileşte optimul ecologic la un pH de 4,5-5,1. O întâlneşte şi la un pH slab alcalin 6,4-6,9 pe soluri mezobazice până la eubazice humifere (V % = 91 - 95) slab aprovizionate în P2O5 şi K2O schimbabil.
În Depresiunea Liteni - Suceava, CIUBOTARU C., 1974 descrie specia Agrostis tenuis într-o repartizare aproape constantă la un pH cuprins între 4,5-7,5 pe soluri slab aprovizionate în P2O5 şi K2O, cu un conţinut mai scăzut în humus (3,2 – 10 %).
Aceste rezultate pun în discuţie generalizarea valorii indicatoare a speciei Agrostis tenuis pentru caracteristicile solului. Dacă ne referim la aciditatea solului în Carstul Bănăţean valoarea indicatoare este destul de bine conturată iar în Derpresiunea Liteni specia Agrostis tenuis este aproape lipsită de această calitate. Facem această comparaţie întrucât autorii celor două lucrări au studiat comportarea speciei Agrostis tenuis independent de fitocenozele în care s-a întâlnit, respectiv în întreaga zonă luată în studiu din care lipsesc însă nardetele.
În lucrarea de faţă s-a studiat dependenţa speciei Agrostis tenuis faţă de caracteristicile solului într-un cadru limitat al fitocenozelor edificate de Nardus stricta specie care după cum se ştie are o influenţă determinantă şi asupra solului. În aceste condiţii specifice nardetelor, prezenţa şi participarea mai mare a speciei Agrostis tenuis în covorul ierbos ne dau suficiente elemente de diferenţiere a unei subasociaţii faţă de altele. Este ceea ce şi-a propus dealtfel acest studiu în direcţia stabilirii pe baze ecologico-floristice a fitocenozelor edificate de specia Nardus stricta. Din punct de vedere al evoluţiei, Nardo-Juncetum agrostidetosum tenuis, reprezintă stadiul cel mai avansat de degradare a fitocenozelor de Agrostis tenuis care s-a instalat după defrişarea pădurilor de fag şi gorun. Specia Nardus stricta se poate instala direct în fitocenozele edificate de Calamagrostis arundinacea care reprezintă vegetaţia ierboasă după defrişare. Prezenţa speciei Calamagrostis arundinacea în subasociaţie atestă această posibilitate directă de instalare a nardetelor în condiţiile păşunatului cu oile. Producţia de masă verde se estimează la cca. 5,1 t/ha, fiind de calitate slabă. Valoarea furajeră este de 33, fiind de asemernea scăzută. 2.4.3.2. Nardo - juncetum conglomerati agrostidetosum canini, Turcu 68.
(Syn.Nardetum strictae collinum agrostidetosum canini Turcu 68;) A fost întâlnită în 14 staţiuni situate între 520 - 630 m altitudine, pe terenuri
plane sau cu o uşoară înclinaţie de pânî la 40 din depresiunile Făgăraş, Vlădeni şi Homoroadelor.
Vegetaţia primară este reprezentată de pădurile de gorun. Solurile sunt podzolice argiloiluviale, superficial-profund oligomezobazice
(Spo).

107
O caracteristică a acestor soluri este drenajul mai bun datorită unui conţinut mai ridicat în fragmente de roci care adeseori se întâlnesc la suprafaţă sub formă de pietriş rulat.
Modul de folosinţă al acestor pajişti este prin păşunat cu bovine din care predomină bubalinele la care se adaugă şi ovinele. Pe alocuri se folosesc mixt, în apropierea localităţilor.
În această subasociaţie specia Nardus stricta participă în proporţie de 47 % din biomasă urmată de Agrostis tenuis cu 22 %, Juncus conglomeratus care cu cei 5 % realizează cea mai ridicată participare medie în nardete şi alte specii.
Prezenţa acestei fitocenoze în ţara noastră a fost pusă în evidenţă în ultima vreme, datorită faptului că o parte din exemplarele de Agrostis caninai au fost atribuite speciei Agrostis stolonifera (alba), după cum remarcă TURCU, 1968.
Astfel, se face că în Depresiunea Vlădeni BUIA, 1963, descrie subasociaţia Nardetum strictae collinum agrostidetosum albae, care ulterior nu a fost reîntâlnită deşi s-au făcut pe parcursul anilor mai multe investigaţii. Este posibil să existe o confuzie mai ales că alături de BUIA, 1963 şi alţi autori SAFTA, 1943; RESMERIŢĂ,1971, nu menţionează existenţa speciei Agrostis canina în nardetele din ţara noastră, indicând în schimb Agrostis stolonifera ca frecventă pe terenuri cu exces de umiditate. Cu toate acestea nu se poate trage o concluzie definitivă asupra unei eventuale confuzii întrucât în nardetele din apropierea luncilor râurilor se întâlneşte sporadic specia Agrostis stolonifera în proporţie de până la 12 % participare în biomasă. Destul de recent mai mulţi autori indică prezenţa speciei Agrostis canina în nardete, adeseori alături de Agrostis stolonifera (SERBĂNESCU,1964; TURCU, 1968; MOTCA, 1972; PAUN, 1973; ANGHEL, 1977).
Ecodiagrama speciei Agrostis canina indică optimum de participare şi prezenţă în nardete pe soluri cu un pH de 4,9-5,2 oligomezobazice, extrem oligotrofice, conţinut în humus de 9-12 % foarte sărace în K2O schimbabil şi conţinut în Al mobil la limita inferioară a toxicităţii. Condiţiile staţionale în care întâlnim această fitocenoză au un caracter mai pronunţat mezohigrofit decât subasociaţia precedentă.
Din subasociaţia Nardo - Juncetum agrostidetosum canini lipsesc speciile: Molinia coerulea, Carex stellulata, Gentiana pneumonanthe, Ranunculus flammula, Ajuga reptans, Lysimachia nummularia şi alte specii care indică un grad mai pronunţat de umiditate a solului.
Producţia de masă verde este estimată la cca. 4,7 t/ha fiind de o calitate slabă. Indicele de valoare furajeră este de 32.
2.4.3.3. Nardo – Juncetum conglomerati molinietosum Gergely 56. (Syn. Nardo-Molinietum, Gergely 56 )
A fost identificată în 27 staţiuni situate între 480-620 m altitudine pe terenuri
plane până la 60 înclinaţie. Subasociaţia este rezultatul invadării fitocenozelor de Molinia coerulea de
către specia Nardus stricta.

108
Dacă se consideră molinietele ca fitocenoze ierboase relicte RESMERIŢĂ I. 1969, atunci problema vegetaţiei primare este destul de clară. Dar evoluţia molinietelor este destul de greu de stabilit în condiţiile influenţei factorilor atropici. Pe de o parte se constată restrângerea arealului fitocenozelor de Molinia coerulea prin invadarea lor de către specia Narduss stricta cât şi a speciilor lemnoase Salix cinerea, Alunus glutinosa, Betula Verrucosa, Populus nigra şi în cele din urmă Querqus robur care se întâlneşte sub formă de indivizi izolaţi, uneori bine dezvoltaţi. În alte condiţii Molinia coerulea se întâlneşte în luminişurile de stejar de la Dumbrava Vadului. Este dificil de constatat dacă stejarul s-a instalat în moliniete în urma unui proces de drenare a terenurilor mlăştinoase. Cercetări de lungă durată în staţionar sunt necesare pentru a lămuri această evoluţie.
Solurile mai reprezentative sunt argiloiluviale pseudogleice şi soluri podzolice argiloiluviale pseudogleizate (IP).
În această subasociaţie Nardus stricta participă în proporţie de 53 % din biomasă urmată de Molinia coerulea cu 8 % şi alte specii cu participare mai mică.
Ca specii diferenţiale ecologice pe lângă Molinia coerulea se remarcă şi Gentiana pneumonanthe şi Achillea patarmica, specii aproape integral întâlnite numai în această fitocenoză.
Ecodiagrama speciei Molinia coerulea indică optimum de participare şi prezenţă relativă în nardetele răspândite pe soluri puternic acide (pH 4,9-5,2), oligomezobazice, oligotrofice, cu un conţinut în humus de 4-8 %, 20-30 mg/a00 g sol Al mobil, foarte sărace în K2O schimbabil (Fig. 2.17).
Analizând comportarea speciei Molinia coerulea din diferite fitocenoze, în afară de nardete CIUBOTARU C. 1974, stabileşte optimum de participare relativă la pH 5,0-5,5 şi cel de prezenţă la pH 6,0-6,5 fiind întâlnită de la pH 4,5 până la pH 7,5. Faţă de humus P2O5 şi K2O acelaşi autor stabileşte exigenţele ecologice ale acestei specii din Depresiunea Liteni-Suceava, în limite apropiate cu cele din judeţul Braşov.
Şi în acest caz se constată unele deosebiri mai ales în ceea ce priveşte aciditatea solului faţă de care Molinia coerulea are exigenţe mai largi.
Ecodiagrama speciei Gentiana pneumonanthe indică soluri cu un pH cuprins între 4,9-5,2, un conţinut în humus de 4-12 % şi valori ale Al mobil de 6-20 mg/100 g sol (Fig.2.18.).
Aceste valori restrânse ale diferitelor constituenţi ai solului fac din această specie una din cele mai valoroase plante indicatoare ale fitocenozelor edificate de Nardus stricta.
Comportarea speciei Gentiana pneumonanthe este destul de apropiată de cea a speciei Molinia coerulea, fiind întâlnite adeseori împreună. Prezenţa speciei Gentiana pneumonanthe în nardete chiar în absenţa speciei Molinia coerulea ne indică provenienţa acestei fitocenoze din moliniete.
Producţia de masă verde este estimată la 4,8 t/ha având o calitate slabă. Valoarea furajeră este de 25 fiind de asemenea scăzută.
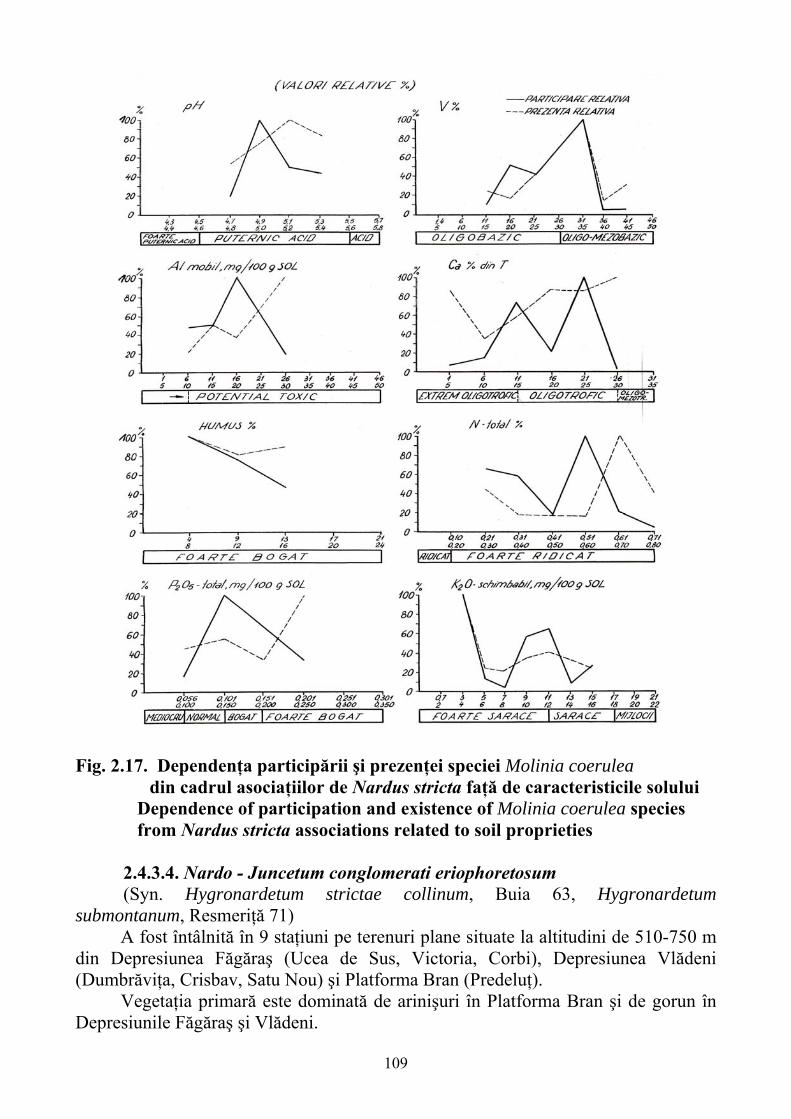
109
Fig. 2.17. Dependenţa participării şi prezenţei speciei Molinia coerulea
din cadrul asociaţiilor de Nardus stricta faţă de caracteristicile solului Dependence of participation and existence of Molinia coerulea species from Nardus stricta associations related to soil proprieties 2.4.3.4. Nardo - Juncetum conglomerati eriophoretosum (Syn. Hygronardetum strictae collinum, Buia 63, Hygronardetum
submontanum, Resmeriţă 71) A fost întâlnită în 9 staţiuni pe terenuri plane situate la altitudini de 510-750 m din Depresiunea Făgăraş (Ucea de Sus, Victoria, Corbi), Depresiunea Vlădeni (Dumbrăviţa, Crisbav, Satu Nou) şi Platforma Bran (Predeluţ). Vegetaţia primară este dominată de arinişuri în Platforma Bran şi de gorun în Depresiunile Făgăraş şi Vlădeni.

110
Fig. 2.18. Dependenţa participării şi prezenţei speciei Gentiana pneumonanthe din cadrul asociaţiilor de Nardus stricta faţă de caracteristicile solului
Dependence of participation and existence of Gentiana pneumonanth species from Nardus stricta associations related to soil proprieties

111
Solurile sunt mai ales argiloiluviale brune podzolite (SVr) şi soluri aluviale frecvent gleizate (SA). În aceste condiţii pe marginea terenurilor mlăştinoase cu exces permanent de umiditate se instalează şi specia Nardus stricta care în unele situaţii reuşeşte să domine în covorul ierbos, edificând higronardetele. Excesul aproape permanent de umiditate face ca această fitocenoză să fie folosită sporadic prin păşunat iar în perioade cu precipitaţii mai abundente animalele să ocolească aceste terenuri. În subasociaţie specia Nardus stricta participă în proporţie de 50 % urmată de Deschampsia caespitosa 6 % dintre graminee şi 18 % ciperacee şi juncacee care au cea mai ridicată participare în biomasa nardetelor din judeţul Braşov. Ca specii diferenţiale ecologice se enumără: Eriophorum latifolium, Juncus articulatus, Pycreus flavescens şi alte plante higrofile. La acestea adăugăm participarea cea mai mare în nardete a speciei Juncus effusus 9 % cât şi prezenţa speciilor din genul Sphagnum în 4 din cele 9 releveuri descrise.
Toate aceste specii dau o notă particulară fitocenozelor de Nardus stricta, care sunt prin excelenţă indicatoare de exces de umiditate a solului la limita terenurilor cu vegetaţie specifică de mlaştină. Producţia de masă verde se estimează la 4,2 t/ha, fiind de o calitate foarte slabă. Valoarea furajeră este de 24 fiind din cele mai scăzute înregistrate pe nardete. În concluzie fitocenozele edificate de specia Nardus stricta în judeţul Braşov prezintă o diversitate accentuată, fapt ce a permis o caracterizare unitară floristico-ecologică, cu unele aspecte economice. Datele care au servit la conturarea fitocenozelor de Nardus stricta se regăsesc în continuare la caracterizarea tipurilor şi bonitarea nardetelor. S-a procedat la acest sistem de reprezentare din motive de spaţiu cu evitarea unor repetări.
2.4.4. Indicii sinecologici ai nardetelor
În vederea unei caracterizări sinecologice a asociaţiilor şi subasociaţiilor edificate de specia Nardus stricta s-au calculat indicii de umiditate, temperatură, lumină, reacţie şi azot, utilizându-se metoda preconizată de ELLENBERG H. 1952, expusă şi modificată de CSŰROS St. şi colab. 1966, prezentată mai amplu de LAUER C. 1974, completată de KOVACS At 1979.Datele obţinute permit compararea trăsăturilor ecologice a fitocenozelor de Nardus stricta şi aduc elemente noi de referinţă în caracterizarea geobotanică a nardetelor, faţă de alte fitocenoze (Tab.2.11).
Faţă de factorul lumină (L) toate fitocenozele sunt indicatoare de lumină care suportă slab umbrirea, fiind apropiate de cele care preferă lumină plină. Diferenţe destul de mici sunt înregistrate şi la factorul reacţia solului (pH-ul) (R) unde fitocenozele edificate de Nardus stricta preferă soluri foarte acide spre cele acide, fapt reieşit şi din ecodiagrame.
De asemenea, faţă de cantitatea de azot mineral din sol (N) la nardete se înregistrează fluctuaţii destul de mici cuprinse între soluri sărace în azot mineral spre cele moderate ca şi conţinut în acest element.
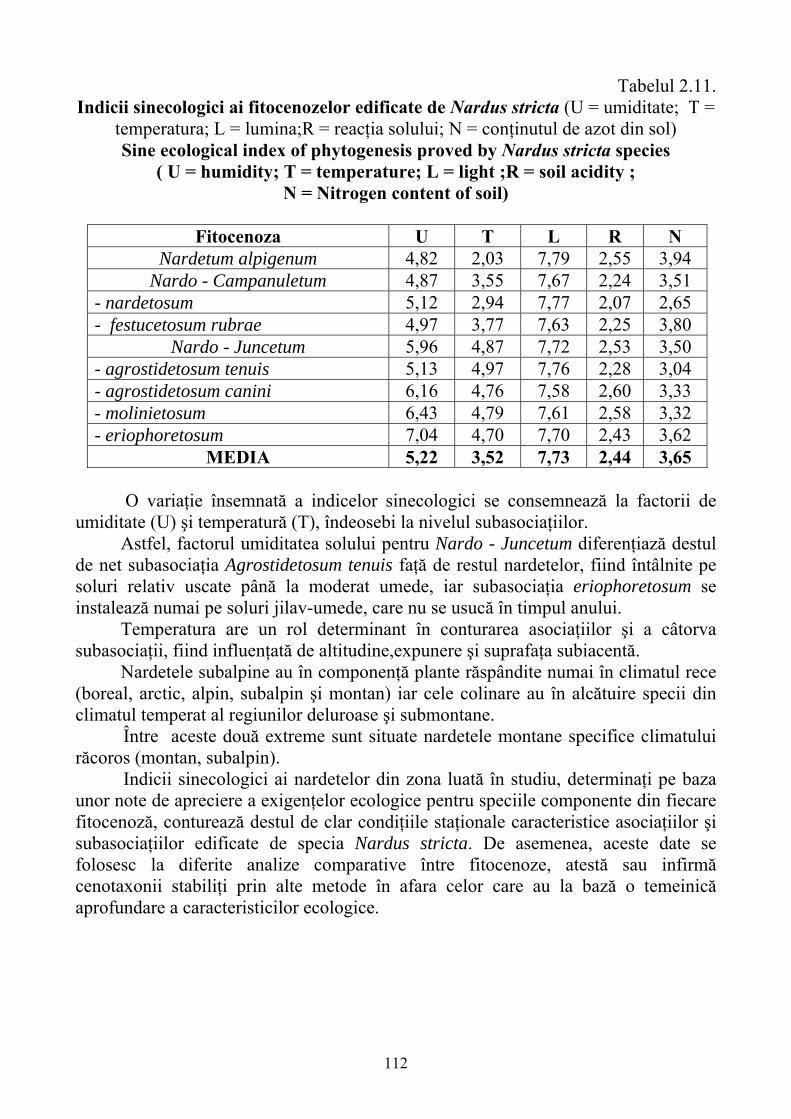
112
Tabelul 2.11. Indicii sinecologici ai fitocenozelor edificate de Nardus stricta (U = umiditate; T =
temperatura; L = lumina;R = reacţia solului; N = conţinutul de azot din sol) Sine ecological index of phytogenesis proved by Nardus stricta species
( U = humidity; T = temperature; L = light ;R = soil acidity ; N = Nitrogen content of soil)
Fitocenoza U T L R N
Nardetum alpigenum 4,82 2,03 7,79 2,55 3,94 Nardo - Campanuletum 4,87 3,55 7,67 2,24 3,51
- nardetosum 5,12 2,94 7,77 2,07 2,65 - festucetosum rubrae 4,97 3,77 7,63 2,25 3,80
Nardo - Juncetum 5,96 4,87 7,72 2,53 3,50 - agrostidetosum tenuis 5,13 4,97 7,76 2,28 3,04 - agrostidetosum canini 6,16 4,76 7,58 2,60 3,33 - molinietosum 6,43 4,79 7,61 2,58 3,32 - eriophoretosum 7,04 4,70 7,70 2,43 3,62
MEDIA 5,22 3,52 7,73 2,44 3,65 O variaţie însemnată a indicelor sinecologici se consemnează la factorii de umiditate (U) şi temperatură (T), îndeosebi la nivelul subasociaţiilor. Astfel, factorul umiditatea solului pentru Nardo - Juncetum diferenţiază destul de net subasociaţia Agrostidetosum tenuis faţă de restul nardetelor, fiind întâlnite pe soluri relativ uscate până la moderat umede, iar subasociaţia eriophoretosum se instalează numai pe soluri jilav-umede, care nu se usucă în timpul anului. Temperatura are un rol determinant în conturarea asociaţiilor şi a câtorva subasociaţii, fiind influenţată de altitudine,expunere şi suprafaţa subiacentă. Nardetele subalpine au în componenţă plante răspândite numai în climatul rece (boreal, arctic, alpin, subalpin şi montan) iar cele colinare au în alcătuire specii din climatul temperat al regiunilor deluroase şi submontane.
Între aceste două extreme sunt situate nardetele montane specifice climatului răcoros (montan, subalpin).
Indicii sinecologici ai nardetelor din zona luată în studiu, determinaţi pe baza unor note de apreciere a exigenţelor ecologice pentru speciile componente din fiecare fitocenoză, conturează destul de clar condiţiile staţionale caracteristice asociaţiilor şi subasociaţiilor edificate de specia Nardus stricta. De asemenea, aceste date se folosesc la diferite analize comparative între fitocenoze, atestă sau infirmă cenotaxonii stabiliţi prin alte metode în afara celor care au la bază o temeinică aprofundare a caracteristicilor ecologice.

113
2.5. DESCRIEREA TIPURILOR DE NARDETE Diferenţierea şi descrierea tipurilor de nardete s-a făcut după anumite criterii în
care compoziţia floristică, factorii staţionali şi posibilităţile de îmbunătăţire a acestor pajişti degradate au avut un rol preponderent în elaborarea unei clasificări unitare.
Clasificarea tipologică aduce multe criterii de referinţă pentru practică şi include posibilităţile de îmbunătăţire şi valorificare a producţiei în funcţie de nivelul cercetărilor şi experienţei extinderii în producţie realizate până acum la noi în ţară şi străinătate. Însăşi termenul de nardete indică covor ierbos degradat, producţie şi calitate furajeră necorespunzătoare, soluri acide, oligobazice şi oligomezobazice, conţinut scăzut în elemente fertilizante, exces temporar de umiditate o perioadă mai scurtă sau mai lungă din an, păşunat permanent cu intensitate moderată, etc., caracteristici prezentate în studiul ecologic şi geobotanic al acestor fitocenoze.
Aceste caracteristici conturate în numeroase lucrări din ţara noastră şi străinătate, fac ca nardetele să fie considerate tipuri de pajişti degradate calitativ şi productiv, ce reprezintă un studiu avansat de invadare de către specia Nardus stricta a unor fitocenoze mai valoroase care ar fi continuat să supravieţuiască în cazul unui sistem de întreţinere şi folosire corespunzător.
Studiul geobotanic caută să fundamenteze pe baze ecologice instalarea şi expansiunea speciei Nardus stricta în diferite staţiuni şi fitocenoze considerate normale în cazul menţinerii unui echilibru favorabil determinat de acţiunea omului şi al animalelor sale.
Studiul tipologic în cazul nardetelor are ca scop principal descrierea condiţiilor care exprimă stadiile degradării legate de vegetaţie, sol, influenţa animalelor,etc., care să stea la baza acţiunilor de corectare a factorilor de degradare şi reinstalării tipurilor de pajişti normale premergătoare sau creării unor tipuri artificiale mai productive şi economice. În acest fel clasificarea tipologică serveşte într-o măsură mai mare practica imediată.
2.5.1. Nardete subalpine situate pe pante foarte puternic înclinate sau abrupte. Reprezintă cel mai răspândit tip de nardete din etajul alpin caracteristic
abrupturilor munţilor Bucegi şi Făgăraş ce aparţin judeţului Braşov, situat pe pante mari, greu accesibile. Fitocenoza este reprezentată de Nardetum alpigenum care preferă pantele puternic însorite.
Condiţiile ecologice au determinat conturarea a două subtipuri şi anume: a. pe soluri relativ profunde; b. pe soluri scheletice cu roci de suprafaţă. Valoarea pastorală a pajiştilor de Nardus stricta subalpine, situate pe pante
puternice şi abrupte este scăzută fiind de 10,6. Variantele de îmbunătăţire pe solurile relativ profunde se reduc la fertilizarea
cu îngrăşăminte organice realizată prin târlire şi în unele cazuri aplicarea îngrăşămintelor chimice cu ajutorul aviaţiei. Aceste măsuri au o eficienţă economică scăzută.

114
Pe solurile scheletice cu roci la suprafaţă nu se recomandă luarea unor măsuri de îmbunătăţire prin fertilizare datorită pierderilor şi a lipsei de eficienţă economică.
Valorificarea producţiei se face în continuare prin păşunat extensiv cu oile. 2.5.2. Nardete subalpine situate pe terenuri plane sau uşor până la mijlociu
înclinate. Sunt răspândite pe Platourile Munţilor Bucegi, Făgăraş şi Bârsei (Piatra Mare)
cât şi pe fundul văilor glaciare ale Făgăraşului şi Bucegilor. Acest tip de nardete are un areal mai restrâns în judeţul Braşov fiind în schimb foarte răspândit în judeţele învecinate, Prahova, Dâmboviţa , Argeş unde munţii Bucegi şi Făgăraş prezintă platouri şi pante uşor înclinate pe întinderi vaste.
Covorul ierbos este de asemenea reprezentat de asociaţia Nardetum alpigenum. Condiţiile edafice, microclimatul şi unele aspecte ale vegetaţiei lemnoase
arbustive au determinat conturarea a 4 subtipuri şi anume: a. pe soluri profunde; b. pe soluri scheletice cu roci la suprafaţă; c. cu muşuroaie înţelenite, d. cu vegetaţie arbustivă în extindere. Între aceste subtipuri se întâlnesc uneori combinaţii cum ar fi ed exemplu
nardetele pe soluri profunde cu muşuroaie înţelenite (a+c) sau nardete pe soluri scheletice cu vegetaţie arbustivă (b+d) sau altele.
Subtipul de nardete subalpine pe soluri scheletice cu roci la suprafaţă se întâlneşte îndeosebi pe fundul căldărilor glaciare sau în apropierea zonelor stâncoase de unde prin dezagregarea rocilor şi rostogolirea lor împânzesc cu materiale scheletice solurile din jurul lor.
Pe platouri mai întinse cum sunt cele din Bucegi întâlnim subtipul de nardete pe soluri profunde. De asemenea, în aceste condiţii apar şi muşuroaiele înţelenite care conturează un subtip de nardete distinct.
În toate aceste condiţii se înfiripă pe suprafeţe destul de întinse o vegetaţie arbustivă reprezentată prin Juniperus communis alpestris şi Pinus mugo.
Valoarea pastorală a pajiştilor de Nardus stricta subalpine situate pe terenuri plane sau uşor până la moderat înclinate,este destul de scăzut fiind de 12,2.
Posibilităţile de îmbunătăţire a acestui tip de nardete sunt mai mari, date fiind condiţiile staţionale mai favorabile introducerii mijloacelor mecanizate
2.5.3. Nardete cu diferite specii de Vaccinum. Sunt bine reprezentate în etajul molidului şi pe soluri puternic acide în munţii
Bucegi, Făgăraş, Bârsa, Ciucaş şi Piatra Craiului. Instalarea masivă a speciilor de Vaccinum myrtillus şi Vaccinum vitis idaea ca
urmare a unui păşunat sporadic pe alocuri abandonat, fac ca aceste nardete să prezinte câteva particularităţi distincte faţă de celelalte tipuri de pajişti cu Nardus stricta din zona montană.
În primul rând acest tip de nardete este situat cel mai adesea pe terenuri cu pante foarte puternic înclinate până la foarte abrupte.

115
În aceste condiţii apar adeseori specii lemnoase cum sunt Juniperus communis, şi Picea abiens care stabilesc în cele din urmă vegetaţia primară de pădure din vecinătate.
Covorul ierbos aparţine asociaţiei Nardo-Camapanulatum nardetosum. În cadrul pajiştilor de Nardus stricta cu diferite specii de Vaccinum se disting
subtipurile: a. pe soluri profunde; b. pe soluri scheletice; d.cu vegetaţie lemnoasă precum şi combinaţiile a+d şi b+d. Valoarea pastorală este de 3,4 fiind cea mai scăzută dintre nardetele luate în
studiu. 2.5.4. Nardete montane cu Festuca rubra şi Agrostis capillaris situate pe
pante foarte puternic înclinate sau abrupte. Sunt de asemenea răspândite în toată zona montană a judeţului Braşov. Vegetaţia ierboasă aparţine subasociaţiei Nardo - Camapanuletum
festucetosum rubrae şi parţial la nardetosum. Nardete cu Festuca rubra şi Agrostis tenuis situate pe pante foarte puternic
înclinate sau abrupte au în componenţă 3 subtipuri si anume: a. pe soluri profunde; b. pe soluri scheletice; d. cu vegetaţie lemnoasă şi două combinaţii dintre acestea (a+d şi b+d). Valoarea pastorală a acestui tip este destul de scăzut fiind de 22,2. Variantele de îmbunătăţire sunt mai restrânse datorită condiţiilor de relief
accidentat şi accesibilitatea redusă. 2.5.5. Nardete cu Festuca rubra şi Agrostis capillaris situate pe pante foarte
slab până la mijlociu înclinate. Sunt răspândite în toată zona montană a judeţului Braşov la etajele pădurilor de
molid, fag şi a amestecurilor dintre acestea. Arealul destul de larg în care se întâlneşte acest tip de pajişte a pus unele
probleme de caracterizare şi conturare a lui. Posibilităţile unitare de îmbunătăţire a acestor pajişti cât şi rezultatul final destul de apropiat al acestor acţiuni obţinute prin numeroase cercetări au uşurat încadrarea lor într-un tip reprezentativ.
Vegetaţia ierboasă aparţine asociaţiei Nardo-Campanuletum cu subasociaţia festucetosum rubrae.
Nardete montane cu Festuca rubra şi Agrostis tenuis situate pe pante slab până la mijlociu înclinate, au în componenţă 4 subtipuri şi anume:
a. pe soluri profunde; b. pe soluri scheletice c. cu muşuroaie şi înţelenite; d. cu vegetaţie lemnoasă şi unele combinaţii dintre acestea. Valoarea pastorală a acestor pajişti ajunge la 23,2 fiind ceva mai ridicată decât
a tipurilor descrise până acum.

116
La fel şi variantele de îmbunătăţire sunt mai numeroase date fiind posibilităţile de mecanizare a lucrărilor.
2.5.6. Nardete colinare, cu diferite specii mezofile şi xeromezofile situate pe
pante mijlocii până la foarte puternic înclinate. Au o răspândire destul de întinsă în zonele de confluenţă a dealurilor cu munţii
şi în zona de deal şi depresionară pe evantaiele din apropierea văilor. Acest tip de nardete a fost descris pentru prima oară în Depresiunea Făgăraş de
către MOTCĂ, 1972, pe care îl numeşte Nardus stricta de dealuri premontane şi evantaie proluviale cu posibilităţi restrânse de mecanizare.
Vegetaţia ierboasă aparţine integral subasociaţiei Nardo-Juncetum agrostidetosum tenuis.
În componenţa acestui tip de nardete se remarcă trei subtipuri şi anume: a. pe soluri profunde; b. pe soluri scheletice d. cu vegetaţie lemnoasă şi două combinaţii dintre acestea a+d şi b+d.
Valoarea furajeră a acestor pajişti este destul de heterogenă, fiind în general scăzută ajungând în medie la 14,2. Variantele de îmbunătăţire sunt mai reduse datorită reliefului cu pante parţial mecanizabile şi al existenţei la mică adâncime în sol al unor materiale parentale dure.
2.5.7. Nardete din depresiuni, cu diferite specii mezohigrofile şi higrofile situate pe terenuri plane sau uşor înclinate.
Acest tip de nardete are cea mai largă răspândire în Depresiunile Făgăraş şi Vlădeni. Este situat pe terenuri cu exces temporar de umiditate cu soluri a căror regim aerohidric este necorespunzător, necesitând ample lucrări de afânare profundă şi desecări.
Descrierea din punct de vedere tipologic al acestor nardete a fost făcută până acum de MOTCĂ, 1972, care le numeşte "Tip Nardus stricta de terase plane, uşor mecanizabile pentru lucrările de îmbunătăţire" şi "Tip Nardus stricta cu Deschampsia caespitosa".
Mai recent KOVACS, 1978 descrie tipul "Pajişti de Nardus stricta situate pe terenuri plane sau uşor înclinate".
În aceste formulări se fac puţine referiri asupra caracteristicilor covorului ierbos, din care cauză s-a procedat la completarea denumirii.
Covorul ierbos aparţine asociaţiei Nardo-Juncetum subasociaţia agrostidetosum canini; molinetosum şi eriophoretosum, situate în întregime pe terenuri cu exces temporar sau permanent de umiditate.
În cadrul tipului se disting subtipurile de nardete: a. pe soluri profunde; b. pe soluri scheletice; c. cu muşuroaie înţelenite; d. cu vegetaţie lemnoasă şi combinaţii a+c, a+d, b+d, a+c+d;
remarcându-se o mare varietate a stadiilor de degradare.

117
Datorită acestor condiţii, variantele de îmbunătăţire şi valorificare a producţiei acestor nardete cuprind o gamă mai largă de măsuri la care încearcă să răspundă experimentările efectuate în staţionarul Vlădeni - Braşov. Valoarea furajeră a acestui tip este de asemenea scăzut, fiind de 9,2. Prin luarea unor măsuri corespunzătoare de îmbunătăţire aceste nardete pot fi transformate în pajişti productive. La acest proces contribuie mai ales posibilităţile de intervenţie cu mijloace mecanizate, aproape fără restricţii.
2.6. ALTE DESCRIERI ALE NARDETELOR 2.6.1. Bonitarea nardetelor
Prin bonitarea vegetaţiei şi a staţiunilor cu nardete din judeţul Braşov se confirmă încă o dată stadiul avansat de degradare a acestor pajişti (Tabelul 2.12). Clasele de vegetaţie sunt cuprinse între VII şi IX în medie VIII, corespunzătoare unor categorii de pajişti slabe-degradate, cu valoarea pastorală medie de 13,4, indicele de umiditate 5,22 şi producţia actuală de masă verde de 4,0 t/ha. Tipul de nardete cu clasa de vegetaţie cea mai ridicată (VII) este cel montan cu Festuca rubra şi Agrostis tenuis situat pe pante foarte slab până la mijlociu înclinate.
În cealaltă extremitate, cu cea mai scăzută clasă de vegetaţie (IX) sunt pajiştile de Nardus stricta alpine situate pe pante foarte puternic înclinate sau abrupte şi pajiştile de Nardus stricta din depresiuni cu diferite specii mezohigrofile şi higrofile situate pe terenuri plane sau uşor înclinate.
Bonitarea staţiunilor cu nardete, scoate în evidenţă prezenţa unor clase de favorabilitate ecologică scăzute cuprinse între VI (moderat favorabil) şi IX (slab favorabil) în medie VII, slab spre moderat favorabil. La o concluzie apropiată privind nardetele ajunge şi KOVACS At.şi colab.1978, care determină pentru vegetaţie clasele VII-VIII şi staţiune clasele VI-VIII după bonitarea unui tip de nardete situat pe terenuri plane sau uşor înclinate din perimetrul localităţii Crizbav - Braşov.
În linii generale există o corelaţie directă între clasele de favorabilitate ale staţiunilor, vegetaţia exprimând destul de fidel condiţiile staţionale.
Totuşi se remarcă o diferenţă între cele două clase, respectiv clasele de favorabilitate ecologică sunt mai ridicate ca valoare decât clasele de vegetaţie. În medie nardetele din judeţul Braşov prezintă o creştere cu o clasă a favorabilităţii ecologice, faţă de vegetaţie. În acest sens, între diferitele tipuri de nardete există diferenţe mai mari sau mai mici, uneori lipsesc.
Diferenţe mari între punctajele acordate staţiunilor şi vegetaţiei sunt înregistrate la tipurile de nardete colinare şi din depresiuni unde ajunge la 24-25 puncte mai mult în favoarea staţiunii, respectiv peste două clase. TUCRA I.şi colab.1980, remarcă diferenţe de 1-2 uneori 3 clase între clasele de favorabilitate a terenurilor pentru pajişti şi clasele şi categoriile de pajişti.
Această constatare pune în evidenţă stadiul foarte avansat de degradare a vegetaţiei în condiţii staţionale relativ bune, cu posibilităţi mari de îmbunătăţire, întreţinere şi valorificare a producţiei acestor pajişti.

118
Tabelul 2.12.
Caracteristicile capacităţii de producţie actuale a tipurilor de nardete din judeţul Braşov Characteristic of the current production capacity of Nardus stricta area type from Brasov County
Tipul de nardete Nr. staţiuni
Vegetative Staţiunea x) Pct. bonitare Dife-renţa (+, -)
Val. past.(Vp)
Ind. de umidit.
(U)
Prod. act.
mv t/ha
Cl. de vegetaţie
Alt. (m)
Înclinaţie
pH în H2O
V % Cl.de favora bilitate
Vege taţie
Staţi une
1.Nardete alpine pe pante foarte puternic încl. sau abrupte
18 0,53 4,61 1,8 IX 1820 2120
200- 450
4,5 4,9
12,5 16,3
IX 17 15 - 2
2. Nardete alpine pe terenuri plane sau uşor până la mijl. încl
12 0,61 5,03 2,3 VIII-IX 1780 2150
00- 150
4,6 4,9
13,1 18,2
VIII- VII
23 29 + 6
3. Nardete cu Vaccinum sp. 17 0,17 5,12 3,2 IX-VIII
1230 1710
100- 400
4,3 4,8
5,7 21,8
VIII 21 23 - 2
4. Nardete montane cu F. rubra şi A. capillaris situate pe pante puternic înclinate sau abrupte
32 1,11 4,91 3,1 VIII-VII 910 1620
200- 450
4,6 5,2
11,3 26,5
VII- VIII
30 31 + 1
5. Nardete montane cu F. rubra şi A. capillaris pe pante foarte slab până la mijlociu înclinate
23 1,16 5,10 3,9 VII 850 1600
20- 150
4,9 5,3
18,2 34,4
VI 36 45 + 9
6. Nardete cu diferite specii mezofile şi xeromezofile situate pe pante mijlociu până la foarte puternic înclinate
40 0,71 4,97 5,2 VIII-IX 540 820
100- 250
4,8 5,7
15,6 49,7
VI- V
23 48 + 25
7.Nardete din depresiuni cu diferite specii mezohigrofite şi hifrofite, situate pe terenuri plana sau uşor înclinate.
63 0,46 6,29 4,8 IX 480 750
00- 50
4,9 5,4
18,7 45,2
VI- VII
18 42 + 24
MEDIA x 0,67 5,22 4,0 VIII 1040 130 5,0 28,0 VII 26 37 + 11 X) Menţiune: pentru bonitarea staţiunii au fost luate în calcul toate datele cuprinse în tabelele 2.5. – 2.10. unde sunt trecute caracteristicile staţionale ale fitocenozei.

119
Diferenţele mai mici între punctajele de bonitare a vegetaţiei şi staţiuni (6-9
puncte în favoarea staţiunii) sunt înregistrate la tipurile de nardete subalpine şi montane situate pe pante foarte slab până la mijlociu înclinate, unde şi posibilităţile de îmbunătăţire şi valorificare a producţiei sunt ceva mai ridicate.
La tipurile de nardete subalpine şi montane, situate pe pante foarte puternic înclinate sau abrupte, datorită condiţiilor staţionale, diferenţele de punctaj între vegetaţie şi staţiune sunt neînsemnate având posibilităţi restrânse de îmbunătăţire şi valorificare a producţiei.
În concluzie, după bonitarea vegetaţiei şi a staţiunilor cu nardete se constată stadiul general avansat de degradare a acestor pajişti şi posibilităţile de creştere a productivităţii actuale, în funcţie de vegetaţia existentă şi condiţiile staţionale ale fiecărui tip de nardete în parte.
2.6.2. Relaţia între fitocenoze, tipuri de pajişti şi bonitare Studiul geobotanic, continuat cu cel tipologic şi de bonitare al nardetelor a scos
în evidenţă câteva caracteristici comune dar mai ales unele diferenţieri în funcţie de metodele aplicate şi obiective propuse a fi elucidate.
În primul rând studiul geobotanic a permis conturarea tipurilor de nardete pe care ulterior s-au efectuat lucrările de bonitare. Din acest punct de vedere, ca metodă de lucru se stabileşte un raport direct între fitocenoză tip-bonitare, atât ca ordine logică de efectuare a lucrărilor cât şi ca interdependenţă între aceste direcţii de studiu al vegetaţiei şi condiţiilor staţionale.
Releveurile floristice, efectuate în diferite staţiuni cu nardete au stat la baza studiilor geobotanice cât şi a părţii de vegetaţie din studiul tipologic şi al bonitării. Alături de analizele floristice datele asupra condiţiilor staţionale au diferenţiat şi conturat fitocenozele, dar mai ales au avut un rol determinant în descrierea tipurilor şi bonitarea nardetelor.
În ce priveşte corespondenţa dintre fitocenoză şi tip de nardete se constată unele diferenţieri destul de accentuate (Tabelul 2.13.).
Astfel, asociaţia Nardetum alpigenum include două tipuri de nardete diferenţiate mai ales după gradul de înclinare a terenului care determină în final posibilităţile de intervenţie pentru îmbunătăţirea şi valorificarea producţiei.
Celelalte două asociaţii Nardo - Campanuletum şi Nardo - Juncetum, înglobează de asemenea mai multe tipuri dar corespondenţa fitocenoză-tip se face la nivelul subasociaţiilor.
Tipul de Nardus stricta cu diferite specii de Vaccinium este situat pe o singură fitocenoză şi anume Nardo - Campanuletum nardetosum, care la rândul ei este reprezentată parţial şi în tipul de Nardus stricta montană cu Festuca rubra şi Agrostis tenuis situate pe pante foarte puternic înclinate sau abrupte. Acest din urmă tip de nardete se constituie în principal pe subasociaţia Nardo - Campanuletum festucetosum rubrae.

120
Tabelul 2.13. Corespondenţa dintre fitocenozele şi tipurile de nardete din judeţul Braşov
Analogy between phytogenesis and types of Nardus stricta areas from Brasov County
Fitocenoza Tipul
Nardetum alpigenum
Nardus stricta subalpin situat pe pante foarte puternic înclinate sau abrupte Nardus stricta subalpin situat pe terenuri plane sau uşor până la mijlociu înclinate
Nardo - Campanuletum nardetosum festucetosum rubrae
Nardus stricta cu diferite specii de Vaccinium Nardus stricta montan cu Festuca rubra şi Agrostis tenuis situat pe pante foarte puternic înclinate sau abrupte. Nardus stricta montan cu Festuca rubra şi Agrostis tenuis situat pe terenuri foarte slab până la mijlociu înclinate
Nardo - Juncetum - agrostidetosum tenui → - agrostidetosum canini → - molinietosum → - eriophoretosum →
Nardus stricta colinar, cu diferite specii mezofile şi xeromezofile situate pe pante mijlocii până la foarte puternic înclinate Nardus stricta din depresiuni cu diferite specii mezohigrofile şi higrofile situate pe terenuri plane sau uşor înclinate
În concluzie, nu există o corespondenţă directă asociaţie tip şi nici subordonare
între acestea. La acest rezultat ajung şi alţi autori (ANGHEL, MOTCĂ, 1975) care în depresiunea Făgăraş, disting pentru asociaţia de Nardus stricta, două tipuri de nardete în funcţie de gradul de mecanizare a terenului. Aceiaşi autori stabilesc mai multe asociaţii pentru un tip de pajişte şi în unele cazuri o asociaţie aparţine unui singur tip de pajişte. Astfel, se ajunge la concluzia că cele două unităţi de vegetaţie - asociaţie şi tipul sunt complet diferite.
În prezenta lucrare au fost făcute câteva încercări de stabilire a unor relaţii între fitocenoză şi subtip. Analizele efectuate nu au dus la concluzii sigure,întrucât subtipurile de nardete determinate după unele criterii cum sunt prezenţa vegetaţiei arborescente, a muşuroaielor, a volumului edafic util şi combinaţii dintre acestea, nu au influenţat decât în mică măsură compoziţia floristică de bază existentă pe ansamblul tipului de nardete analizat. Totuşi se constată că subtipurile cu muşuroaie sunt prezente pe terenuri relativ plane din judeţ, în fitocenozele Nardetum alpigenum, Nardo - Juncetum agrostidetosum canini şi altele.
Subtipurile cu vegetaţie arborescentă sunt semnalate pe tot cuprinsul zonei luate în studiu, dar mai ales în fitocenozele Nardo - Campanuletum nardetosum şi Nardo - Juncetum agrostidetosum tenuis.

121
Între tipurile de nardete şi bonitare există o relaţie directă. Tipul de nardete înglobează caracteristicile de vegetaţie şi staţionale determinate în final prin metoda bonitării.
Prin bonitare se stabilesc clase şi categorii de vegetaţie şi condiţii de staţionale exprimate prin limbajul cifrelor latine, care prezintă în final nivelul actual şi potenţial de producţie.
Clasele de favorabilitate ecologică a staţiunilor cu nardete sunt destul de diversificate fiind cuprinse între V-VI până la IX, respectiv de la moderat favorabile până la slab favorabile.Din acest punct de vedere există diferenţieri aproape nete între un tip de nardete şi altul.
Clasele de vegetaţie pentru nardete sunt mult mai constante variind între VII şi IX în medie VIII, exprimând de altfel caracteristicile unui covor ierbos degradat. Din acest punct de vedere tipul nr.1 de pajişti Nardus stricta subalpine, situate pe pante foarte puternic înclinate sau abrupte care însumează 17 puncte de bonitare, este asemănător tipului 7, de Nardus stricta din depresiuni cu diferite specii mezohigrofile şi higrofile situate pe terenuri plane sau uşor înclinate, care posedă 18 puncte, ambele tipuri fiind încadrate în clasa IX de vegetaţie, aceea a pajiştilor degradate.
În concluzie, deci se poate exprima că din punct de vedere al vegetaţiei, potenţialul actual al unui nardet din etajul subalpin de pe terenuri puternic înclinate poate să fie asemănător cu un nardet din zona colinară-depresionară de pe terenuri plane cu exces pronunţat de umiditate.
Astfel, mai multe tipuri de nardete sunt incluse în aceeaşi clasă de vegetaţie sau în cazuri mai rare unui tip în corespunde o clasă de vegetaţie distinctă. În ceea ce priveşte corespondenţa dintre tipurile de nardete şi clasele de favorabilitate ecologică a staţiunilor se constată o apropiere mai accentuată, respectiv unui tip îi corespunde de regulă o anumită clasă sau apropiat de clase de favorabilitate.
Între fitocenoză şi clasa de vegetaţie stabilită după bonitare există o legătură directă. Este de presupus ca mai multe fitocenoze să aparţină aceleiaşi clase de vegetaţie, aşa cum mai multe tipuri de nardete aparţin unei singure clase de vegetaţie.
Bonitarea vegetaţiei s-a făcut conform metodei de lucru, numai la nivelul fiecărui tip de nardete, unde se prezintă date comparative alături de bonitarea staţiunii.

122
III. ASPECTE PRIVIND ECOLOGIA PAJIŞTILOR DIN ZONA MONTANĂ
Acţiunile de reconstrucţie ecologică a pajiştilor degradate necesită o foarte
bună cunoaştere a ecologiei acestor ecosisteme. Pentru refacerea habitatelor de pajişti degradate este necesară identificarea corectă atât a cauzelor care au determinat degradarea acestora, cât şi a factorilor ecologici care au permis instalarea şi dezvoltarea unor anumitor tipuri de fitocenoze într-o staţiune oarecare.
În materialul de faţă nu ne-am propus o abordare exhaustivă a întregii problematici legate de ecologia pajiştilor, urmărindu-se, în special, evidenţierea rolului pe care diferiţi factori ecologici îl au în structurarea şi dinamica comunităţilor vegetale de pajişti.
Lipsa unor studii ecologice complete şi complexe, la nivel naţional şi pentru toate tipurile de pajişti, face imposibilă elaborarea unor „reţete” universal valabile, dar care ar fi foarte utile, mai ales pentru proprietarii de terenuri dar şi pentru specialiştii implicaţi în activităţi de reconstrucţie ecologică. De aceea, dorim să subliniem importanţa efectuării unor astfel de studii, cât mai aprofundate, atât pentru planificarea acţiunilor de reconstrucţie ecologică cât şi în vederea asigurării unui management optim care să prevină degradarea habitatelor şi necesitatea refacerii acestora.
Adresându-se specialiştilor, care sunt deja familiarizaţi cu teoriile ecologiei generale şi a ecologiei plantelor, în lucrarea de faţă sunt abordate doar acele aspecte care, în opinia noastră, sunt mai puţin cunoscute sau constituie un element de noutate, şi care pot oferi o altă perspectivă asupra ecologiei comunităţilor vegetale în general, şi a celor de pajişti în mod special.
3.1. PRINCIPALII FACTORI ECOLOGICI CARE INFLUENŢEAZĂ STRUCTURA ŞI DINAMICA HABITATELOR DE PAJIŞTI
Vegetaţia actuală este rezultatul interacţiunii continue, în spaţiu şi timp, atât
dintre speciile componente cât şi dintre acestea şi factorii din mediul în care trăiesc. Caracteristicile vegetaţie pot fi considerate ca o funcţie a climatului, materialului parental, topografie, organisme şi timp, aceasta putând fi exprimată astfel (AUSTIN 2005):
V f cl, p,r,o,t , unde:
V – caracteristicile vegetaţiei; cl – climatul; p – materialul parental; r – topografia; o – organismele; t – timpul.

123
Diferitele populaţii de plante care alcătuiesc o anumită comunitate vegetală reprezintă doar un anumit procent din populaţiile prezente în flora locală (GRIME 2001). Una dintre principalele provocări ale studiului covorului vegetal şi a acţiunilor de reconstrucţie ecologică a habitatelor fiind aceea de a identifica procesele şi factorii care determină abundenţa relativă a unei specii în cadrul unei comunităţi vegetale date.
Plantele, ca orice organisme vii, nu trăiesc izolat ci în strânsă interdependenţă atât cu alte organisme (plante sau animale), cât şi cu materia nevie, formând împreună mediul. Variabilele de mediu cele mai importante pot fi împărţite în patru mari categorii şi anume: factori topografici, factori climatici, factori edafici şi factori biotici care includ şi componenta antropică.
3.1.1. Factorii topografici (relieful)
Factorii topografici influenţează în mod indirect structura (specifică şi spaţială)
şi distribuţia fitocenozelor prin modificarea regimului climatic şi a factorilor edafici. Dintre aceştia, fără a minimaliza rolul celorlalţi factori, ne vom opri doar asupra altitudinii şi a indicelui topografic de umiditate (TMI), indice care estimează umiditatea potenţială a solului pe baza topografiei zonei luate în studiu.
a) Altitudinea este un factor de mediu care influenţează în mod indirect structura şi dinamica fitocenozelor, prin modificarea factorilor climatici, edafici şi biotici (POP 1977; BANATICLA et BUOT 2005). Astfel, odată cu creşterea altitudinii scade temperatura şi creşte nivelul precipitaţiilor, ceea ce duce la o creştere a umidităţii solului reflectată şi în viteza de descompunere a litierei (SCHINNER 1982), fapt care determină o creştere a substanţei organice din sol (AMUNDSON et al. 1989) şi o scădere a pH-ului (BANATICLA et BUOT 2005). De asemenea, creşterea altitudinii influenţează în mod direct durata sezonului de vegetaţie ca o consecinţă a duratei acoperirii cu zăpadă (DOLEZAL et SRUTEK 2002; PIPER et al. 2006).
Analiza relaţiilor existente între bogăţia specifică şi altitudine a demonstrat existenţa unor modele diferite (GRYTNES 2003; BRUUN et al. 2006), dintre care cele mai comune sunt modelele unimodale şi monoton descrescătoare (NOGUÉS-BRAVO et al. 2008). Una dintre ipotezele care încearcă să unească toate aceste modele este aceea că variaţia bogăţiei specifice în funcţie de altitudine urmează un model unimodal ca urmare a faptului că productivitatea şi rezerva locală de specii scad odată cu creşterea altitudinii (BRUUN et al. 2006). Din punct de vedere al speciilor rare, numărul acestora creşte odată cu creşterea altitudinii ca urmare a creşterii frecvenţei cu care apar speciile endemice şi slab competitive, care sunt specializate în ocuparea nişelor libere din medii în care valoare factorilor ecologici tinde spre extreme (WHITE et MILLER 1988).
Influenţa altitudinii asupra structurii comunităţilor vegetale se reflectă şi prin variaţia abundenţei-dominanţei diferitelor specii în funcţie de variaţia altitudinii diferitelor staţiuni, fapt ilustrat şi în fig. 3.1., în care se poate observa faptul că diferite specii realizează o abundenţă-dominanţă maximă la altitudini diferite.

124
Fig. 3.1. Răspunsul ecologic al unor specii de pajişti faţă de altitudine (Festrubr – Festuca rubra, Agrocapi – Agrostis capillaris, Calaarun –
Calamagrostis arundinacea, Calaepig – Calamagrostis epijeios, Anthvuln – Anthyllis vulneraria, Bracpinn – Brachypodium pinnatum, Desccaes- Deschampsia
caespitosa, Avenprae – Avenula praeusta, Nardstri – Nardus stricta, Scirsylv – Scirpus sylvaticus, Rumealpi – Rumex alpinus)
Ecological response of some grassland species to altitude Răspunsul ecologic al speciilor analizate în funcţie de variaţia altitudinii diferă
din punct de vedere al formei curbei de răspuns, al amplitudinii maxime şi al valorii optime la care fiecare specie realizează valoarea maximă a abundenţei-dominanţei.
Tot din punct de vedere altitudinal, aşa cum reiese din tabelul 3.1., lărgimea nişelor ecologice realizate de fiecare specie în parte este diferită. În cazul analizat, cele mai largi nişe le realizează speciile Festuca rubra, Nardus stricta şi Agrostis capillaris, acestea prezentând gradul cel mai mare de suprapunere peste nişele celorlaltor specii (94,6; 91,7 şi 86,4%) în timp ce prezintă şi cea mai mică proporţie în care nişele celorlalte specii se suprapun peste al lor (23,1; 25,3 şi 27,8%) (BĂRBOS 2007).
b) Indicele topografic de umiditate (TMI), deşi este corelat cu resursele de apă ale solului (MILLER et FRANKLIN 2002), nu poate fi considerat ca o unitate de măsură fidelă a umidităţii solului, fiind utilizat pentru a caracteriza din punct de vedere al efectului pe care particularităţile topografice îl au asupra distribuţie apei în sol (MEENTEMEYER et MOODY 2000). Totuşi, prin modul în care sunt distribuite valorile de abundenţă-dominanţă al speciilor edificatoare în funcţie de variaţia lui, reflectă destul de bine variaţia umidităţii relative a solului.
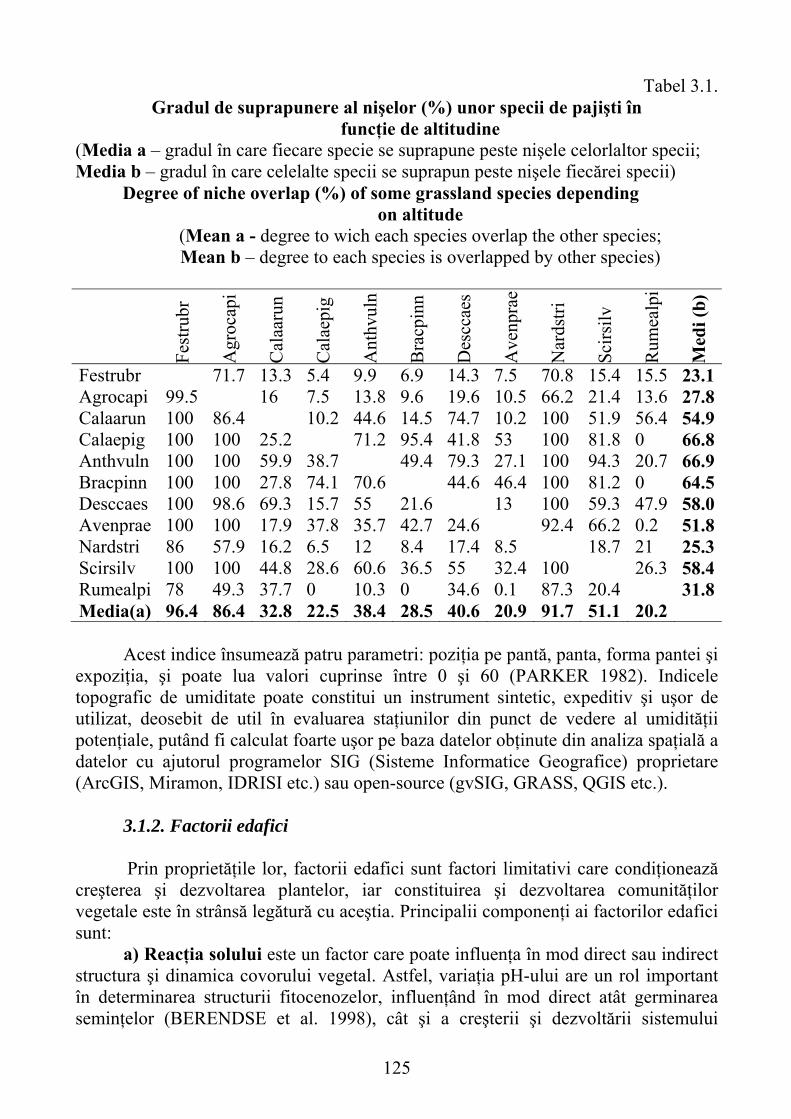
125
Tabel 3.1. Gradul de suprapunere al nişelor (%) unor specii de pajişti în
funcţie de altitudine (Media a – gradul în care fiecare specie se suprapune peste nişele celorlaltor specii; Media b – gradul în care celelalte specii se suprapun peste nişele fiecărei specii)
Degree of niche overlap (%) of some grassland species depending on altitude
(Mean a - degree to wich each species overlap the other species; Mean b – degree to each species is overlapped by other species)
F
estr
ubr
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsil
v
Rum
ealp
i
Med
i (b
)
Festrubr 71.7 13.3 5.4 9.9 6.9 14.3 7.5 70.8 15.4 15.5 23.1Agrocapi 99.5 16 7.5 13.8 9.6 19.6 10.5 66.2 21.4 13.6 27.8Calaarun 100 86.4 10.2 44.6 14.5 74.7 10.2 100 51.9 56.4 54.9Calaepig 100 100 25.2 71.2 95.4 41.8 53 100 81.8 0 66.8Anthvuln 100 100 59.9 38.7 49.4 79.3 27.1 100 94.3 20.7 66.9Bracpinn 100 100 27.8 74.1 70.6 44.6 46.4 100 81.2 0 64.5Desccaes 100 98.6 69.3 15.7 55 21.6 13 100 59.3 47.9 58.0Avenprae 100 100 17.9 37.8 35.7 42.7 24.6 92.4 66.2 0.2 51.8Nardstri 86 57.9 16.2 6.5 12 8.4 17.4 8.5 18.7 21 25.3Scirsilv 100 100 44.8 28.6 60.6 36.5 55 32.4 100 26.3 58.4Rumealpi 78 49.3 37.7 0 10.3 0 34.6 0.1 87.3 20.4 31.8Media(a) 96.4 86.4 32.8 22.5 38.4 28.5 40.6 20.9 91.7 51.1 20.2
Acest indice însumează patru parametri: poziţia pe pantă, panta, forma pantei şi expoziţia, şi poate lua valori cuprinse între 0 şi 60 (PARKER 1982). Indicele topografic de umiditate poate constitui un instrument sintetic, expeditiv şi uşor de utilizat, deosebit de util în evaluarea staţiunilor din punct de vedere al umidităţii potenţiale, putând fi calculat foarte uşor pe baza datelor obţinute din analiza spaţială a datelor cu ajutorul programelor SIG (Sisteme Informatice Geografice) proprietare (ArcGIS, Miramon, IDRISI etc.) sau open-source (gvSIG, GRASS, QGIS etc.). 3.1.2. Factorii edafici
Prin proprietăţile lor, factorii edafici sunt factori limitativi care condiţionează
creşterea şi dezvoltarea plantelor, iar constituirea şi dezvoltarea comunităţilor vegetale este în strânsă legătură cu aceştia. Principalii componenţi ai factorilor edafici sunt:
a) Reacţia solului este un factor care poate influenţa în mod direct sau indirect structura şi dinamica covorului vegetal. Astfel, variaţia pH-ului are un rol important în determinarea structurii fitocenozelor, influenţând în mod direct atât germinarea seminţelor (BERENDSE et al. 1998), cât şi a creşterii şi dezvoltării sistemului
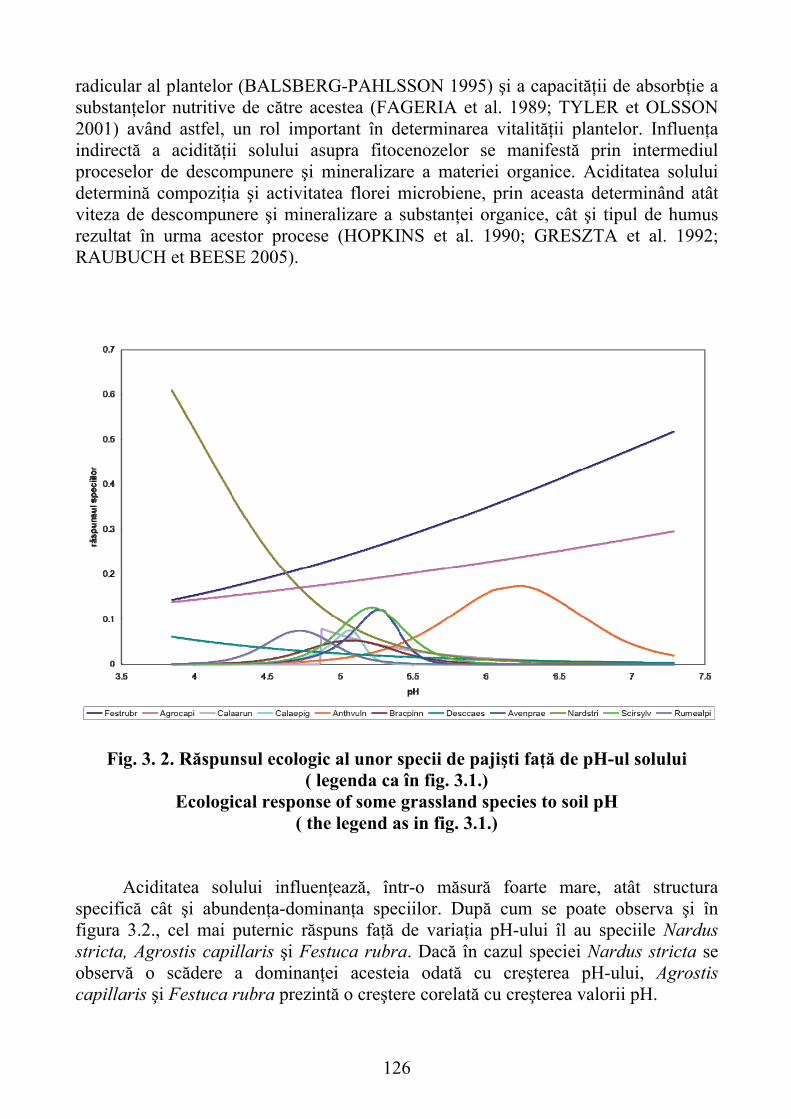
126
radicular al plantelor (BALSBERG-PAHLSSON 1995) şi a capacităţii de absorbţie a substanţelor nutritive de către acestea (FAGERIA et al. 1989; TYLER et OLSSON 2001) având astfel, un rol important în determinarea vitalităţii plantelor. Influenţa indirectă a acidităţii solului asupra fitocenozelor se manifestă prin intermediul proceselor de descompunere şi mineralizare a materiei organice. Aciditatea solului determină compoziţia şi activitatea florei microbiene, prin aceasta determinând atât viteza de descompunere şi mineralizare a substanţei organice, cât şi tipul de humus rezultat în urma acestor procese (HOPKINS et al. 1990; GRESZTA et al. 1992; RAUBUCH et BEESE 2005).
Fig. 3. 2. Răspunsul ecologic al unor specii de pajişti faţă de pH-ul solului ( legenda ca în fig. 3.1.)
Ecological response of some grassland species to soil pH ( the legend as in fig. 3.1.)
Aciditatea solului influenţează, într-o măsură foarte mare, atât structura specifică cât şi abundenţa-dominanţa speciilor. După cum se poate observa şi în figura 3.2., cel mai puternic răspuns faţă de variaţia pH-ului îl au speciile Nardus stricta, Agrostis capillaris şi Festuca rubra. Dacă în cazul speciei Nardus stricta se observă o scădere a dominanţei acesteia odată cu creşterea pH-ului, Agrostis capillaris şi Festuca rubra prezintă o creştere corelată cu creşterea valorii pH.
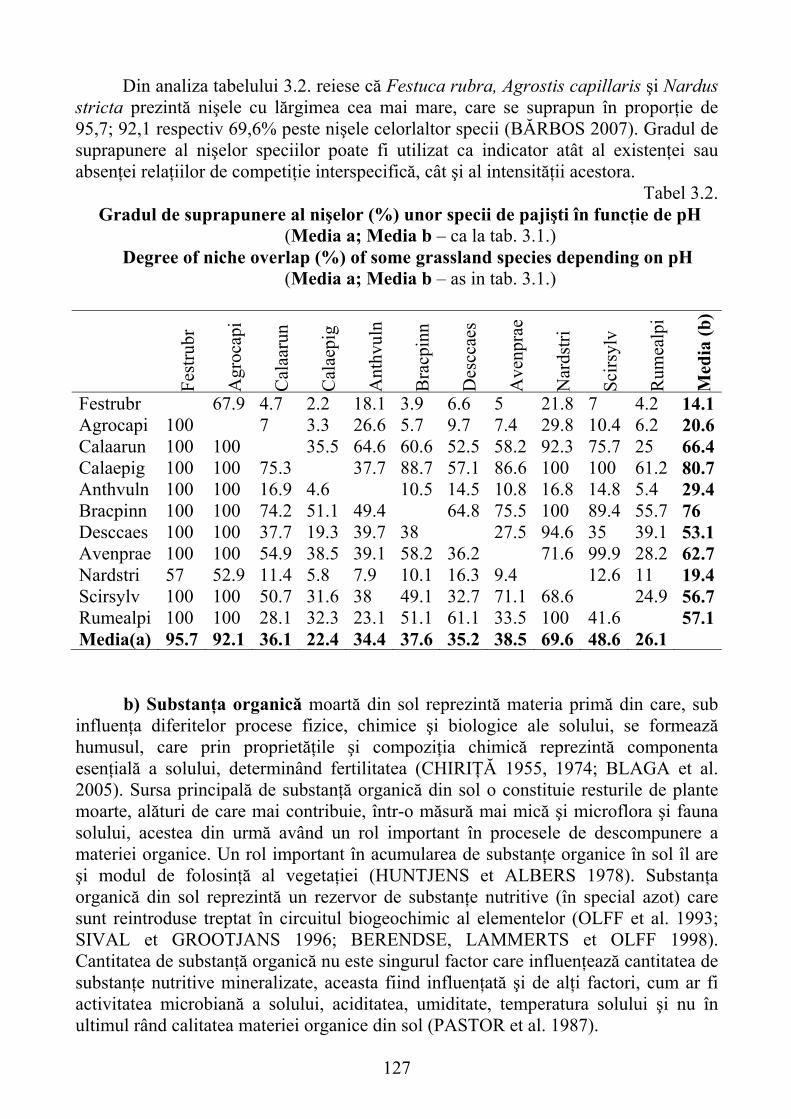
127
Din analiza tabelului 3.2. reiese că Festuca rubra, Agrostis capillaris şi Nardus stricta prezintă nişele cu lărgimea cea mai mare, care se suprapun în proporţie de 95,7; 92,1 respectiv 69,6% peste nişele celorlaltor specii (BĂRBOS 2007). Gradul de suprapunere al nişelor speciilor poate fi utilizat ca indicator atât al existenţei sau absenţei relaţiilor de competiţie interspecifică, cât şi al intensităţii acestora.
Tabel 3.2. Gradul de suprapunere al nişelor (%) unor specii de pajişti în funcţie de pH
(Media a; Media b – ca la tab. 3.1.) Degree of niche overlap (%) of some grassland species depending on pH
(Media a; Media b – as in tab. 3.1.)
Fes
trub
r
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsyl
v
Rum
ealp
i
Med
ia (
b)
Festrubr 67.9 4.7 2.2 18.1 3.9 6.6 5 21.8 7 4.2 14.1Agrocapi 100 7 3.3 26.6 5.7 9.7 7.4 29.8 10.4 6.2 20.6Calaarun 100 100 35.5 64.6 60.6 52.5 58.2 92.3 75.7 25 66.4Calaepig 100 100 75.3 37.7 88.7 57.1 86.6 100 100 61.2 80.7Anthvuln 100 100 16.9 4.6 10.5 14.5 10.8 16.8 14.8 5.4 29.4Bracpinn 100 100 74.2 51.1 49.4 64.8 75.5 100 89.4 55.7 76 Desccaes 100 100 37.7 19.3 39.7 38 27.5 94.6 35 39.1 53.1Avenprae 100 100 54.9 38.5 39.1 58.2 36.2 71.6 99.9 28.2 62.7Nardstri 57 52.9 11.4 5.8 7.9 10.1 16.3 9.4 12.6 11 19.4Scirsylv 100 100 50.7 31.6 38 49.1 32.7 71.1 68.6 24.9 56.7Rumealpi 100 100 28.1 32.3 23.1 51.1 61.1 33.5 100 41.6 57.1Media(a) 95.7 92.1 36.1 22.4 34.4 37.6 35.2 38.5 69.6 48.6 26.1
b) Substanţa organică moartă din sol reprezintă materia primă din care, sub influenţa diferitelor procese fizice, chimice şi biologice ale solului, se formează humusul, care prin proprietăţile şi compoziţia chimică reprezintă componenta esenţială a solului, determinând fertilitatea (CHIRIŢĂ 1955, 1974; BLAGA et al. 2005). Sursa principală de substanţă organică din sol o constituie resturile de plante moarte, alături de care mai contribuie, într-o măsură mai mică şi microflora şi fauna solului, acestea din urmă având un rol important în procesele de descompunere a materiei organice. Un rol important în acumularea de substanţe organice în sol îl are şi modul de folosinţă al vegetaţiei (HUNTJENS et ALBERS 1978). Substanţa organică din sol reprezintă un rezervor de substanţe nutritive (în special azot) care sunt reintroduse treptat în circuitul biogeochimic al elementelor (OLFF et al. 1993; SIVAL et GROOTJANS 1996; BERENDSE, LAMMERTS et OLFF 1998). Cantitatea de substanţă organică nu este singurul factor care influenţează cantitatea de substanţe nutritive mineralizate, aceasta fiind influenţată şi de alţi factori, cum ar fi activitatea microbiană a solului, aciditatea, umiditate, temperatura solului şi nu în ultimul rând calitatea materiei organice din sol (PASTOR et al. 1987).
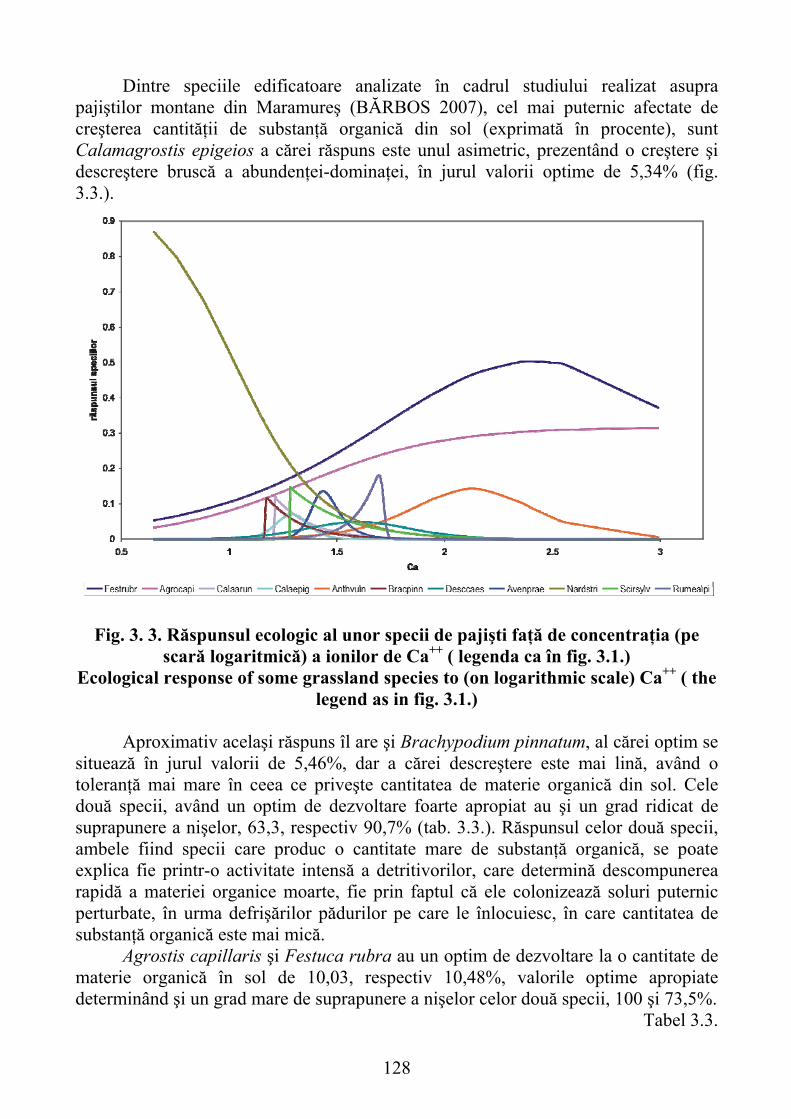
128
Dintre speciile edificatoare analizate în cadrul studiului realizat asupra pajiştilor montane din Maramureş (BĂRBOS 2007), cel mai puternic afectate de creşterea cantităţii de substanţă organică din sol (exprimată în procente), sunt Calamagrostis epigeios a cărei răspuns este unul asimetric, prezentând o creştere şi descreştere bruscă a abundenţei-dominaţei, în jurul valorii optime de 5,34% (fig. 3.3.).
Fig. 3. 3. Răspunsul ecologic al unor specii de pajişti faţă de concentraţia (pe scară logaritmică) a ionilor de Ca++ ( legenda ca în fig. 3.1.)
Ecological response of some grassland species to (on logarithmic scale) Ca++ ( the legend as in fig. 3.1.)
Aproximativ acelaşi răspuns îl are şi Brachypodium pinnatum, al cărei optim se
situează în jurul valorii de 5,46%, dar a cărei descreştere este mai lină, având o toleranţă mai mare în ceea ce priveşte cantitatea de materie organică din sol. Cele două specii, având un optim de dezvoltare foarte apropiat au şi un grad ridicat de suprapunere a nişelor, 63,3, respectiv 90,7% (tab. 3.3.). Răspunsul celor două specii, ambele fiind specii care produc o cantitate mare de substanţă organică, se poate explica fie printr-o activitate intensă a detritivorilor, care determină descompunerea rapidă a materiei organice moarte, fie prin faptul că ele colonizează soluri puternic perturbate, în urma defrişărilor pădurilor pe care le înlocuiesc, în care cantitatea de substanţă organică este mai mică.
Agrostis capillaris şi Festuca rubra au un optim de dezvoltare la o cantitate de materie organică în sol de 10,03, respectiv 10,48%, valorile optime apropiate determinând şi un grad mare de suprapunere a nişelor celor două specii, 100 şi 73,5%.
Tabel 3.3.
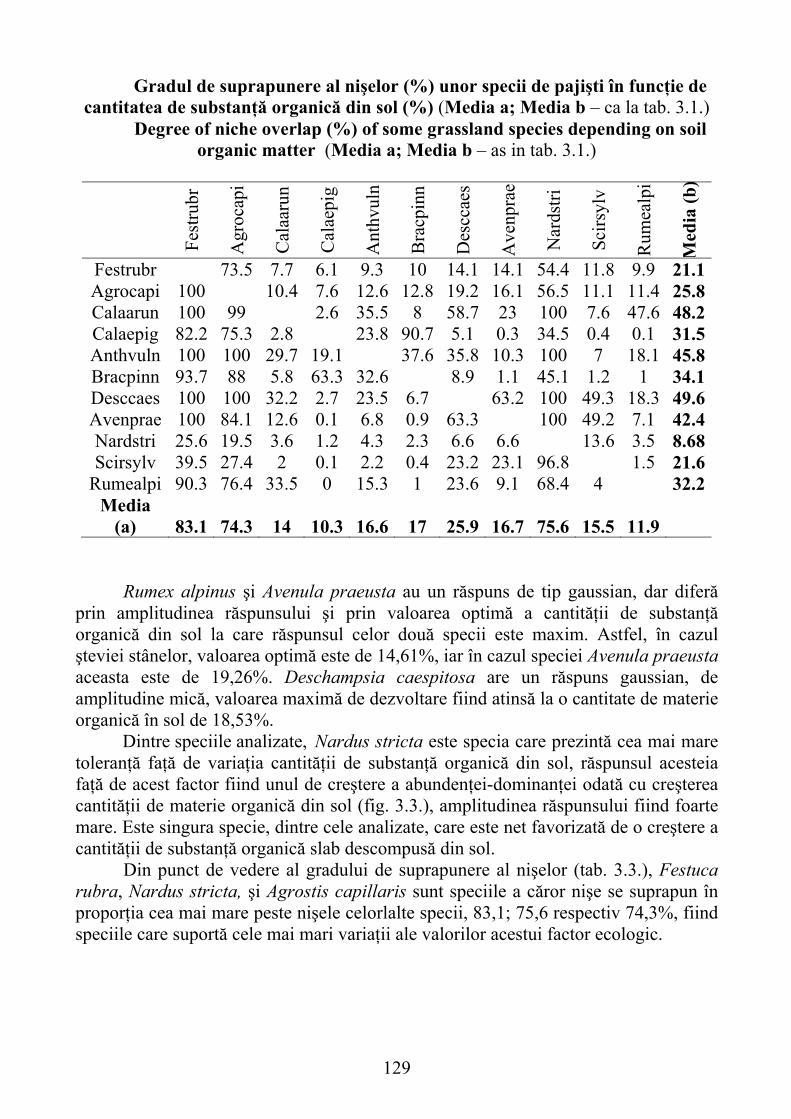
129
Gradul de suprapunere al nişelor (%) unor specii de pajişti în funcţie de cantitatea de substanţă organică din sol (%) (Media a; Media b – ca la tab. 3.1.)
Degree of niche overlap (%) of some grassland species depending on soil organic matter (Media a; Media b – as in tab. 3.1.)
Fes
trub
r
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsyl
v
Rum
ealp
i
Med
ia (
b)
Festrubr 73.5 7.7 6.1 9.3 10 14.1 14.1 54.4 11.8 9.9 21.1Agrocapi 100 10.4 7.6 12.6 12.8 19.2 16.1 56.5 11.1 11.4 25.8Calaarun 100 99 2.6 35.5 8 58.7 23 100 7.6 47.6 48.2Calaepig 82.2 75.3 2.8 23.8 90.7 5.1 0.3 34.5 0.4 0.1 31.5Anthvuln 100 100 29.7 19.1 37.6 35.8 10.3 100 7 18.1 45.8Bracpinn 93.7 88 5.8 63.3 32.6 8.9 1.1 45.1 1.2 1 34.1Desccaes 100 100 32.2 2.7 23.5 6.7 63.2 100 49.3 18.3 49.6Avenprae 100 84.1 12.6 0.1 6.8 0.9 63.3 100 49.2 7.1 42.4Nardstri 25.6 19.5 3.6 1.2 4.3 2.3 6.6 6.6 13.6 3.5 8.68Scirsylv 39.5 27.4 2 0.1 2.2 0.4 23.2 23.1 96.8 1.5 21.6
Rumealpi 90.3 76.4 33.5 0 15.3 1 23.6 9.1 68.4 4 32.2Media
(a) 83.1 74.3 14 10.3 16.6 17 25.9 16.7 75.6 15.5 11.9
Rumex alpinus şi Avenula praeusta au un răspuns de tip gaussian, dar diferă prin amplitudinea răspunsului şi prin valoarea optimă a cantităţii de substanţă organică din sol la care răspunsul celor două specii este maxim. Astfel, în cazul şteviei stânelor, valoarea optimă este de 14,61%, iar în cazul speciei Avenula praeusta aceasta este de 19,26%. Deschampsia caespitosa are un răspuns gaussian, de amplitudine mică, valoarea maximă de dezvoltare fiind atinsă la o cantitate de materie organică în sol de 18,53%.
Dintre speciile analizate, Nardus stricta este specia care prezintă cea mai mare toleranţă faţă de variaţia cantităţii de substanţă organică din sol, răspunsul acesteia faţă de acest factor fiind unul de creştere a abundenţei-dominanţei odată cu creşterea cantităţii de materie organică din sol (fig. 3.3.), amplitudinea răspunsului fiind foarte mare. Este singura specie, dintre cele analizate, care este net favorizată de o creştere a cantităţii de substanţă organică slab descompusă din sol.
Din punct de vedere al gradului de suprapunere al nişelor (tab. 3.3.), Festuca rubra, Nardus stricta, şi Agrostis capillaris sunt speciile a căror nişe se suprapun în proporţia cea mai mare peste nişele celorlalte specii, 83,1; 75,6 respectiv 74,3%, fiind speciile care suportă cele mai mari variaţii ale valorilor acestui factor ecologic.
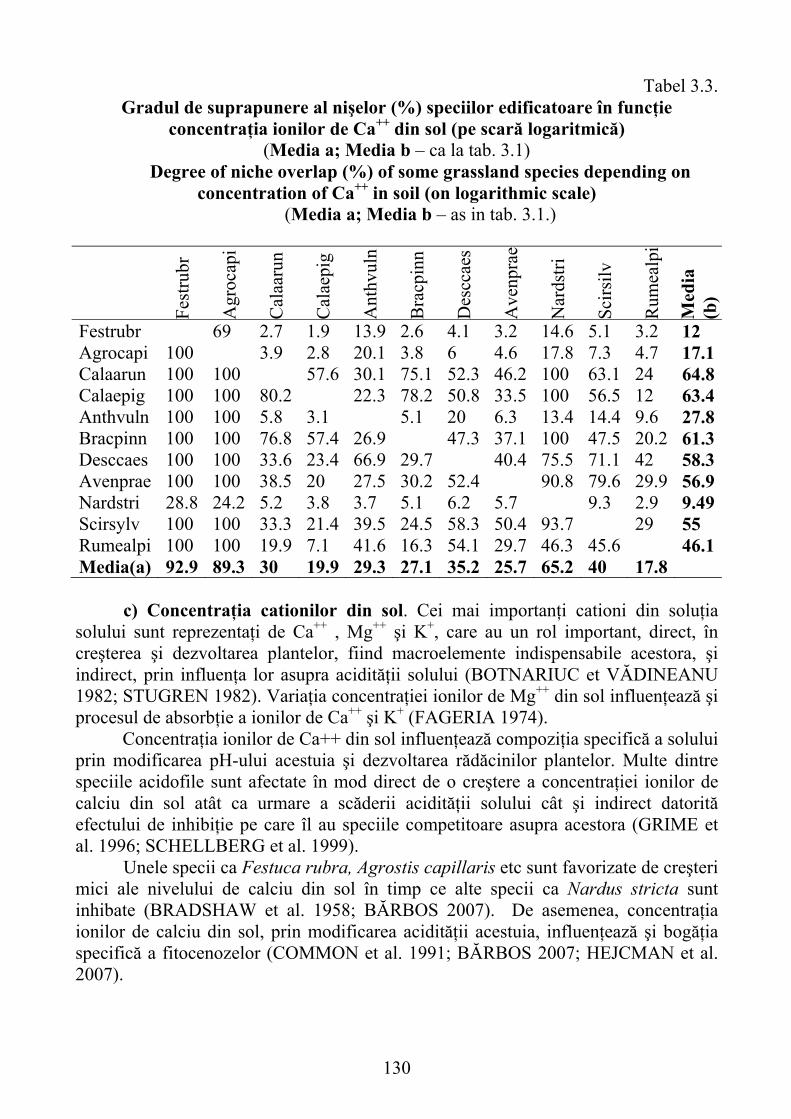
130
Tabel 3.3. Gradul de suprapunere al nişelor (%) speciilor edificatoare în funcţie
concentraţia ionilor de Ca++ din sol (pe scară logaritmică) (Media a; Media b – ca la tab. 3.1)
Degree of niche overlap (%) of some grassland species depending on concentration of Ca++ in soil (on logarithmic scale)
(Media a; Media b – as in tab. 3.1.)
Fes
trub
r
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsil
v
Rum
ealp
i
Med
ia
(b)
Festrubr 69 2.7 1.9 13.9 2.6 4.1 3.2 14.6 5.1 3.2 12 Agrocapi 100 3.9 2.8 20.1 3.8 6 4.6 17.8 7.3 4.7 17.1Calaarun 100 100 57.6 30.1 75.1 52.3 46.2 100 63.1 24 64.8Calaepig 100 100 80.2 22.3 78.2 50.8 33.5 100 56.5 12 63.4Anthvuln 100 100 5.8 3.1 5.1 20 6.3 13.4 14.4 9.6 27.8Bracpinn 100 100 76.8 57.4 26.9 47.3 37.1 100 47.5 20.2 61.3Desccaes 100 100 33.6 23.4 66.9 29.7 40.4 75.5 71.1 42 58.3Avenprae 100 100 38.5 20 27.5 30.2 52.4 90.8 79.6 29.9 56.9Nardstri 28.8 24.2 5.2 3.8 3.7 5.1 6.2 5.7 9.3 2.9 9.49Scirsylv 100 100 33.3 21.4 39.5 24.5 58.3 50.4 93.7 29 55 Rumealpi 100 100 19.9 7.1 41.6 16.3 54.1 29.7 46.3 45.6 46.1Media(a) 92.9 89.3 30 19.9 29.3 27.1 35.2 25.7 65.2 40 17.8
c) Concentraţia cationilor din sol. Cei mai importanţi cationi din soluţia
solului sunt reprezentaţi de Ca++ , Mg++ şi K+, care au un rol important, direct, în creşterea şi dezvoltarea plantelor, fiind macroelemente indispensabile acestora, şi indirect, prin influenţa lor asupra acidităţii solului (BOTNARIUC et VĂDINEANU 1982; STUGREN 1982). Variaţia concentraţiei ionilor de Mg++ din sol influenţează şi procesul de absorbţie a ionilor de Ca++ şi K+ (FAGERIA 1974).
Concentraţia ionilor de Ca++ din sol influenţează compoziţia specifică a solului prin modificarea pH-ului acestuia şi dezvoltarea rădăcinilor plantelor. Multe dintre speciile acidofile sunt afectate în mod direct de o creştere a concentraţiei ionilor de calciu din sol atât ca urmare a scăderii acidităţii solului cât şi indirect datorită efectului de inhibiţie pe care îl au speciile competitoare asupra acestora (GRIME et al. 1996; SCHELLBERG et al. 1999).
Unele specii ca Festuca rubra, Agrostis capillaris etc sunt favorizate de creşteri mici ale nivelului de calciu din sol în timp ce alte specii ca Nardus stricta sunt inhibate (BRADSHAW et al. 1958; BĂRBOS 2007). De asemenea, concentraţia ionilor de calciu din sol, prin modificarea acidităţii acestuia, influenţează şi bogăţia specifică a fitocenozelor (COMMON et al. 1991; BĂRBOS 2007; HEJCMAN et al. 2007).
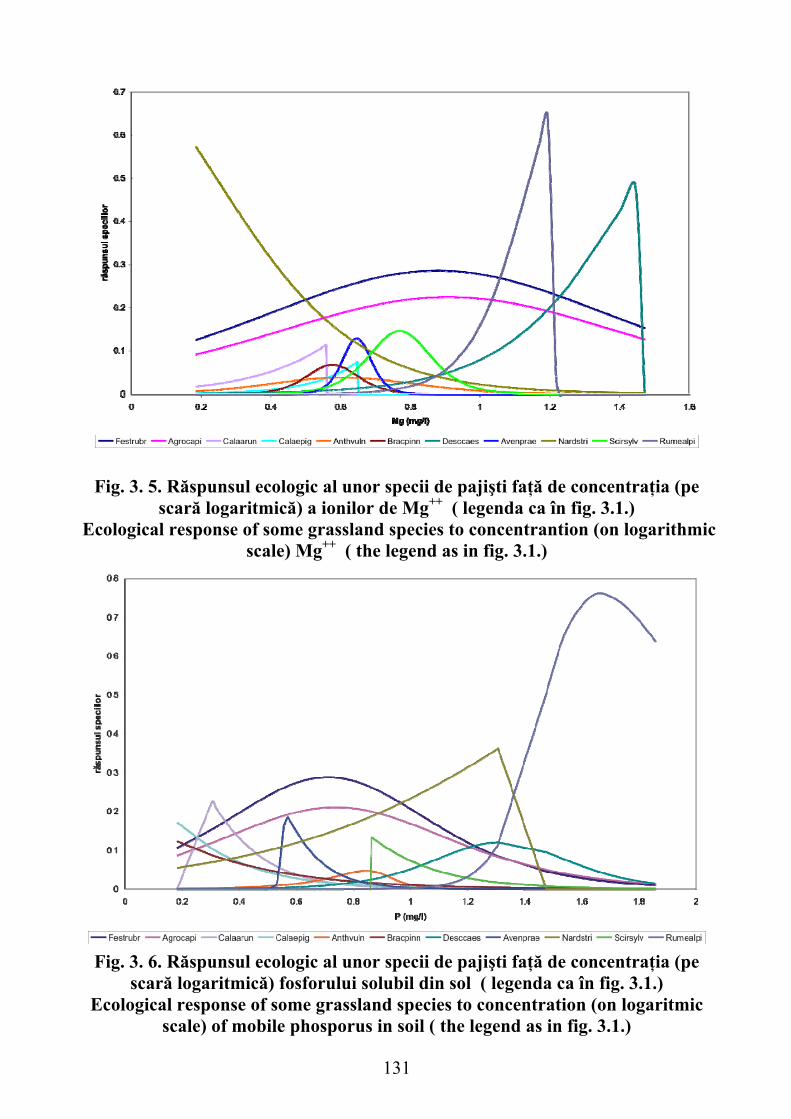
131
Fig. 3. 5. Răspunsul ecologic al unor specii de pajişti faţă de concentraţia (pe scară logaritmică) a ionilor de Mg++ ( legenda ca în fig. 3.1.)
Ecological response of some grassland species to concentrantion (on logarithmic scale) Mg++ ( the legend as in fig. 3.1.)
Fig. 3. 6. Răspunsul ecologic al unor specii de pajişti faţă de concentraţia (pe
scară logaritmică) fosforului solubil din sol ( legenda ca în fig. 3.1.) Ecological response of some grassland species to concentration (on logaritmic
scale) of mobile phosporus in soil ( the legend as in fig. 3.1.)
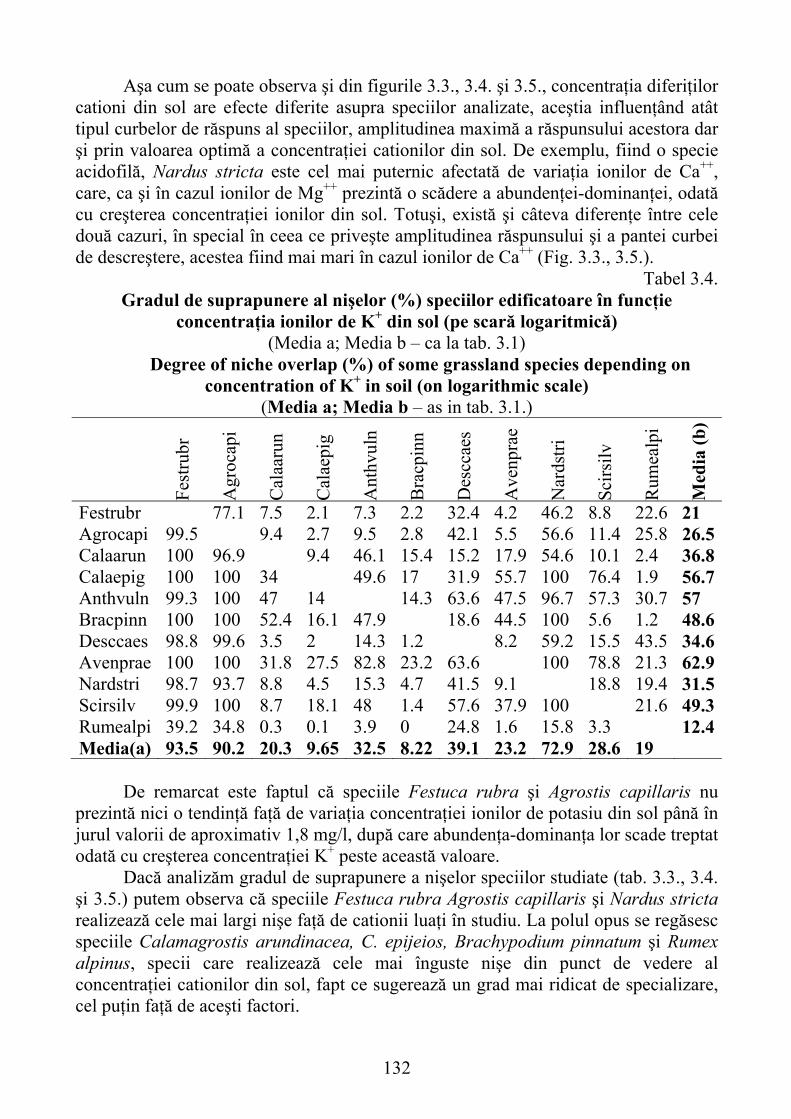
132
Aşa cum se poate observa şi din figurile 3.3., 3.4. şi 3.5., concentraţia diferiţilor cationi din sol are efecte diferite asupra speciilor analizate, aceştia influenţând atât tipul curbelor de răspuns al speciilor, amplitudinea maximă a răspunsului acestora dar şi prin valoarea optimă a concentraţiei cationilor din sol. De exemplu, fiind o specie acidofilă, Nardus stricta este cel mai puternic afectată de variaţia ionilor de Ca++, care, ca şi în cazul ionilor de Mg++ prezintă o scădere a abundenţei-dominanţei, odată cu creşterea concentraţiei ionilor din sol. Totuşi, există şi câteva diferenţe între cele două cazuri, în special în ceea ce priveşte amplitudinea răspunsului şi a pantei curbei de descreştere, acestea fiind mai mari în cazul ionilor de Ca++ (Fig. 3.3., 3.5.).
Tabel 3.4. Gradul de suprapunere al nişelor (%) speciilor edificatoare în funcţie
concentraţia ionilor de K+ din sol (pe scară logaritmică) (Media a; Media b – ca la tab. 3.1)
Degree of niche overlap (%) of some grassland species depending on concentration of K+ in soil (on logarithmic scale)
(Media a; Media b – as in tab. 3.1.)
Fes
trub
r
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsil
v
Rum
ealp
i
Med
ia (
b)
Festrubr 77.1 7.5 2.1 7.3 2.2 32.4 4.2 46.2 8.8 22.6 21 Agrocapi 99.5 9.4 2.7 9.5 2.8 42.1 5.5 56.6 11.4 25.8 26.5Calaarun 100 96.9 9.4 46.1 15.4 15.2 17.9 54.6 10.1 2.4 36.8Calaepig 100 100 34 49.6 17 31.9 55.7 100 76.4 1.9 56.7Anthvuln 99.3 100 47 14 14.3 63.6 47.5 96.7 57.3 30.7 57 Bracpinn 100 100 52.4 16.1 47.9 18.6 44.5 100 5.6 1.2 48.6Desccaes 98.8 99.6 3.5 2 14.3 1.2 8.2 59.2 15.5 43.5 34.6Avenprae 100 100 31.8 27.5 82.8 23.2 63.6 100 78.8 21.3 62.9Nardstri 98.7 93.7 8.8 4.5 15.3 4.7 41.5 9.1 18.8 19.4 31.5Scirsilv 99.9 100 8.7 18.1 48 1.4 57.6 37.9 100 21.6 49.3Rumealpi 39.2 34.8 0.3 0.1 3.9 0 24.8 1.6 15.8 3.3 12.4Media(a) 93.5 90.2 20.3 9.65 32.5 8.22 39.1 23.2 72.9 28.6 19
De remarcat este faptul că speciile Festuca rubra şi Agrostis capillaris nu prezintă nici o tendinţă faţă de variaţia concentraţiei ionilor de potasiu din sol până în jurul valorii de aproximativ 1,8 mg/l, după care abundenţa-dominanţa lor scade treptat odată cu creşterea concentraţiei K+ peste această valoare. Dacă analizăm gradul de suprapunere a nişelor speciilor studiate (tab. 3.3., 3.4. şi 3.5.) putem observa că speciile Festuca rubra Agrostis capillaris şi Nardus stricta realizează cele mai largi nişe faţă de cationii luaţi în studiu. La polul opus se regăsesc speciile Calamagrostis arundinacea, C. epijeios, Brachypodium pinnatum şi Rumex alpinus, specii care realizează cele mai înguste nişe din punct de vedere al concentraţiei cationilor din sol, fapt ce sugerează un grad mai ridicat de specializare, cel puţin faţă de aceşti factori.
133
Tabel 3.5. Gradul de suprapunere al nişelor (%) speciilor edificatoare în funcţie de
concentraţia ionilor de Mg++ din sol (pe scară logaritmică) (Media a; Media b – ca la tab. 3.1)
Degree of niche overlap (%) of some grassland species depending on concentration of Mg++ in soil (on logarithmic scale)
(Media a; Media b – as in tab. 3.1.)
Fes
trub
r
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsil
v
Rum
ealp
i
Med
ia (
b)
Festrubr 77.6 6.8 3.1 6.7 4.5 30.1 5 34.4 12.3 23.2 20.4Agrocapi 100 8.7 4 8.7 5.8 34.5 6.4 37 15.8 26.2 24.7Calaarun 100 100 21.8 40.3 25.5 6.7 3 100 3.3 0.6 40.1Calaepig 100 100 47.2 85.1 99.3 24.6 56.9 100 39 2.8 65.5Anthvuln 100 100 40.5 39.5 52 43.6 37.3 100 58.4 24.9 59.6Bracpinn 100 100 38.1 68.7 77.6 22.8 64.5 100 51.7 3.8 62.7Desccaes 66.7 59.3 1 1.7 6.5 2.3 2.5 9.5 10.5 31.5 19.2Avenprae 100 100 4.2 35.9 50.8 58.8 22.9 96.6 63.1 8 54 Nardstri 62.8 52.4 12.4 5.7 12.3 8.3 7.9 8.7 13.6 5 18.9Scirsilv 100 100 1.8 9.9 32 19 38.8 25.4 60.3 28.9 41.6
Rumealpi 64.1 56.4 0.1 0.2 4.7 0.5 39.4 1.1 7.6 9.8 18.4Media (a) 89.4 84.6 16.1 19.1 32.5 27.6 27.1 21.1 64.5 27.8 15.5
d) Fosforul şi azotul sunt pricipalii nutrienţi necesari pentru creşterea şi dezvoltarea plantelor (PARTON et al. 1988; NIINEMETS et KULL 2005), raportul dintre cele două elemente din ţesutul plantelor indicând care dintre ele are rol de factor limitativ. Astfel, un raport N:P < 10 (GUSEWELL 2004) sau < 14 (MAMOLOS et al. 2005) indică faptul că azotul are rol limitativ, iar un raport N:P >16 (MAMOLOS, VASILIKOS et VERESOGLOU 2005) sau > 20 (GUSEWELL 2004) indică faptul că fosforul are rol limitativ. Dacă din punct de vedere cantitativ, o mare parte a variabilităţii fosforului din sol poate fi atribuită texturii solului (STEWART et COLE 1989), din punct de vedere calitativ un rol important îl are troficitatea solului şi tipul de vegetaţie. Astfel, în solurile oligotrofe, fosforul se găseşte predominant sub formă organică, disponibilitatea sa pentru plante fiind condiţionată de mineralizarea acesteia (WILLIAMS et al. 1999). Din punct de vedere al vegetaţiei, aceasta afectează distribuţia formelor de fosfor din sol, pajiştile conţinând fosfor în combinaţii organice labile sau moderat-labile, care poate fi potenţial utilizabil de către plante (MAGID 1993).
Influenţa fosforului în viaţa plantelor se manifestă atât direct, prin rolul nutritiv, cât şi indirect, fiind unul dintre factorii care condiţionează mineralizarea azotului, fertilizarea solurilor cu fosfor având un impact pozitiv asupra circuitului azotului (PURCHASE 1974). Efectul fosforului asupra mineralizării azotului este amplificat de prezenţa vegetaţiei, datorită stimulării creşterii rădăcinilor, fapt ce
134
determină o creştere a volumului rizosferei şi, implicit, a numărului de microorganisme nitrificatoare (CORNISH et RAISON 1977).
Analizând figura 3.6., se poate observa că, specia cu răspunsul cel mai puternic faţă de variaţia cantităţii fosforului din sol este Rumex alpinus, a cărei dinamică urmează o curbă asimetrică, urmată fiind de speciile Agrostis capillaris, Festuca rubra şi Nardus stricta. Ca şi în cazul celorlalţi factori ecologici analizaţi, păiuşul roşu, iarba câmpului şi ţepoşica sunt speciile cu nişele cele mai largi, acestea suprapunându-se în proporţie de 83,2; 80,5 şi 72,6 % peste nişele celorlaltor specii luate în studiu (tab.3.6.).
Tabel 3.6. Gradul de suprapunere al nişelor (%) speciilor edificatoare în funcţie
concentraţia fosforului solubil din sol (pe scară logaritmică) (Media a; Media b – ca la tab. 3.1)
Degree of niche overlap (%) of some grassland species depending on concentration of mobile phosphorus in soil (on logarithmic scale)
(Media a; Media b – as in tab. 3.1.)
Fes
trub
r
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsil
v
Rum
ealp
i
Med
ia
(b)
Festrubr 78.1 19.5 14.8 6.1 16.9 20.3 10.6 61.4 11.9 9.9 25 Agrocapi 96.7 20.8 18.1 7.5 20.3 28 13.1 72.9 14.8 15.5 30.8Calaarun 86.9 74.9 57.2 12.1 54.6 8.1 15.6 57 1.8 1 36.9Calaepig 100 99.1 86.8 18.8 94.9 12.7 22.5 85.3 3 1.7 52.5Anthvuln 100 100 44.5 45.5 66.1 56.9 55.6 100 31.8 4.2 60.5Bracpinn 100 97.1 72.3 83 23.9 22.4 25.7 84.7 12.2 7.1 52.8Desccaes 74.9 83.5 6.7 6.9 12.8 14 7.6 72.8 29.6 58 36.7Avenprae 100 100 33.1 31.4 32 41 19.6 88.4 5.5 2.1 45.3Nardstri 65.6 63 13.7 13.5 6.5 15.3 21.1 10 12 13 23.4Scirsilv 100 100 3.3 3.7 16.3 17.3 67.3 4.9 93.9 27.8 43.5Rumealpi 7.5 9.5 0.2 0.2 0.2 0.9 11.9 0.2 9.2 2.5 4.23Media(a) 83.2 80.5 30.1 27.4 13.6 34.1 26.8 16.6 72.6 12.5 14
Din punct de vedere al influenţei fertilităţii asupra bogăţiei specifice, în general
este acceptat ca productivitatea ecosistemelor creste odată cu creşterea fertilităţii solului, iar bogăţia specifica are un răspuns unimodal fata de variaţia productivităţii (GRIME 2001). Resursele de azot din sol sunt atât minerale sub formă de azot nitric şi amoniacal, cât şi organice sub formă de aminoacizi simpli care pot fi absorbiţi de către plante (NÄSHOLM et al. 2000). Solurile acide din zona temperată şi arctic-alpină, sunt mai bogate în azot amoniacal decât în azot nitric (GIGON et RORISON 1972; ATKIN et CUMMINS 1994). Preferinţa plantelor pentru azotul amoniacal (NH4
+ ) poate fi pusă pe seama mecanismelor diferite de absorbţie a azotului, absorbţia azotului amoniacal necesitând un consum mai mic de energie decât al azotului nitric (ATKIN et CUMMINS 1994)

135
3.1.3. Factorii climatici Prin climat se înţelege acţiunea complexă a factorilor meteorologici (lumina,
temperatura, umiditatea, precipitaţiile, vântul) pe o anumită suprafaţă a Terrei (POP 1977). În cadrul acestui climat general pot fi deosebite subunităţi mai mici cum ar fi macroclimatul sau climatul regional, mezoclimatul sau climatul local (topoclimatul) şi microclimatul sau climatul microreliefului care include şi fitoclimatul (CRISTEA 1991). Din punctul de vedere al studiilor de vegetaţie, un rol important îl are analiza macro- şi microclimatului, care condiţionează în mod direct structura şi răspândirea fitocenozelor. Din categoria factorilor climatici, pe lângă precipitaţii, temperatură, vânt etc., a căror influenţe asupra covorului vegetal sunt bine cunoscute, un rol important îl au şi radiaţia solară incidentă şi indicele de încălzire.
a) Radiaţia solară incidentă care este de departe sursa dominantă de energie
pentru majoritatea organismelor vii (SCHULZE, BECK et MÜLLER-HOHENSTEIN 2005) şi este singura sursă de energie care poate fi utilizată în activităţile metabolice de către plantele verzi, fiind convertită în energie chimică în procesul de fotosinteză (BEGON et al. 2006). Radiaţia solară este un factor foarte complex care prezintă variaţii simultane ale intensităţii, compoziţiei spectrale şi a distribuţiei spaţiale (EVANS 1969; SCHULZE, BECK et MÜLLER-HOHENSTEIN 2005).
Variaţia zilnică sau sezonieră a radiaţiei solare directe este puternic corelată atât cu umiditatea solului şi a aerului, datorită evapotranspiraţiei (GRANGER et SCHULZE 1977; KIRKPATRICK et al. 1988; FITTER et al. 1998; NEWELL et PEET 1998) , cât şi cu temperatura şi intensitatea luminii, factori care influenţează în mod direct sau indirect structura şi dinamica vegetaţiei (KIRKPATRICK et NUNEZ 1980).
b) Indicele de încălzire (HL), care, chiar dacă nu reprezintă decât o estimare a
cantităţii de energie termică potenţiale pentru o anumită staţiune, totuşi poate oferi informaţii importante privind bugetul termic al solului şi a stratului de aer din imediata sa apropiere. Temperatura joacă un rol important atât în viaţa plantele, fiind unul dintre principalii factori care declanşează sau opresc activităţile metabolice din plante (SMITH 1996; SCHULZE, BECK et MÜLLER-HOHENSTEIN 2005), cât şi în procesele biologice, chimice şi fizice din sol (WEAVER et CLEMENTS 1938). Principalii factori care determină regimul termic al unei staţiunii variază de la radiaţia solară incidentă directă, care este principala sursă de încălzire (KORMONDY 1996) până la culoarea, textura, structura solului, cantitatea de humus din sol, conţinutul de apă al solului şi acoperirea cu vegetaţie a solului (WEAVER et CLEMENTS 1938).
Atât indicele de încălzire cât şi radiaţia solară incidentă sunt două variabile uşor de estimat cu ajutorul programelor SIG, constituind elemente importante în evaluarea condiţiilor staţionale generale ale unei anumite zone.

136
3.1.4. Factorii biotici
Alături de factorii abiotici, factorii biotici (vii) contribuie la constituirea, structurarea şi distribuţia spaţială a fitocenozelor. Dintre aceştia putem aminti acoperirea realizată de stratul de muşchi, grosimea stratului de litieră, relaţiile intra - şi interspecifice de competiţie şi facilitare şi modul de folosinţă al pajiştilor (factorul antropo-zoogen) care, de fapt, se răsfrânge în mod direct asupra primilor trei factori.
a) Acoperirea realizată de stratul de muşchi are un rol important în
determinarea structurii şi a dinamicii comunităţilor de plante vasculare, acţionând în mod direct, ca un filtru, asupra speciilor, unele specii de muşchi putând avea efecte alelopatice asupra cormofitelor (KEIZER et al. 1985) (KEIZER et al. 1985). O altă modalitate prin care muşchii influenţează structura şi dinamica covorului vegetal este prin modificarea condiţiilor microclimatice (chiar nanoclimatice) din fiecare staţiune. Astfel, covorul de muşchi determină o modificare a temperaturii de la suprafaţa solului concomitent cu o creştere a umidităţii (SEDIA et EHRENFELD 2003; OTSUS et ZOBEL 2004; ZAMFIR 2000), dar influenţează şi cantitatea şi calitatea luminii (KEIZER, VANTOOREN et DURING 1985). Aceste modificări ale microclimatului se răsfrâng asupra procesului de germinaţie a seminţelor dar şi în creşterea şi dezvoltarea plantulelor, efectele asupra plantelor vasculare fiind diferite în funcţie de specie şi habitat (ZAMFIR 2000). Păturile compacte de muşchi joacă şi un rol de barieră fizică, putând să împiedice astfel germinaţia seminţelor, dar mai ales creşterea şi dezvoltarea plantulelor, dar, odată stabilite, plantele vasculare creează un microhabitat mult mai propice dezvoltării lor (SEDIA et EHRENFELD 2003).
De asemenea, muşchii au un rol important şi în circuitul biogeochimic al elementelor, în special al azotului (CURTIS et al. 2005). Prezenţa covorului de muşchi favorizează activitatea microbiană şi acumularea de materie organică, fapt ce determină crearea unor microstaţiuni bogate în substanţe nutritive şi azot amoniacal (SEDIA et EHRENFELD 2005). Variaţiile spaţiale şi temporale în structura covorului de muşchi pot constitui unul dintre motivele variaţiei diversităţii comunităţilor vegetale (KEIZER, VANTOOREN et DURING 1985).
b) Litiera, este un factor de mediu cu un rol foarte complex, având o
importanţă deosebită în determinarea structurii şi dinamicii fitocenozelor. În funcţie de modul în care influenţează plantele şi comunităţile vegetale, efectele litierei pot fi:
- directe – constituind o barieră fizică pentru creşterea şi dezvoltarea plantulelor (HECTOR et al. 2000), pentru disiparea vaporilor de apă din sol (MCALPINE et DRAKE 2003), împiedică contactul direct dintre seminţe şi sol (ROTUNDO et AGUIAR 2005), împiedică eroziunea solului datorată apelor pluviale şi protejează solul împotriva îngheţului (WALSH et VOIGT 1977), oferă protecţie seminţelor şi plantulelor tinere împotriva granivorelor şi ierbivorilor (QUESTED et ERIKSSON 2006);
- indirecte – determinând schimbări microclimatice care afectează temperatura şi umiditatea solului dar şi cantitatea şi calitatea radiaţiei solare (ECKSTEIN et DONATH 2005; FACELLI ET PICKETT 1991; BOSY et READER 1995;FACELLI

137
et CARSON 1991; NAKAMURA 1996), determinând o scădere a variaţiilor temperaturii (BOSY et READER 1995), ameliorează condiţiile de umiditate a solului, mai ales în perioadele de stres hidric (MARET et WILSON 2005) şi reduce gradul de iradiere al solului cu până la 99% (Bosy et Reader 1995). Proprietăţile de infiltrare, structura şi textura, cel puţin a orizontului superior al solului sunt determinate, parţial, de cantitatea şi calitatea litierei (WALSH et VOIGT 1977). De asemenea, litiera are un rol important în circuitul biogeochimic al substanţelor nutritive (HECTOR et al. 2000; ECKSTEIN et DONATH 2005; QUESTED et ERIKSSON 2006; AERTS et al. 2003; WALSH et VOIGT 1977).
În funcţie de mecanismele care stau la baza influenţelor exercitate de gosimea stratului de litieră asupra comunităţilor vegetale putem deosebi următoarele efecte:
- efecte fizice – sunt rezultatul proceselor care duc la modificarea condiţiilor microclimatice, determină interacţiunile plantelor cu mediul fizic (NAKAMURA 1996;ROTUNDO et AGUIAR 2005). Acumularea de litieră prezintă o heterogenitate spaţială datorată heterogenităţii vegetaţiei, a redistribuirii litierei şi descompunerii ei inegale, ceea ce determină o heterogenitate a condiţiilor microclimatice (FACELLI et PICKETT 1991);
- efecte chimice – care se datorează în primul rând proceselor de descompunere la care este supusă litiera, acesta fiind un determinant major al înmagazinării carbonului şi al altor nutrienţi şi al fluxului de materie în ecosisteme (QUESTED et ERIKSSON 2006). Descompunerea litierei şi eliberarea nutrienţilor este controlată de o mare varietate de proprietăţi chimice ale acesteia, dintre care putem aminti concentraţia de N, raportul C:N, concentraţia de P, raportul C:P, raportul N:P, cantitatea de lignină, concentraţia compuşilor fenolici (AERTS, de CALUWE et BELTMAN 2003). Proprietăţile chimice ale litierei sunt determinate de compoziţia calitativă a acestei care se datorează, în mare parte, speciilor dominante (HOORENS et al. 2003; HOORENS et al. 2002). Litiera speciilor de dicotiledonate poate avea un efect chimic mult mai puternic decât cea a gramineelor datorită fitotoxinelor şi a eliberării de nutrienţi într-o perioadă de timp mai scurtă, viteza de descompunere a litierei produsă de graminee fiind mai mică (XIONG et al. 2003). De asemenea, descompunerea litierei afectează pH-ul şi conţinutul chimic al apelor de percolare şi conduce la formarea acizilor prehumici, humici şi fulvici (WALSH et VOIGT 1977), având un efect de fertilizare a solului (MONK et GABRIELSON 1985);
- efecte biologice – dintre care menţionăm reducerea densităţii şi a numărului de specii (CARSON et PETERSON 1990; MONK et GABRIELSON 1985; FOSTER et GROSS 1998), influenţează germinaţia seminţelor, creşterea şi dezvoltarea plantulelor, incidenţa bolilor fungice, prezenţa şi intensitatea fenomenelor alelopatice (ROTUNDO et AGUIAR 2005; XIONG et al. 2003) şi dinamica populaţiilor de nevertebrate din sol (XIONG et NILSSON 1999). De asemenea, afectează şi relaţiile intra – şi interspecifice ale indivizilor populaţiilor de plante (NAKAMURA 1996), cu rol important în structurarea fitocenozelor. Dacă analizăm rolul litierei asupra populaţiilor de plante şi a comunităţilor vegetale din punct de vedere calitativ, efectele acesteia pot fi:
- efecte pozitive – care se pot datora îmbunătăţirii condiţiilor hidrice şi reducerea stresului termic, aportului de substanţe nutritive şi a diseminării seminţelor

138
ca urmare a redistribuirii ei (ECKSTEIN et DONATH 2005;QUESTED et ERIKSSON 2006; XIONG et NILSSON 1999; FOSTER et GROSS 1998; MARET et WILSON 2005), iar în stadiile iniţiale ale succesiunii, acumularea de litieră promovează dezvoltarea plantulelor (NAKAMURA 1996);
- efecte negative – determinate în general de creşterea cantităţii de litieră ceea ce duce la o reducere a intensităţii luminii la nivelul solului (FACELLI et PICKETT 1991), creşterea numărului de bacterii şi ciuperci patogene ca urmare a creşterii umidităţii şi a temperaturii (MARET et WILSON 2005; BOSY et READER 1995), inhibă germinaţia seminţelor, în special a celor mici, a căror germinaţie este redusă cu până 100%, şi creşterea şi dezvoltarea plantulelor (FOSTER et GROSS 1998; BOSY et READER 1995; MCALPINE et DRAKE 2003). Toate aceste modificări cauzate de acumularea de litieră determină o scădere a bogăţiei specifice, a biodiversităţii şi a productivităţii fitocenozelor (ECKSTEIN et DONATH 2005; MONK et GABRIELSON 1985).
Litiera este un factor fundamental care controlează structura, stabilitatea şi dinamica comunităţilor vegetale (GRIME 2001; CARSON et PETERSON 1990), având un rol mai important în organizarea fitocenozelor decât în circuitul biogeochimic al nutrienţilor (FACELLI et CARSON 1991).
Analiza curbelor de răspuns al speciilor edificatoare (Fig. 3.7) şi a gradului de suprapunere al nişelor (Tab. 3.7.) permite evidenţierea distribuţiei valorilor de abundenţă-dominanţă a speciilor edificatoare studiate. Astfel, răspunsul speciilor Anthyllis vulneraria şi Agrostis capillaris se caracterizează printr-o scădere a valorilor de abundenţă-dominanţă odată cu creşterea grosimii stratului de litieră, vătămătoarea fiind mult mai afectată de acest factor decât iarba câmpului.
Răspunsul speciei Festuca rubra, răspunsul ecologic este asimetric, amplitudinea maximă a acestuia fiind atinsă la o grosime de 4,3 mm a stratului de litieră.
În cazul speciilor de Nardus stricta şi Scirpus sylvaticus, se înregistrează un răspuns asimetric dar acestea diferă atât prin amplitudinea maximă a răspunsului, cât şi prin grosimea stratului de litieră la care se situează aceasta. Astfel, în cazul speciei Nardus stricta amplitudinea maximă este atinsă la o grosime a litierei de 39,25 mm, iar în cazul speciei Scirpus sylvaticus la o grosime de 56,93 mm.
Deşi, şi în acest caz speciile Festuca rubra, Nardus stricta şi Agrostis capillaris realizează cele mai largi nişe, gradul de suprapunere al acestora peste nişele celorlaltor specii fiind de 72,1; 68,7 şi 59,5%, totuşi se poate remarca o îngustare a nişelor realizate comparativ cu lărgimea nişelor realizate în cazul celorlalţi factori ecologici analizaţi, ceea ce subliniază importanţa litierei în structurarea fitocenozelor. La polul opus, cele mai înguste nişe sunt realizate de speciile Anthyllis vulneraria şi Calamagrostis arundinacea, a căror grad de suprapunere peste nişele celorlaltor specii este de doar 7,69 şi 14,3%.
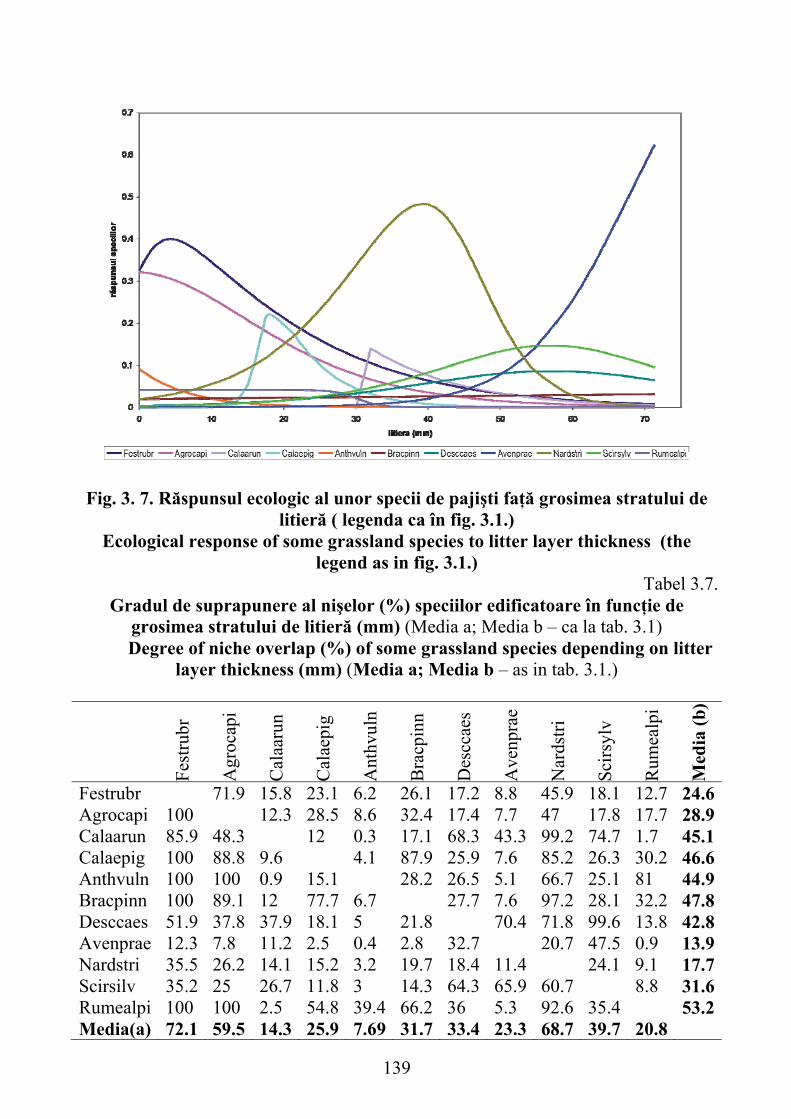
139
Fig. 3. 7. Răspunsul ecologic al unor specii de pajişti faţă grosimea stratului de litieră ( legenda ca în fig. 3.1.)
Ecological response of some grassland species to litter layer thickness (the legend as in fig. 3.1.)
Tabel 3.7. Gradul de suprapunere al nişelor (%) speciilor edificatoare în funcţie de
grosimea stratului de litieră (mm) (Media a; Media b – ca la tab. 3.1) Degree of niche overlap (%) of some grassland species depending on litter
layer thickness (mm) (Media a; Media b – as in tab. 3.1.)
Fes
trub
r
Agr
ocap
i
Cal
aaru
n
Cal
aepi
g
Ant
hvul
n
Bra
cpin
n
Des
ccae
s
Ave
npra
e
Nar
dstr
i
Sci
rsyl
v
Rum
ealp
i
Med
ia (
b)
Festrubr 71.9 15.8 23.1 6.2 26.1 17.2 8.8 45.9 18.1 12.7 24.6Agrocapi 100 12.3 28.5 8.6 32.4 17.4 7.7 47 17.8 17.7 28.9Calaarun 85.9 48.3 12 0.3 17.1 68.3 43.3 99.2 74.7 1.7 45.1Calaepig 100 88.8 9.6 4.1 87.9 25.9 7.6 85.2 26.3 30.2 46.6Anthvuln 100 100 0.9 15.1 28.2 26.5 5.1 66.7 25.1 81 44.9Bracpinn 100 89.1 12 77.7 6.7 27.7 7.6 97.2 28.1 32.2 47.8Desccaes 51.9 37.8 37.9 18.1 5 21.8 70.4 71.8 99.6 13.8 42.8Avenprae 12.3 7.8 11.2 2.5 0.4 2.8 32.7 20.7 47.5 0.9 13.9Nardstri 35.5 26.2 14.1 15.2 3.2 19.7 18.4 11.4 24.1 9.1 17.7Scirsilv 35.2 25 26.7 11.8 3 14.3 64.3 65.9 60.7 8.8 31.6Rumealpi 100 100 2.5 54.8 39.4 66.2 36 5.3 92.6 35.4 53.2Media(a) 72.1 59.5 14.3 25.9 7.69 31.7 33.4 23.3 68.7 39.7 20.8

140
3.2. FITODIVERSITATEA HABITATELOR DE PAJIŞTI DIN ZONA MONTANĂ Măsurarea biodiversităţii unei fitocenoze sau a unui ecosistem a devenit o temă
centrală a studiilor de ecologie din ultimul deceniu, oferind informaţii importante în ceea ce priveşte complexitatea, stabilitatea şi valoarea conservativă a sistemelor analizate.
Diversitatea ecologică poate fi văzută ca o măsură a informaţiei, ca varietatea mesajelor combinată cu numărul de indivizi care aparţin diferitelor specii (MARGALEF 1994) sau ca media rarităţii într-o comunitate (PATIL et TAILLIE 1982).
Analiza arborilor de regresie şi clasificare pentru numărul de specii, indicii de diversitate şi echitabilitate calculaţi pentru fiecare releveu, evidenţiază principalii factori de mediu care determină variaţia lor (BĂRBOS 2007).
Variaţia indicelui de diversitate Shannon (H’) este influenţată semnificativ de către variaţia grosimii stratului de litieră şi cea a concentraţiei de fosfor solubil din sol (Fig. 3.8).
Valoarea prag a grosimii stratului de litieră este de 17,5 mm. În cazul unui strat de litieră cu o grosime egală sau mai mare cu valoarea prag, valoarea medie a indicelui H’ este de 0,753. Dacă grosimea stratului de litieră este mai mică decât valoarea prag, un rol important în variaţia indicelui H’ îi revine concentraţiei fosforului solubil din sol, a cărui valoare prag este de 1,368 mg/l (pe scară logaritmică)
Astfel, la o concentraţie a fosforului egală sau mai mare decât valoarea prag se observă o influenţă negativă asupra valorii indicelui de diversitate Shannon, valoarea medie a acestuia fiind de doar 0,423. O concentraţie a fosforului solubil din sol sub valoarea prag favorizează instalarea unor fitocenoze cu o diversitate mai mare, valoarea medie a indicelui de diversitate H’ fiind de 1,61.
Deci, cu cât grosimea stratului de litieră şi concentraţia fosforului solubil din sol sunt mai mici cu atât diversitatea fitocenozelor este mai mare.
Cei doi factori analizaţi explică 63,46% din varianţa totală a valorii indicelui de diversitate Shannon, fapt ce sugerează implicarea, dar mult mai redusă, şi a altor factori în reglarea diversităţii fitocenozelor studiate.
Analiza arborilor de regresie pentru varianta exponenţială a indicelui de diversitate Shannon (N1) relevă faptul că, şi în acest caz, litiera este principalul factor care ifluenţează variaţia acestui indice, dar de această dată alături de variaţia pH-ului (Fig. 3.9.).
Valoarea prag a grosimii stratului de litieră este, de 16,66 mm, o valoare foarte apropiată de cea înregistrată în cazul indicelui de diversitate H’. În cazul în care grosimea stratului de litieră este egală sau mai mare decât valoarea prag, valoarea medie a indicelui de diversitate N1 este de 2,34.
Dacă grosimea stratului de litieră este sub valoarea prag, atunci un rol important în variaţia indicelui N1 îi revine pH-ului, a cărui valoare prag este de 5,22. La valori ale pH-ului mai mici decât valoarea prag, valoarea medie a variantei

141
exponenţiale a indicelui de diversitate Shanon este de 4,38, în timp ce la valori egale sau mai mari decât valoarea prag, valoarea medie a acestuia este de 6,47.
.
Fig. 3.8. Principalii factori care influenţează variaţia indicelui de diversitate Shannon (H’)
(P – concentraţia fosforului solubil din sol exprimată în mg/l pe scară logaritmică;
n – numărul de relevee prezente în fiecare nod terminal; r2 – coeficientul de determinare) Main factors that influence the variation in the Shannon diversity index (H’)
(P – concentration of soluble phosphorus in the soil expressed in mg/l on a logarithmic scale; n – number of releves present in each terminal node; r2 –
determination coefficient)
Deci, diversitatea cea mai mare se regăseşte la fitocenozele cu un strat de litieră a cărui grosime este mai mică de 16,66 mm şi cu un pH al solului mai mare de 5,22.
Variaţia grosimii stratului de litieră şi a pH-ului solului explică doar 58,23% din varianţa totală a indicelui de diversitate N1, fapt ce sugerează prezenţa şi a altor factori implicaţi în controlul diversităţii fitocenozelor, factori care fie au un rol mai puţin important, fie nu au fost analizaţi în studiul nostru.

142
Fig. 3.9. Principalii factori care influenţează variaţia indicelui de diversitate N1 (pH – aciditatea solului; n – numărul de relevee prezente în fiecare nod terminal;
r2 – coeficientul de determinare) Main factors that influence the variation in the exponential diversity index N1
(pH – soil acidity; n – number of releves present in each terminal node; r2 – determination coefficient)
Analizând variaţia numărului de specii/releveu (Fig. 3.10.), se poate observa că
principalii factorii răspunzători de oscilaţiile acestuia sunt grosimea stratului de litieră, modul de utilizare a pajiştilor şi pH-ul solului. În condiţiile în care grosimea stratului de litieră din fitocenoze este egală sau mai mare de 21,66 mm (valoarea prag), numărul mediu de specii/releveu este de 18,5.
Dacă grosimea stratului de litieră este mai mic decât valoarea prag, un rol important îi revine şi modului de utilizare a pajiştilor
Astfel, dacă pajiştile sunt utilizate ca păşuni pentru vite, oi sau păşuni mixte sau sunt târlite, numărul mediu de specii/releveu este de 21,7. În condiţiile în care pajiştile sunt utilizate ca fâneţe sau ca fâneţe păşunate primăvara şi toamna, pH-ul devine factorul principal care condiţionează numărul de specii din fitocenoze. Dacă pH-ul solului este mai mic de 5,134, numărul mediu de specii/releveu este de 31,8, iar dacă pH-ul are valori egale sau mai mari decât 5,134, numărul mediu de specii/releveu creşte la 39,3.
Grosimea stratului de litieră, modul de utilizare al pajiştilor şi pH-ul solului explică 58,60% din varianţa totală a numărului de specii/releveu.

143
Analizând figura 3.11 se poate constata faptul că principalii factori care influenţează variaţia indicelui de echitabilitate în cadrul fitocenozelor studiate sunt cantitatea de substanţă organică din sol (LOI) şi pH-ul solului.
În condiţiile în care cantitatea de substanţă organică din sol este mai mică de 14,06% (valoarea prag), valoarea medie a indicelui de echitabilitate este de 0,463.
Valoarea pH-ului solului influenţează variaţia indicelui de echitabilitate doar atunci când cantitatea de substanţă organică din sol este egală sau mai mare decât valoarea prag. La o valoare a pH-ului mai mică decât 5,15 (valoarea prag), valoarea medie a indicelui de echitabilitate este de 0,344, în timp ce la un pH ≥ 5,15, valoarea medie a indicelui de echitabilitate este de 0,44. Deoarece varianţa explicată de către cele două variabile de mediu este de doar 29,05%, putem considera că echitabilitatea fitocenozelor este condiţionată în primul rând de particularităţile biologice ale speciilor şi mai puţin de ceilalţi factori de mediu.
Fig. 3.10. Principalii factori care influenţează variaţia numărului de specii/releveu (past_c – utilizarea pajiştilor ca păşuni pentru vite; past_cs – utilizarea
pajiştilor ca păşuni mixte; past_s – utilizarea pajiştilor ca păşuni pentru oi; stbl – pajişti târlite; medw – fâneţe; mdw_p – fâneţe păşunate primăvara şi toamna; n –
numărul de relevee prezente în fiecare nod terminal; r2 – coeficientul de determinare) Main factors that influence the variation in species richness (mdw – hayfield; mdw_p – grazed hayfield; pst_c – cattle pasture; pst_cs – mixed pasture; pst_s – sheep pasture; stable – sheep pen pasture; pH – soil acidity; n – number of releves present in each terminal node; r2 – determination coefficient)

144
Fig. 3.11. Principalii factori care influenţează variaţia indicelui de echitabilitate Smith-Wilson
(LOI – cantitatea de substanţă organică din sol exprimată în procente; pH – reacţia solului; n – numărul de relevee prezente în fiecare nod terminal; r2 – coeficientul de determinare)
Main factors that influence the variation in the Smith-Wilson evenness index (LOI – soil organic matter; pH – soil acidity; n – number of releves present in
each terminal node; r2 – determination coefficient) În concluzie, rezultatele obţinute în urma analizelor de tipul celor folosite de
noi evidenţiază faptul că acumularea de litieră determină o scădere a bogăţiei specifice şi a biodiversităţii, fiind un factor fundamental care controlează structura, stabilitatea şi dinamica fitocenozelor, rolul său în organizarea fitocenozelor fiind mai important decât cel al circuitelor biogeochimice ale nutrienţilor, confirmând astfel rezultatele obţinute şi de alţi cercetători (MONK et GABRIELSON 1985; ECKSTEIN et DONATH 2005; CARSON et PETERSON 1990; GRIME 2001; FACELLI et PICKETT 1991; FACELLI et CARSON 1991).

145
3.3. AMENINŢĂRI POTENŢIALE Ca orice sistem suprapopulaţional de organizare a materiei vii şi pajiştile
realizează schimburi permanente de materie şi energie cu celelalte sisteme vii, căutând să-şi păstreze cât mai mult timp posibil o anumită stare de echilibru dinamic. Această stare de echilibru este temporară ea putând fi perturbată de modificări de amplitudini diferite ale valorilor diferiţilor factori ecologici. De aceea, un prim pas, necesar şi obligatoriu în activitatea de conservare, îl constituie identificarea potenţialilor factori perturbatori, cu alte cuvinte, identificarea ameninţărilor potenţiale care pot produce dezechilibre grave în structura, dinamica şi funcţionarea habitatelor.
Principalele ameninţări potenţiale la adresa stabilităţii pajiştilor din zona montană sunt:
1. Suprapăşunatul şi subpăşunatul, sunt activităţi care pot produce perturbări
majore în cadrul acestui tip de habitat, pajiştile, în general, şi acestea în mod special, sunt dependente de un păşunat raţional. Un management pastoral adecvat, realizat în condiţii normale în ceea ce priveşte numărul de animale/unitatea de suprafaţă, corelat cu o intensitate moderată a păşunatului, are efecte benefice asupra habitatelor de pajişti, efecte ce se manifestă la nivelul tuturor compartimentelor acestuia (structural, funcţional, dinamic). În condiţiile unui suprapăşunat, apar modificări structurale şi funcţionale majore, acestea manifestându-se prin reducerea bogăţiei specifice a pajiştilor şi invadarea acestora de către specii colonizatoare şi invazive. Aceasta se datorează, în primul rând, eliminării speciilor bune furajere, fapt ce permite dezvoltarea speciilor de buruieni, dar şi ca urmare a modificărilor la nivelul structurii solului datorită tasării şi implicit a scăderii aeraţiei solului dar şi datorită îmbogăţirii solului în fosfor şi azot din cauza dejecţiilor animale (eutrofizare).
Prin subpăşunat, speciile ruderale competitive şi cele invazive vor elimina speciile autohtone, determinând astfel o schimbare lentă, dar radicală a structurii habitatului.
2. Abandonul pajiştilor, reprezintă una dintre cele mai mari ameninţări la
adresa pajiştilor montane, mai ales în regiuni greu accesibile. Din punct de vedere ecologic, abandonul are aceleaşi efecte cu subpăşunatul, cu deosebirea că, în cazul abandonului, acestea se manifestă mult mai rapid. Dat fiind faptul că pajiştile din zona montană sunt, în majoritatea cazurilor, pajişti secundare care au apărut şi s-au menţinut datorită unui anumit mod de folosinţă, abandonul acestora determină iniţierea unui proces dinamic de succesiune spre tipul de comunităţi vegetale pe care aceste pajişti le-au înlocuit.
3. Împădurirea reprezintă un caz particular al efectelor subpăşunatului şi al
abandonului. În acest caz, succesiunea generată de un management defectuos duce la instalarea rapidă a unei vegetaţii lemnoase. În unele cazuri (terenuri improprii pentru agricultură), împădurirea se efectuează în mod conştient de către om, grăbindu-se astfel procesul de succesiune spre habitatea de păduri.

146
Pajişte suprapăşunată Overgrazed grassland
Stână de oi în Munţii Suhard Sheepfold in Suhard Mt.

147
Pajişte abandonată recent în Munţii Semenic
Abandoned meadow in Semenic Mt.
Pajişte invadată de vegetaţie lemnoasă în Munţii Rarău
Grassland invaded by trees in Rarău Mt.

148
Pajişte invadată de vegetaţie lemnoasă în Munţii Călimani
Grassland invaded by trees in Călimani Mt.
Aspectul unei pajişti după supratârlire invadata de Rumex alpinus The aspect of grassland after sheepfold was kept in one place for too long

149
4. Supratârlirea, reprezintă o altă ameninţare pentru conservarea habitatelor de pajişti montane, determinând schimbări majore ale principalilor factori edafici şi implicit iniţierea unor succesiuni secundare spre habitate ruderalizate, puternic eutrofizate. În zonele supratârlite se instalează şi domină o vegetaţie nitrofilă formată din Rumex alpinus, R. obtusifolius, Urtica dioica, Veratrum album, Colchicum autumnale şi altele.
5. Fertilizarea chimică sau organică (gunoi de grajd, compost, purin, etc.) în
doze mari are aceleaşi efecte ca şi supratârlirea. Prin fertilizare intensivă, efectul limitativ al nutrienţilor dispare iar speciile cresc mai mari, lumina devenind un factor limitativ.
Din punctul de vedere al compoziţiei chimice, tratamente chimice cu doze mari de azot aplicate 5 ani consecutiv au determinat acumularea unor cantităţi din ce în ce mai mari de azot total, funcţie de mărimea dozei de îngrăşământ. În cazul azotului proteic se observă aceeaşi evoluţie, creşterile însă fiind realizate prin valori mai mici ceea ce dovedeşte o neproporţionalitate între mărimea dozei de îngrăşământ şi a azotului proteic din plante, valori mai mari s-au înregistrat la azotul neproteic care la martorul netratat este de peste două ori mai mic decât la doza maximă de îngrăşământ.
Azotul nitric se găseşte în cantitate mare la tratamentele cu îngrăşăminte chimice, valoarea maximă (0,090%) înregistrându-se după 5 ani de experimentare la varianta fertilizată anual cu N250 kg/ha. Acesta este un indiciu al nemetabolizării azotului nitric, în sensul reducerii lui la amoniac şi antrenării sale în reacţiile biochimice. Proteina solubilă, cea mai accesibilă din punct de vedere metabolic rămânând la aceleaşi valori cu ale martorului nefertilizat la variantele fertilizate cu doze mai mari de N150 kg/ha. Aceasta dovedeşte că sinteza albuminelor şi globulinelor, proteide cu cea mai ridicată valoare alimentară, nu este favorizată de dozele mari de îngrăşăminte.
6. Turismul şi activităţile recreative constituie o ameninţare serioasă mai ales
în cazul pajiştilor de la altitudini mari care sunt atractive pentru activităţi recreative, cum ar fi sporturile de iarna. Deşi practicarea acestora nu afectează în mod direct covorul vegetal, totuşi, prin tasarea solului şi modificarea regimului de aeraţie şi hidric al solului se produc modificări mari în structura covorului ierbos.
De asemenea, prin tasarea zăpezii se întârzie topirea acesteia micşorându-se astfel perioada de vegetaţie a plantelor.
Omul, în mod voit sau nu, constituie o ameninţare permanentă la adresa stabilităţii şi conservării habitatelor de pajişti, contribuind în mod decisiv la degradarea acestora dacă acţionează în mod necorespunzător.
***
Datele prezentate în cadrul acestui material, aşa cum s-a subliniat şi în partea introductivă a cestui capitol, se referă doar la acele aspecte privind ecologia pajiştilor care am considerat noi că prezintă un anumit grad de noutate. Deoarece aceste date se bazează pe rezultatele unui număr mic de relevee floristice, aproximativ 200, care nu

150
au surprins toate habitatele de pajişti din zona montană a României, recomandăm ca acestea să fie privite doar cu titlu informativ.
Suntem conştienţi că nu am reuşit să oferim răspunsurile aşteptate probabil de multă lume, dar sperăm că am sugerat măcar o parte dintre mijloacele prin care aceste răspunsuri pot fi găsite.
De asemenea, sperăm că, pe baza datelor prezentate, am reuşit să subliniem faptul că, un plan de management sau de reconstrucţie ecologică a unor habitate degradate, trebuie să aibă la bază studii ecologice complexe, care să permită identificarea factorilor şi mecanismelor ce determină structura, funcţiile şi dinamica comunităţilor vegetale. Datele obţinute în urma derulării unor astfel de studii, oferă argumentele ştiinţifice necesare luării deciziilor privind măsurile de management necesare pentru conservarea habitatelor, decizii care sunt influenţate de factori politici, economici şi sociali.
În vederea elaborării unor planuri de management sau de reconstrucţie ecologică adecvate, ţinând cont de particularităţile staţionale şi structurale ale fiecărui tip de habitat, fără a subestima importanţa rolului pe care fiecare specie îl are într-o anumită fitocenoză, se recomandă analiza răspunsului ecologic al principalele specii edificatoare (monodominante sau codominante) al habitatelor de pajişti. Importanţa mare acordată speciilor dominante rezidă atât în faptul că realizează cea mai mare parte a fitomasei, cât şi prin influenţarea (în mod direct şi indirect) a identităţii, abundenţei şi distribuţiei locale a celorlalte specii (GRIME 1984; GRIME 1998; GIBSON et al. 1999).

151
IV. INVELIŞUL DE SOLURI ALE PAJIŞTILOR MONTANE Munţii Carpaţi prezintă pe toată întinderea sa o mare varietate a învelişului de
soluri. Având prezente în cadrul lor toate etajele bioclimatice de la pajiştile alpine la pădurile de foioase, o gama extraordinara de forme de relief de la cel de câmpie aluvioproluvială până la cel glaciar, şi un spectru foarte larg de roci eruptive, sedimentare şi metamorfic, se formează o mare diversitate pedologică.
În acord cu acest fapt, fiecărui etaj bioclimatic din Munţii Carpaţi le sunt proprii, atunci când nu se manifestă influenţele excesive ale rocilor, materialelor parentale de soluri sau ale apei anumite asociaţii zonale de soluri, după cum urmează:
etajului alpin al pajiştilor secundare alpine cu Carex curvula îi corespund soluri cu orizont A umbric semiturbos, acid şi profund (nigrisoluri), fără orizont spodic sau cu orizont spodic relativ slab exprimat (humosiosol).
etajului subalpin al jnepenişurilor, tufărişurilor de ericacce şi juniperetelor - îi sunt proprii spodosolurile cu orizont Bhs humico-feriiluvial şi orizont Es clar exprimat (podzoluri);
etajului boreal al pădurilor de molid - îi corespund soluri cu orizont Bs feriiluvial (spodosoluri), în general fără orizont Es-eluvial-podzolic bine exprimat (prepodzoluri şi podzoluri feriiluviale);
etajului nemoral al pădurilor de foioase - îi corespund, pe suprafeţele de teren plane şi slab înclinate, stabile, vechi, soluri cu orizont Bt argiloiluvial, în general luvisoluri (luvosoluri tipice şi albice), iar pe suprafeţele plane tinere sau pe cele în pantă cu bilanţ de denudare activ, soluri cu orizont B, cambic - de alterare (cambisoluri) debazificate şi acide (districambosoluri şi eutricambosoluri);
Zonalitatea solurilor poate fi perturbata prin efecte de extra şi intrazonalitate.
În cadrul unui etaj bioclimatic pot apărea - extrazonal - soluri caracteristice altui etaj bioclimatic, sub influenţa unor condiţii compensatoare de ordin climatic şi a unor asociaţii vegetale extrazonale. Astfel, în etajul pădurilor de foioase, pe versanţii umezi, umbroşi şi reci, mai ales pe substrat permeabil sub asociaţii de ericacce sau pâlcuri de molid, pot apărea spodosoluri, care se dezvolta în mod normal în etajul boreal sau pe versanţi însoriţi, calzi şi relativ uscaţi. Pot apărea sub asociaţii vegetale ierboase molisoluri (faeoziomuri) caracteristice condiţiilor de limita spre silvostepa.
Extrazonalitatea se manifestă în zonele montane, cu precădere în etajul pădurilor de foioase, unde poate apărea o gama larga de topoclimate în raport cu expoziţia versanţilor şi de asemenea o gama larga de asociaţii vegetale. În etajele superioare condiţiile climatice tot mai aspre determină variaţii mai puţin semnificative ale topoclimatelor şi o gamă tot mai mică de formaţiuni vegetale, fapt care împiedică diversificarea învelişului de soluri prin fenomene de extrazonalitate. (BĂLĂCEANU şi MARIAN, 1985).
Intrazonalitatea, concepută ca un fenomen în virtutea căruia într-un etaj bioclimatic pot apărea tipuri de soluri nespecifice, care se îndepărtează de la direcţia de evoluţie zonală, sub influenţa unor condiţii particulare de litologie,

152
hidrologie sau relief, se manifesta pretutindeni în munţi. Totuşi fenomenul este mai intens în etajul pădurilor de foioase, unde condiţiile climatice mai puţin drastice determina tendinţe mai slabe de debazificare şi acidifiere ce pot fi cu uşurinţa contracarate de alte direcţii de solificare imprimate de roci şi materiale parentale cu texturi sau alcătuiri chimico-mineralogice particulare şi unde exista o foarte mare diversitate de forme de relief şi condiţii hidrologice care, la rândul lor, pot influenţa procesele pedogenetice. În acest etaj apare o gama largă de soluri litomorfe, litohidromorfe şi hidromorfe (FLOREA şi MUNTEANU, 2003).
În celelalte etaje (boreal, subalpin, alpin), apariţia solurilor intrazonale se reduce simţitor. Pe de o parte datorită faptului că influenţele condiţiilor bioclimatice zonale în procesele de solificare sunt atât de puternice încât nu mai pot fi contracarate de cele ale rocilor decât în cazuri extreme (calcare şi parţial andezite), iar pe de alta parte pentru ca gama condiţiilor geomorfologice şi hidrologice ce ar putea influenţa solificarea se reduce considerabil (SÎRCU, 1971). Indiferent de caracterul lor zonal, extrazonal sau intrazonal, solurile montane prezintă o mare varietate de subtipuri, varietăţi şi variante în legătura cu caracterele secundare de hidromorfism, caracterul mai mult sau mai puţin scheletic, grosimea până la roca dură, gradul de eroziune etc. Este de remarcat faptul că subtipurile şi varietăţile gleizate şi stagnogleizate, sunt proprii zonei joase nemorale, iar apariţia caracterului scheletic este legat de substratul din roci dure, indiferent de etajul bioclimatic. Grosimea solurilor descreşte în linii mari odată cu înălţimea. Uneori, în special în zonele înalte, învelişul de sol poate lipsi, la suprafaţă apărând roca dură sau materiale reziduale sau de acumulare.
Învelişul de soluri este foarte diferit de la un sector la altul şi în unele situaţii, mai ales în etajul pădurilor de conifere şi în cel alpin, este dominat de asociaţii de soluri zonale. În alte situaţii însă, mai ales în etajul pădurilor de foioase, învelişul de soluri poate fi dominat de soluri intrazonale, extrazonale, azonale sau roci la zi şi, ca atare, zonalitatea este greu de sesizat. Oricum învelişul de soluri din regiunile montane este deosebit de neuniform, fiind legat de intensitatea eroziunii, caracterul scheletic al materialului parental şi de grosimea acestuia (NEACŞU şi colab., 1986).
4.1. SOLURILE DIN ETAJUL PAJIŞTILOR ALPINE În Carpaţii Româneşti, etajul alpin propriu-zis, al pajiştilor alpine
scunde, este cuprins intre 2000 (2300) şi 2544 m (punctul cel mai înalt), reprezentând de fapt limita de jos a acestui etaj, care în alţi munţi, cu condiţii c1imatice comparabile, se întinde până la aproximativ 2800 m.
Condiţiile geologice şi de relief sunt similare celor ale etajului subalpin, cu menţiunea că, în anumite masive (Făgăraş, Retezat), abundă vârfurile, crestele şi abrupturile stâncoase.
Clima este aspra, cu media anuala a temperaturilor <0°C şi precipitaţii peste 1300 mm anual; în mare parte sub forma de zăpada, şi cu vânturi frecvente şi puternice care, adeseori, provoacă deflaţie.

153
Vegetaţia lemnoasa se reduce la sălcii pitice (Salix reticulata, S. retusa, S herbacea), în general nesemnificative pentru pedogeneză şi la rhodorete de altitudine mare, iar cea ierboasa la specii de Carex curvula, Juncus trifidus, Oreachloa disticha, Festuca bucegiensis etc., care constituie asociaţii încheiate numai pe fragmente de platformă, culmi rotunjite şi unii versanţi mai slab înclinaţi. În proporţie mare, covorul vegetal este discontinuu, fiind afectat - ca şi solul - de eroziune.
Covorul ierbaceu este foarte bine dezvoltat fiind reprezentat prin: păruşcă (Festuca airoides), timoftică (Phleum alpinum), firuşorul (Poa alpina), păiuşul (Deschampsia caespitosa), coarnă (Carex curvula), ţepoşica (Nardus stricta), rugină (Juncus trifidus), clopoţel (Campanula alpina), degetăruşul (Soldanella pussila), piciorul cocoşului alpin (Ranunculus alpestris), brânduşa de primăvară (Crocus heuffelianus), degetăruţul (Soldanella montana), genţiana de zăpadă (Genţiana nivalis), cupele (Gentiana acaulis), clopoţei, (Campanula transsilvanica, C. alpina, C. cochlearifolia), ciuboţica-ochiul găinii (Primula minima), miliţeaua (Silene acaulis), ochiul şarpelui (Eritrichium nanum), floarea reginei (Leontopodium alpinum), muşchi (Polytrichum proliferum, Dicranum albicans), licheni (Cladonia rangiferina, Cetraria islandica, Certraria nivalis).
În arealele cu stâncării şi grohotişuri cresc periniţe de meliţea (Silene acaulis), păruşcă pitică (Festuca bucegiensis), macul pirenaic (Papaver pyrenaicum), floarea reginei (Leontopodium alpinum), iar în arealele mai umede şi în preajma turbăriilor apar pajişti cu salcie pitică (Salix herbacea- pe rocile silicioase şi Salix reticulata - pe rocile calcaroase), păruşcă (Festuca airoides), clopoţel (Campanula alpina), vulturică (Hieracium alpinum), ochiul găinii (Primula minima), coarnă (Carex curvula) (ROŞU, 1973).
Condiţiile c1imatice care inhiba humificarea şi formaţiile vegetale cu vocaţie acidifiantă determina formarea solurilor puternic humifere cu orizont Au - umbric, semiturbos (30-40% humus, slab încorporat părţii minerale) şi puternic acide (pH <4,5). Pe suprafeţele mai stabile se formează humosiosol cambic şi chiar criptopodzoluri litice cu orizonturi spodice evidente, care conduc la ideea unei vechi evoluţii a solurilor sub rhodorete, iar pe suprafeţele mai puţin stabile, humosiosol tipic şi litic, fără orizont B. În general solurile acestui etaj nu depaşesc stadiul litic, iar în cazuri foarte frecvente învelişul de soluri este dominat de litosoluri districe şi rocă la zi.
Efectele de intrazonalitate sunt slabe, numai calcarele şi dolomitele determinând apariţia solurilor rendzinoide acidificate în orizontul superior, cu asociaţii în care se întâlnesc Festuca amethystina, Sesleria haynaldiana, Festuca picta, Carex sempervirens, Festuca versicolor.
Indiferent de tip, subtip etc., solurile acestui etaj sunt sărace în elemente nutritive şi cu condiţii precare de humificare şi mineralizare a substanţelor organice.
Printre cele mai întâlnite soluri în acest etaj sunt humosiosolul şi litosolul.
Humosiosolurile (HS) (Sol humico-silicatic), sunt soluri specifice etajului alpin superior specifice Munţilor Carpaţi (Munţii Rodnei, Maramureşului, Bucegi,

154
Făgăraş, Parâng Retezat) ocupate de pajişti alpine în care domină asociaţii de Carex curvula.
Materialul parental este format din materiale dezagregate şi alterate provenind din roci silicatice necarbonatice (gresii, conglomerate cu liant silicios, şisturi cristaline, roci vulcanice), pe forme de relief mai puţin înclinate, platouri, coame largi. Clima este de tip montan cu Tma 1,20C. Pma 1350 mm (Staţia Bâlea).
Covorul ierbaceu este format de asociaţii de Carex curvula, Poa alpina, Festuca airoides, Primula minima, Soldanela pussila, Campanula alpina, Juncus trifidus etc.
Solurile prezintă un orizont A umbric (Au) cu crome de <2 la umed, dar conţinând materie organică humificată segregabilă de partea minerală silicatică, urmat de un orizont intermediar (AC, AR, B), grad de saturaţie în baze<53% şi culori cu crome şi valori <3,5 la umed. Orizontul A are la suprafaţa solului un strat gros de ţelină (Aţ) gros de 4-7 cm. La nivel de subtip se separă humosiosol: tipic, cambic, litic şi scheletic.
Humosiosolul tipic (HS ti) Se întâlneşte în zonele montane înalte ocupate de pajşti alpine, pe roci acide, cu
textură în general nisipo-lutoasă. Profilul de sol de pe platoul Caraiman- Bucegi (PUŞCARIU şi colab., 1956) prezintă un suborizont Aţ format din pâslă de rădăcini, cu material mineral adus de vânt, gros de 4-7 cm. Urmează un orizont Au de cca. 12-18 cm cu textură nisip-nisipo lutos, culoare brun cenuşiu foarte închis (10YR 3/2 la umed), astructurat, afânat. Urmează un orizont AR gros de 15 cm, nisipos cu fragmente de rocă, brun (10YR 4/3) şi rocă dură R relativ dezagregată la cca. 40 cm adâncime. Este un sol intens humifer pe tot profilul, conţine 20-30 % substanţă organică, cu 28,5 humus în Aţ, 7,0-14,9 % în Au, 1,0% în A/R. Conţine 0,9-1% azot total, raportul C:N>20. Capacitatea de schimb cationic este de 30-50 me/100 g sol în Au şi scade la 10 me/100 g sol în A/R. Reacţia solului este puternic acidă cu pH de 4-5, gradul de saturaţie în baze este foarte scăzut 10-12% în Au, putând ajunge la sub 4-5% pe solurile formate pe materiale puternic acide, dar poate să crească la 70-80% când solul se formează pe depozite bazice. Ca mod de folosinţă aceste terenuri sunt folosite ca pajişti pentru păşunatul oilor în perioada de vară (VASU şi colab., 1986).
Litosolurile (LS), sunt soluri incipiente din clasa protisoluri, având un orizont Ao sau O de cel puţin 5 cm grosime urmat din primi 20 cm de o rocă compactă continuă (Rn) sau de un material scheletic cu sub 10% pământ fin (Rp) sau orizont scheletic, cu sub 25% material fin (care poate continua până la peste 50 cm adâncime sau material (scheletic) calcaric cu peste 40% carbonat de calciu echivalent. Aceste soluri reprezintă un stadiu iniţial de formare a solurilor pe roci în general compacte în regiuni cu relief accidentat îndeosebi în regiunile de munte, dar şi de dealuri şi podişuri unde apar la zi de sub pătura de roci moi (argile, luturi, loessuri).
Formarea şi evoluţia lor este legată în principal de intensitatea fenomenului de eroziune cuplate cu fenomenele de dezagregare şi alterare a rocilor compacte. Litosolurile pot apărea atât în zonele reci şi umede din arealele montane din Carpaţi şi Subcarpaţi, cât şi în arealele mai joase cu roca dură aproape de suprafaţa solului (Dobrogea).
Apa freatică nu afectează profilul de sol, fiind situată la adâncimi mai mari de

155
10 m. La nivel de subtip se întâlnesc LS eutrice, districe, rendzinice, scheletice (prundice) şi histice.
Litosolurile eutrice (LS eu), sunt soluri bazice, formate pe seama dezagregării şi alterării rocilor dure formate în principal din calcare, conglomerate, gresii cu liant calcaros din zonele muntoase şi din Podişul Dobrogei.
Prezintă un orizont Aom sau Ao cu grosimi, care variază între 5-20 cm, textură luto-nisipoasă în primii 10 cm şi lutoasă în rest, culoarea variază de la neagră, până la brună (10YR2/1-5/3), structura este grăunţoasă-glomerulară, conţin o cantitate relativ mare de materie organică şi numeroase fragmente de rocă. LS pot avea şi un orizont de tranziţie A/R, cu mult schelet provenit din roca dezagregată, sau urmează direct un orizont R, format din roca dură, parţial dezagregată.
Litosolurile districe (LS di) sunt soluri acide, formate prin dezagregarea şi alterarea rocilor dure acide reprezentate prin şisturi cristaline, roci vulcanice, gresii silicioase. Ele se întâlnesc în general în zonele montane mai înalte, mai reci şi mai umede din Carpaţi şi Subcarpaţi.
În concluzie, învelişul de soluri este sărac în tipuri genetice, are soluri superficiale şi puternic afectate de eroziunea care scoate la zi stâncăriile. Mai mult decât în alte etaje problemele ameliorative ale solurilor trebuie judecate în contextul eficienţei, ţinând seama şi de randamentele posibile ale speciilor adaptate în condiţiile unei perioade de vegetaţie scurta.
În zonele mai înalte din etajul pădurilor şi pajiştilor alpine pe luncile râurilor pot fi întâlnite specii de ruşuliţă (Hieracium aurantiacum), iarba ciutei (Doronicum austriacum), lăptuca oaiei (Cicerbita alpina), salata iepurelui (Prenanthes purpurea), odogaciul (Saponaria officinalis), captalan (Petasites hybridum), călţunul doamnei (Geum rivale), ferigi ca ruginiţa (Asplenium ruta muraria), acul pământului (Asplenium trichomanes), Dryopteris filix-mas şi Polypodium vulgare (SIMIONESCU, 1961).
Pe rocile acide din Carpaţii Orientali şi Meridionali se găsesc specii de Saxifraga pedemontana, laptele stâncii (Androsace sp.), iarba surzilor (Saxifraga aizoon), Saxifraga carpatica, Semperivivum montanum, roşioară (Silene acaulis), floarea reginei (Leontopodium alpinum), Poa alpina ssp.vivipara, Ranunculus alpestris, Eritrichum nanum, Senecio abrotanifolius ssp.carpathicus, macul pirenaic (Papaver pyrenaicum).
Stâncăriile calcaroase adăpostesc asociaţii vegetale de Seseli rigidum, Saxifraga paniculata, Dianthus petraeus ssp. petraeus , Hieracium villosum, Festuca picta, F. versicolor, Festuca cinerea, Melica ciliata.
În momentul când pe stânci se formează un strat subţire de sol apar mici pajişti de Sesleria coerulans, Carex sempervirens. Tot pe stâncării se găsesc o serie de plante cu caracter endemic cum este Silene zawadskii din Carpaţii Orientali şi Silene saxifraga şi Primula auricola specii carpat-balcanice din munţii Banatului şi Apuseni (BĂLĂCEANU V., MARIAN ELISABETA, 1985).
Grohotişurile şi bolovănişurile care se formează în general la baza unui versant dar şi pe platourile montane sunt la început lipsite de vegetaţie după care încep să fie acoperite de licheni şi muşchi ca după ce se formează o pătură mică se sol să fie populate cu asociaţii de Rumex scutatus, Silene vulgaris, Valeriana montana,

156
Geranium robertianum, Gymnocarpium robutianum, iar în cazul când bolovănişurile sunt umbrite apar asociaţii de Lunaria rediviva, Arabis procurrens, Parietaria officinalis.
În cazul când grohotişurile sunt de origine calcaroasă şi sunt situate în etaşul alpin pe ele cresc Saxifraga moschata, Dryas octopetala, Galium anisophyllum, Papaver pyrenaicum (BOJOI, 2000).
4.2. SOLURILE DIN ETAJUL JNEPENIŞURILOR (SUBALPIN) În etajul jnepenişurilor, gama rocilor şi a tipurilor de soluri se lărgeşte şi
mai mult decât în etajul alpin. Rocile sunt exclusiv dure, chiar si în cazul celor sedimentare iar scoarţa
de alterare este superficială, depăşind numai în cazuri excepţionale 50 cm. Relieful este mai variat decât în etajul alpin datorita dezvoltării largi a
fenomenelor erozive. Clima este aspra, umeda şi rece (Tma cca 0°C, Pma 1200-1300 mm),
zăpezile sunt bogate şi îndelung stagnante în locurile adăpostite, iar vânturile sunt puternice.
Asprimea condiţiilor c1imatice determină instalarea în locul pădurilor a jnepenişurilor şi a altor asociaţii de plante lemnoase scunde, mai ales din ericacce. Pajiştile secundare sunt constituite mai ales din asociaţii de Festuca airoides, Agrostis rupestris cu Deschampsia flexuosa (cele mai tipice pentru etaj) şi din asociaţii de Nardus stricta insinuate pe vâlcele. Având habitus mic şi înrădăcinare superficială speciile ierboase amintite nu modifica substanţial solurile formate sub speciile lemnoase.
Pe calcare şi stâncării se întâlnesc asociaţii cu specii calcifile sau pioniere, fără importanţa în orientarea pedogenezei (NEACŞU şi colab, 1986).
Topoclimatele nu determina apariţia extrazonală a unor asociaţii vegetale ci numai distribuiri între asociaţiile din etaj, în funcţie de umiditate, vântuire şi temperatura.
Pajiştile sunt bogate, deoarece pe suprafeţe întinse arborii şi tufărişurile au fost defrişate în principal prin ardere fiind constituite din: firuţă (Poa media), ţepoşică (Nardus stricta), iarba vântului (Agrostis rupestris), păruşcă (Festuca airoides), cinci degete (Potentila aurea ssp. chrysocraspeda), cimbrişor (Thymus pulegioides), ghinţură (Genţiana clusii, Gentiana nivalis), silur (Euphrasia minima).
Pe versanţii calcaroşi cu expoziţie sudică apar specii ierboase doritoare de căldură că: păruşcă (Festuca saxatilis), rogoz (Carex sempervirens).
În locurile mai umede pe unele văi şi depresiuni apar specii de arin de munte (Alnus viridis), măcriş (Rumex arifolius), timoftică (Phleum alpinum), târsă (Deschamsia caespitosa) (ŢUCRA şi NEACŞU, 1985).
În apropierea turbăriilor cu Sphagnum şi Eryophorum şi arealelor mlăştinoase cresc specii de rogoz (Carex curta, C.dacica).
Condiţiile climatice şi de vegetaţie determina cea mai intensă debazificare şi acidifiere, solurile zonale ajungând la valori ale gradului de saturaţie în baze de 1-4% şi la valori pH<4-4,5. În aceste condiţii, predomina

157
pe toate rocile, cu excepţia calcarelor şi dolomitelor masive, solurile din clasa spodosolurilor, extinderea cea mai mare având-o podzolul litic (frecvent cu orizontul Bhs dezvoltat în roca subiacenta, acumularea sescvioxizilor şi humusului făcându-se pe fisuri). Cele mai puternice podzoluri se dezvoltă sub jnepenişuri şi loiselcuriete. Ele se menţin însa şi sub asociaţii de Festuca airoides şi Agrostis rupestris, care nu au capacitatea de a modifica solul moştenit
Gama spodosolurilor este relativ mare, în acest etaj întâlnindu-se pe lângă podzoluri, şi nanopodzoluri (cu orizonturi subcentrimetrice foarte nete), podzoluri brune alpine cu orizont Es difuz etc.
Intrazonalitatea este foarte slab exprimata. Aşa cum s-a arătat, chiar pe conglomeratele calcaroase sau pe gresiile cu ciment calcaros se dezvoltă spodosoluri. Numai pe calcare şi dolomite apar rendzine şi mai ales soluri rendzinice, uneori cu turbificare la suprafaţă (turba uscata).
Cele mai întâlnite soluri în acest etaj sunt podzolurile. Podzolurile sunt soluri puternic acide caracteristice subetajului subalpin al
jneapănului şi subetajului molidului (BARBU, 1976). Ca procese pedogenetice se acumulează puternic humusul brut puternic acid,
silicaţii sunt puternic alteraţi şi migrează sub forma unor produşi de alterare (complexe humico-minerale) care se acumulează într-un orizont iluvial (Bhs şi Bs).
Sunt întâlnite în principal în Carpaţii Meridionali ( Munţii Bucegi, Făgăraş, Cibin, Parâng, Sebeş, Retezat şi Godeanu). În Carpaţii Orientalii sunt întâlniţi în Munţii Rodnei şi Maramureşului iar în Munţii Apuseni apar insular la limita pădurii şi în subetajul subalpin sub jnepenişuri.
Clima este umedă şi rece cu Tma de 2-50C, cu temperaturi de 12-140C vara în iulie şi -5 la -70C iarna în ianuarie. Cantitatea de Pma variază cu altitudinea între 800-1400 mm. Materialul parental este format din alterarea rocilor acide silicioase formate din gresii, conglomerate cu ciment silicios, şisturi cristaline.
Podzolurile au un orizont O sau A ocric sau umbric (Ao, Au) urmat de un orizont eluvial spodic Es şi un orizont B spodic humico-feriiluvial sau feriiluvial (Bhs, Bs). Pot prezenta la suprafaţa solului un orizont organic nehidromorf O (folic) mai gros de 50 cm şi proprietăţi criostagnice.
La nivel de subtip de sol au fost deosebite podzoluri: tipice, umbrice, feriiluvice, histice, criostagnice, litice şi scheltice.
Podzoluri tipice (PD)-(podzol humico feriiluvial) Au ca principala caracteristică prezenţa orizontului Bhs de migrare a
humusului. Aceste soluri sau format în condiţii de relief montan pe areale cu drenaj mai
slab, situate pe platforme, coame şi depresiuni, pe depozite acide. Orizontul superior Ao sau Au la podzolurile umbrice, este închis la culoare cu structură grăunţoasă slab exprimată, restul profilului fiind nestructurat, urmează un orizont Es puternic sărăcit în materie organică şi sescvioxizi şă îmbogăţiţi în silice, albicios, gros de 5-20 cm. Orizontul Bhs de acumulare a sescvioxizilor şi a humusului, gros de 30-70 cm, brun-ruginiu, după care urmează roca dură R sau afânată C.
Profilul de sol din căldarea glaciară a Văii Bâlea, lângă lacul Bâlea (Păunescu şi Roşu, 1979) este situat la baza versantului pe un tăpşan de acumulare, pantă de 5-

158
100 , la 1700 m altitudine. Tma 1,80C, Pma 1200 mm (Staţia Bâlea). Vegetaţia este formată din păşune în subetajul jnepenişurilor cu asociaţii ierbacee de Festuca rubra, Poa annua, Agrosis capillaris, A. rupestris, Soldanella major, Potentilla ternata. Materialul parental este format din depozite coluviale de şisturi clorito-filitoase. Din punct de vedere morfologic profilul prezintă un suborizont organic O de 3 cm cu pâslă de rădăcini de plante ierboase cu foarte puţin material mineral. Urmează un orizont Ao de 3cm cu multă materie organică, nisipo-lutos, cu 25% pietriş, culoare neagră în stare umedă (10YR2/1), glomerular foarte mic-poliedric mic, friabil, rădăcini frecvente. Urmează un orizont Ea cu grosime variabilă de-a lungul profilului până la 18 cm, textură nisipoasă, şi 50% pietriş, culoare cenuşie (10YR 6/1), nestructurat, foarte friabil, slab coeziv în stare uscată, rădăcini frecvente. Orizontul Bhs între 18-25 cm, are textură nisipoasă, 60% pietre şi pietriş, culoare neagră la umed (5YR 2/1), nestructurat, friabil, urmează în continuare un orizont Bs între 25-48 cm, nisipos, cu 75% pietriş şi pietre, culoare brun închisă la partea superioară şi brun gălbui la partea inferioară (7,5YR 3/2-10 YR 5/4, nestructurat, şi roca formată din pietriş şi pietre şi material mineral.
Podzolurile au textură mijlociu-grosieră, nediferenţiată pe profil, au proprietăţi fizice, hidrofizice şi de aeraţie puţin favorabile.
Sunt soluri puţin profunde, cu schelet foarte mult, volum edafic redus, drenaj rapid, capacitate de apă utilă foarte mică.
Bine asigurate cu humus 36,8 % în suborizontul O, şi A, 8,79% în Ea, 24,3 în Bhs, 5-15% în Bs. Raportul C:N este 23 în O şi Ao, 18 în Ea şi Bhs. N total 1,1% în O şi Ao şi 0,3% în Ea, 0,9 în Bhs şi 0,5% în Bs.
Foarte puternic acid cu ph 4,3 în Ao, 4,1 în Ea, 4,3 în Bhs şi 4,6 în Bs. Conţinut extrem de mic în baze de schimb iar gradul de saturaţie în baze foarte mic (5%).
Datorită condiţiilor vitrege de climă şi relief aceste soluri se folosesc ca păşuni şi fâneţe cu productivitate slabă (BĂLĂCEANU şi MARIAN, 1985).
Datorită instabilităţii reliefului crionival, eroziunea capătă uneori forme acute, determinând extinderea extraordinara a litosolurilor şi stâncăriilor reziduale pe creste si versanţi si acumulative pe conuri şi pânze de grohotişuri la baza abrupturilor
Limitările de ordin agrochimic sunt foarte mari, dar, din punctul de vedere al producţiei, limitările cele mai mari se referă la volumul edafic şi la integritatea învelişului de sol în ansamblu. Productivitatea ecosistemelor de pajişti din acest etaj depinde însa foarte mult şi de condiţia c1imatica şi potenţialul biologic al speciilor (NEACŞU şi colab, 1986).
4.3. SOLURILE DIN ETAJUL MOLIDIŞURILOR (BOREAL) Condiţiile de solificare. În etajul pădurilor de molid, diversitatea condiţiilor
fizico-geografice, de sol şi de vegetaţie este mult mai mare decât în cel al jnepenişurilor.
În acest etaj, dispar sau ocupa suprafeţe restrânse rocile moi şi afânate cum ar fi argilele moi, marnele moi, lehmurile, depozitele eoliene, nisipurile, depozitele de terasa şi evantaie de împrăştiere etc.
Gama formelor de relief se restrânge şi ea prin dispariţia reliefului acumulativ

159
de mari proporţii şi restrângerea celui acumulativo-eroziv. Dispar luncile largi, sistemele de terase cu poduri plane, dispar conurile de împrăştiere şi glacisurile de mari proporţii. Domina relieful de eroziune, versanţii şi culmile mai mult sau mai puţin înguste. Relieful aşezat este dat numai de resturile vechilor nivele de eroziune.
Apele freatice îşi pierd din importanta, influenţând solificarea numai local, în tinoave.
Condiţiile climatice sunt mai aspre decât în etajul nemoral (Tma 1.5°C, Pma 900-1200 mm), determinând un excedent mai important de umiditate în sol.
Datorita climei mai severe, s-a dezvoltat masiv pădurea de molid pretutindeni, indiferent de expoziţie, modificările topoclimatice fiind insuficient de drastice pentru a provoca pătrunderea extrazonală a speciilor de foioase în masive semnificative pentru pedogeneză. Apariţia diseminata a paltinului de munte şi scoruşului sau chiar a pâlcurilor de făgete de altitudine este neînsemnată din acest punct de vedere. Pe expoziţiile nordice în schimb, în luminişurile pădurilor sau în pajiştile secundare, au pătruns adeseori specii din etajele superioare, semnificaţia cea mai mare pentru pedogeneză.
Asociaţiile vegetale din pajişti cunosc şi ele o diversitate mai mare. Domina cele mezofile de Festuca rubra şi Nardus stricta şi, în legătură cu
calcarele, apar asociaţii cu plante calcifile sau termofile. Asociaţiile cu plante xerofile şi higrofile îşi pierd importanţa. Efectele instalării vegetaţiei secundare, mai ales a asociaţiilor vechi de Festuca şi Nardus, sunt însa mult mai clar exprimate în pedogeneză. decât în etajul fagului (BORZA, 1960).
În subetajul de amestec de molid cu brad covorul ierbaceu din pădurile de conifere şi mai ales din poienile din pădure sunt reprezentate de: păiuş roşu (Festuca rubra), păiuşcă (Agrostis capillaris), rogoz (Carex ovalis), târsa (Deschampsia caespitosa), horşti (Luzula luzuloides), clopoţei (Campanula patula ssp. abietina), sunătoare (Hypericum maculatum), crinul de munte (Lilium martagon), oiţe (Pulsatilla alba), trestia de pădure (Calamagrostis arundinacea), măcrişul iepurelui (Oxalis acetossela), ferigi (Polypodium vulgare, Athyrium filix-femina), licheni reprezentaţi de barba-ursului (Isnea barbata), ciuperci foarte numeroase: mânătârca sau hribii (Boleotus edulis), râşcovul (Lactarius deliciosus), pâinişoarele (Russula integra) toate comestibile iar flocuşeii (Lactarius torminosa) şi Rossula emetica sunt ciuperci otrăvitoare (SIMIONESCU, 1961).
Procesele de solificare sunt mult mai puţin influenţate de condiţiile climatice decât în etajul jnepenişurilor. Temperatura scăzută condiţionează o activitate biologica slabă în sol. Datorita acestui fapt şi datorita constituţiei chimice a materiei organice furnizate de molid, în sol se acumulează o cantitate însemnata de humus de tip moder sau moder-humus brut, puternic acid, între ai cărui constituenţi predomina acizii fulvici mobili şi agresivi. Solificarea decurge sub influenţa fulvoacizilor în sensul debazificării şi acidifierii puternice, până la valori V <20-25% şi la valori pH <4,7 (4,0-4,5), la care nu mai este posibila argilizarea mineralelor primare decât ca faze intermediare către destrucţia acestora şi eliberarea de oxizi de fier şi aluminiu. Sub influenţa excesului de apa şi a curentului percolant format de acesta, sescvioxizii complexaţi de materia organica mobili sunt antrenaţi în profunzime şi depuşi într-un orizont Bs feriiluvial sau Bhs humico-feriiluvial (spodic). S-au format în acest fel, pe

160
plan zonal, în proporţie de masă, spodosoluri. Procesul de podzolire menţionat, deşi pregnant prin formarea orizontului B
spodic, nu este totuşi, în limitele acestui etaj, suficient de intens pentru a provoca spălarea totală din orizontul mineral de suprafaţa a sescvioxizilor şi humusului şi dezvoltarea neta a unui orizont eluvial Es albicios, ca în cazul podzolurilor propriu-zise. EI se dezvolta numai până la stadii intermediare şi determina existenţa zonala prepodzolurilor tipice (spodosoluri fără orizont Es alb). În etajul respectiv se întâlnesc, ca termeni iniţiali ai evoluţiei câtre solul brun feriiluvial, criptopodzol tipic care sunt lipsite de orizont Bs exprimat morfologic, dar prezintă acumulare de Al2O3 la nivelul orizontului B, şi ca termeni hiperevoluaţi către podzol tipic, caracterizate prin realizarea unui orizont Es foarte slab exprimat morfologic, discontinuu, vizibil clar numai la solul în stare uscata.
Sub pajiştile secundare recente, podzolurile feriiluvice se regăsesc morfologic ca atare, căpătând doar caracteristicile specifice înţelenirii în orizontul A. Sub pajiştile secundare vechi insă, mai ales sub cele de Festuca rubra, se produc în soluri modificări puternice, induse mai ales de modificarea regimului hidrotermic, a naturii humusului şi de masa radiculara puţina ceste modifidri pot merge până la completa estompare morfologica a orizontului spodic, ce mai poate fi însa decelat analitic prin identificarea sescvioxizilor preexistenti. Se formează în acest fel soluri criptospodice tipice ca o creaţie a pajiştilor, care ar putea fi confundate morfologic cu districambosolurile. Aceste soluri, care pot fi considerate zonale, se regăsesc pe suprafeţe însemnate în subetajul boreal (la limita de sus a pădurii).
În general podzolurile feriiluvice, ca şi cele criptospodice tipice sunt scurte, litice sau semilitice şi scheletice, având volum edafic mic. Sub raport agrochimic, ele sunt, fără excepţie, deficitare la toate capitolele.
În etajul boreal, extrazonalitatea se manifesta mai ales prin dezvoltarea podzolurilor tipice pe roci hiperacide, sub influenţa vaccinietelor sau a excesului de apa sub sphagnete, cazuri deja amintite.
Intrazonalitatea se manifesta mai rar şi mai slab decât în etajul nemoral, întrucât pe de o parte Condiţiile climatice mai aspre diminuează rolul rocilor în determinarea sensului pedogenezei, iar pe de alta parte, pentru ca, în acest etaj, dispare o gamă larga de roci moi capabile sa devieze sensul evoluţiei prin crearea de regimuri hidrice specifice etc. Totuşi calcarele, marnele, dolomitele determina apariţia eutricambosolurilor şi a rendzinelor, iar sub influenta andezitelor bazaltoide şi microdioritelor, apar soluri andice şi chiar andosoluri, cu tot cortegiul lor de probleme agrochimice menţionat anterior. De amintit, în sfârşit, existente histosolurilor districe (turbării acide sau înalte) determinate de ape strânse în microdepresiuni (BĂLĂCEANU şi MARIAN, 1985).
Eroziunea care acţionează şi ea, mai ales în domeniul pajiştilor, determina existenţa litosolurilor, a solurilor trunchiate şi a rocilor la zi, în proporţii mai reduse decât în etajul nemoral, în care umanizarea mai puternica devine adesea cauza principala a eroziunii, şi decât în etajul alpin cu covor vegetal mai slab şi cu relief glaciar şi crionival.
Printre principalele soluri în acest areal vegetal se întâlnesc, podzolurile care coboară din arealul jnepenişurilor, prepodzolurile, criptopodzolurile, andosolurile, iar

161
pe areale cu stâncărie aproape de suprafaţa solului litosolurile şi rendzinele. Prepodzolurile (EP) (Sol brun feriiluvial) se întâlnesc în regiunile montane
înalte în subetajul pădurilor de molid, uneori în amestec de fag cu molid şi cu covor de ericacee.
Aceste soluri se formează pe roci acide formate din gresii silicioase, conglomerate, şisturi crisataline şi roci eruptive. Clima este umedă şi rece cu Tma 3-40C şi Pma >1000 mm.
Prepodzolurile au un orizont ocric sau umbric (Ao, Au) urmat de un orizont B spodic feriiluvial (Bs). Poate apărea un orizont de tranziţie Es discontinuu şi un orizont organic nehidromorf O (folic) în general la suprafaţa solului.. La nivel de suptip de sol deosebim prepodzoluri: tipice, umbrice, histice, litice, scheletice.
Prepodzoluri tipice (EPti) Au orizontul Ao (ohric) se întâlnesc în zonele montane sub păduri de molid cu
Luzula silvatica. Profilul de sol de lângă Hotelul Alpin, la altitudinea de 1400 m (N. Florea şi colab., 1968), Folosinţa pădure de molid, vegetaţia este formată de molidişuri cu Luzula sylvatica, Oxalis acetosella, Rubus idaeus, Polygonatum verticillatum etc.
Prezintă la suprafaţă un suborizont O gros de 3 cm cu humus moder de culoare negru spre brun negricios şi uneori din humus brut gros de 5-6 cm cu pâslă de rădăcini de plante ierboase. Urmează un orizont Ao de 15 cm, cu iviri în puncte de orizont Es cu textură nisipo-lutoasă culoare brun închis-negricios la umed (7,5YR3/2), astructurat, conţine grăunciori de cuarţ, fără pelicule de humus sau oxizi de fier cu pete mici cenuşii difuze friabil, rădăcini frecvente. Orizontul Bs gros de 30-80 cm, nisipo-lutos, brun - ruginiu (5YR 3/2), şi brun roşcat - gălbui spre bază (5YR 6/4-6), astructurat. În bază este roca mamă sub adâncimea de 40-80 cm. Scheletul apare pe tot profilul şi creşte spre bază.
Acest sol prezintă un conţinut ridicat de humus 25% în orizontul (O, 9,2%) în Ao+Es şi 1.7-2,3% în Bs. Raportul acizi huminici/acizi fulvici este de 0,3-0,7, conţinutul de azot total ridicat (0,39-1,0%) iar raportul C:N depăşeşte valoarea de 20.
Capacitatea de schimb cationic este de 30-40 me/100 g sol în orizontul A+Es şi scade spre adâncime. Reacţia solului este puternic acidă cu ph 4,2 în O. 4,6 în Ao+Es şi 5,1 în Bs.
Gradul de saturaţie în baze este coborât cu 10-45% în A şi 20-25% în Bs. Se remarcă o migrare a sescvioxizilor din orizontul A şi acumularea lor în Bs
Criptopodzolurile (CP). (Sol brun acid criptopodzolic) sunt întâlnite la contactul dintre districambisoluri (soluri brune acide) şi prepodzoluri în zonele montane înalte în etajul pădurilor de conifere şi de amestec de fag cu răşinoase.
Aceste soluri poate prezenta un orizont organic O şi un orizont A foarte humifer sau numai unul dintre ele, urmat de un orizont B criptospodic (Bcp) humifer. Uneori poate prezenta şi un orizont organic nehidromorf O (folic) sub 50 cm grosime.
La nivel de subtip, pot fi întâlnite criptopodzoluri tipice, histice, litice, scheletice.
Solul criptopodzolic tipic(CP ti) Este întâlnit în zonele montane şi subcarpatice înalte la contactul între pădurile
de conifere cu cele de fag..

162
Profilul de sol situat la 3 km nord de Cascada Bâlea (PĂUNESCU, ROŞU, Ghidul excursiilor, 1979) este situat pe un versant înclinat cu pantă de 150 şi altitudine absolută de 1100 m, în subetajul bioclimatic de amestec de fag cu răşinoase. Tma este de 5.50C iar Pma 950 mm. Vegetaţia este formată de brădete şi făgete cu floră de mull: Lamiastrum galeobdolon, Oxalis acetosella, Galium odoratum, Pulmonaria rubra (CĂLINESCU şi colab., 1969).
Materialul parental este format din depozite deluviale de şisturi clorito-sericitose. Prezintă o textură luto nisipoasă pe tot profilul de sol. Culoarea variază de la brun cenuşiu foarte închis la umed (10 YR 3/2) În orizontul Ao sao Au, la brun gălbui la umed (10YR 5/6) în orizontul Bs. Structura este glomerular mediu în Ao şi poliedric subangular mic şi foarte mic în Bs. Cantitatea de schelet creşte cu adâncimea (!00 cm) până în suborizontul BR când atinge 70% din total.
Este un sol semischeletic, volum edafic mijlociu, drenaj normal, capacitate de apă utilă mijlocie, intens humifer la suprafaţă cu 22,3-32,2% în orizonturile O (de 2cm) şi Au (5 cm grosime), 7,3% în Bs, cu humus mull-moder, puternic acid cu 4,4 în Au şi 4,3 în Bs, conţinut mijlociu de baze schimbabile la suprafaţa solului datorită biopacumulării şi extrem de mic în restul profilului. Aceste soluri sunt cu o productivitate mijlocie pentru fag şi brad şi superioară pentru molid (ŢUCRA şi NEACŞU, 1985).
Andisoluri (AND) (Andosoluri) se găsesc în perimetrele montane vulcanice sau unde sau depus cenuşile vulcanice, în special în Carpaţii Orientali în lanţul vulcanic Oaş, Gutâi, Ţibleş, Căliman, Gurghiu, Hărghita şi în Munţii Apuseni.
Andisolurile se dezvoltă atât pe andezite, în special pe cele bazaltoide, pe microdiorite şi alte roci bazice din familia dioritelor, ca efect al predominării substanţelor amorfe, inclusiv de tip allofanic, între materialele de alterare ale acestor roci, cât şi pe cenuşi vulcanice, piatra ponce şi alte derivate vulcanice de diferite compoziţii. Alterarea rapidă a materialului vulcanic poros determină acumularea de complexe amorfe şi prezenţa de minerale incomplet cristalizate ca allofanul şi imogolitul. Morfologic, andosolurile sunt caracterizate printr-un orizont vitric sau andic.
Orizontul vitric este un orizont dominant de sticlă vulcanică, în timp ce orizontul andic cuprinde orizonturi de sol constituite în principal din allofane în care predomină complexele humico-aluminice.
Andisolurile prezintă un orizont A (Am, Au, Ao) urmat de orizont intermediar (AC, AR, Bv) la care se asociază proprietăţi andice pe cel puţin 30 cm grosime începând din primii 25 cm ai solului mineral. Poate avea un orizont organic O sa T. Dintre acestea va fi prezentat un andosul umbric litic din Caldeira Luci din Munţii Harghita, la 8 km sud-vest de Sâncrăieni (V. PEREPELIŢĂ, M. SPIRESCU, Ghidul Excursiilor, 1979).
Profilul pedologic este situat pe un versant cu pantă de 100, altitudine de 1270 m, expoziţie nordică. Climatul este montan cu Tma 3,50C, Pma 900 mm.
Vegetaţia este de pădure de molid cu pajişte de Oxalis acetosella, Agrostis capillaris, Calamagrostis arundinacea, Campanula patula ssp. abietina, Luzula luzuloides.

163
Materialul parental este format de deluviu din andezite cu piroxeni. Prezintă la suprafaţa solului o pâslă de rădăcini O de 3 cm, un orizont Au de 35 cm cu textură lut nisipoasă în partea superioară şi lut nisipos-lut spre baza orizontului, culoare brun foarte închis la umed (10 YR 2/2), structură glomerular mic-mediu, friabil, slab coeziv la uscat, schelet format din pietre şi bolovani mai abundent spre baza orizontului (40-60%). Urmează un suborizont BR gros de 18-20 cm, cu textură lut-lut nisipos cu schelet 75%, culoare brun închis (10 YR 3/3, structură poliedric subangular, friabil. Sub 52 cm urmează orizontul RB format din 85% pietre şi bolovani printre care se găseşte material mineral, luto-nisipos, culpare brun închis la umed (10YR 3/3).
Andosolul umbric litic este mijlociu profund, scheletic-excesiv scheletic, volum edafic mic, drenaj raopid, capacitate de apă utilă mică. Sol intens şi profund humifer cu moder (18,6-25,6%) în orizontul Au, 11,6% în BR şi 10,0 în RB, puternic acid cu ph 4,7-4,8 în Au, 4,8 în BR şi 4,9 ân RB., conţinut extrem de mic de baze de schimb. Gradul de saturaţie în baze este extrem de mic pe tot profilul de sol V% 1,9-2,7. (R. LĂCĂTUŞU şi colab. Ghidul excursiilor, 1979). Sunt mijlociu favorabile pentru molid şi larice, şi slab favorabile pentru fag (GĂRBACEA,1956).
Aceste soluri sunt dominate morfologic de acumularea puternica de humus într-un orizont A închis la culoare, debazificat, acid (orizont Au-umbric) şi pot fi, ca şi rendzinele, tipice (fara orizont B), cambice (cu orizont B cambic) sau litice (mai subţiri de 50 cm). Se pare ca dezvoltarea lor este optima la limita de sus a subetajului pădurilor de fag, unde condiţiile climatice sunt cele mai propice tipului de alterare menţionat. Comparativ cu districambosolurile în etajul cărora se dezvoltă, andosolurile sunt mai puţin debazificate (opun o rezistenta mai mare la debazificare), au capacitate de schimb cationic (dependentă de pH) mai mare. Datorita conţinutului ridicat de hidroxizi de aluminiu şi fier amorfi, aceste soluri au o mare capacitate de fixare, în forme greu solubile şi greu asimilabile, a ionilor fosfat, borat şi molibdat, fapt care poate induce carenţe de nutritie la plante şi animale. Pe unele roci bazaltice, în legătură cu proporţia mare de silicaţi feromagnezieni, poate apărea o cantitate mare de magneziu şi chiar predominarea ionului Mg
2+ asupra celui de Ca2+ în complexul de schimb al acestor soluri, respectiv un dezechilibru al raportului Ca2+/Mg
2+ şi al nutriţiei vegetale şi animale. Ca şi în cazul districambosolurilor la pH<5, apare şi la aceste soluri Al3+ de schimb în proporţii toxice pentru plante (ŢUCRA şi NEACŞU, 1985).
Litosolurile eutrice (LS eu) Sunt soluri bazice, formate pe seama dezagregării şi alterării rocilor dure
formale în principal din calcare, conglomerate, gresii cu liant calcaros din zonele muntoase şi din Podişul Dobrogei. Prezintă un orizont Aom sau Ao cu grosimi, care variază între 5-20 cm, textură luto-nisipoasă în primii 10 cm şi lutoasă în rest, culoarea variază de la neagră, până la brună (10YR2/1-5/3), structura este grăunţoasă-glomerulară, conţin o cantitate relativ mare de materie organică şi numeroase fragmente de rocă. LS pot avea şi un orizont de tranziţie A/R, cu mult schelet provenit din roca dezagregată, sau urmează direct un orizont R, format din roca dură, parţial dezagregată.

164
LS eutrice din Podişul Dobrogean s-au format în urma proceselor de eroziune, care au îndepărtat materialul mai fin, în general loesoid, de la suprafaţa solului şi au permis apariţia, mai aproape de suprafaţă, a rocilor dure în stadii diferite de dezagregare. În zonele mai înalte, situate în Munţii Măcinului, LS s-a format prin alterarea în situ a rocii dure, dar şi printr-un aport eolian, pe care s-a aşezat vegetaţie ierboasă şi lemnoasă, alături de muşchi şi ciuperci, prin descompunerea cărora s-a acumulat materia organică într-un orizont Ao subţire.
Vegetaţia naturală a litosolurilor dobrogene este formată din vegetaţie lemnoasă şi ierboasă, respectiv de pădure mezofilă cu Quercus petraea, Carpinus betulus, Acer campestre, Prunus mahaleb, în zonele mai înalte, şi vegetaţie de stepă cu specii de Poa bulbosa, Artemisia austriaca, Euphorbia nicaeensis şi Festuca valesiaca, în zonele mai joase, iar în zona montană carpatică covorul vegetal corespunde cu treapta altitudinală de vegetaţie.
Nu are carbonaţi de calciu, solul având o reacţie slab acidă în Aom (6.3) şi A/R (6.2). Au un conţinut mijlociu în humus în Aom (4.48 %) şi A/R (3.43 %). În cazul preluării în cultură conţinutul în humus scade. Conţinutul în N total este mijlociu în Aom (0.25%) şi A/R (0.20%), iar conţinutul în fosfor (P2O5 %) este de 0.12 %. Raportul C:N este 12.0 în Aom şi 10.0 în A/R. T este mică în Aom (19.43 me/100g sol) şi mijlocie în A/R (22.40 me/100g sol), iar V este mezobazic în Aom (74.9%) şi eubazică în A/R (77.4%), la sud-est de Greci, Dobrogea de Nord (FLOREA ş.a., 1968; ANA CONEA, 1970).
Litosolurile districe (LS di) Sunt soluri acide, asmănătoare cu litosolurile eutrice, formate prin
dezagregarea şi alterarea rocilor dure acide reprezentate prin şisturi cristaline, roci vulcanice, gresii silicioase. Ele se întâlnesc în general în zonele montane mai înalte, mai reci şi mai umede din Carpaţi şi Subcarpaţi.
Litosolurile redzinice (LS rz) Reprezintă o altă categorie de LS, specifice Podişului Dobrogei, dar şi zonelor
înalte carpatice sunt asemănătoare cu cele tipice, dar în care orizontul R este format din calcare, roci metamorfice şi eruptive bazice şi ultrabazice, care prin alterare nu formează materiale amorfe. Prezintă o textură luto-nisipoasă, conţinut mare de carbonaţi (12.8-13.3%), o reacţie slab alcalină (pH = 8.0), sunt mijlociu aprovizionate cu humus în Ao (4.5%) şi A/Rrz (3.6%) şi un conţinut mare în N total în Ao (0.30%) şi A/Rrz (0.20%), la Osmancea, Dobrogea de Sud (C. GRIGORAŞ, 1999).
Litosolurile prundice (LS qq) Sunt acele litosoluri în care roca dură o reprezintă pietrişurile. Aceste soluri se
pot întâlnii pe trenele de pietrişuri montane şi piemontane. Litosolurile histice (LS tb) Sunt acel soluri care în orizontul de suprafaţă au o mai mare cantitate de
materie organică mai puţin alterată, care poate ajunge la stadiul de turbă. Deşi sunt relativ bine aprovizionate cu humus LS, nu pot fi valorificate în
cultură, datorită cantităţii mari de schelet, a rocii dure situate la mică adâncime şi uneori a pantei mari şi sunt folosite ca păşuni foarte slabe.
Rendzinele (RZ), se găsesc pe suprafeţe relativ reduse în ţara nostră fiind

165
răspândite în special în zona montană în condiţii de climă umedă ale zonei forestiere în arealul pădurilor de gorun, fag şi fag în amestec cu conifere. În Carpaţii Occidentali se găsesc în munţii: Bihor, Gilău, Trascău, Codru. Pădurea Craiului, Poiana Ruscă, în Carpaţii Orientali: munţii Ciucului, Hăghimaşului, iar în Carpaţii Meridionali: munţii Bucegi, Lotrului, Sebeşului, Vâlcan. Local se mai găsesc în Subcarpaţii Getici şi în Dobrogea (BLEAHU şi RUSU, 1965). Rendzinele apar pe reliefuri fragmentate, culmi înguste şi versanţi cu înclinări şi expoziţii diferite, pe roci compacte bogate în calcar, ca efect al eliberării unor cantităţi importante de Ca2+
,
se dezvoltă, atât sub păduri cât şi sub pajişti. Rendzine, soluri humifere de culoare închisă, cu humus calcic, stabil
(molisoluri litomorfe), cu complexul argilo-humic saturat în proporţie de 80-100%, cu pH neutru slab alcalin şi aprovizionare relativ buna cu elemente nutritive. Ele pot fi tipice (fără orizont B), cambice (cu orizont Bv de alterare), litice (cu profil scurt de 25-50 cm) sau semilitice (cu profil de 50-100 cm grosime). Restricţia principală a acestor soluri în legătură cu fertilitatea este data de volumul edafic mic
Marnele determina, prin acelaşi mecanism al eliberării Ca2+, apariţia fostelor pseudorendzine în vechile clasificări, în prezent, variante ale rendzinelor pe roci carbonatice moi, cu însuşiri similare, dar volum edafic mai mare.
Ele se caracterizează prin prezenţa unui orizont Am, cu crome <3, un orizont Arz şi Bv (cambic), cu culori având cel puţin la partea superioară valori şi crome de <3.5 la umed şi <5.5 în stare uscată şi un orizont Rrz în primii 150 cm. Caracteristica dominantă este prezenţa orizontului diagnostic Rrz rendzinic bogat în schelet dar şi a orizontului Am, închis la culoare, bogat în humus, cu structură bine dezvoltată glomerulară. Treptat rendzinele pot evolua spre solurile zonale după apariţia unui orizont Bv cambic. La nivel de subtip se întâlnesc RZ calcarice, eutrice, cambice şi scheletice.Profilul de sol este de tipul Am-AR-Rrz. Textura variază de la mijlocie la fină, fiind uniformă pe profil.
Rendzinele din zona montană (Culoarul Branului, CONEA, 1968) din punct de vedere morfologic prezintă un orizont Am, argilos, de culoare neagră-brun foarte închisă în stare umedă (10YR 2/1-2), structură grăunţoasă, bine dezvoltată, fragmente de rocă, face efervescenţă pe fragmentele de rocă (tratat cu HCl). Urmează un orizont de tranziţie spre rocă (AR), argilos, culoare mai deschisă sau asemănătoare cu partea superioară, fragmente dese de rocă alterată. Urmează orizontul Rrz care reprezintă roca dură de multe ori dezagregată (BĂLĂCEANU şi MARIAN, 1985).
Deşi au profilul scurt şi scheletul calcaros poate apărea chiar din orizontul Am aceste soluri sunt bine structurate, poroase, cu aeraţie relativ favorabilă. Sunt soluri foarte bogate în humus (>10%), rezervă foarte mare de humus 200-300 t/ha, cu humus de tipul mull calcic. Sunt soluri saturate în baze sau foarte slab debazificate (V 70-100%), capacitatea de schimb cationic (T) în orizontul A este de 50-70 me/100 g sol, sunt soluri neutre-slab alcaline şi uneori slab acide în arizontul Am (5,5-7,5), şi ating 7,0-8,0 în orizontul AR, sunt bine aprovizionate cu substanţe nutritive, raportul C:N este de 12,7, iar N total este de 0,92-1,19%, dar greu mobilizabile în zona umedă. Pentru a se obţine recolte mai mari se recomandă îngrăşăminte chimice (NPK) şi organice în cantităţi mari (IONESCU SISEŞTI şi COCULESCU, 1939).

166
4.4. SOLURILE DIN ETAJUL FOIOASELOR (NEMORAL) În etajul pădurilor de foioase se întâlneşte cea mai bogata gama de condiţii
fizico-geografice şi de asociaţii vegetale din spaţiul carpatic şi, drept urmare, cel mai larg spectru de soluri.
În acest etaj se întâlnesc roci de durităţi foarte diferite, de la argilele şi marnele moi, până la gnaisuri şi cuarţite, roci cu toate constituţiile mineralogice şi chimice cunoscute, de la cele u1trabazice - carbonatice, până la cele hiperacide, în sfârşit, o mare varietate de roci sub raportul originii (aluvii, deluvii, coluvii, proluvii).
Relieful nu este nici el mai puţin complex, atât ca geneza cât şi ca forma, vârstă etc. Se întâlnesc reliefuri fragmentate erozive, reliefuri acumulative de câmpii piemontane nefragmentate, reliefuri acumulativo-erozive piemontane diferit fragmentate, platforme etc. Nu lipsesc nici reliefurile carstice şi nici măgurile vulcanice. La toate acestea trebuie adăugată diversificarea extraordinară provocata de fenomenele de pantă, acumulări în lunci etc.
Apele freatice îşi manifesta influenţa asupra solificării în sistemele de lunci, la capetele conurilor de împrăştiere şi uneori pe pante prin infiltraţii laterale, iar apele de stagnare intervin pe reliefurile plate cu depozite fine.
La modul general, clima este moderat răcoroase (Tma 6-70 C, în subetajul gorunetelor, 60 C în subetajul făgetelor de a1titudine mai mare) şi moderat umeda (Pma 700-1000 mm). La nivelul suprafeţelor orizontale sau slab înc1inate, cu drenabilitate mai slaba, se înregistrează în sol un excedent de umiditate anual, faţa de evapotranspiraţia potenţiala, semnificativ pentru înregistrarea unei tendinţe de evoluţie zonala. În limitele reliefului fragmentat, care este predominant, în relaţie cu înclinarea şi expoziţia versanţilor şi cu gradul de acoperire cu vegetaţie de păduri sau pajişti, se realizează topoclimate locale de la cele calde, relativ uscate, comparabile cu cele din antestepa, cu bilanţ deficitar al apei în sol, favorabile humificării şi mineralizării materiei organice, până la cele reci, umede, cu bilanţ puternic excedentar al apei din sol, nefavorabile humificării şi mineralizării materiei organice, asemănătoare celor din etajele montane superioare.
Multitudinea condiţiilor naturale amintite provoacă diversificarea covorului vegetal, exprimata prin apariţia extrazonală unor asociaţii vegetale din alte etaje bioclimatice, influenţând şi ele de o manieră specifică, prin calitatea humusului furnizat, procesul de solificare.
În limitele etajului, pădurile de gorun ocupa sectoarele joase, intrând în munţi sub forma unor enclave pe golurile depresionare, iar fagul ocupa versanţii umbriţi si sectoarele mai înalte de 600-700 m. Asociaţiile ierboase din plante mezofile influenţează solurile prin natura humusului, tipul de înrădăcinare, distribuţia materiei organice moarte, schimbarea regimului hidrotermic al solurilor etc., aducând uneori modificări notabile în sensul evoluţiei genetice.
Pe versanţii nordici, umbriţi, umezi, reci, coboară extrazonal din etajul superior molidul şi ericaceele, în asociaţii cu mare putere acidifiantă asupra solului, În timp ce pe versanţii expuşi insolaţiei pot apărea asociaţii cu plante termofile şi xerofile, cu putere slaba de influenţare a solificării. Învelişul ierbos este reprezentat în ochiurile de pajişti din pădure prin rogoz (Carex

167
pilosa), fiuţa de pădure (Poa angustifolia), obsiga (Brachipodium sylvaticum), mărgeluşa (Melica uniflora), horştiile (Luzula luzuloides) pe solurile mai sărace şi mai acide, vinariţa (Galium odoratum), urzica moarta galbenă (Lamiastrum galeobdolon), laptele câinelui (Euphorbia amygdaloides), golomăţul (Dactylis glomerata ssp. aschersoniana ) pe solurile mai fertile şi păiuşca (Agrostis capillaris), sadina (Chrysopogon gryllus) pe solurile mai umede din vestul ţării. (ROŞU, 1973).
Sub influenţa excesului de umiditate de natura freatica se dezvolta asociaţii de plante higrofile ce pot provoca turbificări în orizonturile superioare ale solurilor şi chiar pot determina formarea solurilor organice hidromorfe, iar sub influenţa excesului de apa stagnantă, asociaţii higrofile şi higrofil-acidifile specifice, cu Nardus stricta, Molinia caerulea, Deschampsia caespitosa, specii de Juncus etc.
Prezenţa carbonatului de calciu determină apariţia pajiştilor secundare cu specii calcifil-termofile, provocând dezvoltarea orizonturilor humifere, molice, cu humus saturat, stabil. În cadrul etajului pădurilor de fag din cadrul pădurilor de foioase se găsesc pajişti secundare în care învelişul ierbos este relativ bine dezvoltat fiind reprezentat prin specii de: firuţa de pădure (Poa nemoralis), mărgeluţa (Melica uniflora), păiuş (Festuca altissima), păiuş roşu (Festuca nigrescens), vioreaua (Scilla bifolia), ceapa ciorii (Gagea lutea), ghiocel (Galanthus nivalis), brebenei (Corydalis cava), ferigă (Dryopteria filis mas), rodul pământului (Arum maculatum), urzica moartă galbenă (Lamiastrum galeobdolon).
Etajul pădurilor de fag la rândul său în funcţie de anumite aspecte climatice şi altitudinale prezintă şi un subetaj al pădurilor de amestec de răşinoase şi fag. Aici învelişul ierbaceu este mai puţin bogat ca în pădurile de până acum, învelişul de sol fiind mai sărac în humus calcic şi mai acid, fiind reprezentat prin: leurdă (Allium ursinum), urzica moartă galbenă (Lamiastrum galeobdolon), laptele cucului (Euphorbia cyparissias) pe solurile mai bogate cu humus cu mull şi măcrişul iepurelui (Oxalis acetosella), hurşti (Luzula luzuloides), păiuş de pădure (Festuca altissima), păiuş roşu (Festuca nigrescens), afini pe solurile mai acide şi mai scheletice.
În relaţie cu multitudinea extraordinara a condiţiilor naturale, învelişul de soluri din etajul nemoral prezintă cea mai mare diversitate de tipuri, subtipuri, familii şi varietăţi de soluri din toate zonele şi etajele bioclimatice ale ţarii.
Aşa cum s-a arătat mai înainte, procesul general de formare şi evoluţie a solurilor, desfăşurat sub influenţa condiţiilor bioclimatice zonale (bilanţ uşor excedentar al apei în sol şi ca atare regim hidrologie percolativ profund, condiţii moderate-bune de humificare şi mineralizare a materiei organice) are ca tendinţă generală debazificarea si acidifierea. Pe reliefurile relativ stabile şi vechi, pe materiale parentale fine-mijlocii, fenomenele amintite sunt însoţite de argiloiluviere şi formare a orizonturilor B texturale şi a orizonturilor eluviale El sau Ea (argiluvisoluri), iar pe versanţi şi culmi înguste, precum şi pe terasele şi evantaiele de împrăştiere recente, orizonturi de alterare, translocare şi acumulare de argilă, cu doar

168
cel mult cu orientări locale ale argilei (orizonturi cambice-cambisoluri) (PARASCHIV, 1965).
Luvisolurile pot fi, în raport cu vârsta, înclinarea şi gradul de stabilitate a reliefului, preluvosoluri şi luvosoluri tipice şi albice uneori chiar glossice cu degradare podzolica. În cadrul lor se pot întâlni subtipuri slab, mediu sau puternic pseudogleizate, cu menţiunea ca sub pajiştile secundare se înregistrează întotdeauna o înrăutăţire a regimului hidric, solurile fiind, prin comparaţie, mai intens pseudogleizate decât cele de sub păduri.
Pe versanţi, sub influenţa proceselor de denudare, se pot întâlni preluvosoluri şi luvosoluri tipice fără orizonturi eluviale, ce pot fi considerate ca stadii întinerite prin eroziune.
Prin înţelenire secundara, regimul lor hidric se înrăutăţeşte şi, odată cu aceasta, se schimbă nu numai profilul solului care devine tot mai intens pseudogleizat, ci şi asociaţiile vegetale pe care le suportă, trecâdu-se de la pajiştile cu Agrostis capillaris, Festuca rubra, Cynosurus cristatus etc., la cele cu Deschamphsia caespitosa, Agrostis canina, Nardus stricta, Juncus conglomeratus, Molinia coerulea, care degradează valoarea pajiştilor.
Aprovizionarea lor cu substanţe nutritive este în general deficitara, cu atât mai mult cu cât sunt mai acide şi mai debazificate. Pe substratele provenite din cristalin pot prezenta şi carenţe de microelemente.
Solurile specifice pădurilor cu frunză căzătoare sunt foarte variate. Se întâlnesc prepodzolurile din etajul molidului, nigrisolurile, cambisolurile, luvisoluri, alosolurile, litosoluri şi regosoluri, rendzine şi în arealele depresionare, hidrisoluri, histosoluri, azonal faeoziomurile şi intrazonal aluviosoluri districe şi eutrice
Cambisolurile soluri cu orizont B cambic - de alterare - putând avea sau nu orizonturi A humifere umbrice sau molice, relativ scurte, reprezintă o clasă de soluri care ocupă suprafeţe relativ mai reduse pe teritoriul ţării noastre întâlnindu-se în special în zonele mai înalte carpatice, subcarpatice, de podiş şi dealuri înalte. La nivel de tip de sol cambisolurile sunt formate din:
Eutricambosolurile (EC) (brune eumezobazice) larg răspândit în mai muIte etaje bioclimatice în relaţie cu un substrat fin, saturat, frecvent carbonatic, considerat ca tip intrazonal, litomorf.
Sunt soluri având orizont A ocric sau molic (Ao, Am) urmat de orizont intermediar cambic (Bv) cu valori şi crome peste 3,5 (la umed) cel puţin pe feţele agregatelor structurale începând din partea superioară; proprietăţi eutrice în ambele orizonturi. Nu prezintă orizont Cca în primii 80 cm. Pot prezenta orizont O şi orizont vertic sau pelic şi proprietăţi stagnice, gleice şi andice. Au profilul de sol de tipul Ao-Bv-C. au o textură mijlociu-grosieră până la fină dar de obicei lutoasă, fără diferenţiere texturală pe profil. Structura este grăunţoasă-poliedric mic moderat dezvoltată în Ao poliedric angular mediu-prismatică moderat-bine dezvoltat. Ele apar în dependenţa de substraturi fine cu carbonaţi sau cel puţin puternic saturate, cu calciu dominant în complexul adsorbtiv (argile, lehmuri, argile reziduale din calcare, argile marnoase, marne etc.), sau de substraturi mai puţin fine, dar cu schelet calcaros (proluvii calcaroase). Sunt soluri tinere, unele ca vârstă absolută, fiind formate pe reliefuri acumulative recente, în cadrul depresiunilor intramontane (evantaie de

169
împrăştiere şi terase holocene), altele ca vârstă relativa, fiind condiţionate de eroziune şi alunecări şi reprezentând fie vechi soluri trunchiate până în orizontul B, reintrate într-un nou ciclu de solificare, fie un stadiu de solificare relativ evoluat al unui regolit scos la zi prin eroziune sau alunecări (SECELEANU, 1982, 2004).
Se întâlnesc atât sub păduri cât şi sub pajişti secundare, fără ca asociaţiile vegetale de pajişte sa le schimbe semnificativ morfologia. Pot fi ochrice (cu acumulare slaba-moderata de humus intr-un orizont Ao-ochric, deschis la culoare) sau molice. Pe argilele sedimentare montmoril1onitice pot avea caractere vertice (argiloase, gonflate în perioadele umede, cu crăpături mari, în cele uscate), iar pe argilele reziduale sau pe proluviile calcaroase au caractere rendzinice. Frecvent prezintă pelicule de argila şi se asociază cu soluri brune argiloiluviale, având tendinţa de evoluţie către acestea.
Ele sunt formate pe roci bogate în calciu; luturi, argile, marne, calcare, conglomerate şi gresii. Apa freatică nu influenţează în general profilul de sol fiind situată la adâncimi >5m, drenajul fiind în general bun. Proprietăţile fizice şi hidrofizice sunt relativ bune, condiţionate de textură şi adâncimea apei freatice. Conţinutul de humus este diferit dar se situează în jur de 2-4%, rezerva de humus fiind de 65-125t/ha. Reacţia solului este slab acidă-neutră, gradul de saturaţie în baze variază între 55-90%, sunt bine aprovizionate cu substanţe nutritive.
Districambosoluri-DC (Soluri brune acide), sunt cambisoluri care ocupă zonele montane şi de dealuri respectiv Dealurilor Subcarpatice şi Munţii Carpaţi. Caracteristica acestor soluri este existenţa orizontului diagnostic Bv şi gradul de saturaţie în baze V<55%.
Spre deosebire de solurile brune eu-mezobazice, acestea sunt formate pe un material parental în general mai acid, şisturi cristaline, roci eruptive dar şi luturi, nisipuri, gresii, conglomerate, acidifiate prin procesul de pedogeneză.
Profilul de sol este de tipul Ao-Bv-C sau R. Textura este uniformă pe profil, şi poate fi de la grosieră până la mijlocie. Structura solului este grăunţoasă-poliedric mic subangular slab dezvoltat în orizontul Ao şi poliedric angular-subangular mediu slab dezvoltat în Bv. Proprietăţile fizice sunt relativ favorabile iar cele de fertilitate slabe. Ele ocupa primul loc ca frecvenţa şi extindere între solurile etajului nemoral, dezvoltându-se pretutindeni unde apar roci silicatice intermediare şi acide, dure, şi materiale parentale derivate din acestea sau acolo unde apar roci sedimentare afânate, uşor percolabile. Se întâlnesc atât pe reliefuri acumulative tinere (terase, conuri de împrăştiere holocene), dar şi pe reliefurile vechi, cu bilanţ de denudare activ, care menţine solurile într-un stadiu de tinereţe relativa. Ele au texturi variate, de la grosiere la mijlocii (mai rar mijlociu fine), şi frecvent prezintă caracter scheletic şi caracter litic sau semilitic (grosime 20-50 cm sau, respectiv, 50-100 cm), intensitatea caracterului respectiv fiind determinata de panta reliefului, substrat, eroziune etc.
În funcţie de alcătuirea granulometrică şi panta, ele au în cele mai multe cazuri un drenaj global asigurat şi, în general, nu prezintă fenomene de stagnare a apei în profil. Totuşi, în legătura cu alternanţele de straturi de gresii şi argile, pot apărea uneori, pe reliefuri cu alunecări soluri brune acide cu influenţe de apă laterale.

170
Pe reliefurile joase de câmpii proluviale, districambosolurile prundice (dezvoltate pe prundişuri) pot suferi influenţe freatice propriu-zise, clare şi semnificative pentru pajişti (VELCEA, 2001).
În relaţie cu asociaţiile vegetale sub care se dezvolta, cu natura chimico-mineralogica a substratului şi drenajul intern etc., districambosolurile prezintă diverse grade de saturaţie în baze, pH, aprovizionare cu elemente nutritive şi alte particularităţi care le pot diferenţia la nivel de varietate şi chiar subtip. Pe substrate din roci acide şi intermediare, sub păduri de fag şi chiar gorun, se întâlnesc districambosoluri tipice sau litice, cu mull, oligomezobazice (V = 30-50%), cu pH în jur de 4,8-5,2, adeseori cu Al3+ schimbabil în cantităţi apreciabile (6-10 me), fără acumulare de sescvioxizi în orizontul B. Pe substrate hiperacide şi unele roci acide cu cuarţ, uşor percolabile, sub paduri de fag cu Luzula sp. şi sub asociaţii de ericacce (Bruckenthalia spiculifolia, Calluna vulgaris , Vaccinium sp.) se dezvolta soluri brune acide cu moder, oligobazice şi distrioligobazice (V <30%), puternic acide (pH 4,5-4,8), cu acumulare iluvială de Al203 (soluri brune acide criptospodice). În cele mai multe cazuri aceste soluri au orizont A humifer acid (umbric), atât sub pădure cât şi sub pajişte. Pe rocile bazice şi ultrabazice fără carbonaţi, şi uneori chiar pe roci intermediare din familia dioritelor sau pe depozite de fliş, se întâlnesc, până la altitudini de 900-1000 m, soluri brune acide de limită. inferioara, oligomezobazice şi mezobazice (V =50-60%) cu profil tipic de sol brun acid, nediferenţiat textural, fără pelicule argiloase. Ele constituie capul de serie în categoria solurilor acide şi nu au afinităţi genetice cu eutricambosolurile brune eubazice (eumezobazice) sau cu cele argiloiluviale. Un caz aparte îl constituie districambosolurile formate pe andezite, microdiorite etc., care aparţin adeseori subtipului andic, prezentând particularităţi mineralogice (proporţie mare de substanţe amorfe de tip allofan) şi chimice (capacitate mare de reţinere fosfatica etc.), determinate de tipul de alterare (MUNTEANU, 1988, 1998).
Districambosolurile sub pajişti secundare prezintă particularităţi morfologice legate de înţelenire şi au un regim hidrotermic mai favorabil decât cele de sub pădure. În rest, pajiştea secundara conservă ca atare districambosolurile.
Aprovizionarea acestor soluri cu elemente nutritive este cu atât mai deficitara cu cât ele sunt mai debazificate şi mai acide. Cele mai acide au Al3+ de schimb în cantităţi toxice, iar cele pe cristalin şi cele cu caracter andic pot avea carente de microelemente. Ca şi la alte soluri, caracteristicile chimice şi agrochimice ale acestor soluri sunt variabile în cadrul aceluiaşi tip, subtip etc. în diversele provincii geochimice ale Carpaţilor. Aşa cum s-a arătat anterior, în acest etaj este posibilă şi frecventă apariţia extrazonalităţii unor formaţiuni vegetale specifice altor zone, ca efect al topoclimatelor locale de pe povârnişurile umbrite şi reci sau al celor însorite şi calde. Este frecventa mai ales apariţia extrazonală a molidişurilor şi vaccinietelor, asociaţii cu mare potential acidifiant care, pe substraturi hiperacide, dau naştere la spodosoluri, în special podzolurile feriiluvice.
În limitele acestui etaj, diversificarea cea mai importantă a solurilor este determinată de efectele de intrazonalitate create de influenţa excesiva în pedogeneza a rocilor ultrabazice mai ales carbonatice, a celor cu texturi extreme sau cu constituţii mineralogice particulare, precum şi de excesul de apa sau efetul eroziunii.

171
Sunt în mod obişnuit eubazice (grad de saturaţie în baze V > 75%, frecvent 80-90%), dar atunci când provin prin eroziune din vechi luvisoluri sau când sunt mai puternic evoluate câtre acestea, pot fi şi mezobazice (frecvent cu V> 60).
Prezintă un conţinut de humus scăzut format în special din acizi fulvici cu grad de saturaţie în baze mic, acesta împreună cu o cantitate de materie organică parţial humificată în special în zona de pădure atinge de la 4-20%, rezerva de materie organică este foarte mare 250-300t /ha. Gradul de saturaţie în baze este scăzut şi poate atinge V 35%, iar pH <5, sunt slab aprovizionate cu substanţe nutritive. sunt slab favorabile pentru culturile agricole şi sunt folosite ca pajişti şi folosinţă silvică. (PUIU, 1980). În cadrul solurilor montane, se situează între cele mai bine aprovizionate cu elemente nutritive. Ridica mai mult probleme de conservare şi combatere a eroziunii decât probleme agrochimice (TEACI, 1980).
Hidrisolurile reprezintă o clasă de soluri intrazonale formate din cauza unui exces permanent sau temporar de umiditate. Aşezarea lor este determinată de condiţii locale de rocă şi relief care favorizează prezenţa apei în exces, fie datorită pânzei freatice situate la mică adâncime în cazul solurilor hidromorfe de natură freatică, fie la suprafaţa solului prin stagnarea apei din precipitaţii în cazul solurilor superficial stagnante.
Hidrisolurile sunt prin excelenta soluri de pajişti, generând asociaţii cu specii higrofile şi fiind în acelaşi timp influenţate de acestea. Dominant apar ciperacee, juncacee, graminee higrofile, ranunculacee, typhacee etc., în general specii puţin valoroase. Hidrisolurile sunt formate în arealele montane pe de o parte pe terenuri joase, slab drenate, lunci, terase inferioare şi depresiuni şi pe de altă parte pe suprafeţe plane mai înalte acoperite cu depozite argiloase din cuprinsul zonei umede.
Din punct de vedere morfologic hidrisolurile se caracterizează prin existenţa unui orizont de glei tipic la baza profilui de sol sau de pseudoglei în imediata apropiere a suprafeţei solului.
Excesul de umiditate produce în ambele cazuri, datorită condiţiilor de anaerobioză, procese de reducere, ce au ca efect, apariţia în sol a compuşilor reduşi de Fe şi Mn şi a unor culori caracteristice vineţii, cenuşii şi negricioase.
Hidrisolurile apar în proporţii semnificative numai în depresiunile intramontane, la capătul conurilor de împrăştiere, în zona de efilare a apelor freatice şi la contactul diferitelor trepte din sistemele luncilor. În spaţiul montan propriu-zis, influentele freatice se manifesta semnificativ pe suprafeţe mici, numai în tinoave.
În funcţie de gradul de oxigenare a apelor, de natura şi cantitatea cationilor conţinuţi, de viteza de curgere etc., apele freatice determina direcţii diferite de evoluţie în procesele pedogenetice. De asemenea, în funcţie de asociaţiile vegetale şi condiţiile de humificare, pot apărea tipuri diferite de soluri hidromorfe.
În raport cu conţinutul de materie organică, cu gradul ei de humificare şi cu profunzimea acumulării, în depresiunile intramontane se întâlnesc hidrisoluri de tipul gleiosolului distric şi cernic şi limnisoluri de tipul histosolurilor eutrice şi districe. Toate aceste tipuri genetice au apa freatica la adâncimi mai mici de 1,25 m şi în proportie de 90% sunt acoperite de fâneţe.
Hidrisolurile cuprind, în raport cu gradul de acumulare a humusului şi cu saturaţia în baze, următoarele subtipuri şi varietăţi:

172
gleiosolurile (deschise la culoare, cu orizont de humus relativ slab dezvoltat) care, în raport cu saturatia în baze, sunt districe (V <55%), eutrice (V> 55%) şi carbonatice (V 100% cu CaC03); Gleiosolurile sunt soluri având orizont O şi /sau orizont A (Am, Ao,Au) şi proprietăţi gleice (orizont Gr) care apar în profil din primii 50 cm ai solului mineral. gleiosolurile eutrice (cu orizont Ao relativ bogat în materie organică, deschis la
culoare). gleiosolurile molice (cu orizont A molic - bogat în humus, închis la
culoare, cu V> 55%) ; gleiosolurile umbrice - idem, cu V <55%; gleiosolurile histice - cu orizont T de turba<50 cm grosime.
Gleiosol cernic (lăcovişte) - soluri profund humifere, cu V> 55%, pot fi tipice, cambice, carbonatice etc.
Atât solurile gleice cât şi lacoviştile sunt considerate mlăştinoase când au apa la <50 cm adâncime.
Gleiosolurile eutrice Vegetaţia naturală este formată din asociaţii de Agrostis capillaris, Elymus
repens, Poa pratensis, Alopecurus pratensis, Phragmites australis, Cirsium arvense, Carex ligerica.
Apa freatică afectează profilul de sol, fiind situată la adâncimi 1-2 m. Procesul de formare al GS eu este asemănător cu cel al GS ce (lăcovişti), fiind determinat de prezenţa apei freatice la adâncime mică, dar apa este mai săracă în carbonat de calciu. Acumularea mai slabă de humus calcic, cât şi excesul de apă, care produce condiţii de anaerobioză, determină o mai slabă mineralizare a materiei organice şi o culoare mai deschisă a orizontului Ao (BĂLĂCEANU şi MARIAN, 1985).
Excesul de umiditate produce fenomene de reducere şi apariţia orizontului de glei (Gr) în primii 125 cm, bogat în minerale secundare de tipul fero-silicaţilor, cu Fe feros colorat în verzui sau albăstrui. La suprafaţa solului, unde excesul de umiditate este periodic, se formează hidroxizi ferici şi manganici, care se depun sub formă de bobovine şi pete colorate în brun gălbui sau brun roşcat. În general, textura este uniformă pe profilul de sol.
GS eu prezintă un orizont Ao cu grosimi care variază între 20-30 cm, de culoare cenuşie închisă-brun cenuşie închisă (10 YR4/1-2) cu pete brune gălbui închise (10 YR4/4) la umed, structură grăunţoasă moderat dezvoltată. Urmează un orizont de tranziţie A/Go, de cca 20-50 cm, de culoare brun oliv-brun cenuşie (2.5Y 4/4-5/2) în stare umedă, cu o structură poliedrică sau masivă, bobovine numeroase. Orizontul Gr situat sub 60-80 cm, are culoare cenuşie-verzuie (N5-5GY5/1) cu pete oliv (5Y5/4) şi gălbui ruginii (7.5YR6/8) în stare umedă, masiv, bobovine dese (GRIGORAŞ, 1999).
Textura variază de la un areal la altul, de la nisipoasă, la argiloasă. Gleiosolurile molice sunt relativ des întâlnite având un orizont Am închis la culoare, iar gleiosolurile histice prezintă un orizont de turbă la suprafaţă, desupra orizontului A.

173
Gleiosolurile histice (GS tb) Prezintă la suprafaţa solului un orizont Ts gros de 24 cm, format din depozite
organice, culoare (5YR 6.5/1) neagră-brună foarte închis (10YR 2/1.5) în stare umedă, cenuşie foarte închisă-brună cenuşie foarte închis (10YR 3/1.5) în stare uscată, masiv-nestructurat, foarte friabil, cu tulpini nedescompuse frecvente, un orizont A+G gros de 10-15 cm, argilos, negru-brun foarte închis (10YR 2/1.5) în stare umedă, cenuşie foarte închisă-brună cenuşie foarte închisă (10YR 3/1.5) în stare uscată, cu pete cenuşiu verzui închis (5GY 4/1), structură masivă spre nestructurat, foarte friabil, cu resturi vegetale puternic descompuse (MUNTEANU ş.a., 1994). Orizontul Ck/Grk sub adâncimea de 35 cm, cu 6 suborizonturi până la adâncimea de 200 cm, cu textură lutoasă cu stratificaţii nisipoase şi luto-argiloasă în Ck2/Grk, culoarea variază de la cenuşiu închis (5YR 5/1) în stare umedă şi cenuşiu deschis (5YR 6.5/1) în stare uscată, la partea superioară şi cenuşiu închis (N 4) în stare umedă şi cenuşiu verzui deschis (7.5YR 4/4) în stare uscată, structură masivă, friabil, cu oxizi/hidroxizi şi pelicule, efervescenţă puternică (MUNTEANU;1994). Orizontul de turbă nu conţine carbonaţi, cantitatea de CaCO3 total variind pe profil de la mică în A+G (0.4 %), la mijlocie-mare pe profilul de sol (11.9-15.4 %).
Solul are o reacţie slab acidă în Ts (pH=6.4) şi slab alcalină pe profil (7.6-8.0). GC au un conţinut foarte mare în materie organică în Ts, conţinut mijlociu de humus în A+G (6 %) şi de la mic la foarte mic pe profil (1.0-1.5 %). Conţinut foarte mare de N total (1.11-1.29 %) în Ts, mare în A+G (0.39 %) şi foarte mic în restul profilului (0.093 %). Conţinutul în P mobil este mare în orizontul Ts (44-63 ppm) şi foarte mare în orizontul A+G (78 ppm), iar conţinutul în K mobil este mare-foarte mare în Ts (268-682 ppm) şi mijlociu în A+G (199 ppm). T este foarte mare în orizontul turbos (56.7-65.5 me/100g sol), mare în A+G (40.1 me/100g sol) şi mică în restul profilului de sol (15.6-20.6 me/100g sol).
Legat de aportul de microelemente se poate spune că gleiosolurile histice au un conţinut mare de Zn în Ts (6.28-7.40 ppm) şi în A+G (7.21 ppm) şi de la mic la mijlociu pe profil (0.84-1.59 ppm). Un conţinut mare în Cu în Ts (9.8-8.1 ppm) şi în A+G (9.8 ppm) şi mijlociu pe profil (5.8-7.3 ppm), conţinut miljlociu-mare de Mn activ (40.0-54.4 ppm) în Ts, mijlociu în A+G (32.0 ppm) şi Ck2-4/Grk, respectiv mic în Ck1/Grk (15.0 ppm). Conţinutul în B este de la mare la foarte mare (conţinut toxic) în Ts (1.75-4.32 ppm), foarte mare în A+G (2.37 ppm) şi foarte mic în rest (0.41-0.75 ppm), iar conţinutul de Mo este de 0.88 ppm în Ts şi 0.53 ppm în A+G.
Analiza fracţiunii argiloase ne arată preponderenţa smectitului care variază pe profilul de sol, de la 52 la 58%, în detrimentul ilitului (37-39%) şi a caolinitului (3-5%), deci, prin uscare pot apărea crăpături mari. În general, profilul de sol al GC prezintă o textură lutoasă peste tot, cu tendinţa de creştere a conţinutului în argilă în orizontul Ck2/Grk la 32.9%, DA este extrem de mică-foarte mică pe tot profilul de sol (0.45-1.2 g cm-3), implicit PT este extrem de mare-foarte mare pe tot profilul (55.5-80.2 %). PA este de asemenea foarte mare în Ts (32.6-26.2 %), mijlocie în A+Gs
(18.2 %) şi mică în Ck1/Grk şi Ck3/Grk (11.4 %) şi extrem de mică în Ck4-6/Grk. CO este extrem de mare-foarte mare în Ts (21.0-30.2 %), foarte mare în A+Gs
(19.7 %), mijlocie pe tot profilul de sol (8.7-11.3 %). CC este extrem de mare în Ts şi A+Gs (80.0-120.0 %) şi mijlocie în rest (24.0-26.3 %), ca şi CU care este extrem de

174
mare în Ts şi A+Gs (60.4-89.9 %) şi foarte mare în rest (15.8-17.4 %). Ksat este foarte mare pe tot profilul de sol (41.6-108.8 mm h-1). RP este de la mică la foarte mică pe tot profilul de sol (3-15 Kgf cm-2), solurile fiind slab maturate şi foarte afânate, gradul de tasare fiind negativ de la (-14.0 la –44.3).
GS ce au un potenţial de fertilitate ridicat, dar care nu poate fi valorificat decât prin lucrări de îmbunătăţiri funciare, care să elimine excesul de umiditate şi să îmbunătăţească însuşirile fizice şi fizico-chimice ale acestora. În condiţii naturale, neameliorate, se folosesc ca păşuni şi fâneţe slabe.
Histosoluri TB-solul turbos, sunt soluri organice, cu orizont de turba >50 cm grosime în primii 100 cm adâncime, fără ca stratul mineral situat în primii 25 cm, să atingă 20 cm grosime. Pot fi eutrice sau districe (tinoave) (POP,1960).
Indiferent de cantitatea de humus sau materie organica, solurile hidromorfe prezintă carenţe de nutriţie cu azot, întrucât excesul de umiditate împiedică nitrificarea. Ele au un potenţial de fertilitate mai mic sau mai mare, după cum sunt districe sau eutrice. Principalul factor limitativ rămâne excesul de umiditate.
Solurile cu exces de umiditate stagnanta respectiv stagnosolurile - tipice, luvice sau albice se dezvolta pe suprafeţe vechi plane şi, în cele mai multe cazuri, provin din luvosoluri tipice cărora li se înrăutăţeşte drenajul în urma defrişării şi instalării pajiştilor secundare. În majoritatea cazurilor sunt debazificate şi acide şi ridică probleme complexe de ordin agrochimic şi hidroameliorativ.
TB, cunoscute în clasificările mai vechi sub denumirea de turbă şi plaur, s-au format în condiţiile unui relief cu caracter depresionar de lunci, delte şi văi joase, pe depozite cu textură diversă, predominant carbonatice, în condiţiile unui climat cu caracter continental pronunţat (zona climatică călduroasă-secetoasă), dar şi în condiţiile unui climat umed şi rece, din arealele cu caracter depresionar din zonele montane (tinoavele şi bahnele).
Vegetaţia naturală este formată din vegetaţie higrofilă de baltă, Phragmites australis, Carex sp., Juncus sp., Typha şi păduri de Salix cinerea, Populus tremula, Betula pendula.
Apa freatică este situată la suprafaţa solului sau în imediata apropiere. Procesul de formare se caracterizează prin existenţa unui mediu saturat în apă, bogat în vegetaţie hidrofilă. După moarte, resturile acestor plante se acumulează şi se descompun parţial, ca rezultat al mineralizării lente datorate mediului anaerob. Procesul de bioacumulare trebuie să aibă un conţinut de materie organică mai mare de 35%, dacă partea minerală este argiloasă (peste 60% argilă <0.002 mm), 20-35 % materie organică, la conţinut proporţional de argilă şi cel puţin 20%, dacă partea minerală nu conţine argilă.
Stratul de turbă rezultat poate avea grosimi relativ mari, care poate ajunge de ordinul metrilor (10 m). Excesul de umiditate produce fenomene de reducere şi apariţia orizontului de glei, bogat în minerale secundare, de tipul bicarbonatului de tip feros şi manganos, colorat în cenuşiu-verzui sau albăstrui. Adesea în cadrul orizontului de turbă apar inercalaţii minerale, reprezentând aluviuni depuse la viituri mai mari (C. GRIGORAŞ, 1999).

175
La nivel de subtip, în funcţie de gradul de descompunere a materiei organice, şi
de aciditate se întâlnesc TB districe, eutrice. Din punct de vedere al reacţiei solului au fost deosebite şi soluri turboase eutrofe, cu reacţie alcalină.
Histosoluri districe TB di Sunt soluri turboase oligotrofe, cu reacţie puternic acidă (PH <4), cu un
conţinut foarte mare în materie organică, slab descompusă (93-99%) TB hemic sulfatic acid (MUNTEANU, 1996) prezintă un orizont Ts gros de 18
cm, format din material organic sapric, brun foarte închis (10 YR2/2), nestructurat, cu o reţea deasă de rădăcini vii şi moarte. Urmează un orizont Th gros de 40 cm, format din material organic hemic, brun-brun închis (10 YR3-4/3), rădăcini groase vii 3-5 cm diametru, foarte afânat, apoi un orizont Th/Gr gros de 25 cm, cu textură argiloasă, cenuşiu (N5), rădăcini fine semidescompuse, între 82-97 cm adâncime, după care urmează orizontul T/Gr, format din turbă hemică, cu material lutos de culoare cenuşie deschisă (N4), rădăcini şi frunze descompuse, frecvente, foarte moale, sub această adâncime urmând orizonturi minerale de glei Grk, stratificate de la nisip fin, la luturi, culoare cenuşie-cenuşie oliv (5 Y5/1.5), cu carbonaţi. Orizontul de turbă nu are carbonaţi, cantitatea de CaCO3 total sub adâncimea de 97 cm este mijlocie (10.6-12.7%).
Solul are o reacţie moderat acidă în primii 40 cm (PH=5.2-5.6), puternic acidă (4.6), în Th2, pe adâncimea 40-57, extrem de acidă-foarte puternic acidă (3.5-4.3) pe grosimea de 57-97 cm, în orizonturile ThGr şi Tgr şi slab alcalină pe profil, sub această adâncime (7.9-8.4). Are un conţinut foarte mare de materie organică până la adâncimea de 98 cm (20.4-42.8 %), conţinut foarte mare în N total (1.680-1.730 %) în orizontul turbos. T este foarte mare în orizontul turbos (95.6-139.9 me/100g sol) şi mică în rest (11.8 me/100g sol). SB este foarte mare-extrem de mare în orizonturile de turbă (57.8-113.3 me/100g sol) şi mică în rest, iar SH este mare în orizonturile fără carbonaţi (26.6-44.5 me/100g sol) şi lipseşte, în rest. Conţinutul total în săruri este mare în orizontul turbos (624-364 mg/100 gr sol) şi mai redusă, în orizonturile nisipoase din bază (76-137 mg/100 gr sol), predominând sulfaţii solubili (SO4--), cu valori de 14.8-21.8 me/100g sol în turbă şi 1.3-1.7 me/100g sol în stratele de nisip din bază.
TB pun probleme deosebite în cazul luării lor în cultură, în incinte desecate şi îndiguite, datorită poziţiei în areale microdepresionare, cu pericole permanente de ridicare a nivelului freatic şi de salinizare accentuată, lipsa portanţei datorită gradului slab de maturare a solului.
În condiţii naturale, când nu au apa de la suprafaţă, TB sunt ocupate de stuf sau păşuni foarte slabe. Turbele pot fi folosite, ca îngrăşământ natural pentru alte soluri sărace în humus, în pepiniere şi grădini, la răsaduri.
Stagnosoluri (SG) Sunt soluri având orizont A ocric (Ao) sau orizont a ocric şi orizont eluvial E
(Ao+El sau Ea) urmate de orizont B argic (Bt) la care se asociază proprietăţi stagnice intense (orizont W) începând de la suprafaţă sau din primii 50 cm ai solului mineral şi care continuă pe cal puţin 50 cm grosime. În mod frecvent apar concreţiuni ferimanganice. Nu prezintă schimbare texturală bruscă (între E şi Bt) pe cel mult 7,5

176
cm (diagnostică pentru planosoluri). Pot prezenta orizont vertic asociat orizontului B, orizont histic (sub 50 cm grosime) şi proprietăţi gleice sub 50 cm adâncime.
Stagnosolurile se întâlnesc pe forme de relief cu drenaj natural foarte slab, suprafeţe practic orizontale şi microdepresiuni, acoperite cu depozite fine foarte puţin permeabile mai ales pe terasele râurilor şi pe câmpia piemontană din vestul ţării (Câmpia Crasnei, Câmpia piemontană şi terasele Crişurilor, Câmpia piemontană şi terasele Timişului), în unele sectoare din Depresiunea Braşov, compartimentul Tg.Secuiesc, în Moldova în depresiunile Liteni şi Rădăuţi şi terasele inferioare ale Siretului, Moldovei şi Someşului, în platformele Cotmeana şi Cândeşti. Sunt caracteristice în special zonei forestiere cu păduri de Quercus robur, Q. frainetto şi cu un covor erbaceu format din Juncus, Agrostis şi Carex. Materialul parental este format din argile deluvio-proluviale de vârstă pliocenă care acoperă pietrişuri şi nisipuri stratificate torenţial caracteristice piemonturilor. Se mai pot forma pe depozite argiloase mai recente, aluviale şi deluvio-proluviale caracteristice câmpiilor piemontane iar în cadrul Câmpiei Române sunt formate pe depozite loessoide lutoase şi luto-argiloase (CIOCÂRDEL, 1957).
Stagnosolurile au un conţinut mic de humus (2-4%) şi azot (0,10-0,15%), conţinutul de fosfor total este mic (sub 0,10%), reacţia solului este slab-moderat acidă în partea superioară, uneori mai acidă în BW, iar gradul de saturaţie în baze V nu scade sub 60-70%. Are un conţinut ridicat al capacităţii de schimb cationic T de 25-35% (Florea şi colab.1968).
Nigrisolurile. Apariţia lor, sporadica, pare legata de rocile sedimentare percolabile
(gresii) cu ciment calcaros. Se mai dezvoltă pe calcare cristaline şi roci magmatice bazice. Apar sub pajişti mezofile sau mezofil-higrofile, la 900-1200 m. În general sunt litice, au profil puternic dominat de humus, sunt nesaturate şi acide, prezentând carenţele specifice districambosolurilor şi prezintă în afara orizontului diagnostic Au şi un orizont Bv cambic.
Nigrisolul cambic de la Ileni - Depresiunea Făgăraşului (MARIAN, 1979) este format pe un evantai aluvio-proluvial, la altitudinea de 450 m, teren arabil, Tma-8,20C,
Apariţia lor, sporadica, pare legata de rocile sedimentare percolabile (gresii) cu ciment calcaros. Se mai dezvoltă pe calcare cristaline şi roci magmatice bazice. Apar sub pajişti mezofile sau mezofil-higrofile, la 900-1200 m. În general sunt litice, au profil puternic dominat de humus, sunt nesaturate şi acide, prezentând carenţele specifice districambosolurilor. Ele ocupă suprafeţe neînsemnate în munţi (BUZA, 2000).
Pma-691 mm, material parental format din luturi nisipoase de origine aluvio-proluvială cu prundişuri cu elemente silicatice în substrat, apa freatică la 3-4 m.
Din punct de vedere morfologic orizontul Au cu o grosime de 48 cm, prezintă la suprafaţă un suborizont Aa (arat), are o textură lutoasă, culoare neagră (10YR 2/1 în stare umedă, structură poliedrică subangulară mică moderat dezvoltată, slab coeziv, friabil în stare umedă, poros. Urmează un orizont de tranziţie AuB, mai deschis la culoare cu textură lutoasă-lut-nisipoasă, după care sub adâncimea de 84 cm urmează un orizont Bv, lut nisipos de culoare brun cenuşiu foarte închis (10 YR

177
3,5/2) în stare umedă, structură masivă gros de cca. 40 cm, după care urmează roca mumă.
Sunt soluri permeabile cu drenaj bun, aciditate moderată (5,3-5,9) în Au şi 6,0 în Bv, conţinut de humus ridicat (6,9-7,3%) în orizontul Au şi 0,8% în orizontul Bv. Rezerva de humus până la 50 cm extrem de mare (350 t/ha). Bine asigurat cu azot total, dar cu o valorificare mai slabă, cu 0,279-0,294 % în Au, asigurare moderată cu fosfor şi potasiu şi normală în forme mobile de microelemente, mobilizare slabă a aluminiului.
Pentru obţinerea unor recolte ridicate în special de cartof, orz, sfeclă de zahăr şi in fuior se recomandă îngrăşăminte cu fosfor şi potasiu şi amendamente calcaroase pentru combaterea acidităţii.
Luvisolurile sunt alcătuite în principal din preluvosoluri şi luvosoluri. În această clasă au fost cuprinse o gamă foarte variată de soluri care au ca
trăsătură comună o diferenţiere texturală accentuată pe profil între suprafaţa solului şi secţiunea de control, respectiv de la textura luto-nisipoasă la textura luto-argiloasă în orizonturile Ap, Ao, El sau Ea la textură luto-argiloasă şi argilă în secţiunea de control în orizonturile Bt, Btw sau Btyw. Ele se definesc prin existenţa orizonturilor El sau Ea mai grosier şi un orizont Bt îmbogăţit în argilă deci mai impermeabil, paralel cu acidifierea şi scăderea progresivă a fertilităţii solurilor pornind de la solurile brune luvice la luvisolurile albice.
Gradul de eluviere este pusă în evidenţă de valorile mari ale indicelui de diferenţiere texturală pe profil. Ca urmare cea mai mare partea argilei din orizontul Ao migrează către orizontul Bt.
Preluvosolurile (EL) Preluvosolurile fac parte din clasa Luvisolurilor şi se întâlnesc pe un areal
relativ larg pornind din Subcarpaţi, Dealurile Banato-Crişene, Podişul Getic, Podişul Central Moldovenesc, Podişul Sucevei, Podişul Someşan, Podişul Târnavelor, Podişul Dobrogei, unele depresiuni submontane şi extramontane.
Sunt soluri având orizont A ocric sau molic (Ao, Am) urmat de orizont intermediar argic (Bt) având culori cu valori peste 3,5 (la umed) cel puţin pe feţele agregatelor structurale începând din partea superioară şi grad de saturaţie în baze (V) peste 53%.
Pot prezenta orizont vertic, orizont Cca sau concentrări de carbonaţi secundari în primii 125 cm, orizont O şi proprietăţi stagnice intense (W) sub 50 cm sau proprietăţii gleice (Gr) sub 50 cm. La nivel de subtip au fost deosebite în zona montană preluvosoluri: tipice, molice, rodice, stagnice, gleice, calcice, litice, scheletice.
Preluvosoluri tipice (EL ti) se caracterizează prin prezenţa unui orizont Ao slab humifer şi a unui un orizont Bt (argic). Specific acestor soluri este coloritul brun gălbui (10YR 4-5/4), cu nuanţe roşcate în orizontul iluvial.
Preluvosoluri tipice de multe ori în asociaţie cu luvosolurile tipice (brune luvice) ocupă suprafeţe relativ întinse în regiunile deluroase şi piemontane cum sunt: Podişul Transilvaniei, Subcarpaţi, Piemontul Getic, Piemonturile Vestice. Dobrogea de Nord, în jurul culmilor Munţilor Măcinului, Podişului Niculiţelului şi a Dealurilor Tulcei.

178
Preluvosoluri tipice sau format în condiţii de climă temperată umedă cu influenţe oceanice. Media anuală a precipitaţiilor este mai mare sau cel puţin egală cu evapotranspiraţia potenţială oscilând între 600-1000 mm. Temperatura medie anuală variază între 7,6-10,40 C.
Vegetaţia naturală este formată din pădure mezofilă cu asociaţii de Quercus petraea, Q.robur în amestec cu Fagus sylvatica în zonele mai înalte Carpinus betulus, Acer campestre, Prunus mahaleb şi specii ierboase de Brachypodium sylvaticum, Viola reichenbachiana, Carex spicata, Anthriscus sylvestris şi Hedera helix. Altitudinal aceste soluri pot fi întâlnite între 150-800 m în condiţii de relief fragmentat, pe culmi şi versanţi cu diferite înclinări, pe interfluvii cu relief mai domol, piemonturi, câmpuri şi terase.
Materialul parental este format din depozite foarte variate sub raportul originii compoziţiei mineralogice, texturii şi vârstei: loessuri cu textură lutoasă şi luto-argiloasă, luturi roşcate, nisipuri, gresii, argile, marne, dar şi din depozite deluviale şi eluviale, iar apa freatică nu influenţează profilul de sol, depăşind 15 m, drenajul global fiind moderat-bun, putând să apară fenomene de pseudogleizare slabă (FLOREA şi colab., 1968). Excepţie fac preluvosolurile tipice situate în cîmpia joasă a Someşului care prezintă probleme de exces de umiditate freatică (23.288 ha). Din punct de vedere pedogenetic preluvosolurile tipice beneficiază de un surplus de umiditate din precipitaţii şi temperaturi relativ mai scăzute faţă de CZ ar, procesul de bioacumulare este puternic, dar datorită faptului că s-a format predominant pe seama litierei şi a vegetaţiei lemnoase de pădure, duce la formarea unei humus mai slab calitativ, alcătuit din procente egale de acizi huminici şi fulvici. Datorită substratului calcaros bogat în ioni de calciu, humusul este saturat, de tipul mull forestier.
Caracteristica cea mai importantă este procesul mai avansat de debazificare a complexului coloidal al solului, formarea unei cantităţi mai mari de argilă şi depunerea acesteia în orizontul Bt (argic). Carbonaţii de calciu sunt levigaţi sub adâncimea de 120 cm, fiind acumulaţi sub forma unui orizont Cca mai slab exprimat.
Prezintă un profil de tipul Ol-Ao-Bt-Cca la subtipul tipic, Ol-Am-Bt-Cca la subtipul molic, cu Btw la subtipul gleic şi cu Bty la subtipul vertic. Orizontul Ao are o grosime de cca 30 cm, culoare brun cenuşie foarte închis (10YR 3/2.5) la umed şi brun cenuşie închis (10YR 4.5/2) la uscat, textură lutoasă, structură poliedrică subangulară medie, bine dezvoltat, friabil, poros. Urmează un orizont de tranziţie A/B gros de 15 cm, culoare brun gălbuie închis (10YR 4/4) în stare umedă, brun gălbuie (10YR 5/3.5) în stare uscată, structură poliedrică subangulară mare-prismatică medie, bine dezvoltată, friabil, cu concreţiuni-oxizi/hidroxizi, pori mici, compact. Orizontul Bt situat între 45-165 cm are culoare brun gălbuie închisă (10YR 4/4) la umed şi brun gălbuie (10YR 5.5/4) la uscat în partea superioară, respectiv brun gălbuie (10YR 5/4) în stare umedă şi brun gălbuie deschis-brună foarte pal (10YR 6.5/4), cu textură lutoasă-luto argiloasă, structură prismatică-columnoidă medie, bine dezvoltată, ferm în stare umedă şi dur în stare uscată, moderat compact. Între 45-65 cm adâncime apare un suborizont Bt melanic, mai închis la culoare, brun gălbui închis (10YR 3.5/4) în stare umedă, brun gălbui (10YR 4,5/4) în stare uscată. Orizontul Ck este situat sub adâncimea de 165 cm, având culoare brun gălbuie-brun gălbuie deschisă (10YR 5.5/4) în stare umedă, brună foarte pal (10YR 7.5/4) în stare uscată, cu textură

179
lutoasă, masiv spre nestructurat. Prezintă schelet, acumulare de CaCO3 sub formă de eflorescenţe. Are un conţinut moderat de carbonaţi (10.0 %) sub 165 cm adâncime.
Reacţia solului este slab acidă în Ao (PH=6.8), A/B (6.5) şi Bt2 (6.4), moderat acidă în Bt3 (5.3) şi Bt4, (5.7), moderat alcalină în Ck (8.6). Este saturat în baze, V cifrându-se la 91 % în Ao şi 86 % în A/B, 81 % în Bt3 şi 100 % în Cca. SH atinge 2.3 me/100g sol în Ao, 2.0 me/100g sol în Bt1 şi creşte în Bt3 la 3.1 me/100g sol, având un conţinut mijlociu în SB, cu 23.9 me/100g sol în Ao, un conţinut mic în Bt1 (14.9 me/100g sol), Bt4 (12.6-21.7 me/100g sol) şi în Ck (14.7 me/100g sol). T este mijlocie în Ao (26.2 me/100g sol) şi mică în restul profilului. EL ti au un conţinut mare în humus în Ao (până la 7.0 %) şi foarte mic în Bt1 (0.7 %). Rezerva de humus este mijlocie (125 t/ha). Solurile sunt bine aprovizionate în N total (0.375 %) în Ao şi foarte slab aprovizionate în Bt1 (0.079 %), foarte bine aprovizionate cu P mobil în Ao (42 ppm), foarte bine asigurate cu K mobil (325 ppm).
Microelementele sunt prezente printr-un conţinut mare în Zn în Ao (4.64 ppm) şi foarte mic în AB (0.64 ppm) şi Bt2 (0.54 ppm). Sunt mijlociu aprovizionate cu Cu în Ao (4.4 ppm), A/B (4.7 ppm), Bt1 (4.4 ppm) şi mic în Bt3 (3.7 ppm), având conţinut foarte mare de Mn activ în Ao (123 ppm), mic în A/B (14.5 ppm), mijlociu în Bt1 (28.5 ppm) şi foarte mic în Bt3 (7.5 ppm). Conţinutul în B este mare în Ao (1.18 ppm) şi mic (0.26 ppm) în rest în A/B şi Bt2, iar conţinutul în Mo este de 0.10 ppm în Ao şi 0.17 ppm în A/B.
EL ti prezintă o textură lutoasă pe tot profilul de sol, cu tendinţa de creştere a conţinutului în argilă în orizontul Bt. DA este extrem de mică în Ao (0.97 g cm-3) şi A/B 1,17, mică în Bt1, mijlocie în Bt2 (1.41 g cm-3) şi foarte mică în Ck (1.17 g cm-3). PT este extrem de mare în Ao (63.3 %), foarte mare în A/B (56.3 %), mare în Bt1 (52.2 %), mijlocie în Bt2 (47.7 %) şi foarte mare în Ck (56.6 %). PA este foarte mare în Ao (36.3 %), mare în A/B (26.9 %), mijlocie în Bt2 (16.0%) şi foarte mare în Ck.
CO este mijlociu pe tot profilul de sol, în orizontul Ao (11.1 %), în A/B (10.1 %), în Bt4 (9.9 %) şi în Ck (8.7). CC este mare în Ao (27.8 %) şi mijlocie în Bt (21.2-23.7 %) şi în Ck (20.6 %). CU este foarte mare în Ao (16.7 %), mare în A/B (15.1 %) şi mijlocie în rest, în Bt2 (12.0 %), Bt4 (11.3 %) şi în Ck (11.9). Ksat este foarte mare în Ao (46.9 mm h-1), mare în A/B şi Bt1 (21.3 mm h-1), mijlocie în Bt3 (5.4 mm h-1) şi în Ck (10.2 mm h-1). RP este mică în Ao (20 Kgf cm-2), mijlocie în A/B (32 Kgf cm-
2), Bt1 (29 Kgf cm-2), Bt2 (32 Kgf cm-2) şi mică (25 Kgf cm-2) în Bt4 şi în Ck (19 Kgf cm-2).
EL ti nu pun probleme deosebite în situaţia luării lor în cultură. În cazul folosirii irigaţiilor, se recomandă norme de udare mici şi dese, pentru a preîntâmpina eventuala stagnare a apei pe profil. La solurile situate pe pante eroziunea poate fi combătută prin arături pe curba de nivel, prin alegerea sortimentului adecvat de culturi, benzi înierbate, ş.a. Sunt soluri relativ favorabile unei game largi de folosinţe şi culturi, dar cu limitări legate de poziţia pe versant, fertilitatea mai slabă, lipsa precipitaţiilor atmosferice în perioada de vară.
Luvosolurile (LV) Aceste soluri se întâlnesc pe un areal larg pornind din partea centrală şi nordică
a sectorului în regiunile piemontane Cotmeana şi Cândeşti, Olteţului, Dealurile Subcarpatice, Dealurile Banato-Crişene, Podişul Central Moldovenesc, Podişul

180
Sucevei, Podişul Someşan, Podişul Târnavelor, Podişul Dobrogei, unele depresiuni submontane şi extramontane, în special în zonele cu caracter depresionar.
Media anuală a precipitaţiilor este mai mare sau cel puţin egală cu evapotranspiraţia potenţială oscilând între 600-1000 mm. Temperatura medie anuală variază între 7,6-10,40.
Vegetaţia naturală este formată din pădure mezofilă cu asociaţii de Quercus petraea, Q. robur în amestec cu Fagus silvatica în zonele mai înalte Carpinus betulus, Acer campestre, Prunus mahaleb şi specii ierboase de Brachypodium sylvaticum, Viola reichenbachiana, Carex spicata, Anthriscus sylvestris şi Hedera helix. Altitudinal aceste soluri pot fi întâlnite între 150-800 m în condiţii de relief mai domol, piemonturi, câmpuri şi terase.
Materialul parental este format din luturi, depozite loessoide, argile, marne şi gresie.
Din punct de vedere mineralogic, la luvosolurile tipice formate pe loess la suprafaţa solului, în orizontul Ao predomină ilitul 51%, urmat de smectit cu 45%, caolinit 4% şi urme de clorit. În orizontul El predomină tot ilitul 54%, urmat de smectit 42%, caolinit 4% şi urme de clorit; pe profil, smectitul devine predominant, cu 52-60% în Bt şi 64% în Ck, în dauna ilitului, care deţine 35-44% în Bt şi 32% în Ck, respectiv a caolinitului, cu 4-5% în Bt şi 4% în Ck. Dintre minerale predomină micele, cu 13% în primii 65 cm, în Ao, El şi Bt1 (10%), în Bt2 şi în Ck (14%), amfibolii cu 2% în primii 65 cm, 10% în Bt2 şi 5% în Ck, piroxenii care apar în Bt1
(6%), Bt2 (15%) şi în Ck (6%); mai apar cantităţi mici de granaţi (2-4%) pe tot profilul de sol şi disten şi staurolit, sporadic (2-3%) (CRĂCIUN şi POPESCU; Ghidul excursiilor, 1994).
Profilul de sol este de tipul Ap-Ao-El-EB-Bt-C la luvosolurile tipice (brune luvice) (6.124.876 ha ) şi cu Ea la luvosolurile albice (luvisolurile albice) (779.481 ha) şi cu Bty la subtipul vertic.
La nivel de subtip se deosebesc luvosoluri: tipice, umbrice, roşcate, rodice, calcice, rezicalcarice, psamice, vertice, albice, glosice, planice, stagnice, gleice, litice, scheletice şi solodice (sodice). În cazul planosolurilor la nivel de subtip acestea pot fi: tipice, albice, vertice, stagnice şi solodice (sodice).
Profilul de sol are o structură grăunţoasă-poliedric subangular mic în Ao, poliedric angular mic-mediu spre lamelar în El sau Ea şi prismatic mare în Bt.
Însuşirile fizice devin mai nefavorabile de la luvosolurile tipice la luvosolurile albice în relaţia acestor soluri cu apa, respectiv devin mai tasate şi mai puţin permeabile. Astfel densitatea aparentă devine ridicată, porozitatea totală este mijlocie la suprafaţă şi mică pe profil, porozitatea de aeraţie este mică la suprafaţă şi foarte mică pe profil, indicele de contracţie este mare şi foarte mare şi foarte mare în secţiunea de control la subtipul vertic. Coeficientul de ofilire este mare-foarte mare, capacitatea de câmp este mare iar capacitatea de apă utilă este mijlocie. Permeabilitatea acestor soluri este mică şi foarte mică, cu o viteză de infiltraţie de 0,18 mm/h. Aceasta explică fenomenul de pseudogleizare care în funcţie de condiţiile locale poate fi de la slab-moderat la excesiv şi regimul aerohidric deficitar (LUPAŞCU,1986, 1996).

181
Sunt soluri mai puţin fertile, aceasta scăzând de la solurile luvosolurile tipice şi planosolurile tipice la luvosolurile albice. Conţinutul de humus este scăzut variind de la 1,2-2,4% la luvosolurile tipice şi 1,2-1.6% la luvosolurile albice. Reacţia solului variază de la moderat la puternic acidă pH 4,8-5,8 în orizontul Ao la luvosolurile tipice şi puternic acidă pH 4,5-5,5 la luvosolurile albice, sunt soluri oligobazice şi mai rar oligomezobazice. Aluminiul mobil variază în primi 20 cm de la 0,4-34,4 mg/100g sol cu efecte toxice asupra plantelor de cultura. Gradul de saturaţie în baze poate ajunge la 50%, CT este relativ ridicată 12,2-22,2 me dar cu predominarea ionilor de H. (SECELEANU şi colab 1998)
Sunt soluri mai puţin favorabile, fertilitatea acestora scăzând cu creşterea gradului de eluviere, culturile agricole suferă mult de lipsa apei vara şi are apă în exces primăvara şi toamna. Pentru reglarea regimului aerohidric se preconizează realizarea de lucrări hidro şi pedoameliorative cum sânt: canale deschise permanente, drenaje cu prismă filtrantă înaltă pentru arealele depresionare, afânare adâncă, scarificare, arătură în benzi cu coame. Debazificarea înaintată a complexului argilo-humic, aciditatea acestora asociată cu prezenţa aluminiului mobil şi a manganului, adesea cu doze nocive pentru creşterea plantelor, deficitul mare de calciu şi magneziu conduc la condiţii puţin favorabile dezvoltării plantelor (BADEA, 1967).
Se recomandă aplicarea de amendamente calcaroase şi încorporare de gunoi de grajd pentru suplinirea necesarului de azot, fosfor şi potasiu.
Fertilitatea acestor soluri este relativ scăzută şi scade cu cât ne îndreptăm spre regiunile de piemont şi subcarpatice legate de însuşirile fizice şi chimice mai puţin favorabile şi condiţiile de mediu mai austere.
Alosolurile-AL ( sol brun luvic şi luvisol albic holoacid). Sunt soluri având orizont A ocric sau umbric (Ao, Au) urmat direct sau după
un orizont eluvial (E), de orizont B argic (Bt) având proprietăţi alice pe cel puţin 50 cm, între 25 şi 125 cm adâncime (sau cel puţin jumătate din orizont dacă apare orizont R sau C la adâncime mai mică). Poate prezenta orizont organic sau proprietăţi stagnice moderate (w) sau intense (W) sub 50 cm adâncime. Alosolurile sunt soluri acide, cu conţinut mediu - mare de argilă, bine drenate, culoare brună la suprafaţă, structură poliedrică mijlocie sau prismatică, crapă la uscare, culoare roşcată în orizontul Bt. Are un orizont B argic cu o capacitate de schim cationic mai mare sau egală cu 24 me/100 g argilă în primii 100 cm.
Este specific regiunilor tropicale şi sub tropicale umede şi temperate calde, la noi în ţară se găsesc pe suprafeţe reduse în la poala Carpaţilor şi Subcarpaţilor şi în special în piemonturile şi glaciesurile piemontane formate din depozite acide Piemontul Getic (Cândeşti, Cotmeana, Jiului) şi piemontul din faţa Munţilor Poiana Ruscă.
Pelosolurile (PE) Sunt soluri având orizont pelic la suprafaţă sau de la cel mult 20 cm (sub stratul
arat) ce se continue până la cel puţin 100 cm; conţin peste 30% argilă în toate orizonturile până la cel puţin 100 cm adâncime.
La nivel de subtip se deosebesc pelosoluri: tipice, brunice, argice, gleice, stagnice.

182
Pelosolurile sunt întâlnite în areale asemănătoare cu cele ale vertosolurilor respectiv câmpii înalte platouri piemontane şi deluroase şi de terase medii şi superioare ale unor râuri dar şi în zone de câmpie joase cu exceas de umiditate şi de foarte multe ori asociate cu vertisolurile. Aceasta se realizează în condiţiile în care argilele au o cantitate mai redusă de minerale expandabile şi nu se pot realiza vertosoluri sau când umiditatea mai mare nu permite producerea fenomenului de contracţie şi gonflare. Pelosolurile sunt întâlnite în cadrul Câmpiei Române de Vest, Piemontului Getic, Câmpiei şi Dealurilor de Vest, Podişului Transilvaniei şi Moldovei (PĂLTINEANU şi colab., 2003).
Pelosol gleic-stagnic (PE gl-st) Este întâlnit în arealele de câmpii joase şi luncile râurilor mai importante din
ţară în special în zona mai umedă. Sunt formate pe argile de origine aluvială sau fluvio-lacustră. Un profil de sol reprezentativ este cel realizat de MUNTEANU şi colab. (Ghidul excursiilor, 2003).
Prezintă un orizont Ao de 20 cm, argilo-prăfos (48,7% argilă < 0,002 mm), brun cenuşiu închis (10YR 3,5/2 la umed), structură poliedrică angulară mică şi medie puţin dezvoltată, crăpături de 2-3 cm lărgime. Urmează un orizont AB de 15 cm, argilo-prăfos (45,8%), brun cenuşiu închis, (10YR 4/2 la umed), structură poliedrică angulară mare şi medie foarte bine dezvoltată, separaţii ferimanganice şi bobovine mici, compact, crăpături de 2-3 cm lărgime. În continuare urmează un orizont B cambic (Bv)care are şi caractere pelice (z) şi proprietăţi stagnice şi gleice (w, G) format din trei suborizonturi, respectiv: Bz1wac de 10 cm, lut argilo-prăfos (43,7%), brun cenuşiu foarte închis (10YR 3/2 la umed), structură mare-masivă, foarte dens, foarte compact, crăpături de 2-3 cm lărgime, bobovine foarte mari; Bz2Grac de 37 cm, argilă lutoasă (60,0%), cenuşiu oliv închis (5Y3/2 la umed), masiv, foarte dens, compact, crăpături de 1-3 cm lărgime, concreţiuni silico-calcaroase gălbui mari rare, bobovine mari; Bz3Grac de 13 cm, argilă medie (61,2%), cenuşiu oliv (5Y 5/2), masiv, compact, crăpături rare, semiplastic, concreţiuni silico-calcaroase rare, bobovine frecvente. În baza profilului se găseşte un orizont Ck rezidual CkGrac de 22 cm, argilă prăfoasă (57,3%), cenuşiu închis (5Y 4/1), concreţiuni silico-calcaroase albe cenuşii foarte numeroase, bobovine frecvente, semiplasti
Este un sol relativ tânăr format pe depozite fluvio-lacustre fine, care a evoluat în regim de exces de umiditate, în prezent semidrenat funcţionează în continuare ca sol hidromorf datorită împachetării dense care determină stagnarea apei din precipitaţii. Alcalizarea şi concreţiunile calcaroase foarte numeroase provin dintr-o fază anterioară de evoluţie. Este un sol cu textură fină, dens şi compact care fisurează profund la uscare, aeraţie slabă şi permeabilitate redusă, exces de apă stagnantă şi alcalizare moderată în orizonturile inferioare. Afânarea adâncă pare să fie o soluţie de ameliorare a condiţiilor fizice.
Conţinut mic de humus în orizontul Ao (3,90%), A/B (4,75%), Bz1wac (3,54%), foarte mic în Bz2Grac (1,14%) şi extrem de mic în Bz3Grac (0,65). Conţinutul de azot total este mijlociu în Ao(0142%) şi A/B (0,147%) şi mic în Bz1wac (0,137%). Conţinut mic de P mobil (ppm) în Ao (9,0) şi A/B 10,1. Conţinutul de K mobil (ppm) este mijlociu în Ao (191), A/B (177) şi Bz1wac (200).

183
Na schimbabil me/100 g sol este în cantitate foarte mică în Ao, A/B şi Bz1wac, (0,64-1,32), mică în rest (2,57-3,06).
Este slab acid în Ao cu pH în apă 5,98, A/B pH 6,19 şi Bz1wac pH 6,45, slab alcalin în B2zGrac cu pH 7,72, moderat alcalin în Bz3Grac cu pH 8,54 şi puternic alcalin în CkGrac cu pH 9,16.
SB me/100 g sol variază între 12,23-16,07 în primii 50 cm şi între 24,68-36,52 sub această adâncime, iar H schimbabil me/100 g sol are valori de 7,13 în Ao, 7,04 în A/B şi 5,60 în Bz1wac. Gradul de saturaţie în baze V8,3 %T variază între 63,5-73,7 în primii 50 cm şi este 100 în restul profilului. Însuşirile fizice sunt asemănătoare cu ale vertosolurilor (SECELEANU şi colab., 1975, 1988).
Vertosolurile (VS) (vertisoluri) Sunt soluri având orizont vertic de la suprafaţă sau de la cel mult 20 cm (sub
stratul arat ce se continue până la cel puţin 100 cm; conţin peste 30% argilă în toate orizonturile până la cel puţin 100 cm adâncime. Din punct de vedere geomorfologic, răspândirea acestor tipuri de sol este legată de câmpii înalte, platouri piemontane şi deluroase şi de terase medii şi superioare ale unor cursuri de apă, cu altitudini care variază între 100-400 m.
Pot fi întâlnite şi la altitudini situate peste 500m în arealele subcarpatice şi de dealuri înalte. În sudul ţării suprafeţe întinse se găsesc în Piemontul Getic respectiv în partea centrală şi sudică a Platformei Cotmeana, Cândeşti, Olteţului şi Motrului în câmpiile Piteştiului, Găvanu-Burdea, Boianului şi Pleniţei. Cea mai mare densitate a vertosolurilor şi pelosolurilor se găseşte pe arealul ocupat de interfluviile râurilor Olt şi Argeş (CRĂCIUN, 1998, 2000).
În vestul ţării ele se întâlnesc în Câmpia Tisei la Oraviţa, Gătaia, Recaş-Izvin, Salonta, Marghita şi Satu Mare şi la altitudini foarte variate pe terasele vechi ale râurilor la Hunedoara, Orăştie-200 m, Sebeş-400 m Sf. Gheorghe-Ilieni-600 m etc. în Transilvania. (Teaci şi colab. 1956). Pe suprafeţe relativ restrânse ele se mai găsesc în Nordul Moldovei în Câmpia Moldovei la Mileanca şi Păpăuţi, în sud estul Moldovei în Dealurile Fălciului, şi pe piemonturile şi câmpiile înalte care limitează spre est Subcarpaţii de Curbură.
Sunt soluri formate pe depozite argiloase cu caracter gonflant (smectite) prezentând o textură care în Ao variază de la luto-argiloasă până la argilo-lutoasă şi de la argilo-lutoasă la argilă în secţiunea de control în orizontul By. Profilul de sol este de tipul Ao-By-C sau Byw la tipul pseudogleizat.
Prezintă un aspect morfologic diferit al profilului de sol şi un comportament diferit în relaţia cu apa, diferit de alte soluri vertice. Comportamentul diferit al acestor soluri se datoreşte faptului că aceste argile sunt expandabile şi la uscăciune se contractă puternic formând crăpături late de 5-10 cm până la 150 cm adâncime, iar la umezeală se dilată puternic executând o împingere puternică pe verticală şi laterală. Se creează nişte suprafeţe oblice de alunecare (slickensides) cu luciu metalic, cu înclinare de cca. 450 la nivelul cărora se produce presarea şi mişcarea diferitelor compartimente ale solului.
Un alt proces care se produce la suprafaţa solului realizat tot prin procesul de uscare şi gonflare este automulcierea care duce la granularea solului în particule mici şi tari alcătuind un strat de până la 5 cm grosime (in general 2-3 cm).

184
Grosimea stratului de mulch depinde şi de folosinţă, numărul de arături şi poziţia pe versant.
Din punct de vedere morfologic profilul de sol prezintă la suprafaţă un strat de mulch, feţe oblice de alunecare cu luciu metalic pe profil cu atât mai dese cu cât caracterul vertic este mai evident, o structură masivă sau poliedric angular-subangular mediu-mare în secţiunea de control. Culoarea închisă (negru-10YR 2/1), în general, pe adâncimi care pot să depăşească 1m, excepţie făcând vertosolurile brunice şi brunice stagnice care au culori mai deschise. În cazul vertosolurilor stagnice deoarece apa stagnează o perioadă mai îndelungată de timp în profilul de sol la nivelul orizontului Byw apar pete de oxido-reducere şi concreţiuni fero-manganice (bobovine) şi implicit însuşiri fizice mai nefavorabile.
Luate în general vertisolurile sunt soluri relativ fertile, slab-moderat aprovizionate cu humus în Ap (2,5-4,0%), cu o rezervă de humus în primii 50 cm de la mare la foarte mare (170-217t/ha, slab-moderat aprovizionate cu fosfor (11-25 ppm), mijlociu-bine aprovizionate cu potasiu (121,0-181,3 ppm). Sunt moderat acide în Ap (pH 5,1-5,6), neutru la slab alcalin în By şi gradul de saturaţie în baze variată de la mezobazic la eubazic. Însuşirile fizice sunt mai puţin favorabile, densitatea aparentă variază de la 1,41 la vertosolurile cu textoră luto-argiloasă în Ap la 1,39 la cele argilo-lutoase în Ap. Porozitatea totală este mijlocie în Ap (47,0-59,0%) şi de la mică la foarte mică în secţiunea de control (40,7-49,0%). Porozitatea de aeraţie este de la mijlocie la foarte mică în Ap (29,0-1,37%) şi foarte mică în rest (8,00-1,59%) inhibând dezvoltarea plantelor mai ales în ariile micro depresionare în sezonul ploios şi umed.
Coeficientul de ofilire este mare pe tot profilul de sol variind de la 18,8% la vertisolurile cu Ap luto-argiloase şi 15,7% la cele cu orizontul Ap argilo-lutos. Capacitatea de câmp este mijlocie pe tot profilul de sol şi variază de la 27,2% la vertosolurile cu Ap luto-argiloase la 25,6% la cele cu orizontul Ap argilo-lutos. Permeabilitatea solului este de la mică la mijlocie în Ap (1,9-3,8mm/h) şi de la foarete mică la excesiv de mică (0,49-0,20mm/h) în secţiunea de control.
Indicele de contracţie este de la mare la foarte mare pe tot profilul de sol (0,0091-0,015) fiind în concordanţă cu comportamentul argilelor gonflante pe care s-au format vertisolurile. (SECELEANU şi colab, 1992, 1999, 2003)
Pentru o exploatare agricolă cât mai favorabilă se preconizează acolo unde este cazul realizarea de lucrări de combatere (eliminare) a excesului de umiditate datorat stagnării apei la suprafaţa solului, prin drenaj de suprafaţă, prin canale deschise, nivelare pentru asigurarea pantei de scurgere către acestea, cu decopertare selectivă pentru a diminua scăderea fertilităţii solului. Local în zonele microdepresionare ocupate cu vertisoluri pseudogleizate moderat-puternic se pot asocia cu lucrări de drenaj închis (tuburi cu prizmă filtrantă înaltă). Ca lucrări agropedoameliorative se pot realiza lucrări de scarificare, rigole şi drenaj cârtiţă, fertilizare (PARICHI şi SECELEANU,1989). Vertosolurile sunt relativ favorabile pentru folosinţa păşune şi fâneaţă şi pentru cultivarea unei game largi de plante furajere, aceasta favorabilitate scăzând odată cu creşterea gradului de argilozitate şi legat de aceasta a creşteri intensităţii fenomenului de pseudogleizare.

185
Faeoziomurile (FZ) Sunt soluri având orizont A molic (Am), orizont intermediar (Bt, Bv, AC) cu
culori cu crome şi valori sub 3,5 (la umed) cel puţin în partea superioară (pe cca. 10-15 cm) şi cel puţin pe feţele agregatelor structurale şi fără orizont Cca sau concentrări de carbonaţi secundari în primi 125 cm (sau primii 200 cm în cazul texturii grosiere). Pelicule argilo-humice în orizontul B şi adesea caractere de hidromorfie când există orizont Bt. Sunt excluse solurile formate pe materiale parentale calcarifere sau roci calcaroase care apar între 20 şi 50 cm. Pot avea orizont vertic, proprietăţi gleice (Gr) sub 50 cm şi proprietăţi stagnice (w sau sub 50 cm, W).
Ele pot fi întâlnite în Podişul Moldovei, Câmpia Română de Vest, Câmpia de Vest şi Podişul Transilvaniei. Fiind situate în perimetrele mai umede şi mai reci decât solurile similare care au rămas încadrate la cernoziomuri. (BRÂNDUŞ,1981).
Faeoziomurile argice (FZar) Sunt formate în general pe depozite loessoide şi luturi cu un profil de tipul Ap-
Am-AB-Bt-Cca, cu textură lutoasă sau luto-argiloasă diferenţiată pe profil, orizontul Bt argiloiluvial fiind îmbogăţit în argilă.
În unele sectoare ale câmpiei ele prezintă un drenaj mai slab cu apariţia fenomenului de pseudogleizare cu intensitate de la slab la moderat. Structura acestor soluri este glomerulară bine dezvoltată în orizontul Am şi prismatică moderat dezvoltată în Bt. Structura bună şi textura favorabilă se reflectă în însuşirile fizice în general bune ale acestor soluri, care sunt totuşi mai tasate şi mai puţin permeabile ca cernoziomurile tipice şi cambice.
Sunt soluri fertile cu conţinutul de humus de 3-4,5%, reacţia solului este slab acidă în Ap- 6,4-6,6 şi neutră-slab alcalină 7,1-7,4 pe profil, SB este de 20-28 me/100gr.sol, SH-1-8,2me/100gr.sol. Gradul de saturaţie în baze V% este eubazic 80-97% şi mai rar mezobazic cu 70-75%. Aceste însuşiri devin mai puţin favorabile cu creşterea gradului de eroziune şi de pseudogleizare a acestor soluri.
Faeoziomurile argice vertice şi vertice stagnice Sunt formate pe argile şi se evidenţiază în teren prin prezenţa crăpăturilor şi pe
profilul de sol prin existenţa feţelor oblice de alunecare pînă la adâncimea de 1m datorat caracterului vertic.
În perioada cu deficit de umiditate aceste soluri devin compacte şi dure. Profilul de sol este de tipul Ap-Am-ABy-Bty-Cy. Textura solului este luto-argiloasă în Ap şi argilo-lutoasă în Bty iar structura este glomerular-poliedric subangular mic în Am şi poliedric angular mediu în Bty.
Sunt soluri fertile, moderat aprovizionate cu humus în Ap 3,9-4,5%, cu o rezervă foarte mare de humus pe primii 50 cm adâncime, bine aprovizionate cu fosfor mobil 40-70 ppm, bine şi foarte bine aprovizionate cu potasiu mobil 150-250 ppm. Reacţia în Ap este slab acidă 6,4-6,8 şi creşte pe profil. Pe unele suprafeţe agricole unde se folosesc intens îngrăşămintele chimice, pH în Ap este moderat acid.
Însuşirile fizice şi hidrofizice sunt mai puţin favorabile, porozitatea totală Pt este mare la mijlocie în Ap (46-57%) şi mică şi foarte mică în secţiunea de control, porozitatea de aeraţie Pa de la mijlocie la foarte mică în Am (21,8-8,7%) şi foarte mică în secţiunea de control (9,7-4,0%).Coeficientul de ofilire CO variază de la foarte mare 18% la mijlociu 8,6% în Ap şi mare şi foarte mare 13-18% în Bty.

186
Capacitatea de câmp este CC este mijlocie pe tot profilul de sol 23-24%. Permeabilitatea variază de la medie la mare 5,5-12,3 în Ap şi de la mică la excesiv de mică 0,15-5,8 în orizontul Bty şi nu permit o infiltraţie bună pe profil în perioadele umede. Caracterul vertic este reliefat şi de indicele de contracţie Ic care este mare în Ap (0,0081-0,0114) şi de la mare la foarte mare în Bty (0,0088-0,0142).
Pentru corectarea unor însuşiri mai puţin favorabile se recomandă realizarea de lucrări hidroameliorative care constau în folosirea judicioasă a irigaţiilor prin aplicarea de norme mici şi dese, realizarea de canale deschise pentru evacuarea apei din microdepresiuni, iar ca lucrări pedoameliorative afânare adâncă şi arături cu adâncimi variabile şi local amendare calcică.
Faeoziomurile argice sunt soluri favorabile culturilor agricole, gradul de favorabilitate scăzând cu creşterea argilozităţii, a gradului de pseudogleizare şi creşterii eroziunii.
Faeoziomurile greice(FZgr)(Solurile cenuşii de pădure). Prezintă un orizont diagnostic Ame care are atât caractere de orizont molic cât
şi de orizont eluvial prin acumularea de grăunţe de cuarţ care dau un aspect mai albicios acestui orizont
Materialul parental este format din depozite lossoide şi luturi. Spre deosebire de cernoziomurile argice şi argice vertice cu care se învecinează prezintă o tentă mai cenuşie negru-brun cenuşiu foarte închis până la brun cenuşiu foarte închis (10YR 2,5/1,5-10YR 3/1,5). Orizontul Ame prezintă o pudrare de silice şi structură grăunţoasă-poliedric mic subangular, orizontul Bt este mai ferm, cu structură poliedric angular -subangular mare -prismatic. Orizontul Cca apare sub 165 cm şi este bogat în concreţiuni de CaCO3.
Faeoziomurile greice stagnice Ocupă arealele de crovuri unde datorită neuniformităţii terenului, apa din ploi
stagnează o perioadă mai îndelungată şi produce o pseudogleizare slabă-moderată, producând la nivelul orizontului Bt pete de reducere şi un grad mai mare de tasare. Profilul de sol ete de tipul Ap-Am-Ame-ABtw-Btw-Cca
Faeoziomurile greice sunt soluri fertile, mijlociu aprovizionate cu humus în Ap (3,7-3,9), cu o rezervă foarte mare de humus în primi 50 cm ( 218-231t/ha), mijlociu aprovizionate cu fosfor (21,9-26,5 ppm) şi foarte bine aprovizionate cu potasiu (229-260 ppm). Sunt soluri moderat acide în Ap (pH 5,45-5,60 ) şi neutru la slab alcaline în Bt (pH 7,0-7,30). Gradul de saturaţie în baze variază de la mezobazic (73,4 %) la eumezobazic (77,94 %) în Ap şi eumezobazic în Bt cu >90%.
Însuşirile fizice ale acestor soluri sunt relativ favorabile; porozitatea totală variază de la normală în orizontul Ap (51-52%) la mică în Bt (45%), porozitatea de aeraţie este de la mijlocie în Ap (17%) la mică în secţiunea de control (10,0-11,6 %), coeficientul de ofilire variază de la mijlociu la mare în Ap (8,6-14,0 %) şi de la mare la foarte mare în secţiunea de control )15,6-16,5%), capacitatea de câmp este mijlocie pe tot profilul (23 %), permeabilitatea este de asemenea mijlocie pe tot profilul de sol şi variază de la 2,44 mm/h la 7,00 mm/h. Indicele de contracţie este mijlociu în Ame (0,0074) şi mare în Bt (0,0085).
În condiţi normale de expluatare agricolă faeoziomurile greice sunt soluri cu favorabilitate medie pentru agricol.

187
In condiţiile creşterii gradului de pseudogleizare în microdepresiuni favorabilitatea acetor soluri scade. În condiţiile folosiri irigaţiilor şi realizarea unor lucrări pedoameliorative de îmbunătăţire a condiţiilor de aeraţie a solurilor pseudogleizate din microdepresiuni prin canale deschise, afînare adânca, scarificare precum şi aplicarea de amendamente calcice şi îngrăşăminte organice fertilitatea acestor soluri creşte simţitor.
În sfârşit, învelişul de soluri din subetajul fagului este diversificat şi prin efectele eroziunii care, în pajişti, poate avea efecte spectaculoase, apărând soluri divers trunchiate şi chiar roci la zi şi soluri tinere slab evoluate fără a fi propriu-zis erodate, dar împiedicate în evoluţie şi menţinute într-un echilibru precar de câtre un bilanţ de denudare neagresiv. Din aceasta categorie fac parte regosolurile (soluri tinere pe materiale afânate), litosolurile (idem pe roci dure) şi chiar diferitele soluri litice.
Atât solurile erodate cât şi litosolurile şi regosolurile, care au un potenţial foarte mare de eroziune, sunt soluri sărace, dar mai ales soluri care ridica probleme de protecţie şi conservare antierozionala, în special în păşuni.
4.5. CARACTERISTICI GENERALE ALE SOLURILOR Din cele arătate anterior rezulta ca, în limitele spaţiului carpatic din România,
diversitatea tipurilor de soluri şi complexitatea învelişului de sol este diferita în diversele etaje bioclimatice, fiind mai mare în etajul nemoral şi din ce în ce mai mica în etajele superioare, în funcţie de caracterul mai mult sau mai puţin aspru al condiţiilor climatice şi de gama mai bogata sau mai săracă de roci, materiale parentale derivate, forme de relief, condiţii hidrologice şi hidrogeologice, asociaţii vegetale etc.
Cum insa diversele masive, blocuri sau lanţuri muntoase diferă intre ele sub raportul condiţiilor fizico-geografice, mai ales al celor petrografice şi stratigrafice, rezulta ca, atât în ansamblul, cât şi în cadrul aceluiaşi etaj bioclimatic, învelişul de soluri capătă caractere specifice de la un masiv sau grup de masive montane la altul.
Pornind de la acest fapt, în cele ce urmează se face o prezentare succintă a peisajului pedologic, la nivelul marilor subdiviziuni ale Carpaţilor romaneşti - Carpaţii Meridionali, Occidentali şi Orientali - subdiviziuni care au un suport temeinic în aspectele geologico-geografice
4.5.1. Învelişul de soluri al Carpaţilor Orientali
Carpaţii Orientali prezintă, pe întinderi mari, cea mai mare diversitate a învelişului de soluri, având reprezentate toate etajele bioclimatice de la gorun la pajiştile alpine, o gama extraordinara de forme de relief de la cel de câmpie aluvioproluvială, până la cel glaciar, şi un spectru foarte larg de roci eruptive, sedimentare şi metamorfice. Se considera, pentru sistematizarea prezentării pedologice a acestui lanţ muntos, că este utila, mai mult decât subdivizarea lor transversală după criterii geomorfologice, o grupare în raport cu alcătuirea litologică. În funcţie de acest criteriu, în Carpaţii Orientali se disting trei grupe principale: grupa

188
munţilor în roci cristaline, grupa munţilor flişului şi grupa munţilor în roci eruptive. În grupa munţilor flişului se includ şi cele câteva petice de munţi în calcare, molasa etc. La cele de mai sus se mai adaugă ca unităţi distincte seria depresiunilor interne sculptate în roci sedimentare moi sau umplute cu proluvii, aluvii, depozite loessoide etc (DONISĂ,1968).
Cea mai important a grupa de munţi sculptaţi în roci cristaline se constituie ca un nucleu median în Carpaţii Maramureşeni şi cei Moldo-Transilvan. Ea apare ca o imensă culme cuprinzând o bună parte din Munţii Maramureşului, Munţii Rodnei, Suhardului, o bună parte din Munţii Bistriţei şi Munţii Giurghiului şi sfârşeşte la Trotuş.
Sub raport litologic ei prezintă mari afinităţi cu Carpaţii Meridionali, fiind la fel de monotoni ca aceştia, masa lor principală fiind alcătuită din roci intermediare şi acide (şisturi sericitocloritoase, gnaisuri etc.) cu unele intercalaţii locale (semnificative) de şisturi amfibolice, calcare şi dolomite cristaline (SOMEŞAN, 1948).
Este un lanţ puternic dezvoltat atât în suprafaţa cât şi în înălţime. Ei au circa 200 km lungime, lăţimi variabile care pot atinge 50 km (în dreptul Munţilor Rodnei), iar altitudinal ating 2 303 m în Pietrosul Rodnei, punctul cel mai înalt, şi coboară la 1900-1950 m în Munţii Maramureşului şi la 1400-1600 m în Munţii Gurghiului treapta cea mai coborâtă.
Datorita monotoniei lui litologice şi predominării nete a rocilor in-termediare şi acide, acest lanţ montan prezintă condiţii excepţionale pentru manifestarea clara a zonalităţii verticale bioclimatice şi a solurilor. Ei cuprind după cum am arătat toate etajele bioclimatice, de la pădurile de fag la pajiştile alpine, cu toata gama solurilor zonale proprii etajelor respective, de la solurile brune acide până la podzoluri şi humosiosoluri.
Condiţiile climatice mai aspre determină coborârea limitelor etajelor bioclimatice şi de sol, extinderea neobişnuita a pădurilor de conifere şi realizarea morfologică. excepţionala a podzolurilor începând de la altitudini joase, de 1 650-1 700 m ( BARBU,1976).
Învelişul de soluri este şi el monoton, fiind constituit în proporţii de masa din soluri zonale, cu variaţii numai în privinţa grosimii şi caracterului scheletic, ale eroziunii şi alternanţei cu stâncării. Monotonia este întreruptă doar la nivelul suprafeţelor cu calcare şi dolomite, pe care se dezvolta soluri de tip rendzinic.
În cuprinsul Carpaţilor Orientali, mai apar munţi în roci cristaline, cu tot cortegiul problemelor pedologice arătate, 90 km mai la sud, în Perşani şi Leaota. Problemele de agrochimie, potenţial productiv, ameliorare etc., ca şi cele de extrapolare a datelor pedologice din câmpurile experimentale, sunt aceleaşi cu cele din Carpaţii Meridionali. Experienţa din Meridionali poate fi transferată în lanţul cristalin nordic sau invers.
În cuprinsul Carpaţilor Orientali grupa munţilor sculptaţi în depozite de fliş (la care se adaugă şi peticele cu munţi în calcare, depozite de molasa etc.) are ponderea cea mai mare şi prezintă învelişul de sol cel mai complex.

189
Se distinge un fliş intern cretacic constituit predominant din roci ultrabazice frecvent cu carbonaţi şi de constituţii fizice foarte diferite (strate de Sinaia din marne, marne şi calcare, gresii calcaroase, brecii polimictice etc.), un fliş median şistos-grezos intermediar sau bazic şi un fliş extern dominat de roci acide şi intermediare (fliş grezos-şistos. fliş bituminos cu gresii de Kliwa, facies conglomeratic, facies grezos cu gresie de Tarcău etc.). Formaţiunile calcaroase se asociaza mai mult cu flişul intern.
Aparţin flişului intern şi median Culmea Stânişoarei (partial), Ceahlaul, Hăşmaşul, Munţii Ciucului, ai Bodocului şi Baraoltului (din Carpatii Moldo-Transilvani), precum şi Ciucaşul, Munţii Baiului, ai Braşovului şi Perşani (din Carpaţii de curbura), la care se mai adaugă masivele Bucegilor şi Pietrei Craiului, care fac tranziţia la Carpaţii Meridionali (BĂCĂINŢAN,1999).
Flişul extern cuprinde Obcinile Bucovinene (din ramura nordica a Carpaţilor Orientali), Culmea Stânişoara (parţial), Munţii Tarcăului, ai Oituzului, ai Vrancei şi ai Buzăului (BÂLTEANU,1983).
În sfârşit, depozite intermediare de fliş mai apar şi pe latura transilvană a Carpaţilor Maramureşeni, în Munţii Bărgăului şi cea mai mare parte a Ţibleşului.
În vastitatea acestui teritoriu se realizează altitudini de peste 2 000 m numai în Bucegi şi Piatra Craiului (relief glaciar numai în Bucegi), restul munţilor atingând 1 800 m, maximum 1 900 m, numai câteva masive (Ciucaş, Zaganu, Postavaru, Piatra Mare, Siriu, Penteleu şi Carpaţii Curburii, Ceahlăul şi Hăşmaşul in Carpaţii Moldo-Transilvani). În schimb, daca nu au relief glaciar, relieful lor fluvio-torential este extrem de variat, de la formele domoale generate de depozitele mai argiloase, la cele semeţe cu pante abrupte date de gresii şi conglomerate şi, în sfârşit, până la relieful calcaros. Stratigrafia flişului şi rezistenta foarte diferita a rocilor la eroziune au favorizat în zonele neîmpădurite dezvoltarea larga a fenomenelor de panta, cu toate formele generate de eroziune şi alunecări (COTEŢ 1969, 1973). În ceea ce priveşte condiţiile bioclimatice, se remarca faptul că numai Bucegii prezintă toată gama etajelor de la pădurea de fag la cel alpin, restul munţilor în fliş depăşesc limita pădurilor numai în cele câteva mici masive enumerate anterior, care ajung la 1 700-1 900 m, în Carpaţii Curburii pădurile de molid ocupa suprafeţe relativ mici, ele regăsindu-se insa pe suprafeţe mari, mai ales pe munţii flişului intern al Carpaţilor Moldo-Transilvani, o menţiune specială trebuie făcută în legătură cu coborârea puternica extrazonală a molidului şi paj iştilor din acest etaj, pe versanţii vestici ai munţilor flişului intern, la contactul cu depresiunile Ciucurilor şi Gheorghienilor, în legătura cu inversiunile de temperatura (TUFESCU,1966).
Solurile depind în foarte mare măsura de roci şi, datorita acestui fapt, diversitatea lor este mare, dar se desfăşoară pe areale largi, se remarca faptul ca pe flişul intern şi median, dominant ultrabazic şi bazic, au o larga răspândire până la 900-1 000 m altitudine, chiar sub pădurile extrazonale de molid, eutricambosoluri, luvisoluri, uneori solurile brune slab-mediu luvice, iar dintre solurile brune domina cele acide de limita spre mezobazic sau chiar cele mezobazice (V = 40-60%). Pe calcare şi marne apar rendzinel şi eutricambosoluri, molice.

190
În limitele aceluiaşi etaj nemoral, pe flişul extern, domina districambosolurile, oligomezobazice, numai pe gresia de Kliwa şi uneori pe cea de Tarcau districambosolurile oligobazice sau extrem oligobazice şi chiar criptospodice. Comparativ, solurile etajului nemoral din munţii flişului sunt mult mai saturate şi mai profunde decât cele din acelaşi etaj, pe cristalin (BĂLĂCEANU şi MARIAN 1985).
Către limita de sus a etajului capătă preponderenţa districambosolurile tipice, dar pe flişul intern se mai menţin încă suprafeţe notabile cu termeni de limita mezobazici ai acestora şi chiar cu eutricambosoluri. Pe gresii şi conglomerate, chiar pe cele cu ciment sau elemente calcaroase, domina districambosolurile care în multe situaţii sunt semilitice, Frecvent, pe gresii cu ciment calcaros, sub asociaţii de Nardus stricta si Festuca rubra se întâlnesc humosiosoluri. Începând din etajul pădurilor de molid, în sus, complexitatea înveli-şului de sol se restrânge, în limitele flişului intern, pe roci carbonatice şi calcare mai apar încă eutricambosoluri şi rendzine, dar în rest domina net solurile prepodzolice.
În pajiştile din boreal şi alpin inferior se întâlnesc spodosoluri, iar sub pajiştile vechi, soluri criptospodice. Podzoluri bine dezvoltate apar doar pe conglomerate şi gresii, în Bucegi şi Ceahlău. Aşa cum am arătat deja, numai Munţii Bucegi prezintă o zonă alpina bine dezvoltată, cu podzoluri tipice şi feriluviale cu învelişul de sol fragmentat prin acţiunea eroziva a vânturilor, zăpezilor şi apei (PUŞCARU, 1956).
Problema aprecierii capacitaţii productive a terenurilor în raport cu solul este foarte complexa, mai ales în subetajul făgetelor, şi se poate rezolva numai pe baza unui inventar detaliat al acestora, materializat pe hărţi la scara mare sau mijlocie.
Învelişul de soluri al munţilor în roci eruptive din Carpaţii Orientali prezintă o monotonie litologica mai pronunţata chiar decât cea a Carpaţilor Meridionali. Ei sunt, în proporţie covârşitoare, constituiţi din andezite cu piroxeni (bazaltoide) sau cu piroxeni şi amfiboli şi din piroclastite ale acestora, care se comporta ca roci intermediare şi mai ales bazice.
În cadrul ramurii vulcanice au dezvoltare mare în suprafaţa Munţii Gutâi şi mai ales lanţul Călimani-Gurghiu-Harghita. Altitudinal Munţii Gutâi ajung la 1 444 m, menţinându-se în limitele etajelor forestiere (dar având în pajiştile superioare şi accente de subalpin), iar ceilalţi la 2 100 m în Călimani şi 1 800 m în Harghita, prezentând un sector subalpin relativ puternic. Ei prezintă diverse nivele largi relativ plate, dar între ele au suprafeţe de racord şi versanţi foarte puternic înc1inati.
Favorizată de uniformitatea geologica, zonalitatea bioc1imatica este clasica şi clara, răsfrângându-se şi asupra învelişului de sol care trece prin toate etajele, de la eutricambosoluri districe specific pădurilor de fag, până la podzolul humicoferiiluvial specific jnepenişurilor. Determinate de caracterul mai bazic al rocilor, preluvosolurile tipice au, până la altitudini de 800-900 m caractere de limita (oligomezobazic - mezobazic, respectiv V = 50-60%). Particularitatea esenţiala însa a solurilor, derivata din specificul mineralogic al materialelor parentale, este caracterul andic al unei mari parţi a eutricambosoluri districe (solurilor brune acide) şi chiar al unei parţi din spodosoluri. Mai mult încă se remarcă prezenţa andosolurilor, pe suprafeţe relativ semnificative, mai ales la limita superioară a făgetelor.

191
În legătură cu înclinarea puternică a versanţilor, foarte multe soluri din aceste masive au caracter litic sau rămân la stadiul de litosol distric. În zonele înalte, lipsa reliefului glaciar aduce de asemenea o nota de monotonie în învelişul de sol, care este mai puţin sfârtecat.
Pe treptele inferioare, pe vulcanogen sedimentar, se întâlnesc stagnogleiuri luvice.
Din punct de vedere al fertilităţii, problemele solurilor acestor masive sunt similare cu cele din cristalin, cu menţiunea că ele sunt totuşi mai puţin acide şi debazificate. În plus, la andosoluri şi solurile andice se manifestă o capacitate mare de fixare, în forme greu accesibile pentru plante, a ionilor fosfat, molibdat, borat, iar în unele situaţii pot apărea cantităţi mari de Mg, care inhibă nutriţia cu Ca etc.
În spaţiul geografic al Carpaţilor Orientali, depresiunile interne, profund umanizate, aduc o nota cu totul aparte în peisajul geografic, pedologic şi economic.
Sub raportul condiţiilor fizico-geografice, se disting doua grupe de depresiuni, una a celor cu relief fragmentat (Maramureş, Bilbor, Borsec, Glod, Întorsura Buzăului) şi alta a celor cu relief plat de câmpie piemontana (Gheorgheni, Ciuc, Braşov-Tg.Secuiesc).
În general în depresiunile cu aspect deluros se întâlnesc luvosoluri pseudogleizate, luvisoluri albice pseudogleizate, stagnosoluri (pe terase). O nota aparte prezintă marea depresiune a Maramureşului, care are un relief foarte complex, cu piemonturi în structura cărora se întâlnesc depozite provenite din rocile vulcanice, metamorfice sau chiar sediment are din jur, dealuri cu un mozaic foarte mare de roci (argile, marne - uneori salifere - conglomerate) şi în sfârşit terase. Legat de aceasta diversitate a litologiei, apare o gama foarte larga de soluri, anume: luvosoluri pseudogleizate, luvosoluri albice stagnogleice, uneori holoacide şi stagnosoluri pe terase, eutricambosoluri frecvente pe reliefuri cu alunecări, în complex cu soluri erodate şi regosoluri, districambosoluri, şi salsodisoluri pe ivirile de sare de pe unii versanţi înalţi. În sectoarele umede ale luncilor, din zonele înalte se întâlnesc gleiosolurile.
Depresiunile plate au un înveliş de soluri mai uniform, cu luvosoluri stagnice pe treptele vechi, districambosoluri sau eutricambosoluri prundice pe conurile de împrăştiere recente (dupa cum substratul este silicatic sau calcaros) şi gleiosoluri tipice şi cernice la capetele conurilor de dejecţie şi în lunci.
Pajiştile ocupă în general terenurile cu alunecări cu soluri erodate sau eutricambosoluri, piemonturile cu districambosoluri sau luvosoluri stagnice pseudogleizate şi mai ales terenurile cu soluri hidromorfe.
Problemele lor agrochimice, ameliorative etc. sunt cele specifice tuturor lanţurilor muntoase (SPIRESCU şi colab., 1985).
4.5.1. Învelişul de soluri al Carpaţilor Meridionali
Carpaţii Meridionali cuprind trei blocuri montane: Făgăraş, Paring, Retezat, despărţite prin culoare relativ largi şi depresiuni interne (Caransebeş-Mehadiea, Hateg, Petroşani, Titeşti). Din punct de vedere petrografic, aceşti munţi sunt extrem de simpli si monotoni, în constituţia lor predominând net şisturile cristaline

192
intermediare şi acide, străpunse pe alocuri de mase intruzive granitice şi gnaise de injecţie (acide şi ele) şi, local, de calcare cristaline. La poalele munţilor apar uneori brecii sau conglomerate, în general oligomictice, remaniate şi ele din cristalin, sau conuri cu proluvii din aceleaşi roci. Monotonia petrografica este întreruptă numai în etajul pădurilor de foioase de o cuvertura de calcare aşternută peste cristalin în Munţii Vilcanului, de depozite cuaternare fine pe terasele pleistocene din depresiunile Haţegului, Caransebeşului şi Petroşanilor şi de depozite moi marno-argiloase în Depresiunea Titeştilor. Comparate cu şisturile cristaline, toate aceste roci moi, ultrabazice - carbonatice etc. ocupa suprafeţe mici.
Ramura carpatica meridionala este cea mai înalta din România şi cuprinde cel mai vast şi mai bine dezvoltat spaţiu alpin. Relieful sau, monoton şi el din cauza structurii geologice uniforme, are ca trăsături particulare de detaliu, pe de o parte dezvoltarea puternica a platformelor de eroziune (care au favorizat un înveliş continuu de sol şi vegetaţie şi cel mai puternic domeniu pastoral din tara), şi pe de alta parte conservarea excepţionala a formelor reliefului glaciar cuaternar, complicat prin suprapunerea unui alt relief actual crionival, dinamic.
Bioclimatic, se înregistrează întreaga gama a etajelor din Carpaţi, cu menţiunea ca zonalitatea este mai clar exprimata decât oriunde, fiind favorizata de monotonia petrografica. În mod particular efectul de "fata" şi "dos" este foarte puternic, determinând diferenţe de peste 100 m între limitele etajelor bioclimatice pe clinele de nord şi sud ale lanţului montan. Limita superioara a pădurilor este cu toate acestea mai joasa (uneori chiar 1 500-1 600 m) pe povârnişurile sudice, din cauza despăduririlor efectuate în scopul extinderii pajiştilor, fapt cu implicaţii în învelişul de sol. Reflectând monotonia petrografica şi etajarea bioclimatica foarte evidenta, învelişul de sol al spaţiului montan propriu-zis este simplu şi monoton şi el, şi dominat net de solurile zonale, exprimând pregnant zonalitatea verticala. Subetajului pădurilor de fag îi corespunde eutricambosolurile districe (solul brun acid) în ipostazele cele mai debazificate şi mai acide, etajului molidului solul brun feriiluvial şi podzolul brun, jneapănului podzolul, iar caricetelor, solul humicosilicatic litic.
Pe clina nordica, coboară frecvent molidul şi ericaceele în etajul fagului şi, odată cu ele, şi spodosolurile. Variaţiile la nivelul solurilor zonale ţin mai curând de grosimea şi caracterul lor mai mult sau mai puţin scheletic. Ca regula generală, pe conglomeratele din zonele mai joase, solurile sunt mai profunde (semi-litice), în timp ce pe rocile dure sunt litice, asociate adeseori cu litosoluri. În etajul alpin numai platformele au înveliş de sol continuu, În rest, învelişul de soluri este sfârtecat de stâncării reziduale sau îngrămădite sub forma de grohotişuri la baza versanţilor.
În cadrul reliefului montan, monotonia peisajului pedologic este întreruptă de solurile intrazonale. Pe ca1carele cristaline, care se întâlnesc în mai toate etajele bioclimatice, apar rendzine, soluri rendzinice şi humosiosoluri (soluri humico-silicatice) frecvent acidifiate în orizonturile superioare, iar pe ca1carele jurasice din Vâlcan sau de pe rama sudica dinspre Depresiunea Haţegului a Munţilor Şureanului, apar rendzine, eutricambosoluri rodice şi uneori, pe argile reziduale din ca1care, eutricambosoluri. De reţinut însa ca ponderea acestor soluri în totalul suprafeţei Carpaţilor Meridionali este neînsemnata.

193
În sfârşit, o nota de variaţie aduc şi depresiunile interne care apar în subetajul gorunului. În toate depresiunile, cu excepţia celei a Titeştilor domina relieful de terase şi glacisurile piemontane vechi, acoperite respectiv cu luvosoluri tipice şi albice stagnice şi cu alosoluri tipice. În Depresiunea Titeştilor, cu aspect deluros, domina eutricambosolurile eutrice şi districe pe marne şi argile, adeseori asociate cu gleiosoluri de panta, pe reliefuri cu alunecări.
În final trebuie reţinut un lucru esenţial, anume ca învelişul de soluri al Carpaţilor Meridionali este în proporţie de 90-95% constituit din soluri zonale, că în privinţa solurilor zonale este provincia cu cele mai acide şi mai debazificate soluri din ţara, ca toate solurile zonale sunt slab aprovizionate cu elemente nutritive şi adeseori sunt aprovizionate la limita spre insuficient cu microelemente, în sfârşit, ca ele reclama amendamente şi îngrăşăminte care însa, aşa cum s-a arătat, trebuie aplicate cu multă grijă pentru a nu provoca dezechilibre, şi numai acolo unde se justifica prin sporuri de recolte, în arealele mai joase.
Uniformitatea învelişului de sol are avantajul valabilităţii largi şi al posibilităţilor de extrapolare pe întinderi mari a rezultatelor obţinute în câmpurile experimentale, cu condiţia respectării zonelor bioclimatice.
4.5.2. Învelişul de soluri al Carpaţilor Occidentali
Judecaţi prin prisma aspectelor pedologice, Carpaţii Occidentali pot fi subdivizaţi în doua grupe: grupa Munţilor Banatului şi Poiana Rusca şi grupa Munţilor Apuseni.
Munţii Banatului şi Poiana Rusca prezintă mari similitudini cu Carpaţii Meridionali, atât din punct de vedere petrografic cât şi pedologic. Ei sunt constituiţi, în principal, din şisturi cristaline şi graniţe, cu unele intruziuni de ca1care cristaline, mai importante în Poiana Rusca, şi cu o cuvertura de ca1care jurasice în Semenic-Almaj. Altitudinal însă, ei sunt mai coborâţi decât Carpaţii Meridionali cu 1 000-1 200 m, ajungând la 1 400 m în Semenic, 1 220 m în Munţii Almajului şi la 1 374 m în Poiana Rusca. Relieful lor de detaliu se reduce la formele provocate de eroziunea fluviatila. Ei aparţin aproape în întregime subetajului bioclimatic al fagului, pădurea de molid ocupând suprafeţe mici, numai în Semenic şi Poiana Rusca. Circa 70-80% din suprafaţa acestor munţi este acoperită cu districambosoluri în general tipice, oligomezobazice (V=30-50%) şi mai puţin criptopodzolice. Spodosolurile specifice pădurilor de molid ocupa suprafeţe mici în munţii amintiţi, la peste 1 300 m altitudine. Calcarelor cristaline din Poiana Rusca le corespund eutricambosoluri molice şi litice şi mai puţin rendzine, iar calcarelor jurasice din Munţii Banatului, rendzine si uneori eutricambosoluri roşcate. În depresiunile şi culoarele intramontane (Bozovici, Caransebes, Mehadica etc.) domină luvosolurile tipice şi albice stagnogleice.Majoritatea solurilor sunt litice sau cu roca dură aproape de suprafaţă.. Eroziunea prezintă importanţă mai ales în jurul satelor puternic insinuate în spaţiul montan, şi în legătură cu exploatările miniere. Din punct de vedere agrochimic şi al extrapolării datelor din câmpurile experimentale, aceleaşi menţiuni ca în Carpaţii Meridionali. Rezultatele din câmpurile de la Crint-Cibin pot fi extrapolate în Poiana Rusca sau oriunde în solurile pe cristalin, în limitele aceluiaşi etaj bioclimatic.

194
Munţii Apuseni prezintă cea mai mare diversitate de roci şi tipuri de soluri pe unitatea de suprafaţa din întregul lanţ carpatic.
În sectorul nord-nord estic al acestor munţi, se afla un pivot puternic al Bihariei - constituit din patru masive cu altitudini comparabile (Muntele Mare - 1 826 m, Bihorul, Biharia sau Curcubata - 1 849 m şi Vlădeasa 1 836m). Acest nucleu este constituit, predominant, din şisturi cristaline şi graniţe, roci acide şi intermediare care favorizează dezvoltarea solurilor zonale, dar şi din calcare jurasice care rup monotonia petrografica pe suprafeţe relativ mari, creând un peisaj geologic şi pedologic aparte.
În relieful acestui nucleu se deosebeşte o regiune de poduri netede cu martori mai înalţi de forma cornetelor (Vlădeasa), o mare platforma de eroziune la peste 1 700 m altitudine (Muntele Mare), un sector cu relief carstic (Cetăţile Ponorului) şi o prispa mai joasa ce coboară sub 1200-1 300 m (Munceii Dealului).
Vegetaţia cunoaşte o repartiţie zonala, cu menţinerea dominării pădurilor de fag, a restrângerii ariei pădurilor de molid şi a dezvoltării largi, pe poduri şi platforme, a pajiştilor secundare. În relaţie cu condiţiile bioclimatice, solurile cunosc, pe rocile cristaline, aceeaşi repartiţie altitudinala ca în Poiana Rusca. Domina districambosoluri iar prepodzolurile se dezvolta pe suprafeţe restrânse în jurul înălţimilor mai mari. Podzolurile nu apar în acest masiv. Pe suprafeţele plane ale platformelor învelişul de sol este uniform, în timp ce pe versanţi şi mameloane intervin eroziuni, iar solurile au caracter litic. Pe calcarele de la Ponoare apar soluri cutricambosoluri şi rendzine. Tot restul Munţilor Apuseni se dezvolta pe laturile Masivului Bihariei sub forma unor trepte joase tentaculare, în general sub 1 000 m altitudine (dar ajungând uneori la aproape 1 400 m) despărţite prin depresiuni insinuate în perimetrul montan sub forma unor golfuri constituite din câmpii cu terase, profund umanizate şi cultivate agricol. În limitele altitudinilor arătate, toţi aceşti munţi se menţin în etajul pădurilor de foioase, cu incluziuni ale subetajului gorunului, de-a lungul golfurilor depresionare.
În acest context, problema învelişului de soluri nu mai este o chestiune de zonalitate, ci de relaţie roca-relief-sol. În linii mari, situaţiile pedologice în raport cu roca şi relieful sunt următoarele:
golfurile depresionare cele mai importante (Vad-Borod-Beiuş-Zarand) sunt formate din terase mărginite de piemonturi. Acestea sunt constituite la partea superioara, din depozite fine din care cauză se întâlnesc dominant luvosoluri tipice şi albice în general pseudogleizate şi uneori stagnogleiuri;
ramura nordică a Mezeşului şi Muntelui Şes, care se prezintă sub forma unor trepte joase de cca. 900 m fiind constituită din şisturi cristaline, este acoperită cu districambosoluri şi luvosoluri tipice şi uneori alosoluri.
munţii Pădurea Craiului, o masa de calcare mezozoice compacte, acoperite către nord-vest cu fliş cretacic, împreună cu Munceii Codru Moma (< 1 000 m altitudine), care sunt o asociere complicată de formaţiuni permo-carbonifere, fliş cretacic, calcar, andezite etc., prezintă un înveliş de sol extrem de eterogen, cu districambosoluri şi alosoluri, districambosoluri andice, eutricambosoluri şi rendzine etc.;

195
Munceii Zarandului cuprind un compartiment jos (900 m) format din şisturi cristaline, cu districambosoluri şi luvosoluri tipice (Munţii Highiş) şi unul mai înalt, mai complicat, cu fliş cretacic, piroclastite andezitice şi un complex melafiric bazic, pe care s-au dezvoltat, districambosoluri tipice şi districambosoluri andice şi cutricambosoluri, asociate cu preluvosoluri tipice şi litosoluri;
Munţii Metaliferi constituie compartimentul cel mai complex sub toate raporturile. Se caracterizează, din punct de vedere geologic, prin prezenţa a patru complexe litologice, cu rol morfologic şi pedologic diferit: ofiolite mezobazice, calcare jurasico-cretacice (mai ales în Munţii Trascăului), fliş şi eruptiv neogen (VESPREMEANU, 1998). Ele determina forme de relief foarte diferite. Se întâlnesc relief calcaros cu abrupturi spectaculoase golaşe şi poduri cu relief carstic foarte divers, relieful de fliş prezintă forme domoale generate de argile, un relief mai energic dat de gresii şi conglomerate etc. şi în sfârşit cupole de corpuri intrusive dezvelite de eroziune sau platouri mărginite de versanţi abrupţi etc.
Solurile cunosc o diversitate mare de tipuri genetice prezentând o mare varietate ca succesiune şi asociere în spaţiu, deoarece nu numai rocile sunt diferite, dar şi stratigrafia. Se întâlnesc districambosoluri în toate stadiile de debazificare (eutrice şi districe), districambosoluri andice, luvosoluri tipice, alosoluri, nigrosoluri, eutricambosoluri şi rendzine etc. La toate tipurile şi subtipurile arătate mai sus se mai adăugă diversitatea solurilor din culoarele depresionare, sculptate în roci moi şi cea produsă de fenomenele de pantă, frecvente mai ales în depozitele de fliş şi în sectoarele intens umanizate.
Dincolo de varietatea învelişului de sol, apar şi diferenţe de la un loc la altul în ceea ce priveşte aspectele chimice, mineralogice şi fizice, chiar în cadrul aceluiaşi tip sau subtip de sol (BĂLĂCEANU şi MARIAN 1985). În contextul acestei bogăţii de soluri şi a complexităţii deosebite a învelişului de sol, rezultatele obţinute pe solurile din câmpurile experimentale cu pajişti trebuiesc extrapolate cu mult discernământ şi numai pe baza cunoaşterii detaliate a solurilor sub toate aspectele, inclusiv al repartiţiei lor teritoriale. 4.6. REZULTATE DE CERCETARE DIN PUNCTELE CHEIE DE OBSERVAŢII
Solurile regiunilor de munte şi repartizarea lor geografică prezintă o serie de particularităţi prin care se deosebesc de solurile de câmpie şi dealuri.
În regiunea de munte, factorul bioclimatic suferă schimbări rapide, determinate în primul rând de schimbările de altitudine. În mod corespunzător se schimbă solurile, care se succed într-o zonalitate verticală specifică regiunilor montane.
Relieful şi roca intervin, la rândul lor, determinând formarea solurilor de profunzime foarte variată (în general redusă) şi cu caracter scheletic.
În raport cu alcătuirea chimic-mineralogică a rocilor, profunzimea solurilor – în aceleaşi condiţii de relief – poate fi mai mare sau mai mică, iar caracterul de sol scheletic mai slab sau mai puternic. De exemplu, solurile formate în zona flişului carpatic pe gresii slab cimentate sunt mai profunde şi mai slab scheletice ca solurile formate pe roci eruptive sau cristaline mai greu alterabile.

196
Diversitatate pedologică a zonei montane este evidenţiată de hărţile de soluri, în acest scop folosindu-se harta solurilor României, scara 1:1 000 000. Pe baza acestei harti, s-au constituit comunitati de soluri (UPAS) adiacente cu proprietăţi suficient de asemănătoare încât se comportă similar la tehnologii de ameliorare, lucrări ale solului, de fertilizare sau de protecţie. Conceptul nu este nou, a fost introdus ca unitate pedoagrotehnologică – UPAT – de către GHEORGHE ŞI FLOREA, 2006, GHEORGHE, 2007 şi reprezintă unitatea de bază în aplicarea unei agriculturii specifice la nivel local-regional, diferenţiată în teritoriu. Un UPAS este o unitate teritorială cu acelaşi specific calitativ din punct de vedere agricol, care se comportă, răspunde sau evoluează relativ în acelaşi mod la aplicarea unei anumite tehnologii agricole.
Conceptul a fost aplicat la Câmpia Română de Est (GHEORGHE, 2007) pentru condiţii de soluri variate din climat semiarid-semiumed, distingându-se 22 de UPAT-uri. În 2008 conceptul a fost extins la teritoriul agricol al întregii ţări, definindu-se 34 UPAT-uri, precizându-se cerinţele lor agrotehnologice (FLOREA şi GHEORGHE, 2007).
Aplicând criteriile la întreaga suprafaţă a ţării (agricolă şi neagricolă), s-a ajuns la concluzia că unitatea delimitată astfel este mai potrivit să fie denumită unitate de pedopeisaj cu atribute specifice (UPAS), deorece multe unităţi delimitate astfel nu au folosinţă agricolă, cerinţele lor de utilizare fiind de alt ordin.
Criteriile de definire (şi de delimitare în acelaşi timp) sunt reprezentate de caracteristicile de sol ce rezultă din hărţile de soluri şi rapoartele explicative însoţitoare.
Unul din criteriile importante este cel al grupărilor genetice de soluri în funcţie de comportamentul lor similar la diferite folosinţe şi în special la cea agricolă, determinat de conţinutul şi rezerva de materie organică, gradul de dezvoltare a solului şi de diferenţiere pedogenetică (Tabelul 4.1 şi Fig. 4.1)
La acest criteriu, s-au adăugat următoarele: Reacţia solului cu trei subdiviziuni: bazică, neutră-slab acidă, acidă; Textura solului în orizontul superior, diferenţiată în trei clase: grosieră,
mijlocie şi fină; Volumul edafic cu trei subdiviziuni; Excesul de umiditate şi natura lui; Gradul de sărăturare şi natura acesteia (salinizare, alcalizare); Înclinarea terenului şi riscul de eroziune; Prezenţa sau pericolul de alunecări. După aceste criterii, solurile de munte s-au divizat în alte opt UPAS-uri de nivel
superior (Tabelul 4.2 şi Fig. 4.2). Utilizând criteriile de definire şi delimitare ale UPAS-urilor elaborate de
FLOREA şi colab, 2009, au fost separate în final 22 UPAS-uri pentru zona de munte (Tabelul 4.3 şi Fig.4.3). În final se prezintă schema cheie de definire a UPAS-urilor montane (Tabelul 4.4).
197
Tabel 4.1 Distribuţia solurilor din zona de munte în functie de caracteristicile
lor principale şi de natura limitărilor Soil distribution from mountainous area related
to principal characteristics and limit types
Cod Nr. Suprafata poligoane ha %
A – UPAS-uri tabulare sau slab ondulate cu soluri profunde fără limitări majore
96 200022 3.47
B – UPAS-uri tabulare cu soluri profunde cu limitări datorită excesului de apă şi/sau sărăturării
91 165383 2.87
C – UPAS-uri cu soluri profunde cu limitări datorită reliefului
363 5229230 90.76
D – UPAT-uri cu soluri având alte limitări (volum edafic, turbă, rocă la zi).
24 159961 2.78
Lacuri 2 7250 0.13T O T A L 576 5761846 100.00
Tabel 4.2
Distribuţia solurilor din zona de munte in UPAS-uri la nivel superior Distribution of soil from mountainous area in UPAS-es at superior level
Cod Nr.
poligoane Suprafata ha %
100 - Pedopeisaje tabulare sau slab ondulate, cu soluri profunde cu textură mijlocie
85 192033 3.33
300 - Pedopeisaje (UPAS-uri) tabulare sau slab ondulate cu textură fină
11 7989 0.14
400 - Pedopeisaje (UPAS-uri) tabulare cu soluri profunde afectate de exces de umiditate freatic sau stagnant
91 165383 2.87
600 - Pedopeisaje (UPAS-uri) cu soluri profunde fără alunecări
276 4249886 73.76
700- Pedopeisaje (UPAS-uri) cu soluri profunde pe terenuri cu alunecări
86 976165 16.94
800 - Pedopeisaje (UPAS-uri) cu soluri scheletice şi /sau puţin profunde sau extrem de nisipoase
22 159763 2.77
900 - Pedopeisaje (UPAS-uri) cu soluri organice 3 3378 0.061000 - Lacuri 2 7250 0.13
T O T A L 576 5761847 100.00
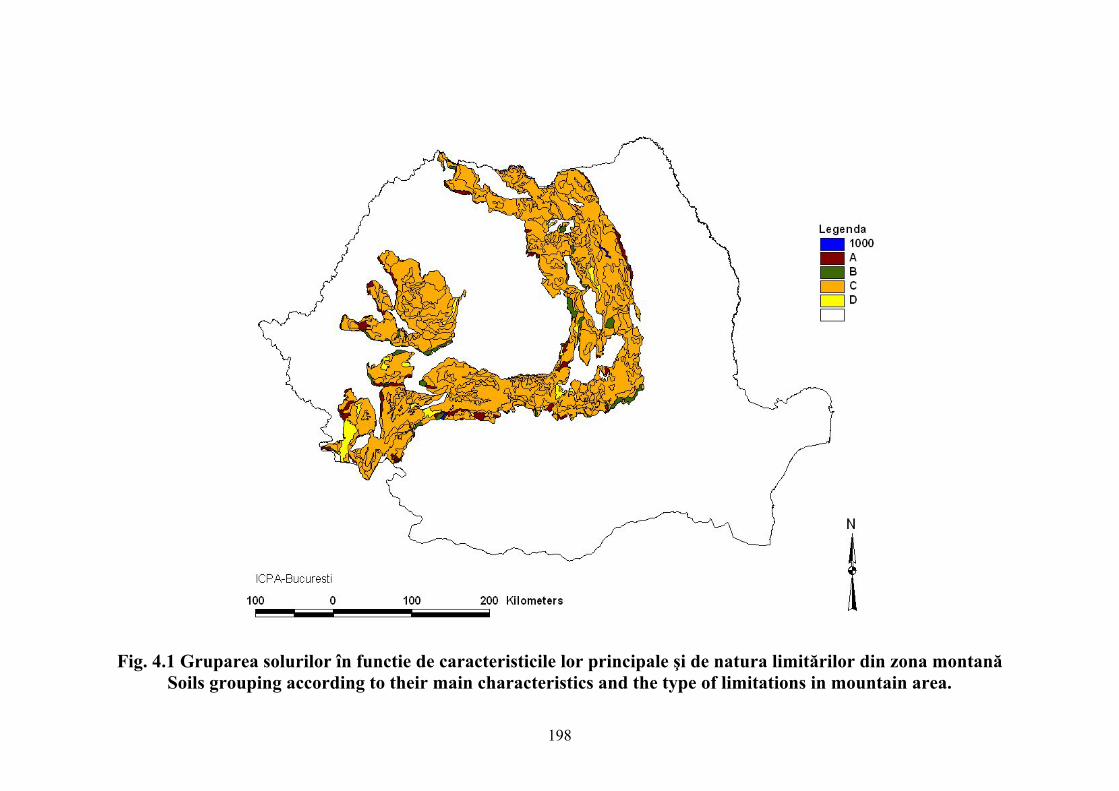
198
Fig. 4.1 Gruparea solurilor în functie de caracteristicile lor principale şi de natura limitărilor din zona montană
Soils grouping according to their main characteristics and the type of limitations in mountain area.

199
Fig. 4.2. Gruparea solurilor din zona de munte in UPAS-uri la nivel superior
Grouping of the soil from the mountain area in UPAS from high level
200
Tabel 4.3 Distribuţia solurilor din zona de munte in Unitati de Pedopeisaj Soil distribution from mountainous area in Pedolandscape units
COD Nr. polig.
Suprafataha %
112 - Soluri cernoziomice bine drenate, cu reacţie neutră-slab acidă (cernoziomuri, faeoziomuri) 10 8209 0.14
113 - Soluri cernoziomice bine drenate, cu reacţie bazică (kastanoziomuri, cernoziomuri calcarice) 1 3184 0.06
122 - Soluri preluvice şi cambice, drenate, cu reacţie neutră-slab acidă 9 12801 0.22
131 - Soluri luvice cu drenaj subteran bun, cu reacţie acidă 65 167838 2.91
312 - Soluri cernoziomice (faeoziomuri) şi preluvosoluri (cu textură mijlocie-fină) bine drenate, cu reacţie neutră la slab acidă
7 3055 0.05
322 - Pelosoluri (vertosoluri) şi soluri pelice cu drenaj subteran relativ bun, cu reacţie neutră-slab acidă 4 4934 0.09
412 - Gleiosoluri şi soluri gleice cu reacţie neutră la slab acidă 5 700 0.01
421 - Stagnosoluri şi soluri stagnice cu reacţie acidă 47 78178 1.36422 - Stagnosoluri, planosoluri şi soluri stagnice cu reacţie neutră la slab-acidă 2 395 0.01
432 - Aluviosoluri şi soluri aluvice cu reacţie neutră slab-acidă. 19 35072 0.61
433 - Aluviosoluri şi soluri aluvice cu reacţie bazică 18 51038 0.89610 - Soluri variate (cambice, luvice) pe versanţi slab moderat înclinaţi 101 488576 8.48
620 - Soluri cambice (cambisoluri, andosoluri) pe versanţi slab la puternic înclinaţi 112 2915473 50.6
630 - Soluri spodice (prepodzoluri, podzoluri, pe versanţi slab la puternic înclinaţi 54 810376 14.06
650 - Soluri de goluri alpine în teritorii puternic accidentate (humosiosoluri, criptopodzoluri, litosoluri) 7 34188 0.59
690 - Soluri puternic erodate pe pante accentuate 3 4453 0.08702 - Faeoziomuri, eutricambosoluri şi soluri erodate pe relief vălurit 4 22332 0.39
704 - Faeoziomuri şi soluri erodate pe relief ondulat 1 17 0 - Soluri variate pe relief ondulat la accidentat, local exces de umiditate 81 953817 16.55
811 - Soluri scheletice pe profil 1 123 0812 - Soluri cu profil scurt (litosoluri, rendzine, soluri litice ) 20 156460 2.72
910- Histosoluri districe (acide) 3 3378 0.061000 - Lacuri 2 7250 0.13
T O T A L 576 5761847 100
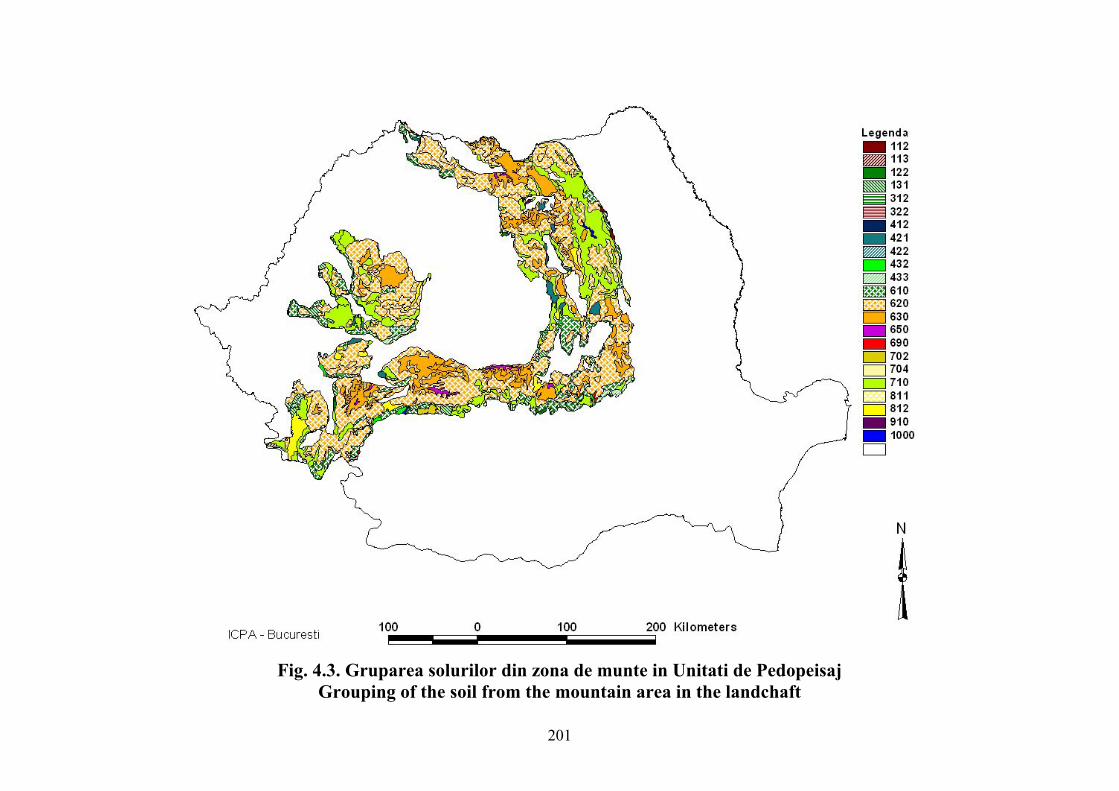
201
Fig. 4.3. Gruparea solurilor din zona de munte in Unitati de Pedopeisaj
Grouping of the soil from the mountain area in the landchaft
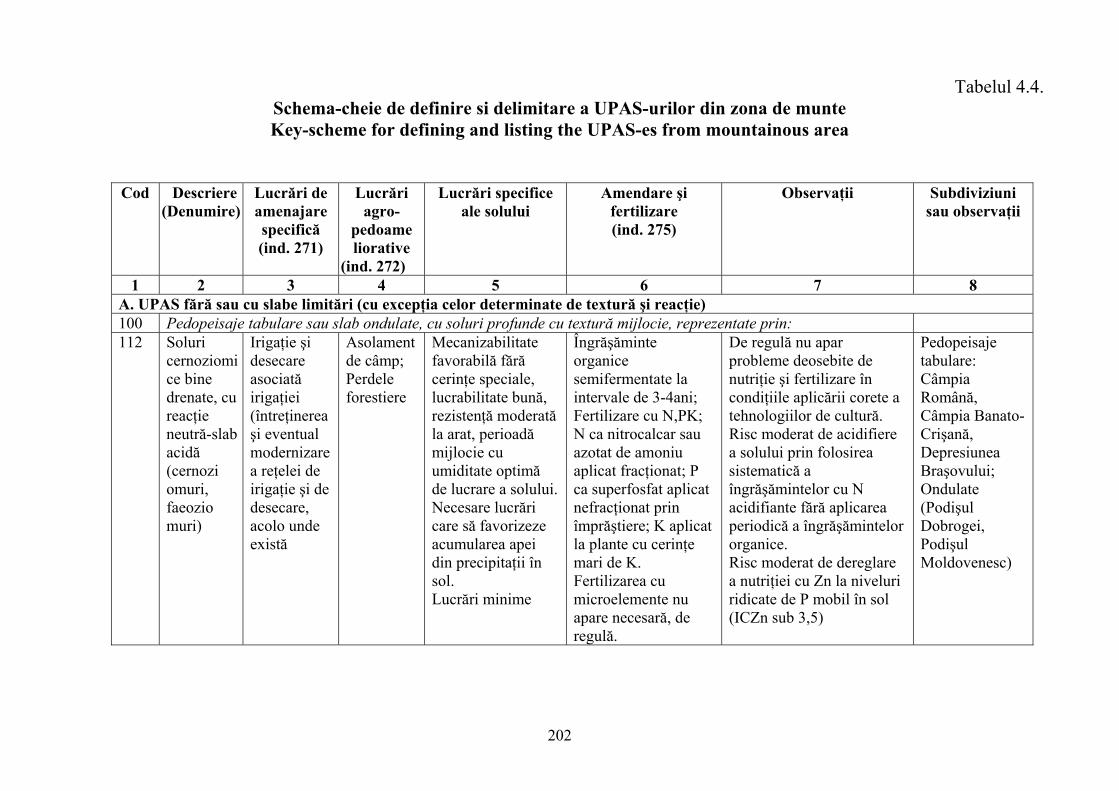
202
Tabelul 4.4. Schema-cheie de definire si delimitare a UPAS-urilor din zona de munte Key-scheme for defining and listing the UPAS-es from mountainous area
Cod Descriere (Denumire)
Lucrări de amenajare specifică (ind. 271)
Lucrări agro-
pedoame liorative
(ind. 272)
Lucrări specifice ale solului
Amendare şi fertilizare (ind. 275)
Observaţii Subdiviziuni sau observaţii
1 2 3 4 5 6 7 8 A. UPAS fără sau cu slabe limitări (cu excepţia celor determinate de textură şi reacţie) 100 Pedopeisaje tabulare sau slab ondulate, cu soluri profunde cu textură mijlocie, reprezentate prin: 112 Soluri
cernoziomice bine drenate, cu reacţie neutră-slab acidă (cernozi omuri, faeozio muri)
Irigaţie şi desecare asociată irigaţiei (întreţinerea şi eventual modernizarea reţelei de irigaţie şi de desecare, acolo unde există
Asolament de câmp; Perdele forestiere
Mecanizabilitate favorabilă fără cerinţe speciale, lucrabilitate bună, rezistenţă moderată la arat, perioadă mijlocie cu umiditate optimă de lucrare a solului. Necesare lucrări care să favorizeze acumularea apei din precipitaţii în sol. Lucrări minime
Îngrăşăminte organice semifermentate la intervale de 3-4ani; Fertilizare cu N,PK; N ca nitrocalcar sau azotat de amoniu aplicat fracţionat; P ca superfosfat aplicat nefracţionat prin împrăştiere; K aplicat la plante cu cerinţe mari de K. Fertilizarea cu microelemente nu apare necesară, de regulă.
De regulă nu apar probleme deosebite de nutriţie şi fertilizare în condiţiile aplicării corete a tehnologiilor de cultură. Risc moderat de acidifiere a solului prin folosirea sistematică a îngrăşămintelor cu N acidifiante fără aplicarea periodică a îngrăşămintelor organice. Risc moderat de dereglare a nutriţiei cu Zn la niveluri ridicate de P mobil în sol (ICZn sub 3,5)
Pedopeisaje tabulare: Câmpia Română, Câmpia Banato-Crişană, Depresiunea Braşovului; Ondulate (Podişul Dobrogei, Podişul Moldovenesc)

203
1 2 3 4 5 6 7 8 113 Soluri
cernoziomice bine drenate, cu reacţie bazică (kastanoziomuri, cernoziomuri calcarice)
Idem Idem Idem Îngrăşăminte organice semifermentate la interval de 3-4 ani. Fertilizare cu N, P, K: N ca azotat de amoniu, aplicat fracţionat; P ca superfosfat aplicat nefracţionat prin împrăştiere; K la plante cu cerinţe mari de K. Fertlizare cu Zn la valori sub 3,5 ale ICZn la plante susceptibile la deficienţe de Zn.
Probabilitate de apariţie a clorozelor determinată de deficienţa de Fe sau Mn la plantele sensibile
Pedopeisaje: Tabulare (Câmpia Română, C. Banato Crişană) Ondulate (Podişul Dobrogei, Pod. Moldovenesc)
122 Soluri preluvice şi cambice, drenate, cu reacţie neutră-slab acidă
Irigaţie de completare în perioade secetoase
Asolament de câmp
Idem Îngrăşăminte organice semifermentate la intervale de 3-4ani; Fertilizare cu N,PK; N ca nitrocalcar sau azotat de amoniu aplicat fracţionat; P ca superfosfat aplicat nefracţionat prin împrăştiere; K aplicat la plante cu cerinţe mari de K.Fertilizarea cu microelemente nu apare necesară, de regulă.
De regulă nu apar probleme deosebite de nutriţie şi fertilizare în condiţiile aplicării corecte a tehnologiilor de cultură. Risc moderat de acidifiere a solului prin folosirea sistematică a îngrăşămintelor cu N acidifiante fără aplicarea periodică a îngrăşămintelor organice. Risc moderat de dereglare a nutriţiei cu Zn la niveluri ridicate de P mobil în sol (ICZn sub 3,5)
Pedopeisaje influenţate de clima: continentală (estică); Balcanică; Central-europeana
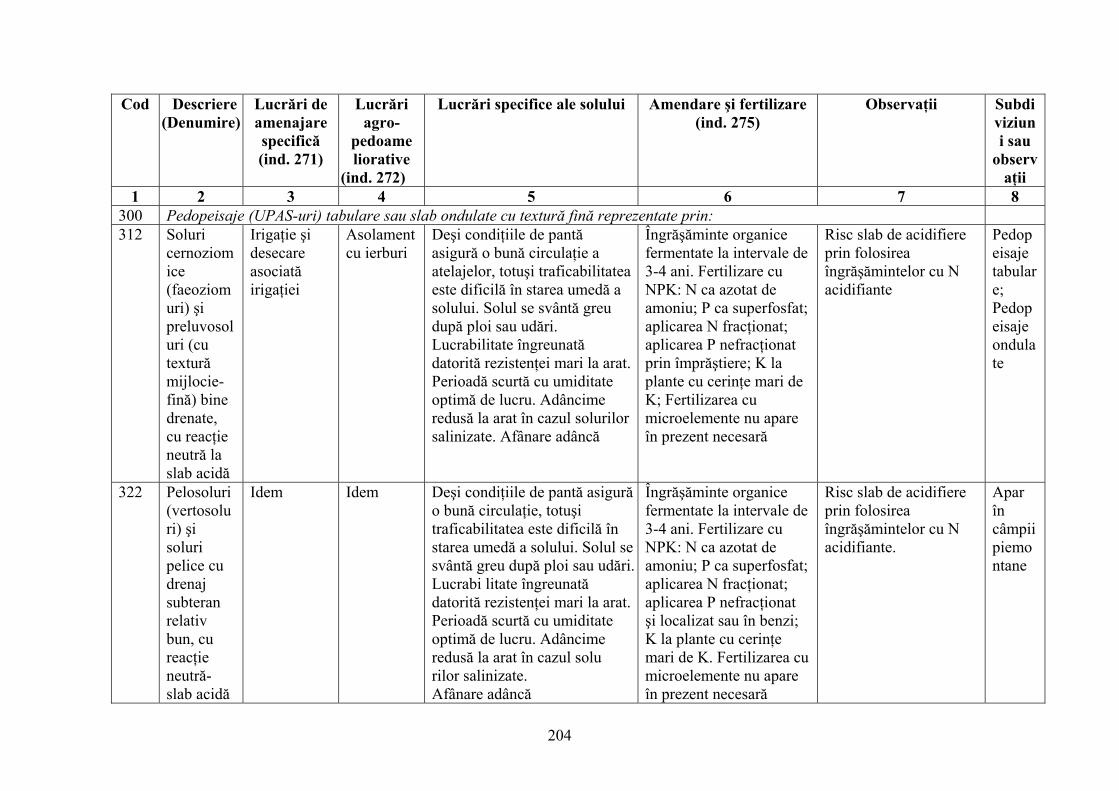
204
Cod Descriere
(Denumire) Lucrări de amenajare specifică (ind. 271)
Lucrări agro-
pedoame liorative
(ind. 272)
Lucrări specifice ale solului Amendare şi fertilizare (ind. 275)
Observaţii Subdiviziuni sau
observaţii
1 2 3 4 5 6 7 8 300 Pedopeisaje (UPAS-uri) tabulare sau slab ondulate cu textură fină reprezentate prin: 312 Soluri
cernoziomice (faeoziomuri) şi preluvosoluri (cu textură mijlocie-fină) bine drenate, cu reacţie neutră la slab acidă
Irigaţie şi desecare asociată irigaţiei
Asolament cu ierburi
Deşi condiţiile de pantă asigură o bună circulaţie a atelajelor, totuşi traficabilitatea este dificilă în starea umedă a solului. Solul se svântă greu după ploi sau udări. Lucrabilitate îngreunată datorită rezistenţei mari la arat. Perioadă scurtă cu umiditate optimă de lucru. Adâncime redusă la arat în cazul solurilor salinizate. Afânare adâncă
Îngrăşăminte organice fermentate la intervale de 3-4 ani. Fertilizare cu NPK: N ca azotat de amoniu; P ca superfosfat; aplicarea N fracţionat; aplicarea P nefracţionat prin împrăştiere; K la plante cu cerinţe mari de K; Fertilizarea cu microelemente nu apare în prezent necesară
Risc slab de acidifiere prin folosirea îngrăşămintelor cu N acidifiante
Pedopeisaje tabulare; Pedopeisaje ondulate
322 Pelosoluri (vertosoluri) şi soluri pelice cu drenaj subteran relativ bun, cu reacţie neutră-slab acidă
Idem Idem Deşi condiţiile de pantă asigură o bună circulaţie, totuşi traficabilitatea este dificilă în starea umedă a solului. Solul se svântă greu după ploi sau udări. Lucrabi litate îngreunată datorită rezistenţei mari la arat. Perioadă scurtă cu umiditate optimă de lucru. Adâncime redusă la arat în cazul solu rilor salinizate. Afânare adâncă
Îngrăşăminte organice fermentate la intervale de 3-4 ani. Fertilizare cu NPK: N ca azotat de amoniu; P ca superfosfat; aplicarea N fracţionat; aplicarea P nefracţionat şi localizat sau în benzi; K la plante cu cerinţe mari de K. Fertilizarea cu microelemente nu apare în prezent necesară
Risc slab de acidifiere prin folosirea îngrăşămintelor cu N acidifiante.
Apar în câmpii piemontane
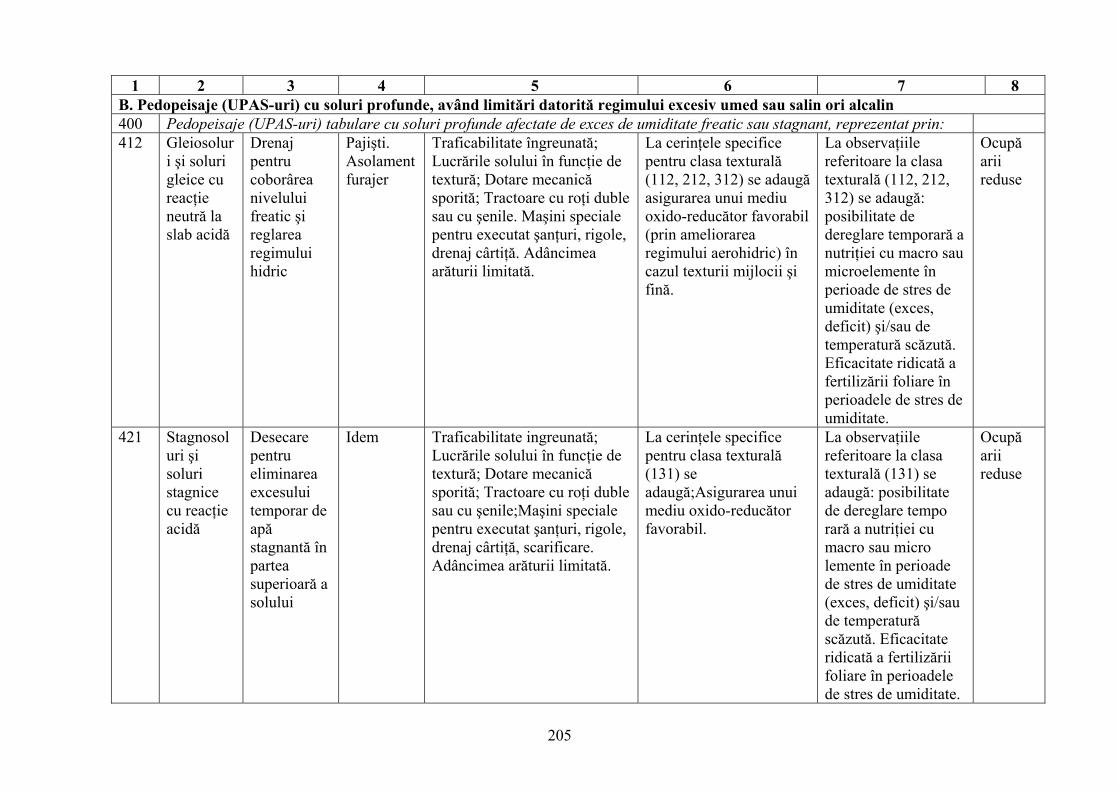
205
1 2 3 4 5 6 7 8 B. Pedopeisaje (UPAS-uri) cu soluri profunde, având limitări datorită regimului excesiv umed sau salin ori alcalin 400 Pedopeisaje (UPAS-uri) tabulare cu soluri profunde afectate de exces de umiditate freatic sau stagnant, reprezentat prin: 412 Gleiosolur
i şi soluri gleice cu reacţie neutră la slab acidă
Drenaj pentru coborârea nivelului freatic şi reglarea regimului hidric
Pajişti. Asolament furajer
Traficabilitate îngreunată; Lucrările solului în funcţie de textură; Dotare mecanică sporită; Tractoare cu roţi duble sau cu şenile. Maşini speciale pentru executat şanţuri, rigole, drenaj cârtiţă. Adâncimea arăturii limitată.
La cerinţele specifice pentru clasa texturală (112, 212, 312) se adaugă asigurarea unui mediu oxido-reducător favorabil (prin ameliorarea regimului aerohidric) în cazul texturii mijlocii şi fină.
La observaţiile referitoare la clasa texturală (112, 212, 312) se adaugă: posibilitate de dereglare temporară a nutriţiei cu macro sau microelemente în perioade de stres de umiditate (exces, deficit) şi/sau de temperatură scăzută. Eficacitate ridicată a fertilizării foliare în perioadele de stres de umiditate.
Ocupă arii reduse
421 Stagnosoluri şi soluri stagnice cu reacţie acidă
Desecare pentru eliminarea excesului temporar de apă stagnantă în partea superioară a solului
Idem Traficabilitate ingreunată; Lucrările solului în funcţie de textură; Dotare mecanică sporită; Tractoare cu roţi duble sau cu şenile;Maşini speciale pentru executat şanţuri, rigole, drenaj cârtiţă, scarificare. Adâncimea arăturii limitată.
La cerinţele specifice pentru clasa texturală (131) se adaugă;Asigurarea unui mediu oxido-reducător favorabil.
La observaţiile referitoare la clasa texturală (131) se adaugă: posibilitate de dereglare tempo rară a nutriţiei cu macro sau micro lemente în perioade de stres de umiditate (exces, deficit) şi/sau de temperatură scăzută. Eficacitate ridicată a fertilizării foliare în perioadele de stres de umiditate.
Ocupă arii reduse
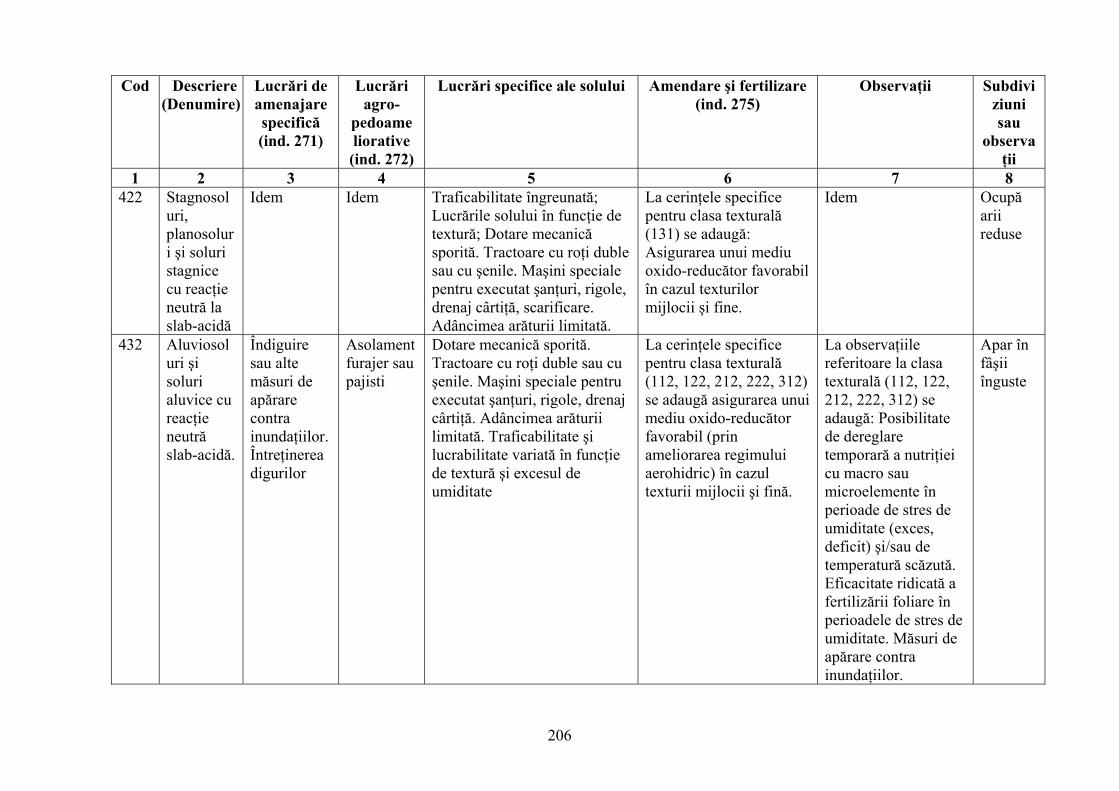
206
Cod Descriere (Denumire)
Lucrări de amenajare specifică (ind. 271)
Lucrări agro-
pedoame liorative (ind. 272)
Lucrări specifice ale solului Amendare şi fertilizare (ind. 275)
Observaţii Subdiviziuni sau
observaţii
1 2 3 4 5 6 7 8 422 Stagnosol
uri, planosoluri şi soluri stagnice cu reacţie neutră la slab-acidă
Idem Idem Traficabilitate îngreunată; Lucrările solului în funcţie de textură; Dotare mecanică sporită. Tractoare cu roţi duble sau cu şenile. Maşini speciale pentru executat şanţuri, rigole, drenaj cârtiţă, scarificare. Adâncimea arăturii limitată.
La cerinţele specifice pentru clasa texturală (131) se adaugă: Asigurarea unui mediu oxido-reducător favorabil în cazul texturilor mijlocii şi fine.
Idem Ocupă arii reduse
432 Aluviosoluri şi soluri aluvice cu reacţie neutră slab-acidă.
Îndiguire sau alte măsuri de apărare contra inundaţiilor. Întreţinerea digurilor
Asolament furajer sau pajisti
Dotare mecanică sporită. Tractoare cu roţi duble sau cu şenile. Maşini speciale pentru executat şanţuri, rigole, drenaj cârtiţă. Adâncimea arăturii limitată. Traficabilitate şi lucrabilitate variată în funcţie de textură şi excesul de umiditate
La cerinţele specifice pentru clasa texturală (112, 122, 212, 222, 312) se adaugă asigurarea unui mediu oxido-reducător favorabil (prin ameliorarea regimului aerohidric) în cazul texturii mijlocii şi fină.
La observaţiile referitoare la clasa texturală (112, 122, 212, 222, 312) se adaugă: Posibilitate de dereglare temporară a nutriţiei cu macro sau microelemente în perioade de stres de umiditate (exces, deficit) şi/sau de temperatură scăzută. Eficacitate ridicată a fertilizării foliare în perioadele de stres de umiditate. Măsuri de apărare contra inundaţiilor.
Apar în fâşii înguste

207
1 2 3 4 5 6 7 8 433 Aluviosol
uri şi soluri aluvice cu reacţie bazică
Îndiguirea sau alte măsuri de apărare contra inundaţiilor. Întreţinerea digurilor. Irigaţie şi desecare. Drenaj în ariile joase
Asolamente de câmp în ariile îndiguite şi irigate; perdele forestiere. Asolament furajer. Pajişti în arii joase.
Idem La cerinţele specifice pentru clasa texturală (113, 213, 223, 313, 323) se adaugă: Asigurarea unui mediu oxido-reducător favorabil în cazul texturilor mijlocii şi fine.
Idem, plus Probabilitate mare de apariţie a clorozelor (Fe, Mn). Măsuri de apărare contra inundaţiilor.
Apar în fâşii înguste în zona deluroasă şi mai largi în regiunea de câmpie.
C. Pedopeisaje (UPAS-uri) cu limitări datorită reliefului şi riscului de eroziune şi alunecare 600 Pedopeisaje (UPAS-uri) cu soluri profunde fără alunecări cu: 610 Soluri
variate (cambice, luvice-) pe versanţi slab moderat înclinaţi
Organizare şi amenajarea teritoriului pentru dezvoltare drabilă
Pădure, pajişte. Pe terenuri cultivate asolament şi structură de culturi adaptate înclinării terenului.
Lucrările solului pe curba de nivel. Adâncimea arăturii limitate pe soluri cu orizont scurt. Scarificarea pe soluri cu orizont B compact. Culturi în benzi (agrotehnică specifică pe terenurile în pantă)
Fertilizarea puţin recomandată. Amendare cu calcar şi dolomit în doze stabilite în funcţie de indicii agrochimici; interval de revenire 5-7 ani. (Fertilizare cu NPK: N ca nitrocalcar; P ca superfosfat; aplicarea N şi K fracţionat; aplicarea P nefracţionat prin împrăştiere). Fertlizarea prin târlire.
Risc de acidifiere rapidă a solului. Probabilitate de dereglare a nutriţiei cu N, P în primăverile reci (datorită diminuării ritmului de absorbţie a P la temperaturi scăzute). Probabilitate de toxicitate de Al la majoritatea plantelor şi de Mn la plante sensibile.
Subdiviziuni ca la 131
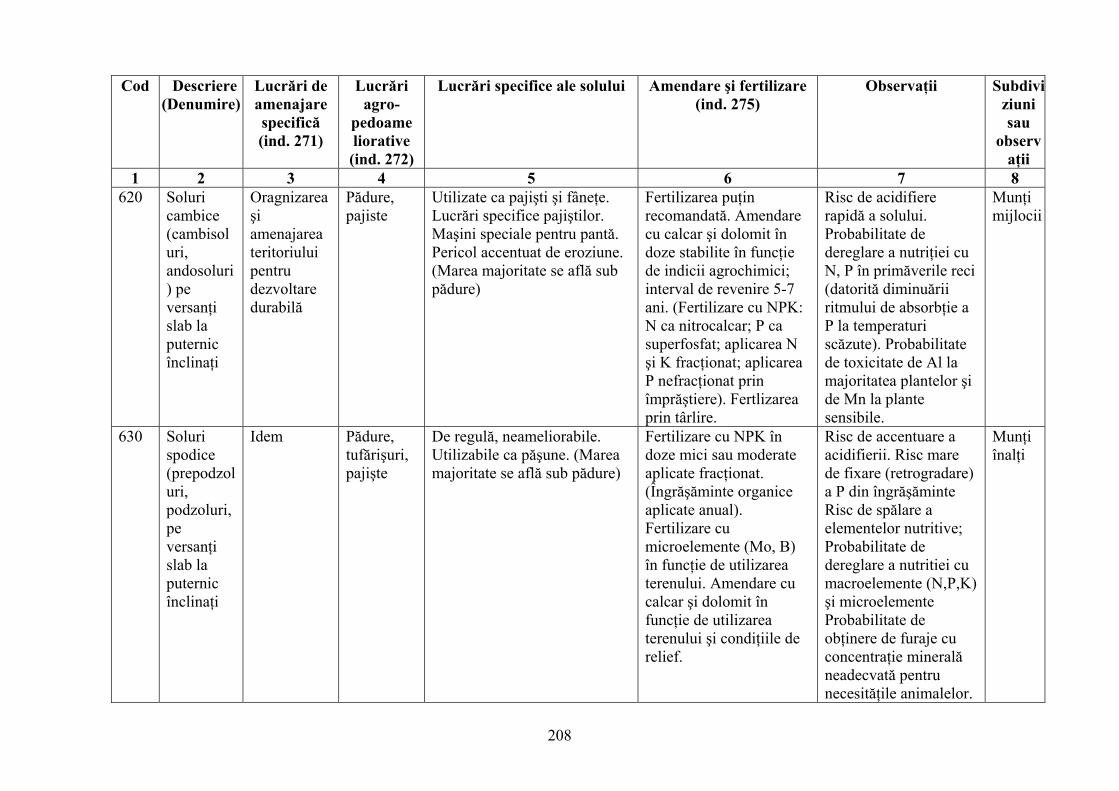
208
Cod Descriere (Denumire)
Lucrări de amenajare specifică (ind. 271)
Lucrări agro-
pedoame liorative (ind. 272)
Lucrări specifice ale solului Amendare şi fertilizare (ind. 275)
Observaţii Subdiviziuni sau
observaţii
1 2 3 4 5 6 7 8 620 Soluri
cambice (cambisoluri, andosoluri) pe versanţi slab la puternic înclinaţi
Oragnizarea şi amenajarea teritoriului pentru dezvoltare durabilă
Pădure, pajiste
Utilizate ca pajişti şi fâneţe. Lucrări specifice pajiştilor. Maşini speciale pentru pantă. Pericol accentuat de eroziune. (Marea majoritate se află sub pădure)
Fertilizarea puţin recomandată. Amendare cu calcar şi dolomit în doze stabilite în funcţie de indicii agrochimici; interval de revenire 5-7 ani. (Fertilizare cu NPK: N ca nitrocalcar; P ca superfosfat; aplicarea N şi K fracţionat; aplicarea P nefracţionat prin împrăştiere). Fertlizarea prin târlire.
Risc de acidifiere rapidă a solului. Probabilitate de dereglare a nutriţiei cu N, P în primăverile reci (datorită diminuării ritmului de absorbţie a P la temperaturi scăzute). Probabilitate de toxicitate de Al la majoritatea plantelor şi de Mn la plante sensibile.
Munţi mijlocii
630 Soluri spodice (prepodzoluri, podzoluri, pe versanţi slab la puternic înclinaţi
Idem Pădure, tufărişuri, pajişte
De regulă, neameliorabile. Utilizabile ca păşune. (Marea majoritate se află sub pădure)
Fertilizare cu NPK în doze mici sau moderate aplicate fracţionat. (Îngrăşăminte organice aplicate anual). Fertilizare cu microelemente (Mo, B) în funcţie de utilizarea terenului. Amendare cu calcar şi dolomit în funcţie de utilizarea terenului şi condiţiile de relief.
Risc de accentuare a acidifierii. Risc mare de fixare (retrogradare) a P din îngrăşăminte Risc de spălare a elementelor nutritive; Probabilitate de dereglare a nutritiei cu macroelemente (N,P,K) şi microelemente Probabilitate de obţinere de furaje cu concentraţie minerală neadecvată pentru necesităţile animalelor.
Munţi înalţi
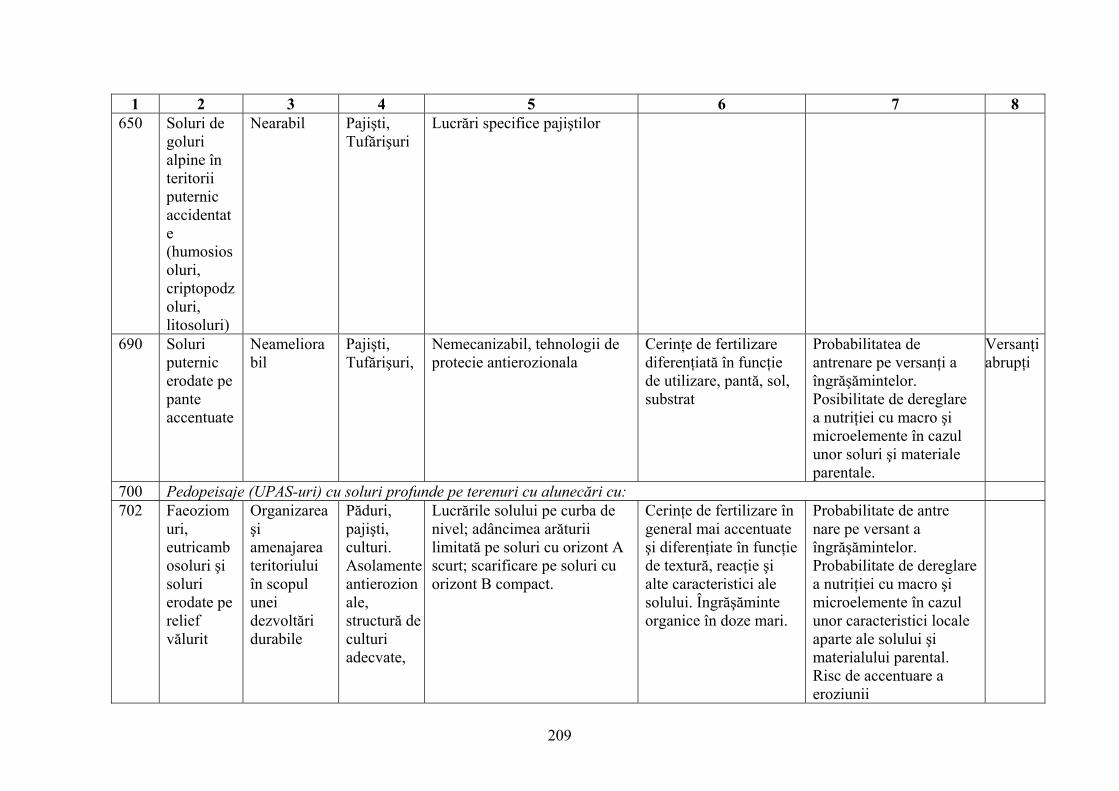
209
1 2 3 4 5 6 7 8
650 Soluri de goluri alpine în teritorii puternic accidentate (humosiosoluri, criptopodzoluri, litosoluri)
Nearabil Pajişti, Tufărişuri
Lucrări specifice pajiştilor
690 Soluri puternic erodate pe pante accentuate
Neameliorabil
Pajişti, Tufărişuri,
Nemecanizabil, tehnologii de protecie antierozionala
Cerinţe de fertilizare diferenţiată în funcţie de utilizare, pantă, sol, substrat
Probabilitatea de antrenare pe versanţi a îngrăşămintelor. Posibilitate de dereglare a nutriţiei cu macro şi microelemente în cazul unor soluri şi materiale parentale.
Versanţi abrupţi
700 Pedopeisaje (UPAS-uri) cu soluri profunde pe terenuri cu alunecări cu: 702 Faeoziom
uri, eutricambosoluri şi soluri erodate pe relief vălurit
Organizarea şi amenajarea teritoriului în scopul unei dezvoltări durabile
Păduri, pajişti, culturi. Asolamente antierozionale, structură de culturi adecvate,
Lucrările solului pe curba de nivel; adâncimea arăturii limitată pe soluri cu orizont A scurt; scarificare pe soluri cu orizont B compact.
Cerinţe de fertilizare în general mai accentuate şi diferenţiate în funcţie de textură, reacţie şi alte caracteristici ale solului. Îngrăşăminte organice în doze mari.
Probabilitate de antre nare pe versant a îngrăşămintelor. Probabilitate de dereglare a nutriţiei cu macro şi microelemente în cazul unor caracteristici locale aparte ale solului şi materialului parental. Risc de accentuare a eroziunii
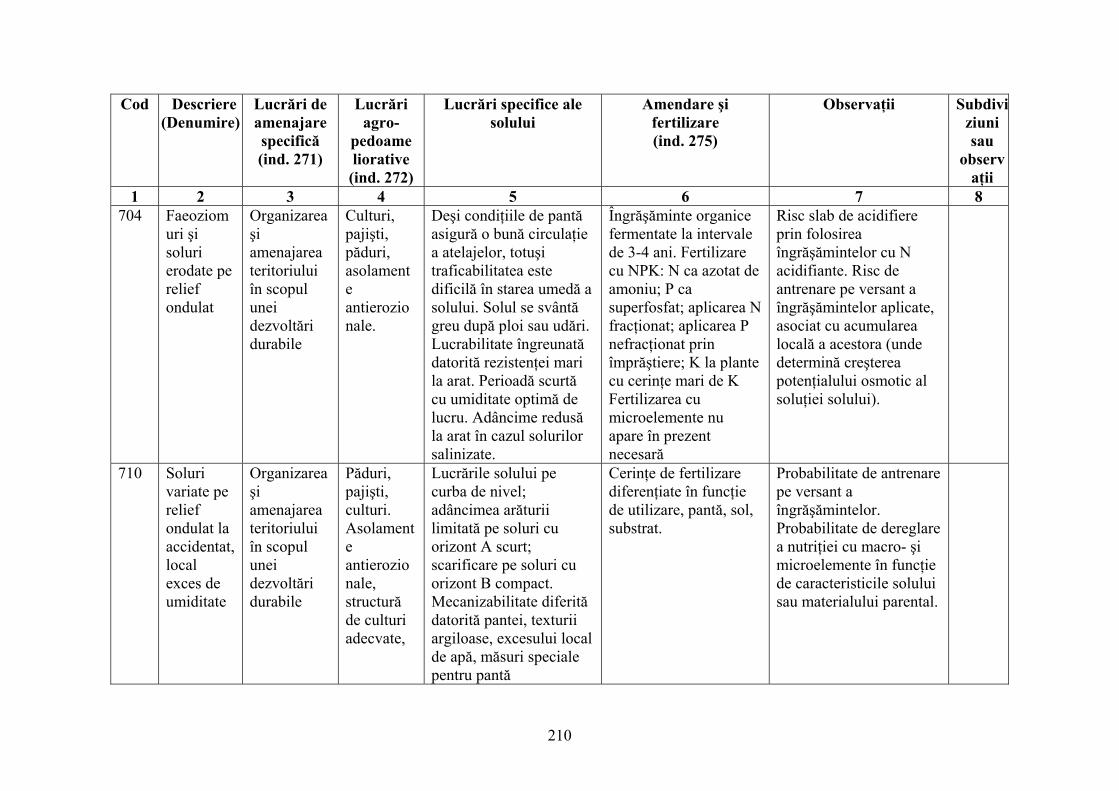
210
Cod Descriere
(Denumire) Lucrări de amenajare specifică (ind. 271)
Lucrări agro-
pedoame liorative (ind. 272)
Lucrări specifice ale solului
Amendare şi fertilizare (ind. 275)
Observaţii Subdiviziuni sau
observaţii
1 2 3 4 5 6 7 8 704 Faeoziom
uri şi soluri erodate pe relief ondulat
Organizarea şi amenajarea teritoriului în scopul unei dezvoltări durabile
Culturi, pajişti, păduri, asolamente antierozionale.
Deşi condiţiile de pantă asigură o bună circulaţie a atelajelor, totuşi traficabilitatea este dificilă în starea umedă a solului. Solul se svântă greu după ploi sau udări. Lucrabilitate îngreunată datorită rezistenţei mari la arat. Perioadă scurtă cu umiditate optimă de lucru. Adâncime redusă la arat în cazul solurilor salinizate.
Îngrăşăminte organice fermentate la intervale de 3-4 ani. Fertilizare cu NPK: N ca azotat de amoniu; P ca superfosfat; aplicarea N fracţionat; aplicarea P nefracţionat prin împrăştiere; K la plante cu cerinţe mari de K Fertilizarea cu microelemente nu apare în prezent necesară
Risc slab de acidifiere prin folosirea îngrăşămintelor cu N acidifiante. Risc de antrenare pe versant a îngrăşămintelor aplicate, asociat cu acumularea locală a acestora (unde determină creşterea potenţialului osmotic al soluţiei solului).
710 Soluri variate pe relief ondulat la accidentat, local exces de umiditate
Organizarea şi amenajarea teritoriului în scopul unei dezvoltări durabile
Păduri, pajişti, culturi. Asolamente antierozionale, structură de culturi adecvate,
Lucrările solului pe curba de nivel; adâncimea arăturii limitată pe soluri cu orizont A scurt; scarificare pe soluri cu orizont B compact. Mecanizabilitate diferită datorită pantei, texturii argiloase, excesului local de apă, măsuri speciale pentru pantă
Cerinţe de fertilizare diferenţiate în funcţie de utilizare, pantă, sol, substrat.
Probabilitate de antrenare pe versant a îngrăşămintelor. Probabilitate de dereglare a nutriţiei cu macro- şi microelemente în funcţie de caracteristicile solului sau materialului parental.

211
1 2 3 4 5 6 7 8 D. Pedopeisaje (UPAS-uri) cu alte limitări 800 Pedopeisaje (UPAS-uri) cu soluri scheletice şi /sau puţin profunde sau extrem de nisipoase sistematizate în pedopeisaje cu: 811 Soluri
scheletice pe profil
Asolament specific (furajer, sau pajişti ori păduri
Mecanizabilitate, traficabilitate şi lucrabilitate îngreunate de caracterul scheletic al solului precum şi de pante
Fertilizare diferenţiată, în raport cu natura scheletului şi volumul edafic, cu aplicare de îngrăşăminte (şi amendamente, dacă este cazul) în doze mici sau moderate, aplicate fracţionat. Fertilizare organică cu doze mari, în special pe soluri formate pe substrate sărace (silicioase).
Risc de spălare în adâncime şi/sau antrenare pe versant a îngrăşămintelor şi amendamentelor.
812 Soluri cu profil scurt (litosoluri, rendzine, soluri litice )
Asolament specific (furajer, sau pajişti ori păduri
Mecanizabilitate, traficabilitate şi lucrabilitate îngreunate de caracterul scheletice al solului precum şi de pante
Fertilizare diferenţiată, în raport cu natura scheletului şi volumul edafic, cu aplicare de îngrăşăminte (şi amendamente, dacă este cazul) în doze mici sau moderate, aplicate fracţionat. Fertilizare organică cu doze mari, în special pe soluri formate pe substrate sărace (silicioase).
Risc de spălare în adâncime şi/sau antrenare pe versant a îngrăşămintelor şi amendamentelor.

212
Cod Descriere (Denumire)
Lucrări de amenajare specifică (ind. 271)
Lucrări agro-
pedoame liorative (ind. 272)
Lucrări specifice ale solului Amendare şi fertilizare (ind. 275)
Observaţii Subdiviziuni sau
observaţii
900 Pedopeisaje (UPAS-uri) cu soluri organice şi anume: 910 Histosoluri
districe (acide)
Se recomandă folosirea în condiţii naturale. Luarea în cultură presupune ameliorare
Pajişte Mecanizabilitate dificilă Desecare prealabilă. Amendare cu calcar şi dolomit în doze mici (0,3-0,5 t/ha la pH=6) la intervale scurte (1-2 ani). Cerinţe ridicate de P şi K. Fertilizare cu NPK: N ca nitrocalcar; P ca superfosfat, fosfaţi bruţi sau fosforite activate; aplicare fracţionată. Fertilizare organică la 4-5 ani. Fertilizare cu Cu la plante sensibile la deficienţa cu Cu sau la plante furajere. Amestec cu material pământos.
Risc de suprafertilizare la aplicarea de doze mari nefracţionate de îngrăşăminte. Risc de spălare a îngrăşămintelor, cu probabilitatea de poluare a mediului. Risc mărit de scădere a rezistenţei plantelor în primele perioade de creştere (datorită temperaturilor scăzute) în condiţiile unei fertilizări neechilibrate. Probabilitate de dereglare a nutriţiei cu Ca şi Mg. Probabilitate de obţinere de furaje cu compoziţie minerală neadecvată pentru necesităţile animalelor. Risc de autoaprindere şi de spulberare eoliană.

213
Din datele de până acum, se poate observa că UPAS-urilor cu soluri profunde
cu limitări datorită reliefului, fără alunecări (cambisoluri, andosoluri) pe versanţi de la slab la puternic înclinaţi (620) ocupa cea mai mare suprafaţă. În cadrul acestor clase, domină brunele acide (BO) si andosolurile (AN).
Solurile brune acide analizate prezintă textura mijlocie-grosieră până la mijlocie, nediferenţiată pe profil, drenaj intern bun şi în consecintă cu regim aerohidric favorabil. Reacţia este acidă, pH <5, în orizontul de suprafată, conţine cantităţi importante de materie organică (humus de tip mull acid cu mineralizare slabă), frecvent peste 4%, conţinut de azot total de la mijlociu la foarte mare, conţinut de P mobil mic şi foarte mic, K mobil in cantitati mici şi foarte mici. Gradul de saturaţie în baze este <50%, frecvent <35%.
Climatul rece şi umed în care se întâlnesc aceste soluri şi reacţia acidă a solului inhibă aprovizionare optimă cu elemente nutritive a plantelor.
Având în vedere aciditatea naturală a acestor soluri, este absolut necesară neutralizarea acidităţii efective la fertilizare, în special la cea cu azotaţi.
Este necesară şi eficientă, ţinând cont de condiţiile bioclimatice, amendarea moderată şi fertilizarea moderată până la intensă cu NPK.
Andosolurile (AN) ocupă suprafeţe mai mici în cadrul acestei grupe. Din profilele analizate se poate observa ca sunt soluri nediferenţiate textural pe profil, conţin cantităţi mari de materie organică, dar slab transformată şi acidă, pH 5,7, frecvent sub 5, grad de saturaţie în baze sub 50%. Sunt puţin active microbiologic şi slab aprovizionate cu substanţe nutritive. Pentru îmbunătăţirea calităţii pajiştilor de pe aceste soluri sunt necesare îngrăşăminte cu NPK şi amendamente calcaroase.
Pe locul doi ca întindere (16.55%) se află UPAS-urile cu soluri variate pe relief ondulat la accidentat (710), local cu exces de umiditate. In cadrul acestei categorii, solurile brune eumezobazice (BM) ocupă suprafeţele cele mai mari. Din profilele analizate, se poate observa ca acestea sunt soluri echilibrate din punct de vedere al însuşirilor fizico-chimice: textură de la mijlociu-grosieră până la fină (de obicei, mijlocie), nediferenţiate pe profil, bine aprovizionate cu humus, pH frecvent peste 6, grad de saturaţie în baze >53%, aprovizionare cu substanţe nutritive şi activitate microbiologică relativ bună.
Potenţialul lor productiv poate fi mai eficient valorificat prin fertilizare slabă până la moderată cu N şi P, însoţită unde este cazul (existenţa Als) în soluri mezobazice, de amendare foarte slabă la solurile cu capacitate de tamponare mică (Te mic), evoluate pe substrate slab tamponate.
Solurile spodice (prepodzoluri, podzoluri, pe versanţi slab la puternic înclinaţi-UPAS- 630) ocupă, de asemenea, suprafeţe importante (14, 06%). Solurile spodice (prepodzoluri şi podzoluri PB, PD) sunt soluri cu conţinut mare şi foarte mare de humus de tip mull, foarte puternic acid, mull-moder, moder şi chiar humus brut, cu grad slab de mineralizare, conţinut mic şi foarte mic de argilă nesaturată, conţinut mare de materiale amorfe, crescând cu stadiul de podzolire. În consecinţă, sunt soluri foarte puternic şi puternic acide cu cantităţi mari şi extrem de mari de Als, predominant oligobazice (V<35%) cu deficit mare şi chiar carenţă de elemente nutritive.

214
Ameliorarea acestor soluri trebuie să aibă în vedere amendarea şi fertilizarea moderată cu NPK. Fertilizarea pe aceste soluri trebuie să ţină cont de competitivitatea în absorbţia elementelor nutritive a cantităţilor mari, cu aport continuu, de materiale amorfe.
O altă unitate importantă este cea a solurilor variate (cambice, luvice) pe versanţi slab moderat înclinaţi – UPAS – 610 (8,48%). Dominante sunt în această unitate solurile luvice (BD, BP, SP), care sunt soluri diferenţiate textural, cu un conţinut ridicat de argilă pe profil, conţinuturi mici de humus saturat sau slab acid, deficit moderat până la mare de elemente nutritive. Gradul de debazificare şi acumulare de aluminiu schimbabil în orizontul eluvial determină reacţie predominant puternic acidă, defavorizează circuitul de substanţe nutritive şi mai ales absorbţia acestora de către plante, mai ales în solurile cu capacitate de tamponare mai scăzută, cu conţinut mic de humus şi evoluate pe substrate mai acide.
Argilozitatea mare şi diferenţierea texturală accentuată afectează drenajul intern mai ales pe suprafeţele plane sau slab înclinate, cu diminuarea conductivităţii hidraulice la valori sub 2 mm/h şi chiar sub 1 mm/h şi scăderi ale potenţialului de oxido-reducere la valori caracteristice pentru regim aerohidric nefavorabil sau puţin favorabil.
Factorii limitativi pentru aceste soluri sunt aciditatea, deficitul de elemente nutritive, drenajul global imperfect. Compensarea deficitului de elemente nutritive trebuie să ţină cont de nivelul adsorbţiei fosfatice în funcţie de argilozitatea solului, de natura mineralogică a materialului parental şi proporţia de potasiu în complexul adsorbtiv, de efectul acidifiant al azotaţilor.
Sunt necesare şi eficiente, pentru aceste soluri, amendarea moderată, fertilizarea moderată până la intensă cu fosfor, moderată cu potasiu şi azot. Solurile luvice cu drenaj subteran bun, cu reacţie acidă – UPAS - 131, ocupă suprafete mai mici, 2,91%. Avand drenaj subteran bun la aceste soluri dispar limitările legate de excesul de umiditate, creşte accesibilitatea nutrienţilor pentru plante. Sunt soluri sărace în materie organică şi elemente nutritive. Raspund cu rezultate foarte bune la aplicarea îngrăşăminetelor organice şi minerale. Amendamentele calcaroase au, de asemenea, efecte benefice asupra stării de nutriţie a acestor soluri.
Solurile cu profil scurt (litosoluri, rendzine, soluri litice) – UPAS - 812, ocupă suprafeţe mici (2,75%). Rendzinele sunt bogate în humus de bună calitate, peste 10%,sunt saturate sau slab debazificate V%=100-70, pH 7,5-6, bine aprovizionate cu elemente nutritive si cu activitate microbiologică intensă. Sunt soluri fertile care răspund foarte bine la aplicarea gunoiului de grajd si a îngrăşămintelor minerale. Necesită îndepărtarea pietrelor, atunci când acestea există, precum şi lucrări de combaterea eroziunii pe suprafeţele înclinate.
Litosolurile sunt ocupate de o vegetaţie slab reprezentată. Utilizarea lor în scopuri agricole necesită aplicarea de îngrăşăminte organice şi minerale, îndepărtarea pietrelor.
Restul UPAS-urilor ocupă suprafeţe foarte mici, împreună însumând 4,7% din zona cercetată.

215
*** Problemele principale ale agriculturii din regiunea de munte privesc îndeosebi
ridicarea stării de fertilitate a solurilor ocupate de pajişti, în scopul ameliorării compoziţiei floristice şi a creşterii producţiei păşunilor şi fâneţelor.
Amendamentele calcaroase şi îngrăşămintele sunt necesare pe toate solurile montane ocupate de pajişti. Aplicarea unor doze convenabile (5-10 t/ha) de amendament calcaros pe pajiştile cu Nardus stricta, ameliorează compoziţia floristică.
Dintre îngrăşămintele minerale, cele mai bune rezultate le dau îngrăşămintele cu azot. Sporuri mari de recoltă se obţin însă prin folosirea de îngrăşăminte complexe (NPK), aplicate împreună cu amendamentul calcaros.
Datorită regimului hidric dominant sau predominant percolativ al solurilor din regiunea de munte, este economică folosirea de doze relativ mici atât de amendament (aplicate toamna), cât şi de îngrăşăminte (aplicate primăvara după topirea zăpezii). Îngrăşămintele cu reacţie fiziologic acidă sunt contraindicate fără amendamente calcaroase. Astfel de îngrăşăminte, deşi determină o creştere apreciabilă a producţiei pajiştilor, au ca efect accentuarea acidităţii şi debazificării solurilor şi o înrăutăţire în perspectivă a fertilităţii.
Gunoiul de grajd aplicat în doze mari are un efect favorabil de durată atât în ceea ce priveşte creşterea producţiei de iarbă, cât şi asupra calităţii nutritive a acesteia. Un procedeu eficace pentru creşterea productivităţii pajiştilor este îngrăşarea prin târlire. Datele din literatura de specialitate evidenţiază înbunătăţirea considerabilă a condiţiilor de reacţie şi saturaţie în baze a solurilor îngrăşate prin târlire. Producţia de masă verde creşte de 3-4 ori şi se ameliorează substanţial valoarea ei nutritivă prin creşterea conţinutului de proteină digestibilă.
Efectul târlirii se menţine mulţi ani, iar când se conbină târlirea cu aplicarea de amendamente calcaroase, rezultatele sunt şi mai bune.

151

216
V. PRINCIPALELE TIPURI DE HABITATE DE PAJIŞTI DIN ZONA MONTANĂ
5.1. ARGUMENT PENTRU CONSERVAREA BIODIVERSITĂŢII
Biodiversitatea (bios gr.= viaţă, diversitas lat. = diversitate) reprezintă
totalitatea organismelor vii (plante, animale, microorganisme), cu alte cuvinte biodiversitatea se referă la întreaga lume vie în ansamblul ei. În funcţie de nivelurile de organizare a materiei vii, se pot distinge următoarele tipuri de biodiversitate (CRISTEA et DENAEYER 2004):
Biodiversitate genetică (intraspecifică) care reprezintă variabilitatea genotipurilor şi genofondului din cadrul populaţiilor unei specii;
Biodiversitate specifică (interspecifică) care cuprinde totalitatea speciilor existente într-un anumit biotop, regiune, ţară, continent etc.;
Biodiversitate ecologică (a ecosistemelor) care reprezintă mozaicarea diverselor comunităţi de organisme, dar şi complexul de relaţii funcţionale din cadrul ecosistemelor;
Biodiversitate culturală care cuprinde totalitatea practicilor, tradiţiilor şi creaţilor umane prin care omul a reuşit atât să creeze biodiversitate (specii hibride, soiuri, rase etc.) cât şi să conserve anumite tipuri de ecosisteme.
Începând cu ultimele decenii ale secolului trecut, odată cu conştientizarea faptului că un număr tot mai mare specii de animale şi plante dispar iar rata de dispariţie a acestora este tot mai mare, s-a pus tot mai mult accent pe găsirea unor măsuri care să permită conservarea acestora. Odată cu creşterea ratei de extincţie a speciilor, pierdem nu doar specii cu o anumită valoare economică (alimentară, medicinală, tinctorială etc.), dar şi un număr mare de specii despre a căror importanţă nu ştim nimic, ceea ce nu înseamnă că ele nu au nici o valoare.
Din punct de vedere al valorii biodiversităţii, se pot distinge 4 categorii de valoare (CRISTEA et DENAEYER 2004):
- valoarea de folosinţă, organismele vii putând fi folosite ca materii prime în diverse procese de producţie artizanală sau industrială (valoare de producţie), pentru activităţi comerciale (valoare de piaţă), pentru alimentaţie prin consumul lor în stare proaspătă sau conservată (valoare de consum). În multe cazuri, biodiversitatea oferă anumite particularităţi ale peisajului care îi conferă o valoarea recreativă, sau reprezintă obiectul unor studii, cercetări, acţiuni de instruire şi educaţie a populaţiei, fapt ce îi conferă o valoare cognitivă.
- valoarea ecologică sau valoare de protecţie şi reglaj, deşi nu se poate cuantifica întotdeauna în bani, constituie un factor important care afectează în mod direct sau indirect întreaga activitate a omului.
- valoarea de opţiune viitoare sau valoarea de moştenire care reprezintă obligaţia noastră de a lăsa generaţiilor viitoare o anumită zestre pe care ei să o poată valorifica în funcţie de propriile necesităţi.

217
- valoarea intrinsecă sau valoarea de existenţă care reprezintă dreptul fiecărei fiinţe vii la perpetuare în timp, la evoluţie şi diversificare prin procese naturale. Conştientizarea importanţei conservării biodiversităţii, deşi cu un rol decisiv pentru motivarea acţiunilor de conservare, nu reprezintă decât jumătate din problemă, cealaltă jumătate fiind reprezentată de găsirea modalităţilor practice prin care se poate realiza acest deziderat. În prezent, principalele două mari sisteme de conservare sunt conservarea „in situ” şi „ex situ”, fiecare dintre ele având particularităle lui. Conservarea „in situ” reprezintă soluţia optimă, ideală pentru conservarea biodiversităţii, deoarece „... singura posibilitate reală de a proteja speciile periclitate cu dispariţia rămâne astfel încercarea de a conserva comunităţile biologice şi ecosistemele din care fac parte” (BOŞCAIU 1985, p. 127 ap. CRISTEA et DENAEYER 2004). Pentru realizarea conservării „in situ” este nevoie să se identifice anumite arii (situri) cu importanţă mare pentru conservarea biodiversităţii, care vor intra în anumite categorii de protecţie (rezervaţii ştiinţifice, rezervaţii ale Biosferei, parcuri naţionale, naturale, situri Natura 2000 etc.) în cadrul cărora se va reglementa intensitatea acţiunii antropice, de la interzicerea totală a oricărei activităţi umane (rezervaţii ştiinţifice), restricţionarea acestora doar la activităţi tradiţionale (situri Natura 2000) până la permisiunea de a desfăşura o gamă mai variată de activităţi economice, dar doar cu aprobarea forurilor competente şi pe baza unor studii de impact. Conservarea „ex situ” reprezintă „... un complex de variate măsuri complementare, dezvoltate într-o altfel de ambianţă ecologică decât cea caracteristică unităţii ţintă, în scopul refacerii şi perpetuării populaţiilor unor specii ameninţate sau a unor soiuri şi rase” (CRISTEA et DENAEYER 2004, p. 69). Fără a intra în detalii privind particularităţile fiecărei metode, în cadrul acestui sistem se pot distinge colecţiile tematice, băncile de gene, conservatoarele in situ. Fără a avea pretenţia de a fi epuizat acest vast subiect, sperăm că am reuşit, într-o oarecare măsură, să subliniem atât importanţa conservării biodiversităţii, cât şi să oferim câteva informaţii, foarte sumare, despre principalele sisteme de conservare.
5.2. CONSIDERAŢII METODOLOGICE Pentru delimitarea arealelor montane care adăpostesc principalele habitate de pajişti cu valoare conservativă mare au fost luate ca puncte de plecare atât observaţiile efectuate pe teren în cadrul proiectului (în măsura în care aceste observaţii au putut fi efectuate ca urmare a reducerii drastice a bugetului proiectului) cât şi a datelor publicate în literatura de specialitate privitor la vegetaţia României (COLDEA 1972, 1984, 1990, 1991; COLDEA et al. 1997; SANDA,POPESCU et BARABAŞ 1998; SANDA,POPESCU et STANCU 2001; POP, CRISTEA et HODIŞAN 2002; SANDA,ÖLLERER et BURESCU 2008; SANDA,VICOL et ŞTEFĂNUŢ 2008).
Pentru evidenţierea distribuţiei pajiştilor cu valoare conservativă mare, datorită lipsei hărţilor de vegetaţie detaliate, s-a optat pentru delimitarea arealelor la nivel de masive muntoase.

218
Pentru habitatele de pajişti cu o răspândire largă în toată zona montană, nu s-au mai realizat hărţi de distribuţie.
Delimitarea masivelor muntoase s-a realizat pe baza hărţii unităţilor de relief din România (CANDREA, CANDREA et NIŢĂ 2008) setul de date vectoriale fiind descărcat de pe situl www.earth.unibuc.ro. Sistemul de proiecţie a datelor este Stereo 70, iar hărţile de distribuţie s-au realizat folosind ArcInfo versiunea 9 (ESRI 2004). Frontiera de stat a României, principalele cursuri de apă şi oraşe, au fost vectorizate având ca bază cartografică harta geologică a României la scara 1:200000.
Pentru clasificarea şi descrierea habitatelor de pajişti s-a optat pentru sistemul de clasificare propus de DONIŢĂ et al. (2005), considerând că acest sistem de clasificare este mult mai accesibil publicului larg, prezentându-se şi corespondenţa cu alte sisteme de clasificare.
Deoarece aceste habitate (în mare parte) necesită măsuri sporite de protecţie, pentru evidenţierea gradului în care acestea au fost incluse într-o arie naturală protejată, s-a comparat distribuţia acestora cu distribuţia siturilor Natura 2000 la nivelul regiunii biogeografice alpine. S-a optat pentru evaluarea gradului în care arealele habitatelor analizate incluse în cadrul siturilor Natura 2000 deoarece, ariile protejate naturale din România se suprapun peste siturile de importanţă comunitară. Limitele siturilor Natura 2000 sunt cele stabilite prin OM 1964/2007, pentru realizarea hărţilor fiind utilizate datele vectoriale existente pe pagina Ministerului Mediului (http://www.mmediu.ro/ protectia_naturii/protectia_naturii.htm).
Pentru stabilirea valorii conservative a fiecărui habitate s-a luat în calcul atât răspândirea fiecărui habitat la nivel european şi naţional, habitatele endemice au fost considerate ca fiind cele mai valoroase din punct de vedere conservativ, dar şi prezenţa/absenţa în structura habitatelor a speciilor care se regăsesc pe Listele Roşii naţionale. Pentru identificarea speciilor rare s-a consultat Lista Roşie elaborată de OLTEAN şi colaboratorii (1994).
5.3. DESCRIEREA PRINCIPALELOR TIPURI DE HABITATE DE PAJIŞTI DIN ZONA MONTANĂ A ROMÂNIEI
5.3.1. Pajişti alpine şi subalpine 5.3.1.1. Habitatul R3601 - Pajişti sud-est carpatice de rogoz pitic (Kobresia myosuroides) şi Oxytropis carpatica NATURA 2000: 6170 Alpine and subalpine calcareous grasslands EMERALD: - CORINE: 36.42 Wind edge naked-rush swards PAL.HAB 1999: 36.425 Carpathian naked-rush swards EUNIS: E4.42 Wind edge (Kobresia myosuroides) swards

219
Asociaţii vegetale: Oxytropido carpaticae - Elynetum (Puşcaru et al. 1956) Coldea 1991(Syn. Elynetum myosuroides Puşcaru et al. 1956; inclusiv Oxytropido carpaticae - Elynetum festucetosum bucegiensis Coldea 1991); Oxytropido carpaticae - onobrychidetum transsilvanicae Täuber 1987. Scurtă descriere: Fitocenozele caracteristice acestui habitat se întâlnesc pe platouri şi creste cu pante moderate din etajul alpin ocupând staţiuni cu diferite expoziţii şi puternic vântuite pe tot parcusrul anului. Acest habitat prezintă cel mai evoluat stadiu de înţelenire din etajul alpin, şi este considerat ca fiind un relict glaciar. Habitatul are o distribuţie insulară în etajul alpin. Condiţii staţionale: Altitudine: 1850-2500 m. Clima: T = 1,0- -2,50C; P = 1300-1450 mm. Relief: platouri, creste şi muchii, acolo unde zăpada este spulberată de vânt din locuri puternic vântuite în timpul iernii. Substrat: calcare, gresii, conglomerate. Soluri: crude sau brune de înţelenire primară, cu pH = 4,8-5,3. Profilul de sol prezintă orizonturi puţin diferenţiate. Specii dominante şi caracteristice: Kobresia myosuroides (Elyna myosu- roides), Oxytropis carpatica, Kobresia myosuroides, Achillea schurii, Minuartia sedoides, Cerastium alpinum ssp. lanatum, Erigeron uniflorus, Dianthus glacialis, Carex atrata, Oxytropis halleri, Minuartia gerardii, Potentilla crantzii, Carex rupestris, Astragalus alpinus, Minuartia verna, Gentiana nivalis, Helianthemum alpestre, Minuartia verna, Leontopodium alpinum, Pedicularis verticillata, Euphrasia salisburgensis, Anthyllis alpestris, Poa alpina, Agrostis alpina, Achillea schurii, Festuca bucegiensis, Onobrychis transsilvanica. Valoare conservativă: are o valoare conservativă mare atât prin faptul că este un habitat endemic cât şi prin prezenţa unor specii rare/endemice/vulnerabile aflate pe listele roşii (Oxytropis carpatica, Onobrichis transsilvanica, Achillea schurii, Dianthus glacialis, Festuca bucegiensis, Pedicularis oederi etc.). Corologie: Carpaţii Meridionali, Munţii Bucegi, Munţii Piatra Craiului, Munţii Făgăraş, Munţii Retezat. Starea actuală de protecţie: - toate masivele muntoase în care acest habitat este răspândit sunt incluse în situri din cadrul Reţelei Natura 2000 (Fig. 5.1).

220
Fig. 5.1. Harta de distribuţie a habitatului R3601 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3601 into Natura 2000 Network 5.3.1.2. Habitatul R3602 - Pajişti sud-est carpatice de coarnă (Carex curvula) şi Primula minima NATURA 2000: 6150 Siliceous alpine and boreal grassland EMERALD: CORINE: 36.341 Carex curvula grassland PAL.HAB. 1999: 36.3413 Carpathian Carex curvula grasslands EUNIS: E4.34 Alpigenous acidophilous grassland Asociaţii vegetale: Primulo-Caricetum curvulae Br.-Bl. 1926 em. Oberd.1957 (Syn. Caricetum curvulae Brockm.-Jer. 1907).

221
Scurtă descriere: Fitocenozele acestui habitat formează pajişti primare în etajul alpin. Habitatul este prezent în aproape toate masivele lanţului carpatic cu vârfuri de peste 2000, dar suprafeţele ocupate sunt destul de reduse. Specia caracteristică şi edificatoare Carex curvula domină masiv, având o acoperire cuprinsă între 65-85%. Primula minima are o prezenţă ridicată, dar o acoperire redusă. Condiţii staţionale: Altitudine: 1980-2500 m. Clima: T = 0,5- -2,50C; P = 1350-1450 mm. Relief: terenuri plane sau slab înclinate Substrat: gresii, conglomerate. Soluri: humosio-soluri puternic acide (pH = 4,2-4,4), cu troficitate redusă şi cu regim chiono-higrofil; profilul solului este redus (10-20 cm). Specii dominante şi caracteristice: Carex curvula, Primula minima, Oreochloa disticha, Potentilla aurea ssp. chrysocraspeda, Loiseleuria procumbens, Pedicularis exaltata, Vaccinium vitis-idaea, Campanula alpina, Phyteuma confusum, Luzula spicata, Armeria alpina, Dianthus glacialis, Sesleria coerulans, Senecio carpaticus, Hieracium alpinum, Pulsatilla alba, Minuartia sedoides, Vaccinium gaultherioides. Valoarea conservativă: este medie, acest habitat adăpostind o serie de specii aflate pe listele roşii (Pedicularis exaltata, Oreochloa disticha, Phyteuma confusum, Dianthus glacialis, Armeria alpina, Vaccinium gaultheroides etc.) Corologie: Munţii Rodnei, Munţii Călimani, Munţii Suhard, Munţii Bucegi, Munţii Piatra Craiului, Munţii Iezer-Păpuşa, Munţii Făgăraş, Munţii Cibinului, Munţii Parâng, Munţii Retezat, Munţii Ţarcu, Munţii Godeanu. Starea actuală de protecţie: - toate masivelor muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.2).

222
Fig. 5.2. Harta de distribuţie a habitatului R3602 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3602 into Natura 2000 Network 5.3.1.3. Habitatul R3603 - Pajişti sud-est carpatice de părul porcului (Juncus trifidus) şi Oreochloa disticha NATURA 2000: 6150 Siliceous alpine and boreal grassland EMERALD: - CORINE: 36.34 Crooked-sedge swards and related communities PAL.HAB 1999: 36.3463 Carpathian Juncus trifidus swards, 36.34632 Eastern Carpathian Juncus trifidus swards EUNIS: E4.3463 Carpathian Juncus trifidus swards

223
Asociaţii vegetale: Oreochloo -Juncetum trifidi Szafer 1927 (Syn.:Juncetum trifidi BUIA 1962; Rhododendro - Juncetum trifidi Resmeriţă 1975; Junceto trifidi -Vaccinietum RESMERIŢĂ 1975). Scurtă descriere: Acest habitat este reprezentat prin pajişti primare, cu caracter xerofil-oligoterm fiind prezent în mai toate masivele muntoase importante din ţară. Speciile caracteristice şi edificatoare Oreochloa disticha şi Juncus trifidus se găsesc, de cele mai multe ori, în raporturi de codominanţă pe suprafeţele cu expoziţie nordică, în timp ce pe platouri domină Juncus trifidus, Oreochloa disticha fiind sporadică. Cele două specii edificatoare realizează o acoperire medie de 50-80%. Condiţii staţionale: Altitudine: 1800-2500 m. Clima: T = 1- -2,50C; P = 1300-1450 mm. Relief: versanţi slab-puternic înclinaţi, cu expoziţii variate, expuse vânturilor. Substrat: gresii, conglomerate. Soluri: humosiosoluri acide (pH = 4,2-4,5) şi foarte sărace în elemente nutritive. Specii dominante şi caracteristice: Oreochloa disticha, Juncus trifidus, Carex curvula, Potentilla aurea ssp. chrysocraspeda, Loiseleuria procumbens, Empetrum nigrum ssp. hermafroditum, Vaccinium vitis-idaea, Vaccinium myrtillus, Campanula alpina, Festuca airoides, Primula minima, Agrostis rupestris, Avenula versicolor, Phyteuma confusum, Luzula spicata, Sesleria coerulans, Senecio carpaticus, Hieracium alpinum, Pulsatilla alba, Minuartia sedoides, Vaccinium gaultherioides. Valoare conservativă: are o valoare conservativă medie, habitatul adăpostind o serie de specii aflate pe listele roşii (Oreochloa disticha, Phyteuma confusum, Vaccinium gaultheroides, Empetrum nigrum ssp. hermafroditum etc.) Corologie: Carpaţii Orientali: Munţii Rodnei (Coasta Netedă, Şaua Gărgălău-Galaţiu, vârfurile Pietrosul Mare, Momaia, Golgota, Buhăiescu, Anieşul Mare, Omul, Galaţiul, Ineu), Munţii Giumalău, Munţii Călimani. Carpaţii Meridionali: Munţii Făgăraş (vârfurile Zârna-Negoiu), Munţii Cibinului, Valea Sadului, Munţii Parâng, Munţii Retezat. Starea actuală de protecţie: - toate masivele muntoase în care acest habitat este răspândit sunt incluse (parţial sau total) în situri din cadrul Reţelei Natura 2000 (Fig. 5.3).
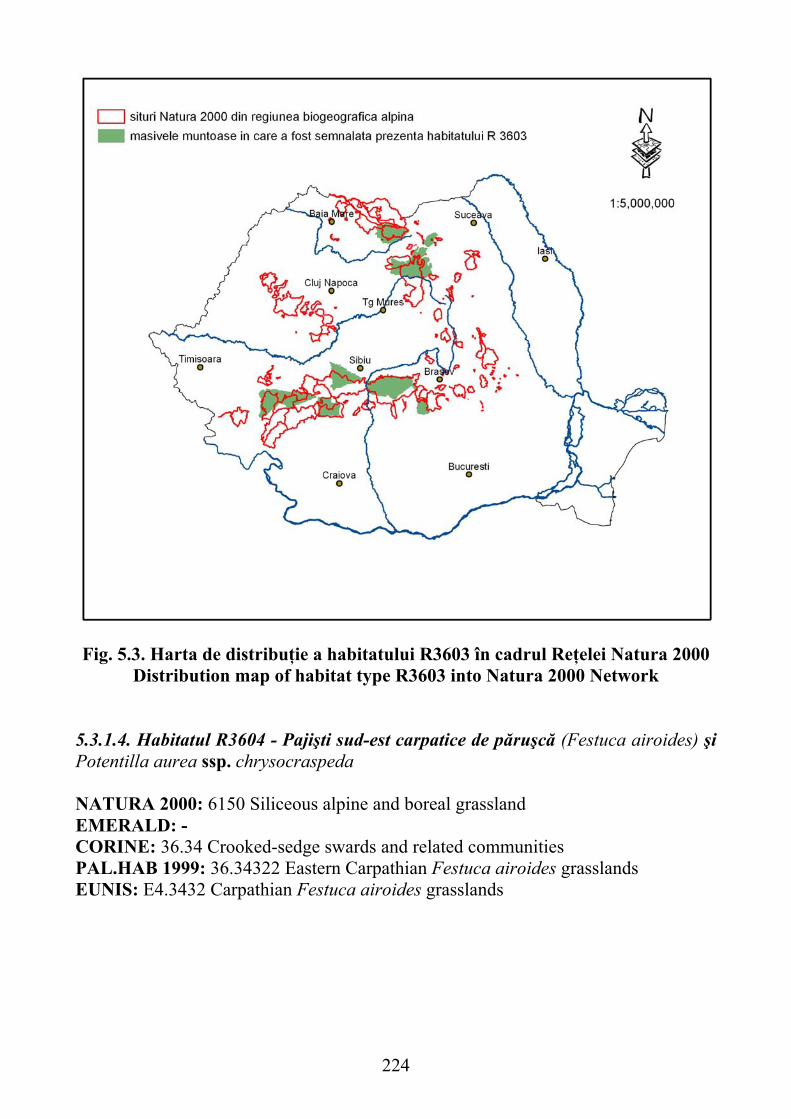
224
Fig. 5.3. Harta de distribuţie a habitatului R3603 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3603 into Natura 2000 Network 5.3.1.4. Habitatul R3604 - Pajişti sud-est carpatice de păruşcă (Festuca airoides) şi Potentilla aurea ssp. chrysocraspeda NATURA 2000: 6150 Siliceous alpine and boreal grassland EMERALD: - CORINE: 36.34 Crooked-sedge swards and related communities PAL.HAB 1999: 36.34322 Eastern Carpathian Festuca airoides grasslands EUNIS: E4.3432 Carpathian Festuca airoides grasslands

225
Asociaţii vegetale: Potentillo chrysocraspedae-Festucetum airoidis Boşcaiu 1971 (Syn.: Festucetum supinae Domin 1933; Potentillo (ternatae) - Festucetum supinae Boşcaiu 1971; Antennario-Festucetum sudeticae Dihoru 1975; As. Festuca supina-Nardus stricta Şerbănescu 1939; As. Festuca supina Puşcaru et al. 1959; Festuca supina- Deschampsia flexuosa-Vaccinium myrtillus Puşcaru et al. 1959; Festuca supina-Festuca rubra fallax Puşcaru et al. 1959; Festucetum supinae Vicol et al. 1971; Festucetum supinae subalpinae Sillinger 1933). Scurtă descriere: Pajiştile edificate de Festuca airoides acoperă suprafeţe mari din etajul alpin . Specia edificatoare Festuca airoides prezintă o acoperire foarte mare. Prin practicarea unui păşunat neraţional, pajiştile edificate de Festuca supina şi Potentilla aurea ssp. chrysocraspeda, se degradează şi evoluează către nardete. Condiţii staţionale: Altitudine: 1550-2500 m. Clima: T = 3,0- -2,50C; P = 800 mm. Relief: locuri plane, versanţi şi coame moderat înclinate până la repezi, uneori chiar pe grohotişuri fixate. Substrat: cristalin, mai rar calcaros. Soluri: spodisoluri sau humosiosoluri, puţin profunde până la superficiale, foarte acide sau slab acide (pH = 4,1-4,5), adeseori bogate în humus, reavene până la uscate. Specii dominante şi caracteristice: Potentilla aurea ssp. chrysocraspeda, Festuca airoides, Campanula alpina, Agrostis rupestris, Avenula versicolor, Hieracium alpinum, Primula minima, Sesleria coerulans, Phyteuma confusum. Valoare conservativă: are o valoare conservativă mare atât prin faptul că este un habitat endemic cât şi prin speciile endemice şi rare pe care le adăposteşte (Phyteuma confusum, Vaccinium gaultheroides, Leontopodium alpinum etc.). Corologie: Munţii Rodnei (Faţa Cătinului, Cobăşel, vârfurile Rebra, Bătrâna, Gardina, Galaţiul, Cormaia, Cişa), Vârfu Goru (jud. Vrancea), Muntele Siriu, Munţii Bucegi (Lăptici, Piatra Arsă, Cocora, vârfurile Baba Mare, Caraiman, Coştila, Morarul, Bucşoiu, Omu, Obârşia, Bătrâna, Vf. Cu Dor, Furnica, Jepii Mari, Jepii Mici), Muntele Gârbova, Munţii lezer-Păpuşa, Munţii Piatra Craiului, Muntele Ciucaş, Munţii Făgăraş, Munţii Cibinului, Munţii Parâng, Munţii Retezat, Munţii Ţarcu, Munţii Godeanu, Munţii Cernei. Starea actuală de protecţie: - aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (parţial sau total) în situri din cadrul Reţelei Natura 2000 (fig. 5.4).

226
Fig. 5.4. Harta de distribuţie a habitatului R3604 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3604 into Natura 2000 Network 5.3.1.5. Habitatul R3605 - Pajişti sud-est carpatice de păiuş de colţi (Festuca versicolor) şi Sesleria rigida ssp. haynaldiana NATURA 2000: 6170 Alpine and subalpine calcareous grasslands EMERALD: - CORINE: 36.4 Alpine and subalpine calciphilous grasslands PAL.HAB 1999: 36.43922 East Carpathian Festuca versicolor grasslands EUNIS: E4.4392 East Carpathian calciphile stepped grasslands Asociaţii vegetale: Seslerio -Festucetum versicoloris Beldie 1967 (Syn.: Festucetum versicoloris Puşcaru et al. 1956; Festucetum versicoloris transsilvanicum Soó 1944).

227
Scurtă descriere: Acest tip de habitat se dezvoltă pe soluri bogate în carbonat de calciu şi grad ridicat de saturaţie în baze de pe crestele şi poliţele din etajele alpin şi subalpin având un caracter mezoterm-heliofil. Condiţii staţionale: Altitudine: 2100-2500 m. Clima: T = -0,6- -2,50C; P = 1400-1450 mm. Relief: pante mai mult sau mai puţin înclinate, într-un microclimat caracterizat printr-o puternică insolaţie, în locurile adăpostite de acţiunea intensă a vânturilor. Substrat: calcare, conglomerate calcaroase. Soluri: rendzine bogate în humus, amestecate cu sfărâmături de calcar, care asigură saturaţia în carbonat de calciu, astfel încât reacţia se menţine neutră sau slab alcalină (pH = 7-7,4). Specii dominante şi caracteristice: Festuca versicolor, Sesleria rigida ssp. haynaldiana, Cerastium transsilvanicum, Bupleurum falcatum ssp. cernuum, Carduus kerneri, Festuca amethystina, Potentilla crantzii, Androsace lactea, Sesleria bielzii, Helianthemum nummularium ssp. tomentosum, Festuca saxatilis, Asperula capitata, Biscutella laevigata, Anthyllis alpestris, Aster alpinus, Campanula cochleariifolia, Carex sempervirens, Cerastium alpinum, Minuartia verna, Nigritella nigra, Scabiosa lucida, Dryas octopetala, Euphrasia salisburgensis, Festuca saxatilis, Festuca violacea, Galium anisophyllon, Gypsophila petraea, Hieracium villosum, Pedicularis verticillata, Phyteuma orbiculare, Poa alpina, Seseli libanotis, Achillea schurii, Dianthus callizonus, Eritrichium nanum ssp. jankae,Dianthus spiculifolius, Dianthus tenuifolius etc. Valoare conservativă: are o valoare conservativă mare fiind un habitat endemic şi adăpostind numeroase specii de pe listele roşii (Festuca amethystina, Nigritella nigra, Scabiosa lucida ssp. barbata, Gypsophila petraea, Achillea schurii, Dianthus callizonus, Eritrichium nanum ssp. jankae, Dianthus spiculifolius, Dianthus tenuifolius, Linum perenne ssp. extraaxilare etc.). Corologie: Munţii Rodnei (Corongişul Mic, Corongişul Mare, Vf. Faţa Cătinului, Vf. Mihăiasa), Munţii Bucegi (Colţii Obârşiei, vârfurile Baba Mare, Jepii Mari, Caraiman, Colţii Morarului, Strunga, Bucşoiu), Munţii Piatra Craiului, Munţii Făgăraş, Munţii Retezat. Starea actuală de protecţie: - toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.5).
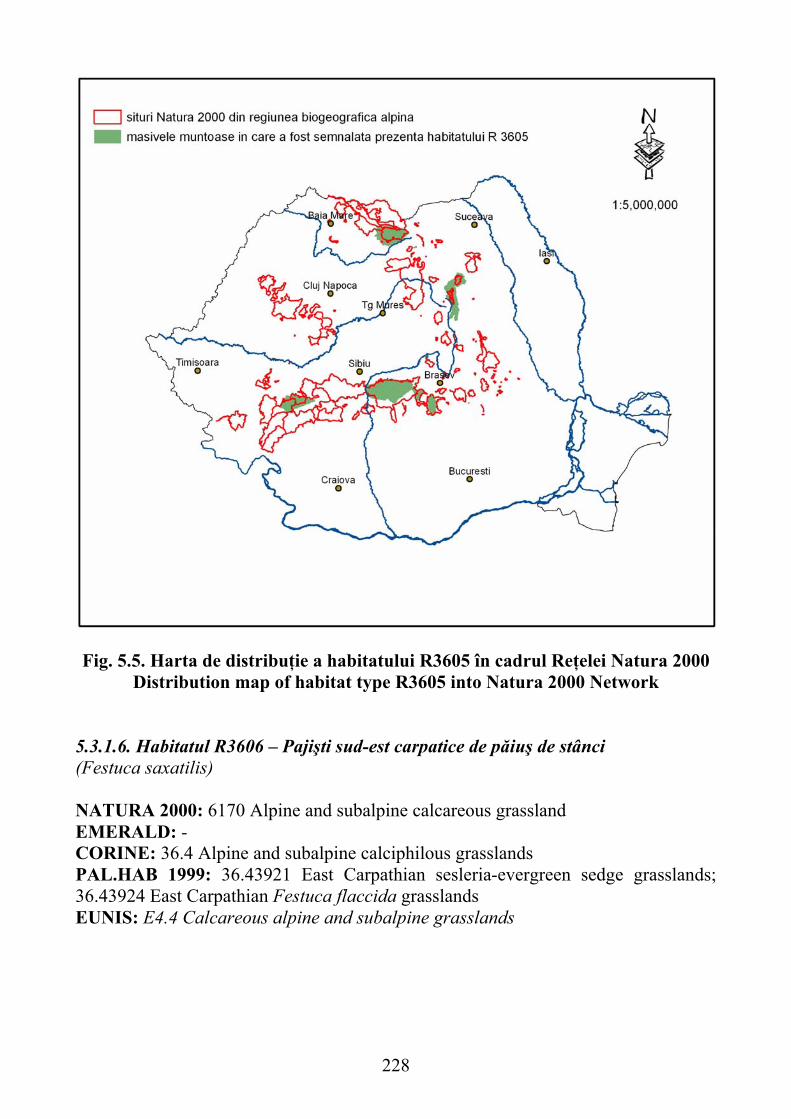
228
Fig. 5.5. Harta de distribuţie a habitatului R3605 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3605 into Natura 2000 Network 5.3.1.6. Habitatul R3606 – Pajişti sud-est carpatice de păiuş de stânci (Festuca saxatilis) NATURA 2000: 6170 Alpine and subalpine calcareous grassland EMERALD: - CORINE: 36.4 Alpine and subalpine calciphilous grasslands PAL.HAB 1999: 36.43921 East Carpathian sesleria-evergreen sedge grasslands; 36.43924 East Carpathian Festuca flaccida grasslands EUNIS: E4.4 Calcareous alpine and subalpine grasslands

229
Asociaţii vegetale: Festucetum saxatilis Domin 1933 (Syn.: Festucetum saxatilis Puşcaru et al. 1956; Seslerio -Festucetum saxatilis Beldie 1967; Festucetum saxatilis subalpinum auct. roman.; Caricetum sempervirentis Beldie 1967 p.p.); Festuco saxatilis - Trisetetum (macotrichum) Popescu et Sanda 1989; Festucetum flaccidae Coldea 1984. Scurtă descriere: Stratul ierbos este dominat de Festuca saxatilis alături de care, la edificarea habitatului mai constribuie şi Carex sempervirens, Sesleria rigida ssp haynaldiana etc. Fitocenozele acestui habitat endemic se întâlnesc pe suprafeţe reduse din etajul subalpin al Carpaţilor Orientali şi Meridionali. Condiţii staţionale: Altitudine: 1200-2300 m. Clima: T = 4,0- -2,50 C; P = 1100-1450 mm. Relief: stănci calcaroase însorite. Substrat: stânci calcaroase. Soluri: rendzine, superficiale, cu o reacţie slab acidă până la neutră (pH = 6,4-6,8), uneori chiar slab bazică. Specii dominante şi caracteristice: Festuca saxatilis (F. rupicola ssp. saxatilis), Festuca nitida ssp. flaccida, Dianthus tenuifolius, Sesleria bielzii, Helianthemum nummularium ssp. tomentosum, Festuca flaccida, Festuca versicolor, Alyssum repens, Carduus kerneri, Festuca amethystine, Sesleria rigida ssp. haynaldiana, Primula elatior ssp. carpatica, Centaurea kotschyana, Cerastium arvense ssp. calcicolum, Potentilla crantzii, Poa rehmanii, Centaurea pinnatifida, Trisetum fuscum, Carex sempervirens, Phyteuma orbiculare, Bartsia alpina, Galium anisophyllon, Polygonum viviparum, Myosotis alpestris, Pedi-cularis verticillata, Hieracium villosum, Helianthemum nummularium ssp. grandiflorum, Carex atrata, Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thesium kernerianum, Thymus pulcherrimus. Valoare conservativă: - fiind un habitat endemic care adăposteşte numeroase specii de pe liste roşii (Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thesium kernerianum, Thymus pulcherrimus etc.) aceste pajişti au o valoare conservativă mare. Corologie: Munţii Rodnei, Munţii Rarău, Muntele Hăşmaşu Mare, Muntele Siriu, Munţii Bucegi, Munţii Ciucaş, Muntele Postăvarul, Muntele Gârbova, Munţii Piatra Craiului, Munţii Făgăraş, Munţii Retezat. Starea actuală de protecţie: - toate masivele muntoase în care acest habitat este răspândit sunt incluse în parcuri naţionale şi situri din cadrul Reţelei Natura 2000 (Fig.5.6).
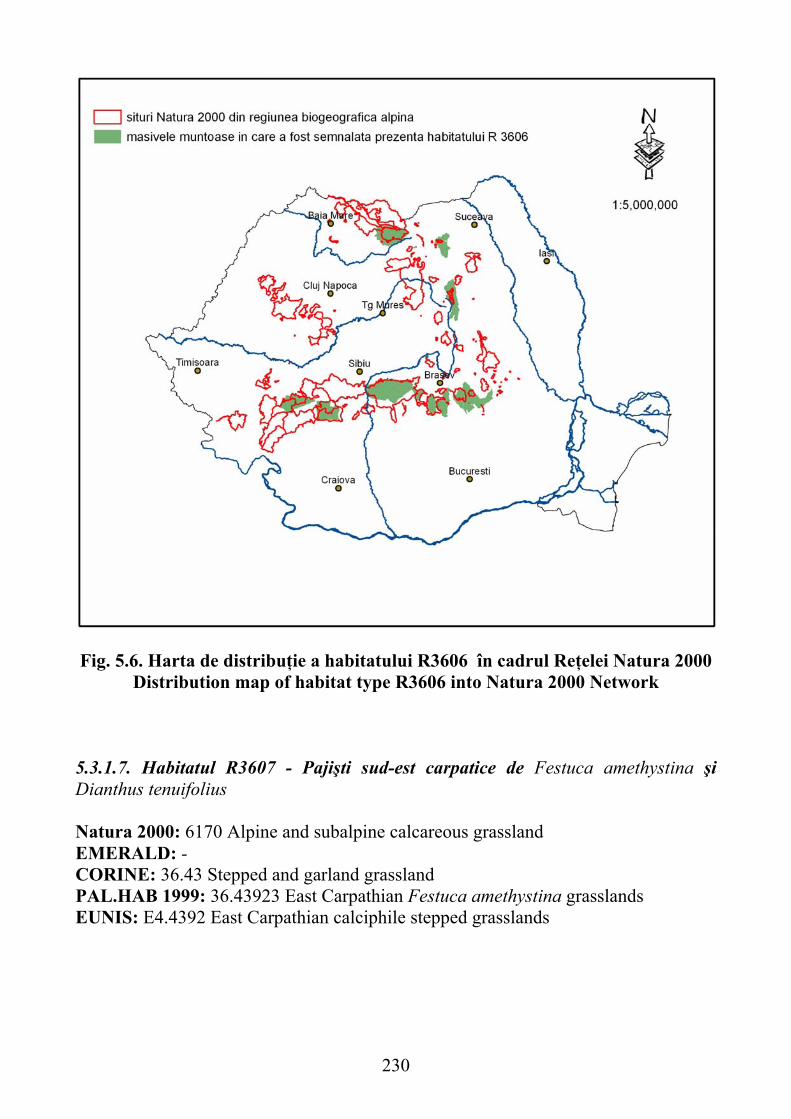
230
Fig. 5.6. Harta de distribuţie a habitatului R3606 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3606 into Natura 2000 Network 5.3.1.7. Habitatul R3607 - Pajişti sud-est carpatice de Festuca amethystina şi Dianthus tenuifolius Natura 2000: 6170 Alpine and subalpine calcareous grassland EMERALD: - CORINE: 36.43 Stepped and garland grassland PAL.HAB 1999: 36.43923 East Carpathian Festuca amethystina grasslands EUNIS: E4.4392 East Carpathian calciphile stepped grasslands

231
Asociaţii vegetale: Diantho tenuifolii -Festucetum amethystinae (Domin 1933) Coldea 1984 (Syn.: Festucetum amethystinae Puşcaru et al. 1956; Festucetum amethystinae transsilvanicum Nyárády 1967). Scurtă descriere: este un habitat xerofil, întâlnit în staţiuni ferite de vânt şi în care speciile caracteristice şi dominante ale asociaţiei sunt endemite carpatice, Festuca amethystina şi Dianthus tenuifolius. Condiţii staţionale: Altitudine: 1500-2200 m. Clima: T = 3,2- -1,50 C; P = 1200-1450 mm. Relief: versanţii calcaroşi, însoriţi, pe pante înclinate de 10-450. Substrat: calcaros. Soluri: rendzine cu pH = 5,8-7,5. Specii dominante şi caracteristice: Festuca amethystina, Dianthus tenuifolius, Sesleria bielzii, Helianthemum nummularium ssp. tomentosum, Festuca saxatilis, Festuca versicolor, Alyssum repens, Carduus kerneri, Sesleria rigida ssp. haynaldiana, Primula elatior ssp. carpatica, Centaurea kotschyana, Cerastium arvense ssp. calcicolum, Potentilla thuringiaca, Poa rehmanii, Cerastium transsilvanicum, Centaurea pinnatifida, Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thymus pulcherrimus. Valoare conservativă: fiind un habitat endemic care adăposteşte şi numeroase specii aflate pe liste roşii (Cerastium transsilvanicum, Centaurea pinnatifida, Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thymus pulcherrimus, Poa rehmanii etc.) are o valoare conservativă ridicată. Corologie: Munţii Rodnei, Munţii Rarău, Muntele Hăşmaşu Mare, Munţii Ciucului, Muntele Siriu, Munţii Bucegi, Munţii Făgăraş. Starea actuală de protecţie: - toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.7).
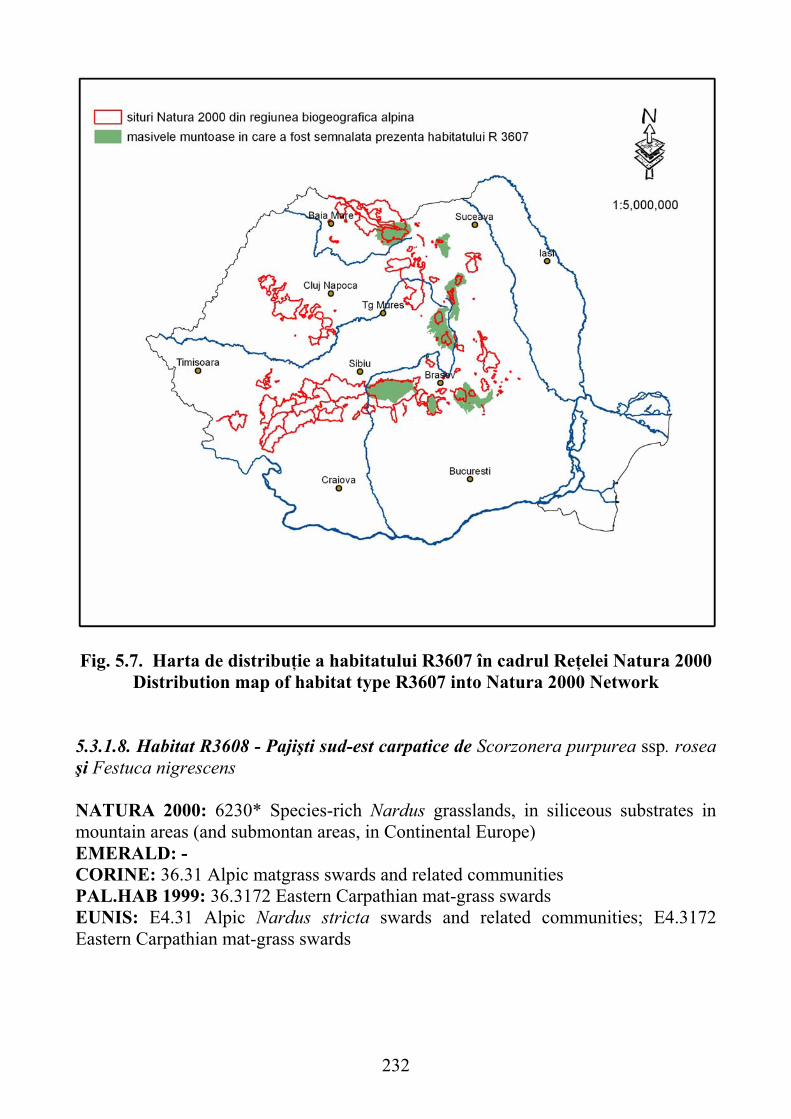
232
Fig. 5.7. Harta de distribuţie a habitatului R3607 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3607 into Natura 2000 Network 5.3.1.8. Habitat R3608 - Pajişti sud-est carpatice de Scorzonera purpurea ssp. rosea şi Festuca nigrescens NATURA 2000: 6230* Species-rich Nardus grasslands, in siliceous substrates in mountain areas (and submontan areas, in Continental Europe) EMERALD: - CORINE: 36.31 Alpic matgrass swards and related communities PAL.HAB 1999: 36.3172 Eastern Carpathian mat-grass swards EUNIS: E4.31 Alpic Nardus stricta swards and related communities; E4.3172 Eastern Carpathian mat-grass swards

233
Asociaţii vegetale: Scorzonero roseae -Festucetum nigricantis (Puşcaru et al. 1956) Coldea 1978 (Syn.: Festucetum rubrae fallax Puşcaru et al. 1956, Festucetum rubrae montanum Csűrös et Resmeriţă 1960). Scurtă descriere: Habitat mezofil şi mezo-higrofil, de pajişte secundară dezvoltată în urma defrişării pădurilor de molid. Stratul arbustiv - foarte redus; în pajişti pătrund specii arbustive, dintre care: Vaccinium myrtillus, Vaccinium vitis-idaea.Stratul ierbos - dominant, speciile caracteristice Scorzonera purpurea ssp. rosea şi Campanula abietina definesc caracterul regional al grupării. Specia edificatoare Festuca nigrescens realizează o acoperire cuprinsă între 35-85%. În structura floristică sunt prezente numeroase specii mezofile, caracteristice pentru Arrhenatheretea. Specia Nardus stricta are o constanţă ridicată şi o acoperire de până la 25-35%. Condiţii staţionale: Altitudine: 900-2340 m. Clima: T = 6,0- -1,50C; P = 950-1450 mm. Relief: versanţi, platouri. Substrat: diferit. Soluri: disticambosoluri, cu profil scurt şi saturate în baze (20-25%) şi pH = 4-4,5. Specii dominante şi caracteristice: Scorzonera purpurea ssp. rosea, Festuca nigrescens, Viola declinata, Nardus stricta, Poa media, Tozzia carpathica, Geum montanum, Potentilla aurea ssp. chrysocraspeda,Campanula serrata, Ligusticum mutellina, Gentiana kochiana, Hieracium aurantiacum, Hipochoeris uniflora, Thymus balcanus, Antennaria dioica, Euphrasia stricta, Hieracium pillosela, Potentilla erecta, Luzula sudetica, Thymus pulegioides, Alchemilla glaucescens, Danthonia decumbens, Hypericum umbellatum, Arnica montana, Luzula campestris, Alchemilla flabellata, Polygala vulgaris, Nigritella rubra. Valoare conservativă: are o valoare conservativă moderată fiind un habitat endemic şi prioritar la nivel european, care adăposteşte specii aflate pe listele roşii (Viola declinata, Scorzonera rosea ssp purpurea, Gymnadenia conopsea, Campanula serrata etc.). Corologie: habitat comun în tot lanţul carpatic în etajul montan superior şi subalpin. Starea actuală de protecţie: - toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.8.).
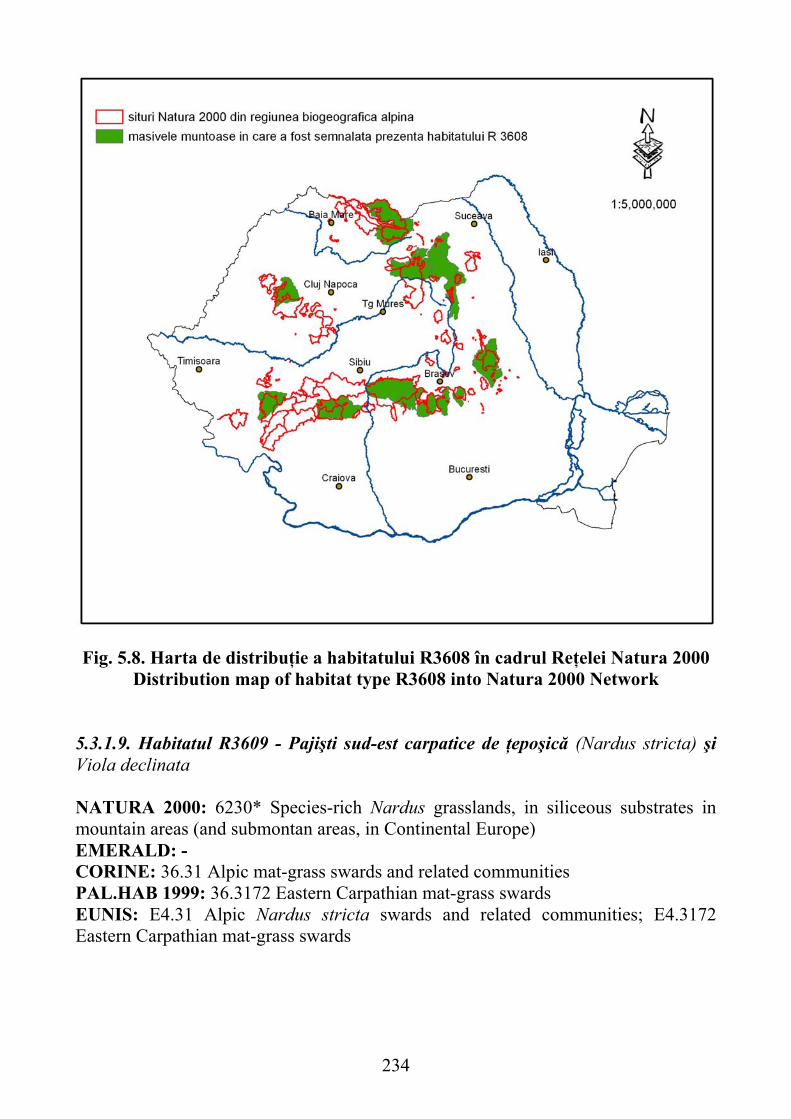
234
Fig. 5.8. Harta de distribuţie a habitatului R3608 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3608 into Natura 2000 Network 5.3.1.9. Habitatul R3609 - Pajişti sud-est carpatice de ţepoşică (Nardus stricta) şi Viola declinata NATURA 2000: 6230* Species-rich Nardus grasslands, in siliceous substrates in mountain areas (and submontan areas, in Continental Europe) EMERALD: - CORINE: 36.31 Alpic mat-grass swards and related communities PAL.HAB 1999: 36.3172 Eastern Carpathian mat-grass swards EUNIS: E4.31 Alpic Nardus stricta swards and related communities; E4.3172 Eastern Carpathian mat-grass swards

235
Asociaţii vegetale: Violo declinatae -Nardetum Simon 1966 (Syn.: Nardetum strictae montanum Resmeriţă et Csűrös 1963, Nardetum strictae alpinum Buia et al. 1962, Nardetum alpigenum austro - carpaticum Borza 1959). Scurtă descriere: Habitat oligotrof, xerofil, acidofil. Stratul arbustiv este foarte redus; în pajişti pătrunzând specii arbustive din fitocenozele învecinate, dintre care Vaccinium myrtillus, Vaccinium vitis-idaea sunt cel mai frecvent întâlnite. În stratul ierbos se întâlneşte frecvent, specia caracteristică carpato-balcanică Viola declinata. Festuca nigrescens are o constanţă ridicată, dar cu o acoperire destul de redusă, mai ales în cazul habitatelor degradate. Pajiştile de ţepoşică, degradate ca urmare a păşunatului neraţional nu sunt incluse în cadrul habitatului Natura 2000 6230*. Au fost descrise subasociaţiile typicum, care este întâlnită în etajul montan superior şi festucetosum airoidis Coldea 1987, prezentă în etajul subalpin, cu numeroase specii microterme şi cu diferenţialele ecologice. Condiţii staţionale: Altitudine: 500-2070 m. Clima: T = 6,0-00C; P = 900-1400 mm. Relief: platouri, versanţi, văi şi coaste domoale până la moderat înclinate. Substrat: acid. Soluri: spodisoluri cu profil scurt, sărace în baze (5-10%), slab aerate şi acide pH = 3,6-4,5. Specii dominante şi caracteristice: Viola declinata, Nardus stricta, Scorzonera rosea, Poa media, Hieracium aurantiacum, Hypochoeris uniflora, Calluna vulgaris, Campanula serrata, Geum montanum, Ligusticum mutellina, Potentilla aurea ssp. chrysocraspeda,Campanula abietina, Leucorchis albida, Genista sagittalis, Festuca nigrescens, Antennaria dioica, Luzula campestris, Carex ovalis, Polygala vulgaris, Euphrasia stricta, Hieracium pilosella, Hieracium lactucella, Potentilla erecta, Luzula sudetica, Alchemilla glaucescens, Alchemilla flabellate, Carex pallescens, Danthonia decumbens, Hypericum maculatum, Arnica montana. Valoare conservativă: are o valoare conservativă moderată fiind un habitat prioritar la nivel european. Acest habitat adăposteşte şi o serie de specii de pe listele roşii (Viola declinata, V. dacica, Campanulla serrata, Scorzonera rosea ssp purpurea, Gymnadenia conopsea, Arnica montana, Anthemis macrantha etc.). Corologie: habitat comun în tot lanţul carpatic în etajul montan şi subalpin. Starea actuală de protecţie: - aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.9.).
236
Fig. 5.9. Harta de distribuţie a habitatului R3609 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3609 into Natura 2000 Network 5.3.1.10. Habitatul R3610 - Pajişti sud-est carpatice de Poa media NATURA 2000: - EMERALD: - CORINE: 36.31 Alpic mat-grass swards and related communities PAL.HAB 1999: 36.3172 Eastern Carpathian mat-grass swards EUNIS: E4.31 Alpic Nardus stricta swards and related communities Asociaţii vegetale: Poetum mediae Csűrös et al. 1956.

237
Scurtă descriere: Pajiştile subalpine edificate de Poa media sunt pajişti întâlnite pe platouri şi versanţi puţin înclinaţi pe care zăpada persistă până primăvara târziu. Caracterul chiono-higrofil este subliniat de prezenţa unor specii ca: Ranunculus crenatus, Gnaphalium supinum etc.. Grupările cu Poa media prezintă numeroase specii caracteristice pentru Potentillo-Nardion şi Caricetalia curvulae.Din punct de vedere dinamic, aceste fitocenoze tind să evolueze spre pajişti de ţepoşică. Condiţii staţionale: Altitudine: 1800-2000 m. Clima: T = 0,7- -0,00C; P = 950-1450 mm. Relief: versanţi, platouri. Substrat: diferit. Soluri: humosiosoluri, bogate în substanţe nutritive şi reacţie acidă pH = 5. Specii dominante şi caracteristice: Poa media, Viola declinata, Nardus stricta, Agrostis rupestris, Avenula versicolor, Geum montanum, Potentilla ternata, Campanula serrata, Campanula abietina, Plantago atrata, Pedicularis verticillata, Leontodon croceus, Veronica alpina, Veronica officinalis, Alchemilla glabra, Cerastium fontanum ssp. triviale, Cardaminopsis halleri ssp. ovirensis, Euphrasia minima, Hypochoeris uniflora, Vaccinium uliginosum, Alchemilla obtusa, Hieracium aurantiacum, Phleum alpinum ssp. commutatum, Anthoxantum odoratum, Alchemilla flabellata, Achillea stricta, Festuca nigrescens, Antennaria dioica, Achillea distans, Alchemilla xanthochlora, Luzula multiflora, Leucorchis albida, Hypericum richeri ssp. grisebachii, Lychnis nivalis, Thlaspi dacicum. Valoare conservativă: moderată dată de prezenţa unor specii endemice sau rare aflate pe listele roşii naţionale, cum ar fi: Viola declinata, Leucorchis albida, Hypericum richeri ssp. grisebachii, Lychnis nivalis, Thlaspi dacicum etc. Corologie: Munţii Maramureşului, Munţii Rodnei, Munţii Bucegi, Munţii Făgăraş, Munţii Retezat, Munţii Parâng. Starea actuală de protecţie: - toate masivele muntoase în care a fost semnalată prezenţa acestui habitat sunt incluse în situri din cadrul reţelei Natura 2000 (Fig. 5.10).

238
Fig. 5.10. Harta de distribuţie a habitatului R3610 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3610 into Natura 2000 Network 5.3.1.11. Habitatul R3611 – Pajişti sud-est carpatice de coada iepurelui (Sesleria rigida ssp haynaldiana) şi rogoz (Carex sempervirens) NATURA 2000: 6170 Alpine and subalpine calcareous grassland EMERALD: - CORINE: 36.4 Alpine and subalpine calciphilous grassland PAL.HAB 1999: 36.43921 East Carpathian sesleria-evergreen sedge grasslands EUNIS: E4.4392 East Carpathian calciphile stepped grasslands Asociaţii vegetale: Seslerio haynaldianae- Caricetum sempervirentis Puşcaru et al. 1956 (Syn.: Seslerietum haynaldianae sempervirentis Puşcaru et al. 1950,1956;

239
Seslerietum rigidae retezaticum Csűrös et al. 1956; Seslerietum rigidae biharicum Csűrös 1963, Resmeriţă 1965); Seslerio haynaldianae - Saxifragetum rochelianae Boşcaiu 1971 (Syn.: Seslerietum rigidae retezaticum Csűrös 1956); Seslerio rigidae - Saxifragetum rochelianae Gergely 1967; Seslerio heuflerianae - Caricetum sempervirentis Coldea 1984 (Syn.: Seslerietum heuflerianae auct. rom.). Scurtă descriere: Habitat mezoterm-heliofil, a cărei compoziţie floristică cuprinde un grup ridicat de specii relictare termofile. Stratul ierbos este dominat de Sesleria rigida ssp. haynaldiana şi Carex sempervirens. Condiţii staţionale: Altitudine: 1500-2400 m. Clima: T = 2,5- -2,50C; P = 1200-1450 mm. Relief: stânci calcaroase, brâne, versanţi diferiţi, însoriţi sau cu expoziţie nordică. Substrat: calcaros. Soluri: rendzine, cu conţinut bogat de carbonat de calciu şi grad ridicat de saturaţie în baze, cu o reacţie neutră (pH = 6,8-7). Specii dominante şi caracteristice: Sesleria rigida ssp. haynaldiana, Carex sempervirens, Saxifraga marginata, Sesleria heufleriana, Dianthus tenuifolius, Helianthemum nummularium ssp. tomentosum, Festuca saxatilis, Alyssum repens, Festuca amethystina, Primula elatior ssp. carpatica, Centaurea kotschyana, Cerastium arvense ssp. calcicolum, Potentilla thuringiaca, Poa rehmanii, Anthyllis alpestris, Hedysarum hedysaroides, Carduus kerneri, Alyssum repens, Geranium coerulatum, Calamintha alpina ssp. baumgarteni, Bromus riparius ssp. barcensis, Trollius europaeus, Centaurea pinnatifida, Cerastium transsilvanicum, Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thesium kernerianum, Thymus pulcherrimus. Valoare conservativă: fiind un habitat endemic şi adăpostind un număr mare de specii care se regăsesc pe listele roşii (Centaurea pinnatifida, Cerastium transsilvanicum, Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thesium kernerianum Thymus pulcherrimus etc.), are o valoare conservativă mare. Corologie: Munţii Rodnei, Munţii Rarău, Muntele Hăşmaşu Mare, Munţii Bucegi, Munţii Piatra Craiului, Muntele Postăvarul, Munţii Ciucaş, Munţii Făgăraş, Munţii Vâlcanului, Munţii Retezat, Munţii Ţarcu, Munţii Godeanu, Munţii Cernei, Masivul Vlădeasa. Starea actuală de protecţie: - aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.11).

240
Fig. 5.11. Harta de distribuţie a habitatului R3611 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3611 into Natura 2000 Network 5.3.1.12. Habitatul R3612 – Pajişti sud-est carpatice de rogoz (Carex sempervirens) şi coarnă mare (Sesleria bielzii) NATURA 2000: 6170 Alpine and subalpine calcareous grassland EMERALD: - CORINE: 36.43 Stepped and garland grassland PAL.HAB 1999: 36.43921 East Carpathian sesleria-evergreen sedge grasslands EUNIS: E4.4392 East Carpathian calciphile stepped grasslands Asociaţii vegetale: Seslerio bielzii -Caricetum sempervirentis Puşcaru et al. 1956 (Syn.: Seslerietum bielzii transsilva-nicum Borhidi 1956, 1958); Poo alpinae - Alyssetum repentis Beldie 1967.

241
Scurtă descriere: Habitat mozaicat, datorat variabilităţii condiţţilor staţionale în care se dezvoltă. Stratul ierboseste edificat de Carex sempervirens şi Sesleria bielzii, alături de care, cu constanţă ridicată, se mai întâlnesc: Astragalus alpinus, Sausurea alpina, Cerastium transsilvanicum, Androsace chamaejasme, Astragalus frigidus, Bartschia alpina, Hedysarum hedysaroides, Biscutella laevigata. În condiţii de umiditate ridicată, în asociaţie se infiltrează adeseori elemente din Salicetum reticulatae. Condiţii staţionale: Altitudine: 1650-2200 m. Clima: T = 2,0- -1,50C; P = 1200-1450 mm. Relief: stânci calcaroase însorite, în unele cazuri, umbrite. Substrat: calcaros. Soluri: soluri rankere, bogate în schelet, adeseori slab înţelenite, cu pH = 5,8-7,5. Specii dominante şi caracteristice: Sesleria bielzii, Carex sempervirens, Dianthus tenuifolius,Cerastium transsilvanicum, Festuca versicolor, Festuca flaccida, Helianthemum nummularium ssp. tomentosum, Carduus kerneri, Festuca amethystina, Sesleria rigida ssp. haynaldiana, Primula elatior ssp. carpatica, Centaurea kotschyana, Cerastium arvense ssp. calcicolum, Potentilla thuringiaca, Poa rehmanii, Centaurea pinnatifida, Alyssum repens, Alchemilla palmata, Calamintha baumgarteni, Anthemis carpatica ssp. pyrethriformis, Cerastium transsilvanicum, Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thesium kernerianum Thymus pulcherrimus. Valoare conservativă: fiind un habitat endemic care adăposteşte un număr mare de specii regasite pe listele roşii (Anthemis carpatica ssp. pyrethriformis, Cerastium transsilvanicum, Dianthus spiculifolius, Dianthus tenuifolius, Koeleria transsilvanica, Linum perenne ssp. extraaxilare, Onobrychis transsilvanica, Thlaspi dacicum, Thesium kernerianum Thymus pulcherrimus etc.), are o valoare conservativă ridicată. Corologie: Munţii Rarău, Masivul Ceahlău, Muntele Hăşmaşu Mare, Munţii Ciucaş, Munţii Bucegi, Muntele Gârbova, Muntele Postăvaru, Munţii Piatra Craiului, Munţii Făgăraş, Munţii Parâng, Munţii Ţarcu. Starea actuală de protecţie: - aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.12).
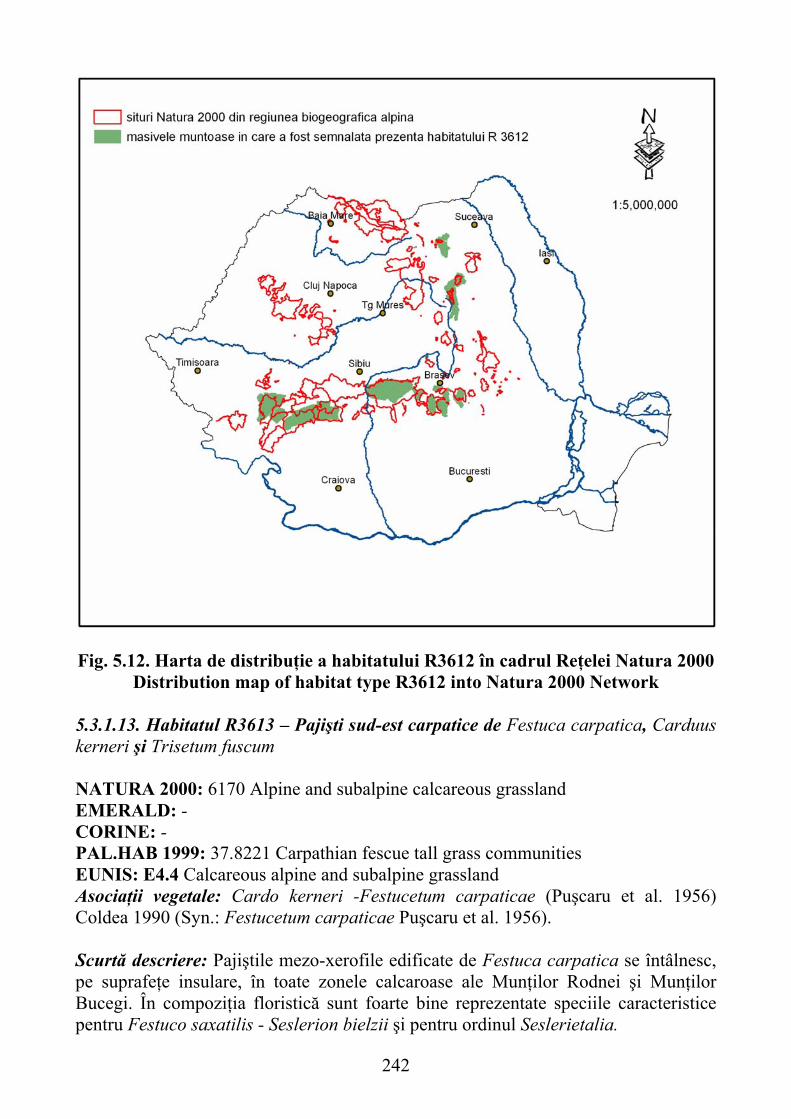
242
Fig. 5.12. Harta de distribuţie a habitatului R3612 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3612 into Natura 2000 Network 5.3.1.13. Habitatul R3613 – Pajişti sud-est carpatice de Festuca carpatica, Carduus kerneri şi Trisetum fuscum NATURA 2000: 6170 Alpine and subalpine calcareous grassland EMERALD: - CORINE: - PAL.HAB 1999: 37.8221 Carpathian fescue tall grass communities EUNIS: E4.4 Calcareous alpine and subalpine grassland Asociaţii vegetale: Cardo kerneri -Festucetum carpaticae (Puşcaru et al. 1956) Coldea 1990 (Syn.: Festucetum carpaticae Puşcaru et al. 1956). Scurtă descriere: Pajiştile mezo-xerofile edificate de Festuca carpatica se întâlnesc, pe suprafeţe insulare, în toate zonele calcaroase ale Munţilor Rodnei şi Munţilor Bucegi. În compoziţia floristică sunt foarte bine reprezentate speciile caracteristice pentru Festuco saxatilis - Seslerion bielzii şi pentru ordinul Seslerietalia.

243
Condiţii staţionale: Altitudine: 1390-2200 m. Clima: T = 3,5- -1,50C; P = 1200-1450 mm. Relief: versanti slab însoriţi. Substrat: grohotişuri calcaroase semifixate şi fixate. Soluri: rendzine, reavăne, superficiale, cu pH = 6,7-7. Specii dominante şi caracteristice: Festuca carpatica, Carduus kerneri, Trisetum fuscum, Sesleria bielzii, Festuca saxatilis, Alyssum repens, Festuca amethystina, Sesleria rigida ssp. haynaldiana, Primula elatior ssp. carpatica, Centaurea kotschyana, Bupleurum diversifolium, Poa rehmanii, Bartsia alpina, Carex sempervirens, Galium anisophyllon, Phyteuma orbiculare, Polygonum vivipa-rum, Ranunculus oreophilus, Myosostis alpestris, Pedicularis verticillata, Biscutella laevigata, Scabiosa lucida, Hieracium villo-sum, Anemone narcissiflora, Helianthemum nummularium ssp. grandiflorum, Hedy-sarum hedysaroides, Ranunculus thora, Astragalus frigidus, Crepis jacquinii, Sausurea discolor, Cerastium transsilvanicum, Linum perenne ssp. extraaxilare, Thymus pulcherrimus. Valoarea conservativă: este dată de caracterul endemic al habitatului şi de speciile regăsite pe listele roşii pe care le adăposteşte (Astragalus frigidus, Crepis jacquinii, Sausurea discolor, Cerastium transsilvanicum, Linum perenne ssp. extraaxilare, Thymus pulcherrimus etc.). Corologie: Munţii Rodnei, Munţii Bucegi. Starea actuală de protecţie: - toate masivele muntoase în care acest habitat este răspândit sunt incluse în situri din cadrul Reţelei Natura 2000 (Fig. 5.13).
244
Fig. 5.13. Harta de distribuţie a habitatului R3613 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3612 into Natura 2000 Network 5.3.1.14. Habitatul R3614 - Pajişti sud-est carpatice de Festuca xanthina NATURA 2000: 6170 Alpine and subalpine calcareous grassland EMERALD: 34.3 Dense perennial grasslands and middle European steppes CORINE: - PAL.HAB 1999: 34.35123 South Carpathian Festuca xanthina grasslands EUNIS: E4.4 Calcareous alpine and subalpine grassland Asociaţii vegetale: Festucetum xanthinae Boşcaiu 1971.

245
Scurtă descriere: Fitocenozele acestor pajişti xerofile vegetează pe versanţii abrupţi şi poliţele de la baza stâncilor calcaroase. Prin speciile caracteristice, pentru alianţa Seslerion rigidae, aceste fitocenoze prezintă particularităţi floristice distincte faţă de gruparea descrisă din estul Serbiei: Festucetum xanthino-variae. în regiunea montană şi etajul subalpin. Condiţii staţionale: Altitudine: 450-1700 m. Clima: T = 8-1,50C; P = 800 mm. Relief: creste şi versanţi abrupţi. Substrat: conglomerat calcaros. Soluri: rendzine, cu humus de tip mull. Specii dominante şi caracteristice: Festuca xanthina, Asperula capitata, Alyssum repens, Sesleria rigida, Seseli gracile, Seseli rigidum, Saxifraga marginata, Primula veris ssp. columnae, Dianthus petraeus, Erysimum witmannii, Athamantha turbith ssp. hungarica, Draba lasiocarpa, Scrophularia heterophylla ssp. laciniata, Phyteuma orbiculare, Aster alpinus, Ranunculus oreophilus, Carduus glaucus, Centaurea pinnatifida, Minuartia verna, Biscutella laevigata, Myosotis alpestris, Thymus comosus, Viola jooi. Valoare conservativă: - mare, atât datorită faptului că este un habitat endemic cât şi prezenţei multor specii endemice şi rare pentru flora României, cum ar fi: Erysimum witmannii, Athamantha turbith ssp. hungarica, Thymus comosus, Viola jooi etc. Corologie: Munţii Retezat, Munţii Ţarcu-Godeanu, Munţii Cernei, Piatra Cloşani (Fig. 5.14). Starea actuală de protecţie: - toate masivele muntoase în care se găseşte acest tip de habitat au fost incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.14).
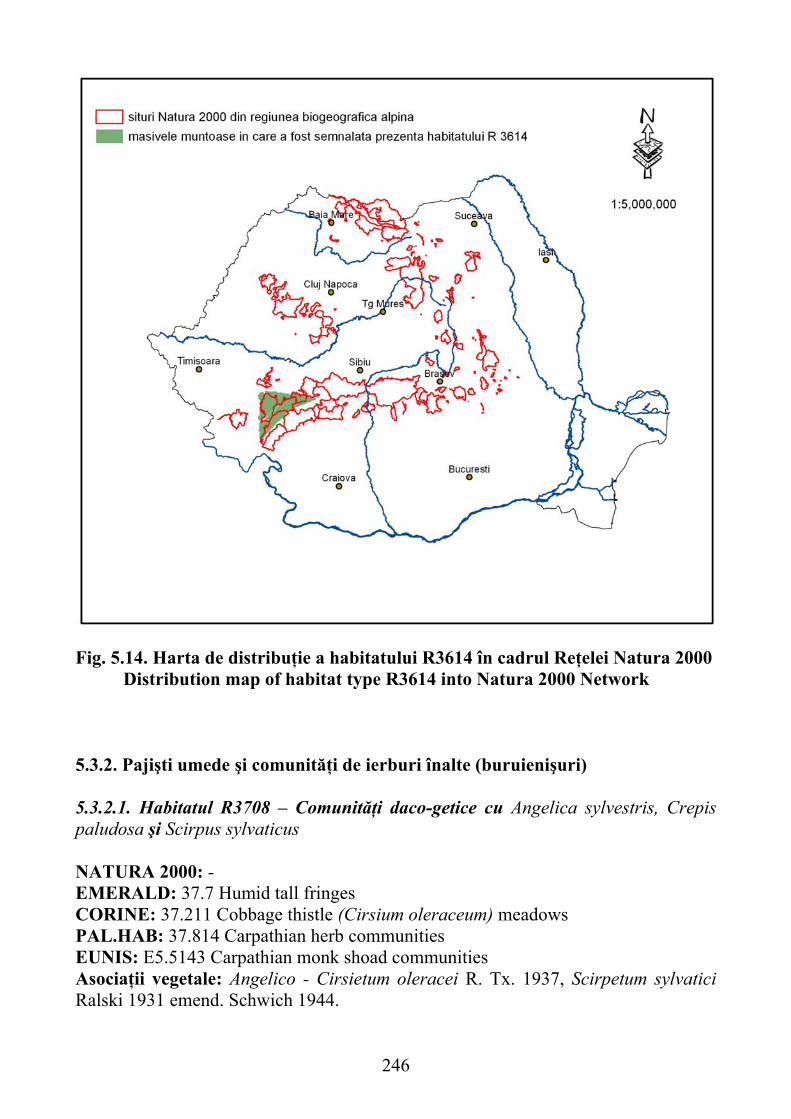
246
Fig. 5.14. Harta de distribuţie a habitatului R3614 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3614 into Natura 2000 Network 5.3.2. Pajişti umede şi comunităţi de ierburi înalte (buruienişuri) 5.3.2.1. Habitatul R3708 – Comunităţi daco-getice cu Angelica sylvestris, Crepis paludosa şi Scirpus sylvaticus NATURA 2000: - EMERALD: 37.7 Humid tall fringes CORINE: 37.211 Cobbage thistle (Cirsium oleraceum) meadows PAL.HAB: 37.814 Carpathian herb communities EUNIS: E5.5143 Carpathian monk shoad communities Asociaţii vegetale: Angelico - Cirsietum oleracei R. Tx. 1937, Scirpetum sylvatici Ralski 1931 emend. Schwich 1944.

247
Scurtă descriere: Sunt întâlnite insular în văile umede şi difuz luminate din etajele colinar şi montan inferior. Speciile edificatoare sunt plante înalte, cunoscute şi sub numele de buruieni din văile de munte, dintre care mai reprezentative sunt: Cirsium oleraceum, Angelica sylvestris, Cirsium rivularis, Filipendula ulmaria. Condiţii staţionale: Altitudine: 350-800 m. Clima: T = 8-70C; P = 650-800 mm. Relief: versanţii văilor, în apropiere de firul văii sau al ochiurilor de apă din lungul acestora. Soluri: aluviale, pseudo-gleice, bogate în substanţe nutritive. Specii dominante şi caracteristice: Scirpus sylvaticus, Angelica sylvestris, Cirsium oleraceum, Angelica archangelica, Crepis paludosa, Juncus articulatus, Ligularia sibirica, Polemonium caeruleum, Trolius europaeus. Valoarea conservativă: este redusă, din punct de vedere sozologic fiind importante doar acele pajişti care adăpostesc specii rare (Ligularia sibirica, Polemonium caeruleum, Trolius europaeus). Corologie: este un habit comun în etajul colinar şi montan inferior. Starea actuală de protecţie: - nu se cunoaşte, nici măcar cu aproximaţie, ce suprafaţă ocupată de acest tip de habitat este inclusă în situri din cadrul Reţelei Natura 2000. 5.3.2.2. Habitatul R3710 – Pajişti dacice de Molinia coerulea NATURA 2000: 6410 Molinia meadows on calcareous, peaty or clayey-siltladen soils (Molinion caeruleae) EMERALD: 37.3 Oligotrophic humid grasslands CORINE: 37.31 Purple moograss meadows PAL.HAB: 37.312 Acid purple moorgrass meadows EUNIS: E3.511 Calcidine purple moorgross meadows Asociaţii vegetale: Junco-Molinietum Preising 1951, Peucedano rocheliani -Molinietum coeruleae Boşcaiu 1965, Molinio - Salicetum rosmarinifoliae Magyar ex Soó 1933. Scurtă descriere: Fitocenozele acestui habitat se instalează pe terenuri cu umiditate în exces şi bogate în substanţe nutritive. În cadrul fitocenozelor participă, în număr mare, plante de talie înaltă ce depăşesc 1 m înălţime. Condiţii staţionale: Altitudine: 300-450(500) m. Clima: T = 9-80C; P = 700-850 mm.

248
Relief: depresiuni, cu pante foarte uşor înclinate, cu expoziţii variate. Substrat: depozite luto-argiloase. Soluri: brune luvice, brun acide de pantă, pe terenurile plate soluri turboase sau gleice cu conţinut ridicat de substanţe nutritive şi umiditate în exces. Specii dominante şi caracteristice: Molinia caerulea, Peucedanum rochelianum, Juncus conglomeratus şi Salix rosmarinifolia, Angelica sylvetris, Cirsium rivulare, Succisia pratensis, Stachys officinalis, Gentiana pneumonanthe, Galium boreale, Carex ovalis, Parnassia palustris, Lathyrus pratensis,Gladiolus imbricatus, Stellaria graminea. Valoare conservativă: are o valoare conservativă moderată prin speciile rare şi periclitate pe care le adăposteşte (Peucedanum rochelianum, Gentiana pneumonanthe, Gladiolus imbricatusetc.). Corologie: Maramureş, Transilvania, depresiunile intramontane din bazinul Oltului superior, Bazinul Giurgeului, Banat, Munţii Apuseni. Starea actuală de protecţie: - aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.15). 5.3.2.3. Habitatul R3712 – Comunităţi dacice cu Deschampsia caespitosa şi Agrostis stolonifera NATURA 2000: - EMERALD: 37.2 Eutrophic humid grasslands CORINE: - PAL.HAB: 37.263 Danubio-Pannonic riverine and humid meadows EUNIS: E2.233 Carpathian submontane hay meadows Asociaţii vegetale: Agrostio stoloniferae -Deschampsietum caespitosae Ujvárosi 1947.
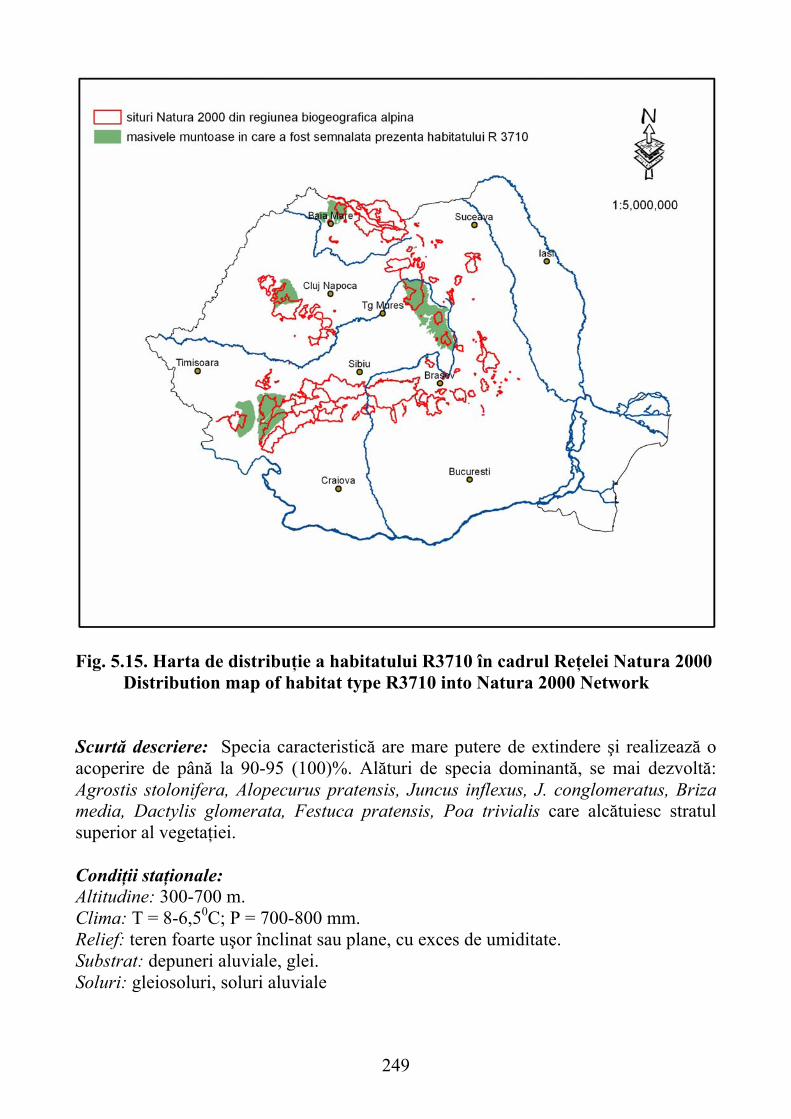
249
Fig. 5.15. Harta de distribuţie a habitatului R3710 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3710 into Natura 2000 Network Scurtă descriere: Specia caracteristică are mare putere de extindere şi realizează o acoperire de până la 90-95 (100)%. Alături de specia dominantă, se mai dezvoltă: Agrostis stolonifera, Alopecurus pratensis, Juncus inflexus, J. conglomeratus, Briza media, Dactylis glomerata, Festuca pratensis, Poa trivialis care alcătuiesc stratul superior al vegetaţiei. Condiţii staţionale: Altitudine: 300-700 m. Clima: T = 8-6,50C; P = 700-800 mm. Relief: teren foarte uşor înclinat sau plane, cu exces de umiditate. Substrat: depuneri aluviale, glei. Soluri: gleiosoluri, soluri aluviale

250
Specii dominante şi caracteristice: Deschampsia caespitosa, Agrostis stolonifera, Juncus conglomeratus, Phleum pratense, Festuca pratensis, Poa trivialis, P. palustris, Ranunculus acris, Alopecurus pratensis, Trifolium pratense, Briza media, Lathyrus pratensis, Cynosurus cristatus, Holcus lanatus, Cirsium canum. Valoare conservativă: are o valoare conservativă redusă. Corologie: Transilvania, iar în Oltenia, Muntenia şi Moldova, numai în zona colinară şi etajul montan inferior. Starea actuală de protecţie: - aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig. 5.16).
5.3.3. Pajişti mezofile 5.3.3.1. Habitatul R3801 Pajişti sud-est carpatice de Trisetum flavescens şi Alchemilla vulgaris NATURA 2000:6520 Mountain hay meadows EMERALD: !38.25 Continental meadows CORINE: 38.3 Montan hay meadows PAL.HAB: 38.2333 Eastern Carpathian yellow oatgrass meadows EUNIS: E2.2333 Eastern Carpathian yellow oatgrass meadows Asociaţii vegetale: Cerastio holosteoidis -Trisetum flavescenti Sanda et Popescu 2001 (Poo - Trisetum flavescetis auct. rom. non Knapp 1951), Trisetetum flavescentis (Schröter) Brockmann 1907. Scurtă descriere: Fitocenozele de Trisetum flavescens ocupă suprafeţe relativ mici pe terenuri slab înclinate din etajul montan inferior şi mijlociu. Acestea au în compoziţie numeroase specii de talie mare (60-80 cm), care realizează o acoperire de 80-95%. Alături de specia dominantă se dezvoltă frecvent: Agrostis capillaris, Phleum montanum, Cynosurus cristatus, Festuca pratensis, Arrhenatherum elatius, Onobrychis viciifolia, Leucanthemum vulgaris, Knautia arvensis, Campanula glomerata, Trifolium pratense, Lotus corniculatus, Luzula campestris, Gymnadenia conopsea, Carum carvi, Trifolium campestre, T. montanum, Cerastium holosteoides, specii care imprimă un caracter mezofil, micro-mezoterm şi eurionic acestor pajişti.
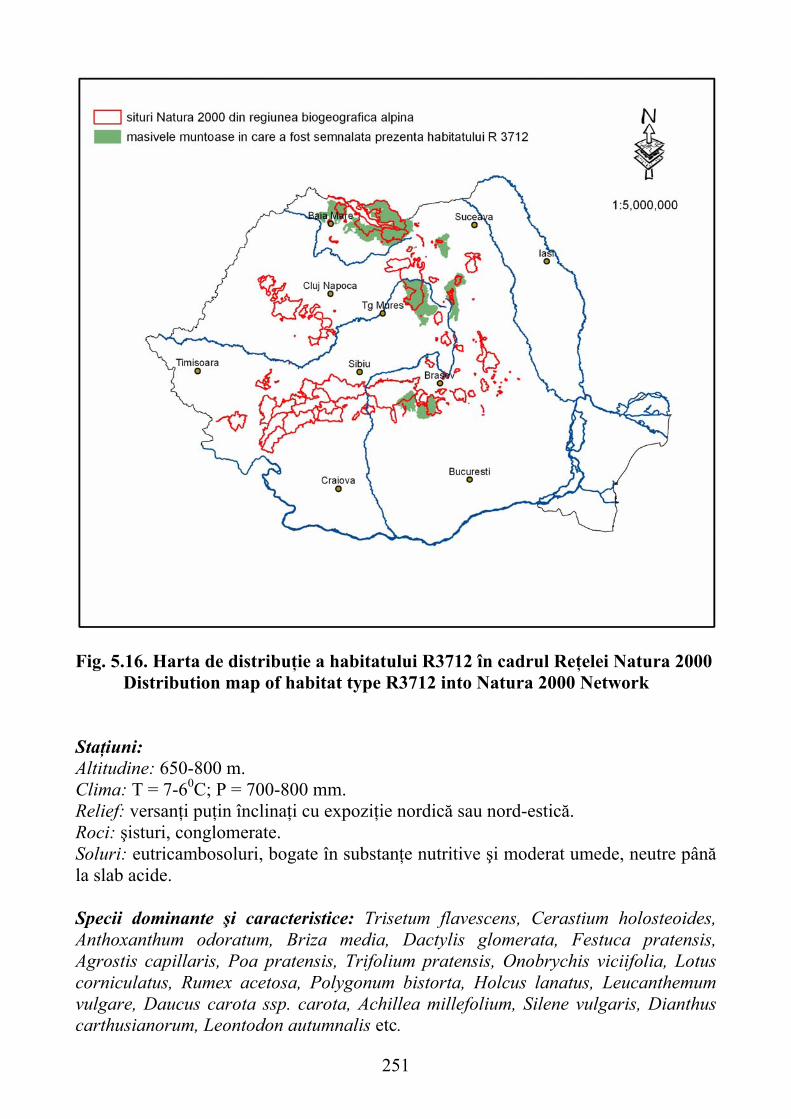
251
Fig. 5.16. Harta de distribuţie a habitatului R3712 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3712 into Natura 2000 Network Staţiuni: Altitudine: 650-800 m. Clima: T = 7-60C; P = 700-800 mm. Relief: versanţi puţin înclinaţi cu expoziţie nordică sau nord-estică. Roci: şisturi, conglomerate. Soluri: eutricambosoluri, bogate în substanţe nutritive şi moderat umede, neutre până la slab acide. Specii dominante şi caracteristice: Trisetum flavescens, Cerastium holosteoides, Anthoxanthum odoratum, Briza media, Dactylis glomerata, Festuca pratensis, Agrostis capillaris, Poa pratensis, Trifolium pratensis, Onobrychis viciifolia, Lotus corniculatus, Rumex acetosa, Polygonum bistorta, Holcus lanatus, Leucanthemum vulgare, Daucus carota ssp. carota, Achillea millefolium, Silene vulgaris, Dianthus carthusianorum, Leontodon autumnalis etc.

252
Valoare conservativă: redusă Corologie: Etajele montan inferior şi mijlociu ale Carpaţilor din România (Sud-Estici). Starea actuală de conservare: - datorită valorii conservative scăzute nu credem că este necesară includerea acestui habitat în arii protejate noi, el fiind deja prezent în multe dintre ariile protejate din zona montană. 5.3.3.2. Habitatul R3802 - Pajişti daco-getice de Arrhenatherum elatius NATURA 2000:6510 Lowland hay meadows (Alopecurus pratensis, Sanguisorba officinalis) EMERALD: 38.2 Lawlands humid grasslands CORINE: - PAL.HAB: 38.233 Carpathian submontane hay meadows EUNIS: E2.233 Carpathian submontane hay meadows Asociaţii vegetale: Arrhenatherum elatioris Br,-Bl. ex Scherrer 1925. Scurtă descriere: Flora pajiştilor de ovăscior este variată şi bogată, etajul superior fiind format din: Arrhenatherum elatius, Dactylis glomerata, Festuca pratensis, Salvia nemorosa, Onobrychis viciifolia, Trifolium pratense, Alopecurus pratense. Înălţimea acestui etaj ajunge la 80-100 cm şi realizează o acoperire mare (peste 70%). Cel de al doilea etaj, este format din plante de talie mai mică, dintre care cele mai reprezentative sunt: Medicago lupulina, Agrostis stolonifera, Trifolium repens, Lotus corniculatus, Campanula patula ssp. patula. În pajiştile din sud-vestul ţării (Oltenia, Banat) apare destul de frecvent specia Moenchia mantica, plantă mai puţin răspândită în flora ţării. Staţiuni: Altitudine: 350-700 m. Clima: T = 9-6,50C; P = 600-800 mm. Relief: versanţi slab şi mediu înclinaţi, expoziţie sudică şi sud-estică. Roci: şisturi cristaline şi mai rar calcare şi gresii. Soluri: brune luvice, moderate în humus, brun-montane slab acid, fertile. Specii dominante şi caracteristice: Arrhenatherum elatioris, Dactylis glomerata, Lotus corniculatus. Cerastium holosteoides, Trisetum flavescens, Trifolium pratense, Onobrychis viciifolia, Agrostis stolonifera, Dactylis glomerata, Alopecurus pratense, Leontodon autumnalis, Lolium perenne, Campanula patula ssp. patula, Phleum pratense.

253
Valoare conservativă: moderată. Corologie: Etajele colinar şi montan inferior din toată ţara. Starea actuală de conservare: - datorită valorii conservative scăzute şi a răspândirii largi pe care o are, el fiind deja prezent în multe dintre ariile protejate din zona montană, nu considerăm că este necesară includerea acestui habitat în arii protejate noi. 5.3.3.3. Habitatul R3803 – Pajişti sud-est carpatice de Agrostis capillaris şi Festuca rubra NATURA 2000: 6520 Mountain hay meadows EMERALD: 37.2 Eutrophic humide grasslands CORINE: 35.12 Agrostis – Festuca grasslands PAL.HAB: 38. 2323 Eastern Carpathian yellow oatgrass meadows EUNIS: E1.721 Nemoral Agrostis -Festuca grasslands Asociaţii vegetale: Festuco rubrae - Agrostetum capillaris Horvat 1951. Scurtă descriere: Acest tip de habitat cuprinde fâneţe montane, bogate în specii, cu o mare amplitudine ecologică. Sunt cele mai răspândite tipuri de pajişti, fiind prezente în tot lanţul carpatic şi ocupă cele mai mari suprafeţe. Sunt utilizate atât ca fâneţe cât şi ca păşuni. Condiţii staţionale: Altitudine: 350-700 m. Clima:T = 8-6,50C; P = 700-800 mm. Relief:versanţi slab înclinaţi, expoziţii sudice, estice. Substrat: roci silicioase, pietrişuri. Soluri: districambosoluri, luvosoluri. Specii dominante şi caracteristice: Agrostis capillaris, Festuca rubra, Trisetum flavescens, Anthyllis vulneraria, Hypochoeris radicata, Cynosurus cristatus, Briza media, Stellaria graminea, Anthoxanthum odoratum, Centaurea phrygia s.l., Linum catharticum, Leontodon hispidus, Trifolium aureum, Peucedanum oreoselinum, Holcus lanatus, Colchicum autumnale, Trifolium montanum, Trifolium pannonicum, Chamaespartium sagittale. Valoare conservativă: are o valoare conservativă moderată, aceasta fiind dată de prezenţa în structura habitatului a unor specii care se regaăsesc pe listele roşii naţionale şi europene, cum ar fi: Gymnadenia conopsea, Orchis maculata. O. morio, Traunsteinera globosa, Anthemis macrantha etc. Corologie: Etajul montan inferior şi mijlociu din întreg arcul carpatic.

254
Starea actuală de protecţie: - datorită valorii conservative scăzute nu credem că este necesară includerea acestui habitat în arii protejate noi, el fiind deja prezent în toate ariile protejate din zona montană. 5.3.3.4. Habitatul R3804 – Pajişti daco-getice de Agrostis capillaris şi Anthoxanthum odoratum NATURA 2000: - EMERALD: 37.2 Eutrophic humide grasslands CORINE: 35.12 Agrostis - Festuca grasslands PAL.HAB: 38.233 Carpathian submontane hay meadows EUNIS: E1.721 Nemoral Agrostis - Festuca grasslands Asociaţii vegetale: Anthoxantho -Agrostetum capillare Silinger 1933. Scurtă descriere: Sunt pajişti foarte asemănătoare din punct de vedere structural cu precedentele fiind edificate de Agrostis capillaris, Anthoxanthum odoratum şi Cynosurus cristatus. Specii dominante şi caracteristice: Agrostis capillaris, Anthoxanthum odoratum, Cynosurus cristatus, Lolium perenne, Luzula campestris, Knautia arvensis, Galium mollugo, Pastinaca sativa, Achillea millefolium, Festuca pratensis, Stellaria graminea, Filipendula vulgaris, Briza media, Linum catharticum, Trifolium pratense. Staţiuni: Altitudine: 300-700 m. Clima: T = 9,0-70C; P = 650-750 mm. Relief: pante moderat înclinate, platouri. Substrat: slab-moderat acid. Soluri: eutricambosoluri, districambosoluri, aluviosoluri. Valoare conservativă: ca şi în cazul precedentului tip de habitat, valoarea conservativă este redusă, ea fiind dată mai ales de către speciile de pe listele roşii naţionale care se regăsesc în structura habitatului. Corologie: etajul montan inferior din Carpaţii Orientali şi Apuseni.. Starea actuală de protecţie: - deşi doar o mică parte din masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în situri din cadrul Reţelei Natura 2000 (Fig.5.17), datorită valorii conservative scăzute nu credem că necesită declararea de noi situri Natura 2000 pentru conservarea acestui habitat.

255
Fig. 5.17. Harta de distribuţie a habitatului R3804 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3804 into Natura 2000 Network

256
5.3.4. Pajişti xerice 5.3.4.1. Habitatul R3401 - Pajişti sud-est carpatice de Asperula capitata şi Sesleria rigida NATURA 2000: 6170 Alpine and subalpine calcareous grasslands EMERALD: - CORINE: 36.4 Alpine and subalpine calci-philous grasslands PAL.HAB 1999: 36.43921 East Carpathian Sesleria-evergreen sedge grasslands EUNIS: E4.4392 East Carpathian calciphile stepped grasslands Asociaţii vegetale: Asperulo capitatae -Seslerietum rigidae (Zólyomi 1939) Coldea 1991 (Syn.: Seslerietum rigidae praebiharicum Zólyomi 1939, Asplenio -Seslerietum rigidae Csűrös 1958, Alysso - Seslerietum rigidae Csűrös et al. 1958). Scurtă descriere: Cenozele edificate de Sesleria rigida ssp. rigida şi Asperula capi-tata, sunt foarte răspândite, regăsindu-se pe curprinsul majorităţăţii masivelor calcaroase din ţară. Pajiştile de Sesleria rigida se întâlnesc pe vesanţi stâncoşi cu expoziţii variabile şi pante medii până la mari. Staţiuni: Altitudine: 400-1550 m. Clima: T = 7,8-30C; P = 800-1200 mm. Relief: se întâlnesc frecvent pe rocile calcaroase umbroase şi semi-umbroase din etajul montan. Soluri: rendzine bogate în schelet şi cu reacţie bazică (pH = 7,3-8). Compoziţie floristică: Sesleria rigida, Asperula capitata, Alyssum repens, Saponaria bellidifolia, Centaurea triumfetii, Anthericum ramosum, Pedicularis comosa ssp. campestris, Helictotrichon decorum, Festuca xanthina, Saxifraga marginata, Bupleurum falcatum, Centaurea atropurpurea, Stipa eriocaulis, Dianthus petraeus, Edrainthus graminifolius, Conioselinum tataricum, Cnidium silaifolium, Seseli gracile, Seseli rigidum, Primula columnae, Aconitum anthora, Dianthus spiculifolius, Thymus comosus, Viola jooi. Valoare conservativă: - mare determinată de caracterul endemic al habitatului şi de prezenţa unor specii endemice şi rare, dintre care amintim Seseli rigidum, Dianthus spiculifolius, Centaurea atropurpurea, Conioselinum tataricum, Saponaria bellidifolia, Helictotrichon derorum, Saxifraga marginata, Thymus comosus, Viola jooi etc. Corologie: Muntele Postăvaru, Măgura Codlei, Munţii Leaota, Cheile Rudăriţei, Cheile Dâmboviţei, Munţii Bucegi, Munţii Piatra Craiului, Munţii Lotru, Valea Călineşti, Munţii Ţarcu-Godeanu, Munţii Apuseni, Munţii Trascău, Cheile Ordâncuşii, Cheile Crăciuneşti, Cheile Bulzeşti, Cheile Feneşului, Cheile Turului, Gilău, Metaliferi, Munţii Bihorului, Munţii Scăriţa-Belioara, Cheile Runcu.

257
Starea actuală de conservare: majoritatea zonelor în care se întâlneşte acest habitat sunt incluse în parcuri naţionale şi situri Natura 2000 (Fig. 5.18) nefiind necesare măsuri suplimentare.
Fig. 5.18. Harta de distribuţie a habitatului R3401 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3401 into Natura 2000 Network

258
5.3.4.2. Habitatul R3402 – Pajişti sud-est carpatice de Helictotrichon decorum NATURA 2000: 6170 Alpine and subalpine calcareous grasslands EMERALD: - CORINE: 34 Dry calcareous grasslands and steppes PAL.HAB 1999: 34.3512 East Carpathian Sesleria rigida grasslands; 34.35122 Apuseni Helictotrichon grasslands EUNIS: - Asociaţii vegetale: Helictotrichetum decori Domin 1932. Scurtă descriere: Pajiştile edificate de Helictotrichum decorum sunt întâlnite în zone restrânse din Carpaţii Orientali, iar în Carpaţii Occidentali mai ales la periferia acestora lipsind din parte lor centrală. Fitocenozele acestui habitat vegetează mai ales pe versanţii însoriţi a căror cu pante variabile, de la mici până la foarte mari. Din punct de vedere fitosociologic, în cadrul asociaţiei Helictotrichetum decori Domin 1932 au fost descrise următoarele subasociaţii: caricetosum humilis Csűrős 1958; seslerietosum rigidae Csuros 1958; melicetosum flavescentis Zolyomi 1939. Staţiuni: Altitudine: 500-1350 m. Clima: T = 7,3-3,50C; P = 800-1150 mm. Relief: stânci, versanţi însoriţi. Substrat: calcaros. Soluri: rendzine superficial, cu o reacţie bazică (pH = 7-8). Specii dominante şi caracteristice: Helictotrichon decorum, Asperula capitata, Alyssum repens, Anthericum ramosum, Pedicularis comosa ssp. campestris, Saponaria bellidifolia, Sesleria rigida, Seseli gracile, Seseli rigidum, Primula veris ssp. columane, Aconitum anthora, Centaurea atropurpurea, Stipa eriocaulis, Erysimum witmannii, Centaurea reichenbachii, Phyteuma orbiculare, Aster alpinus, Ranunculus oreophilus, Carduus glaucus, Acinos alpinus, Minuartia verna, Biscutella laevigata, Myosotis alpestris, Centaurea pinnatifida, Dianthus spiculifolius, Thymus comosus, Viola jooi. Valoare conservativă: - mare determinată atât de caracterul endemic al habitatului cât şi de prezenţa în structura acestuia a unui număr mare de specii endemice şi rare, cum ar fi: Helictotrichon decorum, Saponaria bellidifolia, Seseli gracile, Seseli rigidum, Centaurea atropurpurea, Stipa eriocaulis, Erysimum witmannii, Centaurea reichenbachii, Dianthus spiculifolius, Thymus comosus, Viola jooi, Cephalaria radiata, Allium obliquum etc. Corologie: Suhardul Mic lângă Lacul Roşu, Muntele Postăvaru, Muntele Scăriţa-Belioara, Cheile Întregalde, Piatra Caprei, Cheile Runcu, Cheile Feneş, Cheile Râmeţ, Cheile Turzii, Muntele Trascău.
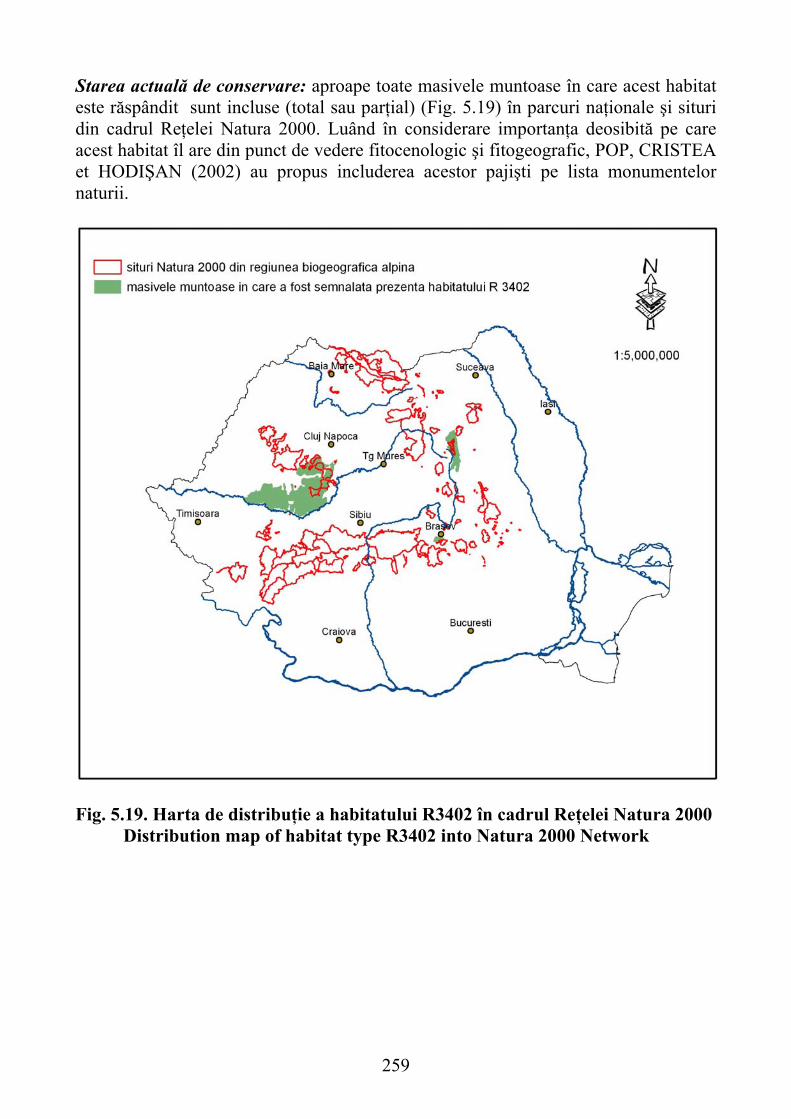
259
Starea actuală de conservare: aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) (Fig. 5.19) în parcuri naţionale şi situri din cadrul Reţelei Natura 2000. Luând în considerare importanţa deosibită pe care acest habitat îl are din punct de vedere fitocenologic şi fitogeografic, POP, CRISTEA et HODIŞAN (2002) au propus includerea acestor pajişti pe lista monumentelor naturii.
Fig. 5.19. Harta de distribuţie a habitatului R3402 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3402 into Natura 2000 Network

260
5.3.4.3. Habitatul R3403 Pajişti daco-getice de Festuca pallens şi Melica ciliata NATURA 2000: 6190 Rupicolous pannonic grasslands EMERALD: 34.3 Dense perennial grasslands and middle European steppes CORINE: 34.35 Pale fescue (Festuca pallens) grasslands PAL.HAB: 34.35223 Pre-Dacic pale fescue grasslands EUNIS: E1.1133 Melica ciliata rock debris swards Asociaţii vegetale: Asplenio rutae-murariae - Melicetum ciliatae Soó 1962, Seseli gracile - Festucetum pallentis (Soó 1959) Coldea 1991, Melico -Phleetum montani Boşcaiu 1966. Scurtă descriere: În cadrul habitatului se regăsesc pajişti xerofile stepizate din etajul montan inferior. Fitocenozele care alcătuiesc acest tip de habitat preferă staţiunile uscate, pe soluri calcaroase cu diferite expoziţii şi grade de înclinare. Staţiuni: Altitudine: 500-800 m. Clima: T = 7,3-60C; P = 750-900 mm. Relief: versanţi moderat înclinaţi până la puternic înclinaţi, cu expoziţie sudică, sud-estică, sud-vestică. Roci: calcare, bolovănişuri calcaroase. Soluri: rendzine superficiale, bogate în humus. Specii dominante şi caracteristice: Festuca pallens, Melica ciliata, Phleum montanum, Stipa joannis, Seseli gracile, Ferula sadleriana, Helianthemum nummularium, Cardaminopsis arenosa, Stachys recta, Jurinea mollis, Linum tenuifolium, Artemisia campestris, Veronica austriaca ssp. jacquinii, Peucedanum oreoselinum, Vincetoxicum hirundinaria, Poa pannonica, Genista januensis, Festuca rupicola, Asplenium trichomanes, A. septentrionale, A. ruta-muraria, Cystopteris fragilis, Sedum hispanicum, Acinos arvensis, Veronica bachofenii, Thymus comosus etc. Valoare conservativă: moderată ca urmare a prezenţei unor specii endemice şi rare cum ar fi: Ferula sadleriana, Seseli gracile, Genista januensis, Poa pannonica, Jurinea mollis, Thymus comosus, Veronica bachofenii etc. Corologie: Munţii Hăşmaş, Masivul Postăvaru, Masivul Leaota, Munţii Cibinului, Munţii Trascăului, Munţii Gilău, Someşul Rece, Munţii Plopiş. Starea actuală de conservare: aproape toate masivele muntoase în care acest habitat este răspândit sunt incluse (total sau parţial) în parcuri naţionale şi situri din cadrul Reţelei Natura 2000 (Fig. 5.20).
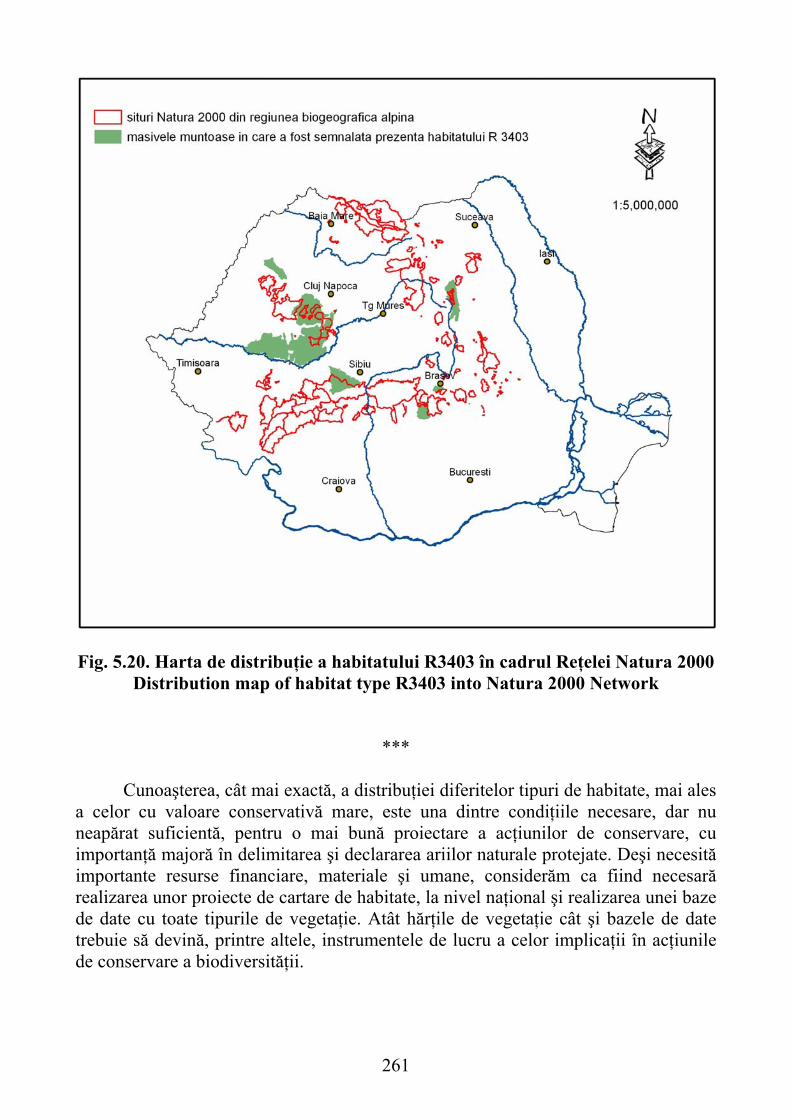
261
Fig. 5.20. Harta de distribuţie a habitatului R3403 în cadrul Reţelei Natura 2000
Distribution map of habitat type R3403 into Natura 2000 Network
***
Cunoaşterea, cât mai exactă, a distribuţiei diferitelor tipuri de habitate, mai ales a celor cu valoare conservativă mare, este una dintre condiţiile necesare, dar nu neapărat suficientă, pentru o mai bună proiectare a acţiunilor de conservare, cu importanţă majoră în delimitarea şi declararea ariilor naturale protejate. Deşi necesită importante resurse financiare, materiale şi umane, considerăm ca fiind necesară realizarea unor proiecte de cartare de habitate, la nivel naţional şi realizarea unei baze de date cu toate tipurile de vegetaţie. Atât hărţile de vegetaţie cât şi bazele de date trebuie să devină, printre altele, instrumentele de lucru a celor implicaţii în acţiunile de conservare a biodiversităţii.

262
VI GOSPODĂRIREA HABITATELOR DE PAJISTI 6.1. GOSPODĂRIREA HABITATELOR CU VALOARE FURAJERĂ ŞI
CONSERVATIVĂ
Modul de folosinţă al pajiştilor (managementul pajiştilor) are implicaţii la nivelul compoziţiei floristice, productivităţii şi calitatea furajului obţinut. În cadrul a numeroase studii realizate pe pajişti, managementul pajiştilor este de multe ori ignorat şi tocmai acesta este cel care a determinat situaţia actuală. Managementul este cel care creează şi menţine o fitodiversitate ridicată, dar tot acesta poate să o reducă drastic într-un timp scurt. Implicaţiile modului de folosinţă merg şi mai departe până la nivelul esteticii landşaftului cultural. Neaplicarea managementului pe anumite pajişti ar determina dispariţia a numeroase elemente de landşaft (grămezi de pietre, garduri de lemn, copaci singuratici etc.), care ar deprecia considerabil particularitatea landşaftului local. Deprecierea landşaftului ar atrage după sine scăderea considerabilă a potenţialului turistic al unei zone (În Alpii francezi şi Pirinei, 65% din păşunile de vară sunt întretăiate de cărări care servesc agroturismului şi ecoturismului, cu adăposturi construite pentru turişti, iar 15% servesc activităţilor de schiat). Absenţa managementului (abandonul pajiştilor) determinat de depopularea zonelor montane, are de asemenea efecte negative asupra fitodiversităţii şi implicit a landşaftului. Suprafeţele respective se vor reîmpăduri şi landşaftul cultural va deveni probabil o „pădure”. Situaţii similare s-au întâmplat şi în ţări ale Comunităţii Europene, spre exemplu în Elveţia în perioada 1985 – 1995, 4% din suprafaţa de pajişti a ţării a fost reîmpădurită (JEANGROS şi THOMET, 2004). Abandonarea suprafeţelor de pajişti din Italia a determinat reîmpădurirea suprafeţelor si, implicit, reducerea biodiversităţii floristice (LOMBARDI, 2001, SABATINI şi colab., 2003, SUSAN şi ZILIOTTO, 2004). Acelaşi lucru este semnalat de BIALA şi ZYSZKOWSKA (2004) în Polonia, HEJCMAN şi colaboratorii (2004) în Cehia.
Nu cu mult timp în urmă, atât la nivel European, cât şi în ţara noastră, obiectivele principale ale managementului pajiştilor erau de a intensiviza sistemele de pajişti în vederea îmbunătăţirii troficităţii solului şi sporirii producţiei de iarbă la unitatea de suprafaţă. Această intensivizare presupunea aplicarea de îngrăşăminte minerale şi organice în cantităţi mari (200 – 300 kg / ha N s.a.), care de multe ori erau combinate cu lucrări de supraînsămânţare. Această situaţie creată a determinat modificări pregnante la nivelul covorului ierbos, înregistrându-se o instalare puternică a speciilor nitrofile în detrimentul speciilor azotofuge şi moderat nitrofile. La momentul actual, când se pune problema păstrării fitodiversităţii şi menţinerea unor habitate importante pentru reţeaua Natura 2000 şi alte programe de conservare, se impun noi abordări ale managementului pajiştilor, care să aibă obiective chiar contradictorii cu cele ale sistemelor intensive.
Din cele expuse mai sus putem afirma că pajiştile sunt strâns legate de modul de folosinţă şi foarte uşor ele pot fi „dirijate” de la o extremă la alta (abandon – exploatare intensivă). De aceea este deosebit de importantă elaborarea unor „reţete” de moduri de folosinţă care să menţină pajiştile naturale primare şi secundare în cadrul landşaftului cultural.

263
Scopul lucrării noastre este de a elabora un sistem de management de menţinere a habitatelor de pajişti şi a fitodiversităţii lor în cadrul landşaftului cultural în conformitate cu cerinţele programului Natura 2000
6.1.1. Pajişti alpine şi subalpine Etajul alpin La noi in tara etajul alpin ocupa terenuri la altitudini de peste 2000 m in nord si
2100-2200 m in sud. Solurile humicosilicatice, rendzine, litosoluri, s-au format pe şisturi cristaline
în Munţii Rodnei, Făgăraş, Parang-Cindrel, conglomerate si calcare in Bucegi, Ţarcu-Godeanu, graniţe in Retezat etc.
Temperatura medie anuala (Tm) este de -1,50C pana la 2,50C, iar precipitaţiile medii anuale (Pm) sunt cuprinse intre 1300 si 1400 mm. Suprafaţa ocupata cu pajişti este de aproximativ 40.000 ha.
Tipul Carex curvula Pe lângă specia dominanta mai întâlnim si alte specii: Agrostis rupestris,
Juncus trifidus, Poa media, Campanula alpina, Hieracium alpinum, Chrysanthemum alpinum, Primula minima, Soldanella pusilla.
Ocupa platouri, coame, versanţi· slab înclinaţi cu soluri puternic acide (humicosilicatice, litice turboase), fiind cel mai important tip de pajişte din zona alpina.
Tipul Juncus trifidus -Agrostis rupestris Alte specii: Festuca airoides, Poa media, Anthemis carpatica, Campanula
alpina, Hieracium alpinum, Soldanella pusilla. Ocupa culmi înguste şi versanţi foarte puternic înclinaţi, abrupţi, expuşi
vanturilor, cu soluri revene, foarte putemic acide. Pajiştile de Carex curvula, Juncus trifidus şi Agrostis rupestris dau producţii
foarte mici de 1,5-3 t/ha masa verde si o capacitate de păşunat de 0,1 – 0,3 UVM/ha.
Etajul subalpin Etajul subalpin ocupa teritorii in munţii înalţi, la altitudini cuprinse intre 1700-
2200 m. Solurile sunt podzoluri, brune feriiluviale, rendzine, litosoluri, formate pe şisturi cristaline, graniţe, calcare.
Temperatura medie anuala (Tm) este scăzuta, fiind cuprinsa Intre 0,5si 1,50C, iar cantitatea de precipitaţii mare: Pm=1200 mm.
Suprafaţa ocupata de pajişti este de aproximativ 66.000 ha.

264
Tipul Festuca airoides Alături de specia principala mai întâlnim: Agrostis rupestris, Poa violacea,
Nardus stricta, Geum montanum, Polygonum viviparum, Potentilla ternata, Pulsatila alba. Acest tip ocupa versanţi de la moderat la foarte puternic înclinaţi, însoriţi, soluri revene-uscat revene, extrem oligobazice, foarte puternic acide.
Tipul Festuca nigrescens - Nardus stricta Pe lângă cele doua specii care domina pajiştea mai întâlnim: Phleum alpinum,
Genista sagittalis, Trifolium repens, Centaurea nervosa, Hieracium aurantiacum, Scorzonera rosea, Viola declinata. Ocupa versanţi de la slab pana la moderat înclinat, însoriţi, soluri revene, puternic acide, in zone cu ericacee, cu humus brut turbos.
Tipul Nardus stricta - Potentilla aurea ssp. chrysocraspeda Alte specii ale releveului floristic: Poa media, Genista tinctoria, Gentiana
punctata, Luzula sudetica, Potentilla aurea ssp. chrysocraspeda. Ocupa versanţi slab pana la moderat înclinaţi, soluri revene-jilave, cu umiditate
excesiva primăvara datorita topirii rapide a stratului de zăpada, având humus brut turbos.
Tipul Festuca amethystina - Festuca versicolor Alte specii: Poa violacea, Koeleria macrantha, Onobrychis montana, Allium
ochroleucum, Iris ruthenica, Knautia longifolia, Thlaspi dacicum. Acest tip de pajişte se dezvolta pe versanţi abrupţi din munţi calcaroşi, pe
soluri afânate, slab acide-neutre. Are mai mica importanţă furajeră şi mai mult botanică.
Acestei serii îi mai aparţin două tipuri cu importanţă praticolă redusă: Sesleria haynaldiana - Carex sempervirens care ocupă versanţi calcaroşi, si Poa violacea pe versanţi însoriţi, abrupţi.
Pajiştile de Festuca airoides (păruşcă) sunt slabe din punct de vedere productiv, 2-4 t/ha masă verde şi o capacitate de păşunat de 0,2-0,5 UVM/ha.
Aşa cum am mai remarcat, pajiştile de Nardus stricta sunt slab productive, 3-5 t/ha masa verde, de calitate slabă, şi o încărcătură de 0,4-0,5 UVM/ha.
Cele aparţinând seriei Festuca amethystina prezintă mai multă importanţă din punct de vedere botanic şi mai puţin furajer.
6.1.1.1. Lucrări de întreţinere Curăţirea pajiştilor constă în strângerea pietrelor de pe pajişti. Această lucrare
se justifică numai acolo unde s-a practicat timp îndelungat (nu se recomandă ca o lucrare nouă pentru regiunile unde nu s-a mai practicat) deoarece, probabil, a avut o contribuţie la aspectul floristic actual şi la crearea unor elemente de landşaft (grămădarele de pietre).
Recomandăm curăţirea pajiştilor de orice alte „materiale” cum ar fi: pungile de plastic, peturile, pungile de hârtie, ş.a. Distrugerea muşuroaielor, mai ales a celor de origine vegetală, se recomandă atunci când au depăşit 40 % din suprafaţa unei pajişti, dar numai cu acordul şi sub supravegherea organelor de stat competente.

265
Combaterea buruienilor. Buruienile ce apar în etajul alpin şi subalpin sunt: Aconitum anthora, Aconicum tauricum, Aconicum toxicum, Colchicum autumnale, Euphorbia cyparissias, Hypericum maculatum, Ranunculus nemorosus, Stellaria graminea, Veratrum album etc. Dintre acestea , o răspândire mai largă şi un pericol mai mare pentru animale o au speciile Veratum album şi Aconitum sp. De asemenea, sunt foarte răspândite speciile nitrofile, ca: Urtica dioica, Rumex alpinus şi Rumex obtusifolius. Există şi alte multe specii de buruieni şi recomandăm combaterea lor manuală (exclus folosirea erbicidelor sau a altor substanţe), dar cu o consultare prealabilă cu organele competente ale statului, pentru a evita distrugerea vreunei specii cu valoare conservativă. Îndepărtarea vegetaţiei lemnoase. În etajele alpin şi subalpin vegetaţia lemnoasă are o importanţă deosebită, cu o tendinţă de extindere, care, uneori poate ajunge la o acoperire foarte mare, diminuând astfel, suprafaţa pajiştii. Recomandăm combaterea manuală a speciilor ca Vaccinium myrtillus şi Vaccinium vitis-idaea în condiţiile unui plan riguros şi prezentat organelor competente. În planurile de defrişare, indiferent de condiţiile în care se găsesc, nu intră speciile periclitate sau declarate Monumente ale Naturii, cum ar fi: Pinus cembra, Pinus sylvestris, Larix decidua, Rhodondendron myrtifolium, Juniperus sibirica, Bruckenthalia spiculifolia, Loiseleuria procumbens etc. Fertilizarea pajiştilor alpine şi subalpine nu se recomandă şi considerăm că sunt suficiente cantităţile de gunoi ce rămân în urma păşunatului. Recomandăm fertilizarea doar pe pajiştile cu un grad de eroziune ridicat, unde acoperirea covorului ierbos este mică (sub 60%). Fertilizarea se poate realiza fie prin târlitul cu oile (1-2 nopţi), fie prin aplicarea a 5-10 t / ha gunoi de grajd o dată la 3-4 ani.
6.1.1.2. Sistemul de exploatare Pajiştile alpine şi subalpine se vor exploata prin păşunat într-un sistem liber
extensiv. Recomandăm păşunatul cu ovinele , liber extensiv îmbunătăţit care presupune împărţirea pajiştilor în 2-3 parcele, iar încărcătura de animale se stabileşte în funcţie de încărcătura ei.
Păşunatul trebuie să înceapă când vegetaţia are 10 cm înălţime şi să se încheie cu 25-30 zile înainte de venirea primelor îngheţuri, astfel încât plantele să aibă timp să-şi acumuleze substanţele de rezervă necesare. Încărcătura de animale la hectar pentru pajiştile alpine şi subalpine o recomandăm între 0,2-0,4 UVM / ha.
6.1.2. Pajişti umede Pajiştile umede sunt suprafeţe ierboase situate pe soluri umede sau umezite în
permanenţă, în luncile râurilor, în văile şi depresiunile dintre dealuri sau din câmpie, la baza pantelor şi pe terenurile cu apa freatică la mică adâncime. Aceste pajişti, în ţara noastră, ocupă o suprafaţă de circa 550.000 hectare, reprezentând 9,2 % din suprafaţa totală de pajişti permanente.
Condiţiile naturale specifice din aceste zone au favorizat instalarea unor pajişti valoroase, cu producţii mari şi de bună calitate furajeră, cu o compoziţie floristică

266
variată, formată din specii mezofile, mezohigrofile şi, pe terenurile cu exces permanent de umiditate, din specii higrofile sau hidrofile.
Suprafeţele cele mai mari de pajişti umede se întâlnesc în luncile şi văile râurilor, unde predomină solurile aluvionare, permeabile, cu textură uşoară (luto-nisipoasă), bine aprovizionate în substanţe nutritive, ca urmare a inundaţiilor periodice prin care se transportă şi se depozitează cantităţi mari de minerale, determinând fertilitatea ridicată a acestor terenuri.
Structura floristică a pajiştilor instalate în luncile râurilor este foarte diversificată, aceasta în funcţie şi de distanţa acestor suprafeţe faţă de albia majoră. Astfel, pe suprafeţele din imediata apropiere a albiei, care reprezintă şi fâşia cea mai înaltă a râului şi cea mai redusă ca suprafaţă, în urma inundaţiilor, se depun nisipuri grosiere, cu aport scăzut în materie organică. Pe aceste terenuri, unde solurile au o capacitate redusă, iar după retragerea apelor de inundaţie scade puternic şi nivelul apei freatice, plantele instalate pot suferi din lipsa apei în cea mai mare parte a verii. În aceste condiţii variabile vegetaţia pajiştilor este formată, în cea mai mare parte, din specii xerofile şi mezoxerofile, caracteristice terenurilor neinundate.
După această fâşie îngustă de teren urmează suprafaţa propriu-zisă ,cea mai mare a albiei râurilor, pe care, în urma inundaţiilor, se depun materiale fine, bogate în minerale, imprimând solului o textură mijlocie luto-nisipoasă, cu o capacitate mai mare de reţinere a apei. Pe aceste terenuri, în urma retragerii apelor, se instalează pajişti valoroase, formate din asociaţii de specii mezofile şi mezohigrofile, unde prezenţa în proporţie mai mare a speciilor de poacee şi fabacee ridică calitatea acestor pajişti.
La periferia albiei majore a râurilor, în porţiunea cea mai îndepărtată, aflată sub nivelul apei, unde excesul de umiditate este aproape în permanenţă, se formează asociaţii vegetale din specii higrofile şi hidrofile, în care predomină speciile de ciperacee, juncacee şi multe alte specii cu valoare furajeră scăzută.
Structura floristică a pajiştilor situate la baza pantelor, în zonele neinundabile, unde starea de umiditate mai mare a solului este dată de apele diluviale (provenite din precipitaţii), care se scurg pe pantele versanţilor, este formată din specii cu valoare furajeră mare, adaptate la soluri cu umiditate şi fertilitate ridicată.
Asociaţiile vegetale de pajişti întâlnite în zonele de lunci şi depresiuni nu sunt specifice numai acestora, acestea se pot forma şi pe terenurile mai înalte, mai ales în zona podişurilor, unde condiţiile de umiditate şi fertilitate a solurilor sunt ridicate.
Tipurile de pajişti formate în aceste condiţii au o structură floristică foarte diversă, în care predomină speciile de poaceae mezofile şi higrofile. Aceste specii s-au asociat în formaţii de pajişti cu alte specii, din care mai importante, ca valoare economică şi furajeră, sunt pajiştile în care proporţia cea mai mare o au următoarele speciile: pajişti de Lolium perenne, de Agropyron repens, de Alopecurus pratensis, de Agrostis stolonifera, de Poa pratensis, de Cynodon dactylon.
Pajiştile formate din specii higrofile şi hidrofile sunt adaptate la condiţii de umiditate în exces, pe tot timpul anului, în care predomină speciile de rogozuri şi pipirig, iar pe terenurile cu apă în permanenţă (bălţi şi mlaştini) speciile de stuf şi papură.

267
Pajiştile de zâzanie (Lolium perenne) sunt răspândite în luncile râurilor, pe terenurile plane sau cu înclinaţie mică, pe soluri fertile şi cu umiditate suficientă. Specia dominantă din această asociaţie este însoţită de numeroase alte specii mezofile sau mezoxerofile, cum sunt:
Poacee: Agrostis stolonifera, Poa pratensis, Cynodon dactylon, Alopecurus pratensis
Fabacee: Trifolium repens, Trifolium pratense, Trifolium fragiferum, Lotus corniculatus, Medicago lupulina etc.
Ciperacee şi juncacee: Carex hirta, Carex vulpina, Juncus gerardii etc. Specii diverse: Taraxacum officinale, Artemisia austriaca, Plantago sp.,
Achillea millefolium, Convolvulus arvensis, Potentilla reptans, Cichorium intybus, Matricaria inodora, Echium vulgare, Lepidium draba, Carduus nutans.
În structura compoziţiei floristice poaceele au o participare de 40-50 %, fabaceele între 5-10 %, iar speciile din alte familii botanice, mai ales pe terenurile ruderalizate, ajung până la 50 %.
Prezenţa în asociaţie a unor specii valoroase de fabacee şi poacee asigură realizarea unor producţii bune de masă verde şi o calitate ridicată a furajului.
Pajiştile de pir târâtor (Agropyron repens), se întâlnesc în toate luncile inundabile ale râurilor, din zonele de câmpie şi de deal, pe solurile aluvionale, permeabile, reavăne.
În structura floristică a acestor pajişti, specia dominantă Agropyron repens ajunge la peste 60% din compoziţia floristică.
Răspândirea largă a acestor pajişti determină şi o structură foarte diversă a speciilor însoţitoare, cu care, în anumite zone, formează asociaţii mixte (de exemplu Agropyron repens + Gallega officinalis).
Speciile mai importante care intră în asociaţie cu Agropyron repens sunt următoarele: Lolium perenne, Alopecurus pratensis, Cynodon dactylon, Poa pratensis, Lotus corniculatus, Lotus tenuis, Trifolium repens, Triofolium hybridum, Medicago lupulina, Plantago lanceolata, Inula britanica, Lepidium ruderale, Polygonum aviculare, Centaurea iberica, etc.
Caracterul stolonifer al plantelor de Agropyron repens duce la formarea unor pajişti cu un grad mai scăzut de acoperire a solului cu vegetaţie, determinând o valorificare a acestora, mai mult, prin cosire pentru producerea fânului.
Pajiştile de moleaţă (Agrostis stolonifera) ocupă suprafeţe mari pe terenuri mai joase din luncile râurilor, frecvent inundate şi preferă solurile grele, luto-argiloase, lăcoviştite, cu apa freatică superficială.
Specia dominantă, Agrostis stolonifera , poate realiza, uneori, o acoperire de 80-100 % a covorului vegetal, determinând o calitate slabă a furajului obţinut de pe aceste pajişti.
În pajiştile asociate compoziţia floristică este formată din specii mai valoroase, cum sunt: Lolium perenne, Poa pratensis, Poa trivialis, Cynosurus cristatus, Holcus lanatus, Agropyron repens, Trifolium repens, Trifolium hybridum, Trifolium fragiferum, Trifolium campestre, Lotus corniculatus, Carex hirta, Scirpus maritimus,

268
Plantago major, Potentilla reptans, Symphytum officinale, Rumex crispus, Lychnis flos-cuculi etc..
Pajiştile de Agrostis stolonifera deşi realizează producţii ridicate, calitatea furajului este scăzută din cauza gradului redus de consumabilitate al speciei dominante, care creşte sau scade în funcţie de ponderea de participare a speciilor de fabacee sau a celor din alte familii botanice.
Condiţiile staţionale şi însuşirile morfologice ale speciei Agrostis stolonifera influenţează şi modul de valorificare al acestor pajişti, în sensul folosirii lor ca fâneaţă.
Pajiştile de firuţă (Poa pratensis) au o largă răspândire în zonele depresionare dintre dealuri şi se instalează pe coastele domoale ale acestora şi pe platouri. Această asociaţie de pajişte preferă solurile reavăne, profunde, fertile, din grupa solurilor aluvionare şi cernoziomice.
În zonele cu condiţiile ecologice cele mai favorabile se întâlnesc pajişti mixte, cu două specii de poacee edificatoare, respectiv: Poa pratensis + Festuca pratensis sau Poa pratensis + Lolium perenne.
Dintre speciile însoţitoare care formează compoziţia floristică a acestor pajişti, cele mai importante sunt: Agropyron repens, Agrostis stolonifera, Festuca pseudovina, Festuca sulcata, Poa bulbosa, Alopecurus pratensis, Festuca arundinacea, Trifolium repens, Trifolium pratense, Trifolium hybridum, Trifolium resupinatum, Medicago lupulina, Lotus corniculatus, Plantago sp, Achillea millefolium, Taraxacum officinale, Euphorbia cyparissias, Potentilla reptans, etc.
Structura floristică a acestor pajişti şi condiţiile staţionale de formare determină valorificarea acestora, în exclusivitate, prin păşunat, deoarece speciile componente rezistă bine la păşunat şi se regenerează repede.
Pajiştile de pir gros (Cynodon dactylon) Asociaţia care formează acest tip de pajişte se întâlneşte în zonele mai înalte
din luncile râurilor, unde predomină solurile nisipoase şi cu umiditate redusă. În general, aceste pajişti ocupă grindurile nisipoase din apropierea albiei râurilor.
Condiţiile ecologice speciale care conduc la formarea şi răspândirea acestor pajişti influenţează negativ asupra dezvoltării covorului vegetal, care, de regulă, se caracterizează printr-o acoperire scăzută a solului.
Cele mai frecvente specii însoţitoare, care participă la alcătuirea covorului vegetal al acestor pajişti, sunt următoarele: Medicago lupulina, Trifolium fragiferum, Capsella bursa – pastoris, Daucus carota, Euphorbia cyparissias, Eryngium campestre, etc.
Valoarea economică a acestor pajişti este redusă, cu producţii relativ mici şi de calitate slabă.
Pajiştile de rogozuri, pipirig, stuf şi papură (Carex sp., Juncus sp., Scirpus sp., Phragmites sp., Typha sp.).
Vegetaţia acestor pajişti este formată din specii higrofile şi hidrofile, adaptate terenurilor cu exces, aproape, permanent de umiditate în sol.
Zonele cu răspândirea cea mai largă a acestor terenuri se întâlnesc în luncile inundabile ale râurilor, în luncile şi bălţile Dunării şi în Deltă.

269
În funcţie de adâncimea apei de pe aceste terenuri se dezvoltă diverse asociaţii de specii higrofile, formate din: Phalaris arundinacea, Poa polustris, Agrostis canina, Beckmania eruciformis, Carex sp., Scirpus sp., Juncus sp. La adâncimi de peste 0,50 m se instalează o vegetaţie hidrofilă, specifică condiţiilor mediului acvatic.
Condiţiile staţionale specifice şi valoarea furajeră scăzută a speciilor din vegetaţia acestor pajişti impun măsuri speciale de protecţie şi conservare a habitatelor de pe aceste terenuri.
6.1.2.1. Lucrări de întreţinere Pajiştile umede sunt considerate ca cele mai importante din ţara noastră, nu
numai pentru valoarea lor economică şi furajeră, dar mai ales, pentru biodiversitatea lor, dată de bogăţia mare de specii botanice şi faunistice. Aceste pajişti formate într-un mediu deschis, în care apa provenită din precipitaţii sau din inundaţia râurilor au un rol foarte important, necesită intervenţia permanentă a omului, atât în privinţa valorificării cât şi a preservării şi conservării acestora.
Gestionarea pajiştilor umede, mai ales a celor aluviale, care ocupă un spaţiu de tranziţie permanentă între apă şi uscat, trebuie să asigure perenitatea acestora, în condiţiile în care asupra acestor terenuri acţionează o serie de factori perturbatori, cum sunt: intensivizarea practicilor agricole pentru valorificarea producţiei în furajarea animalelor, schimbarea destinaţiei acestor terenuri prin cultivarea de specii cerealiere şi „urbanizarea” terenurilor de-alungul râurilor şi văilor.
De asemenea, o valorificare raţională a acestor pajişti nu presupune şi alegerea variantei de abandon, deoarece, aceste terenuri ar evolua spre stadiul pre-forestier, prin proliferarea arbuştilor şi arborilor tineri.
Lucrările tehnologice de întreţinere, care vizează managementul acestor pajişti, trebuie să respecte bunele condiţii agricole şi de mediu (GAEC) şi să asigure păstrarea echilibrului dintre fitodiversitatea vegetaţiei şi menţinerea unor habitate specifice acestor terenuri.
Condiţiile naturale staţionale în care se întâlnesc pajiştile umede influenţează şi structura lucrărilor tehnologice ce se aplică pe aceste terenuri pentru întreţinerea şi folosirea lor. Astfel, starea de umiditate, aproape permanentă a solului, textura şi gradul de troficitate, structura floristică a covorului vegetal, determină aplicarea unui sistem diferenţiat de măsuri organizatorice şi tehnologice de întreţinere în vederea ameliorării şi preservării acestor pajişti.
Suprafeţele de pajişti situate în luncile şi văile râurilor vor fi curăţate după fiecare viitură a apelor de materialele aluvionare grosiere (pietriş, pietre) şi de unele materiale menajere. Această lucrare se aplică şi pe celelalte tipuri de pajişti umede, aflate la periferia albiei majore a râurilor, în urma unor inundaţii majore.
La începutul perioadei de vegetaţie (primăvara devreme), sau toamna târziu (după încheierea vegetaţiei) se vor efectua lucrări generale de curăţenie, printre care şi îndepărtarea resturilor vegetale, executate manual prin cosire.
Pe pajiştile cu exces de umiditate, formate din specii cu tufa deasă (Deschampsia caespitosa) sau specii de Juncus sp., prezenţa muşuroaielor, datorate acestor specii, duce la denivelarea terenului şi la diminuarea suprafeţei de folosinţă. În situaţia în care excesul de umiditate nu poate fi eliminat în totalitate şi pentru a se evita tasarea solului, se recomandă distrugerea muşuroaielor prin lucrări manuale. În

270
condiţiile în care starea de umiditate a solului permite se pot folosi, pentru distrugerea acestor muşuroaie, grape obişnuite (cu colţi reglabili), urmate de nivelarea terenului.
În structura floristică a pajiştilor umede unde gradul de îmburuienare este ridcat, mai ales în cazul terenurilor cu exces de umiditate şi apa freatică la mică adâncime, se vor efectua lucrări specialer de combatere a speciilor de buruieni. În acest sens, ţinând seama de condiţiile staţionale, se vor aplica numai măsuri preventive şi lucrări manuale sau mecanice. Măsurile preventive, prin care se limitează îmburuienarea acestor pajişti sau se evită apariţia unor specii de buruieni puternic invazive, sunt următoarele: cosirea resturilor vegetale neconsumate de animale, cosirea speciilor de buruieni de pe suprafeţele din apropierea pajiştilor luate în folosinţă, aplicarea gunoiului de grajd bine fermentat, împrăştierea dejecţiilor animaliere după păşunat, recoltarea fâneţelor înainte de fructificarea buruienilor (pe suprafeţe de pajişte unde proporţia acestora în covorul vegetal este de peste 50%), pentru a se evita autoînsămânţarea acestora.
În funcţie de frecvenţa, numărul, gradul de acoperire şi biologia speciilor de buruieni se recomandă lucrarea de cosire repetată, pe cale manuală sau mecanică, în condiţiile evitării tasării solului.
Lucrarea de grăpare se recomandă, în special, pe pajiştile de luncă, de o parte şi alta a râurilor, unde s-a depus un strat de nisip sau de mâl în urma inundaţiilor şi pe pajiştile umede, în care dominante sunt speciile stolonifere (pajiştile de Agrostis stolonifera şi Cynodon dactylon), ce necesită o foarte bună aerare a solului.
Combaterea vegetaţiei lemnoase, cu excepţia speciilor de talie joasă, se realizează numai în situaţia în care speciile lemnoase ocupă un spaţiu mai mare din fitocenoza pajiştilor. Această lucrare se va efectua manual pentru a se evita tasarea profundă a solului la folosirea utilajelor mecanice.
Pajiştile umede, situate în luncile râurilor sau în zonele depresionare, indiferent de zona în care se găsesc, se caracterizează prin condiţii naturale asemănătoare, determinate pe de o parte de nivelul apelor freatice şi regimul inundaţiilor, iar pe de altă parte de natura solului, format din depuneri aluvionare, bogate în substanţe nutritive. În aceste condiţii, în care majoritatea speciilor edificatoare a acestor habitate sunt indiferente faţă de troficitatea solului şi îşi procură elementele nutritive din materialele depuse de viituri, nu se recomandă fertilizarea. În luncile neinundabile şi pe platourile mai înalte se pot aplica, în doze moderate, îngrăşăminte minerale cu azot şi potasiu, sub formă de îngrăşăminte complexe.
6.1.2.2. Sistemul de exploatare Condiţiile foarte favorabile pentru dezvoltarea diferitelor plante, inclusiv a
speciilor cu valoare furajeră ridicată, fac ca pajiştile umede, situate în zona luncilor, văilor şi depresiunilor, să fie menţinute timp îndelungat într-o stare bună de productivitate şi calitate, atât prin aplicarea unor lucrări speciale de întreţinere cât şi prin folosirea unui sistem raţional de valorificare.
Folosirea pajiştilor umede se face diferenţiat, în funcţie de următorii factori staţionali: starea de umiditate a solului, nivelul apei freatice, structura floristică a fitocenozei, topografia terenului.

271
Toate tipurile de pajişti, situate de o parte şi de alta a albiilor râurilor, cu exces aproape permanent de umiditate din cauza inundaţiilor periodice, se vor folosi numai prin cosire extensivă, realizându-se 1-2 recolte pe an.
Producţia rezultată poate fi folosită direct în hrana animalelor, sau însilozată, iar pentru producerea fânului, uscarea se realizează numai pe suporţi.
De asemenea, pajiştile în care predomină speciile de Agrostis stolonifera şi Alopecurus pratensis se vor valorifica numai prin cosire, deoarece rezistenţa acestor specii la păşunat este foarte scăzută.
Pentru a prezerva diversitatea fitocenozelor de pajişti umede se recomandă , la intervale de 2-3 ani, cosirea acestor terenuri mai târziu, după fructificarea principalelor specii mai importante, în vederea realizării autoînsămânţării, inclusiv a unor specii de plante de interes conservativ (Angelica archangelica, Trollius europaeus şi unele specii de orhidee).
Pajiştile de Agropyron repens, Lolium perenne şi Poa pratensis pot fi valorificate prin păşunatul cu animalele, în următoarele condiţii: interzicerea păşunatului când solul este umed (inclusiv după retragerea apelor de inundaţie) şi corelarea încărcăturii de animale cu capacitatea de producţie a pajiştii.
Valorificarea mixtă (cosire, urmată de un păşunat uşor spre sfârşitul perioadei de vegetaţie) este posibilă pe pajiştile umede situate la baza versanţilor şi pe platourile mai înalte din depresiuni şi văi.
În vederea prevenirii distrugerii unor habitate de animale şi păsări, ce se întâlnesc în vegetaţia pajiştilor umede (mai ales în zonele de luncă, mlaştini şi bălţi), se recomandă, ca tehnica de lucru, cositul „centrifugal”, începând de la centrul parcelei spre margine, pentru retragerea la timp a acestor vieţuitoare.
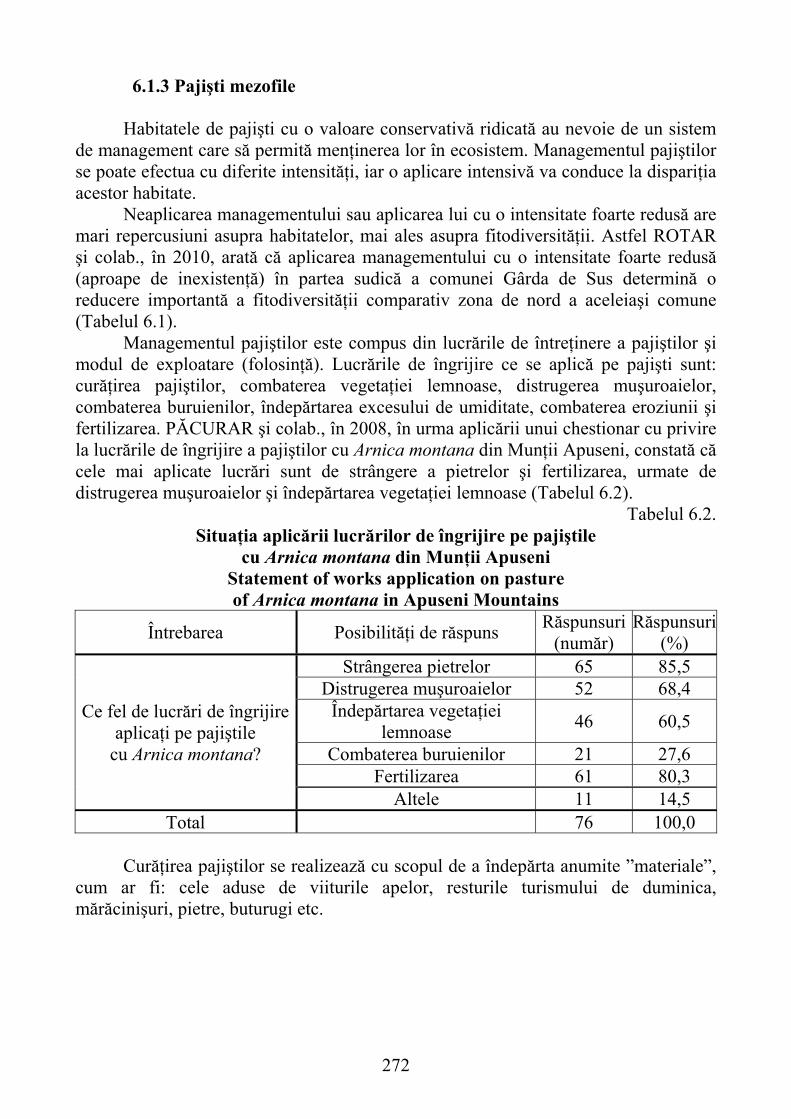
272
6.1.3 Pajişti mezofile
Habitatele de pajişti cu o valoare conservativă ridicată au nevoie de un sistem de management care să permită menţinerea lor în ecosistem. Managementul pajiştilor se poate efectua cu diferite intensităţi, iar o aplicare intensivă va conduce la dispariţia acestor habitate.
Neaplicarea managementului sau aplicarea lui cu o intensitate foarte redusă are mari repercusiuni asupra habitatelor, mai ales asupra fitodiversităţii. Astfel ROTAR şi colab., în 2010, arată că aplicarea managementului cu o intensitate foarte redusă (aproape de inexistenţă) în partea sudică a comunei Gârda de Sus determină o reducere importantă a fitodiversităţii comparativ zona de nord a aceleiaşi comune (Tabelul 6.1).
Managementul pajiştilor este compus din lucrările de întreţinere a pajiştilor şi modul de exploatare (folosinţă). Lucrările de îngrijire ce se aplică pe pajişti sunt: curăţirea pajiştilor, combaterea vegetaţiei lemnoase, distrugerea muşuroaielor, combaterea buruienilor, îndepărtarea excesului de umiditate, combaterea eroziunii şi fertilizarea. PĂCURAR şi colab., în 2008, în urma aplicării unui chestionar cu privire la lucrările de îngrijire a pajiştilor cu Arnica montana din Munţii Apuseni, constată că cele mai aplicate lucrări sunt de strângere a pietrelor şi fertilizarea, urmate de distrugerea muşuroaielor şi îndepărtarea vegetaţiei lemnoase (Tabelul 6.2).
Tabelul 6.2. Situaţia aplicării lucrărilor de îngrijire pe pajiştile
cu Arnica montana din Munţii Apuseni Statement of works application on pasture of Arnica montana in Apuseni Mountains
Întrebarea Posibilităţi de răspuns Răspunsuri
(număr) Răspunsuri
(%)
Ce fel de lucrări de îngrijire aplicaţi pe pajiştile
cu Arnica montana?
Strângerea pietrelor 65 85,5 Distrugerea muşuroaielor 52 68,4
Îndepărtarea vegetaţiei lemnoase
46 60,5
Combaterea buruienilor 21 27,6 Fertilizarea 61 80,3
Altele 11 14,5 Total 76 100,0
Curăţirea pajiştilor se realizează cu scopul de a îndepărta anumite ”materiale”,
cum ar fi: cele aduse de viiturile apelor, resturile turismului de duminica, mărăcinişuri, pietre, buturugi etc.
273
Tabelul 6.1. Influenţa managementului asupra fitodiversităţii pajiştilor semi-naturale din comuna Gârda de Sus
Influence of management of diversity on semi-natural grasslands in Gârda de Sus
Specificare Zona sudică a comunei Gîrda de Sus Zona nordică a comunei Gîrda de Sus
Nr.parcelei 4004 4048 4057 4072 4073 11 26 30 45 52
Localitatea Valea
Buciniş Biharia Biharia Biharia Biharia Marocu Marocu
Gîrda Seacă
Ocoale Ocoale
Altitudinea (m) 1014 1200 1250 860 880 1134 1245 1216 1305 1289
Expoziţia 335 335 200 320 45 300 220 240
Grosimea solului (cm) 12 9 8 7 5 20 10 15 <20 12
Panta % 32 11 25 21 26 15 17 7 7 6
Aplicarea lucrărilor de îngrijire
nu da da da nu da da da da da
Aplicarea fertilizării cu gunoi nu redus nu redus nu 5-6 t 3-5 t 12-16 t 16-17 t 16-20 t
Momentul fertilizării nu p - t nu p - t nu p - t p p p p - t
Păşunat continuu Vara -toamna
vara continuu continuu sept. toamna toamna nu toamna
Cosit nu nu nu nu nu Aug. Aug. Iul. Iul. Aug.
Folosinţă Mixtă nu nu nu nu nu da da da x da
Recolta kg/m2 masă verde 0,52 0,469 0,371 0,279 0,487 0,364 0,552 0,642 0,361 0,79
Nr. de specii (1 m2) 11 10 13 15 15 35 39 28 28 30
Nr. de specii (25 m2) 14 16 20 30 26 52 53 39 46 47
p – primavară t – toamnă p-t = primavară-toamnă

274
Aceste activităţi trebuie, în general, să se realizeze deoarece aplicarea unora dintre ele au creat anumite elemente de landşaft, ca de exemplu ”grămădarele de pietre” apărute în urma activităţii de strângere a pietrelor.
Grămădarele de pietre apar ca nişte movile de pietre de diferite forme (liniare, rotunde şi neregulate), cele mai vechi fiind acoperite de vegetaţie ierboasă sau lemnoasă (GÂRDA, 2010, Fig.6.1).
Fig. 6.1. Grămadă de pietre în fâneaţă (Hănăşeşti, Gîrda de Sus) PPiillee ooff ssttoonneess iinn tthhee mmeeaaddooww (Hănăşeşti, Gîrda de Sus)
Muşuroaiele care apar pe pajişti pot fi de origine animală şi vegetală. Pe
pajiştile montane sunt mai răspândite muşuroaiele de origine vegetală (VÎNTU şi colab.2004). Pe muşuroaie se instalează o vegetaţie specifică, care este nevaloarosă din punct de vedere furajer. De asemenea, prezenţa muşuroaielor îngreunează foarte mult aplicarea lucrărilor de întreţinere şi exploatare. În unele habitate de pajişti, distrugerea muşuroaielor apare ca o lucrare necesară, dar în altele nu se recomandă deoarece aici îşi găsesc locul numeroase specii de origine animală (Fig.6.2).
Îndepărtarea vegetaţiei lemnoase este deosebit de importantă mai ales în cazul pajiştilor semi-naturale deoarece din lipsa unor lucrări de îngrijire corespunzătoare aceste vor fi invadate de vegetaţia lemnoasă specifică zonei sau etajului de vegetaţie (Fig. 6.3).

275
Fig.6.2. Distrugerea manuală a muşuroaielor în vederea exploatării prin
cosit (Vârful Petreasa, comuna Horea, Munţii Apuseni) Destruction of the mounds manually by mowing
(Peak Petreasa, Horea, Apuseni Mountains)
Fig. 6.3. Invadarea vegetaţiei lemnoase pe o păşune din comuna Avram Iancu Invasion of woody vegetation on pasture, Avram Iancu

276
Defrişarea vegetaţie lemnoase se face pe baza unor studii şi documentaţii (amenajamete silvopastorale) în care se prevăd toate detaliile privind organizarea şi efectuarea lucrării (prevenirea eroziunii, loc de refugiu pentru animale etc.).
De asemenea, sunt excluse de la defrişare speciile rare şi cele declarate monumente ale naturii (aceste specii vor fi menţionate mai jos în cazul fiecărui tip de habitat).
În praticultură sunt considerate buruieni plantele fără valoare furajeră, cel care depreciază calitatea produselor obţinute de la animale, cele care dăunează vegetaţie pajiştilor şi cele toxice. Cauzele care conduc la înburuienarea pajiştilor sunt multiple, cele mai importante sunt condiţiile staţionale şi lipsa sistemului raţional de folosire a pajiştii. Combaterea buruienilor se poate face prin diferite metode (preventive şi curative), care vor fi detaliate mai jos pentru fiecare tip de habitat.
Îndepărtarea excesului de umiditate se realizează prin mai multe metode (prin canale deschise, drenajul cârtiţă, drenajul biologic etc.), dar nu se recomandă când este vorba de păstrarea habitatelor de pajişti umede.
Dintre lucrările menţionate mai sus, cea mai mare importanţă o are fertilizarea. Fertilizarea poate determina schimbarea covorului ierbos într-un timp foarte scurt. Chiar şi aplicarea unor cantităţi reduse de îngrăşăminte (10 t gunoi de grajd / ha sau 50N 25P2O5 25K2O + 10 t/ha gunoi de grajd aplicate anual) poate determina schimbări importante la nivelul covorului ierbos şi o reducere puternică a fitodiversităţii. Astfel, PĂCURAR şi ROTAR (2008) arată că prin aplicarea a 10 t / ha gunoi de grajd anual pe o pajişte de Festuca rubra din Munţii Apuseni după o perioadă de 7 ani, fitodiversitatea s-a restrâns cu 10 specii (Tabelul 6.3). Prin combinarea celor două tipuri de îngrăşăminte în cantităţile amintite, diversitatea floristică s-a restrâns şi mai mult cu până la 15 specii. Aplicarea anuală a 10 t/ha gunoi + 100N 50P2O5 50K2O a determinat cea mai mare reducere a fitodiversităţii cu până la 21 de specii. Aşadar, în vederea menţinerii fitodiversităţii se recomandă aplicarea unor cantităţi mai mici de îngrăşământ comparativ cu cele folosite în experienţa de faţă sau cel mult de 10 t/ha gunoi sau 50N 25P2O5 25K2O aplicate o dată la 2-3 ani.
Sistemul de exploatare poate fi prin cosit, prin păşunat sau mixt. Acesta poate să aibă o aplicare intensivă sau extensivă. În recomandarea unui sistem sau a altuia trebuie să ţinem cont de pretabilitatea fitocenozei la cosit, păşunat sau la sistemul mixt.
De asemenea, în cazul în care dorim să menţinem în fitocenoză o anumită specie cu o mare valoare conservativă, atunci modul de exploatare trebuie recomandat cu stricteţe, în funcţie de exigenţele speciei respective cu privire la păşunat, călcat sau cosit. Exploatarea prin cosit trebuie să ţină cont de următoarele elemente: tehnica de cosit, momentul începerii cositului, înălţimea de cosit, frecvenţa cosirilor şi modul de uscare al ierbii.
Exploatarea prin păşunat are în vedere următoarele aspecte: sistemul de păşunat, specia de animale ce păşunează, momentul începerii şi terminării păşunatului, capacitatea de păşunat, valoarea pastorală etc.
277
Tabelul 6.3 Influenţa fertilizării organice, minerale şi organo-minerale
asupra unei pajişti de Festuca rubra
Influence of organic, mineral and organo-mineral fertilization on a meadow of Festuca rubra
Variante experimentale
SPECII martor10t
gunoi/ha
10t gunoi/ha + 50N 25P2O5
25K2O
100N 50P2O5 50K2O
10t gunoi/ha + 100N 50P2O5 50K2O
Acoperire % 96 98 98 100 100 Specii % Poaceae 41,5 41,5 56,5 88,5 82 Festuca rubra 22,5 18,5 4 2 6 Anthoxanthum odoratum 1 18 1 0,2 16 Trisetum flavescens 2,5 10 20 Cynosurus cristatus 1 0,2 1 0,2 Festuca pratensis 0,2 0,2 Briza media 2 Agrostis capillaris 17 41,5 40 65 60 Fabaceae 11,5 26,5 6,5 45 2,5 Lathyrus pratensis 0,5 Lotus corniculatus 3 3,5 0,2 Trifolium repens 6 7 1 0,2 0,2 Trifolium pratense 2 8 1 0,2 0,2 Vicia cracca 0,2 4 3 1,5 Cyperacee and Juncaceae 1 - - - - Luzula multiflora 0,2 Carex pallescens 0,2 Alte familii botanice 56,83 48,8 40 20 13,5 Achilea milefolium 0,2 Alchemila vulgaris 24 8 7 4 7 Campanula patula 0,2 Carlina acaualis 0,2 Carum carvi 0,3 0,3 Centaurea phrygia agg. 8 11 15 6 10 Colchicum autumnale 2 2 2 0,2 1 Crepis biennis 0,2 0,5 0,2
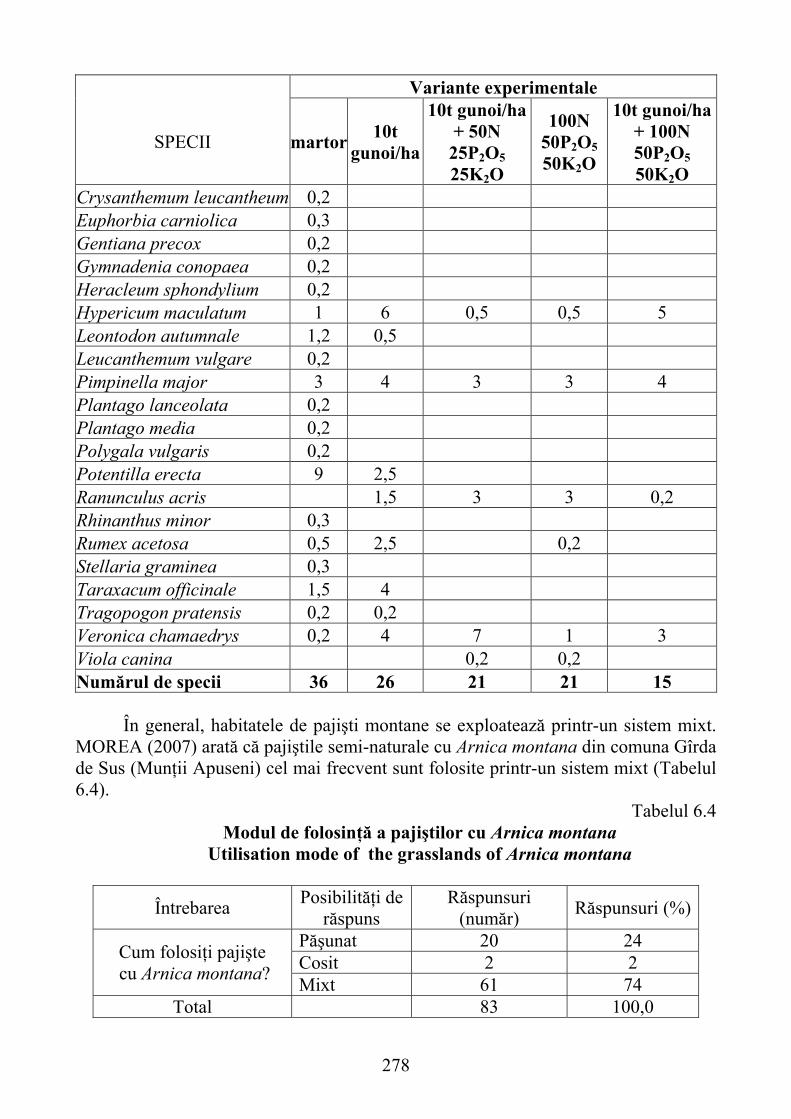
278
Variante experimentale
SPECII martor10t
gunoi/ha
10t gunoi/ha + 50N 25P2O5 25K2O
100N 50P2O5 50K2O
10t gunoi/ha + 100N 50P2O5 50K2O
Crysanthemum leucantheum 0,2 Euphorbia carniolica 0,3 Gentiana precox 0,2 Gymnadenia conopaea 0,2 Heracleum sphondylium 0,2 Hypericum maculatum 1 6 0,5 0,5 5 Leontodon autumnale 1,2 0,5 Leucanthemum vulgare 0,2 Pimpinella major 3 4 3 3 4 Plantago lanceolata 0,2 Plantago media 0,2 Polygala vulgaris 0,2 Potentilla erecta 9 2,5 Ranunculus acris 1,5 3 3 0,2 Rhinanthus minor 0,3 Rumex acetosa 0,5 2,5 0,2 Stellaria graminea 0,3 Taraxacum officinale 1,5 4 Tragopogon pratensis 0,2 0,2 Veronica chamaedrys 0,2 4 7 1 3 Viola canina 0,2 0,2 Numărul de specii 36 26 21 21 15
În general, habitatele de pajişti montane se exploatează printr-un sistem mixt. MOREA (2007) arată că pajiştile semi-naturale cu Arnica montana din comuna Gîrda de Sus (Munţii Apuseni) cel mai frecvent sunt folosite printr-un sistem mixt (Tabelul 6.4).
Tabelul 6.4 Modul de folosinţă a pajiştilor cu Arnica montana
Utilisation mode of the grasslands of Arnica montana
Întrebarea Posibilităţi de
răspuns Răspunsuri
(număr) Răspunsuri (%)
Cum folosiţi pajişte cu Arnica montana?
Păşunat 20 24 Cosit 2 2 Mixt 61 74
Total 83 100,0

279
Pajiştile mezofile de Agrostis capillaris-Festuca rubra şi Agrostis capillaris- Anthoxanthum odoratum din punct de vedere agronomic sunt moderat acidofile, moderat nitrofile, mediu rezistente la cosit, mediu rezistente la călcat şi mediu rezistente la păşunat (GÂRDA, 2010). Recomandarea tehnologiei de întreţinere şi folosire trebuie să aibă în vedere caracterele fitocenozelor deoarece schimbarea condiţiilor de troficitate poate să determine anumit fluctuaţii sau succesiuni nedorite.
6.1.3.1. Lucrări de întreţinere Curăţirea pajiştilor constă în strângerea pietrelor de pe pajişti. Această lucrare
se justifică numai acolo unde s-a practicat timp îndelungat (nu se recomandă ca o lucrare nouă pentru regiunile unde nu s-a mai practicat) deoarece, probabil, a avut o contribuţie la aspectul floristic actual şi la crearea unor elemente de landşaft (grămădarele de pietre). Recomandăm curăţirea pajiştilor de orice alte „materiale” cum ar fi: pungile de plastic, peturile, pungile de hârtie, ş.a. Distrugerea muşuroaielor, mai ales a celor de origine vegetală, se recomandă atunci când au depăşit 40 % din suprafaţa unei pajişti, dar numai cu acordul şi sub supravegherea organelor de stat competente. Combaterea buruienilor. Buruienile ce apar în etajul alpin şi subalpin sunt: Aconitum anthora, Aconicum tauricum, Aconicum toxicum, Colchicum autumnale, Euphorbia cyparisias, Hypericum maculatum, Ranunculus nemorosus, Stelaria graminea, Veratrum album etc. Dintre acestea , o răspândire mai largă şi un pericol mai mare pentru animale o au speciile Veratum album şi Colchicum autumnale. De asemenea, sunt foarte răspândite speciile nitrofile, ca: Urtica dioica, Rumex alpinus şi Rumex acifolius. Există şi alte multe specii de buruieni şi recomandăm combaterea lor manuală (exclus folosirea erbicidelor sau a altor substanţe), dar cu o consultare prealabilă cu organele competente ale statului, pentru a evita distrugerea unei specii cu valoare conservativă.
Îndepărtarea vegetaţiei lemnoase. În etajele alpin şi subalpin vegetaţia lemnoasă are o importanţă deosebită , cu o tendinţă de extindere, care, uneori poate ajunge la o acoperire foarte mare, diminuând astfel, suprafaţa pajiştii. Recomandăm combaterea manuală a speciilor ca Vaccinium myrtilus şi Vaccinium vitis-idae în condiţiile unui plan riguros şi prezentat organelor competente. În planurile de defrişare, indiferent de condiţiile în care se găsesc, nu intră speciile periclitate sau declarate Monumente ale Naturii, cum ar fi: Pinus cembra, Pinus sylvestris, Larix decidua, Rhodondendron koschyi, Juniperus sibirica, Bruckentalia spiculifolia, Loiseleuria precumbens etc.
Fertilizarea habitatelor de pajişti mezofile se realizează numai cu îngrăşăminte organice, în cantităţi cuprinse între 5-10 t / ha gunoi de grajd, administrate o dată la 2-3 ani. Gunoiul de grajd trebuie să fie bine fermentat şi să provină de la bovine, cabaline sau ovine. Aplicarea gunoiului se realizează toamna şi numai în cazuri excepţionale primăvara.

280
6.1.3.2. Sistemul de exploatare Habitatele de pajişti mezofile pot fi exploatate prin cosit sau printr-un sistem
mixt. Cositul se va realiza extensiv, când se ia o coasă pe an şi numai în cazuri excepţionale se vor efectua două coase. Momentul de începere a cositului va corespunde cu faza de fructificare a plantelor din alte familii botanice. Cositul se va realiza fie manual, fie mecanic, la o înălţime de 5-6 cm. Cositul mecanizat se va realiza cu maşini de cosit rotative uşoare, ca să nu determine tasarea solului. Acest mecanism are repercusiunile cele mai mici asupra faunei. Tehnica de deplasare cu maşina de cosit se realizează de la centru spre extremităţi, pentru ca vieţuitoarele să aibă timp să se retragă. Uscatul ierbii se recomandă a se realiza pe sol, pentru că prin manipularea manuală a fânului multe inflorescenţe se vor scutura pe pajişte şi în acest fel unele seminţe vor avea posibilitatea să germineze. De asemenea, existenţa căpiţelor de fân au un rol foarte important în aspectul sezonier al landşaftului cultural. Exploatarea prin sistemul mixt presupune cositul pajiştilor aşa cum s-a descris mai sus şi realizarea unui păşunat extensiv toamna, după ce animalele se întorc de pe păşunea comunală. În acest caz pajiştea se coseşte o singură dată, iar coasa a doua va fi destinată păşunatului. Păşunatul trebuie să înceapă când iarba are 10 – 15 cm înălţime şi să înceteze cu 25 – 30 zile înainte de venirea îngheţurilor permanente. Încărcătura de animale se recomandă a fii de 0,6 – 0,8 UVM / ha 6.2. GOSPODĂRIREA HABITATELOR DEGRADATE DE NARDUS STRICTA
6.2.1. Lucrări biopedoameliorative generale
Răspândirea speciei Nardus stricta (ţepoşica, părul porcului) de la 200 m altitudine în depresiunile din nordul ţării noastre, până la peste 2 200 m altitudine în munţii înalţi, ne demonstrează o plasticitate ecologica deosebită. Întreg lanţul Munţilor Carpaţi, la care se adaugă dealurile şi depresiunile cu climat mai umed ce îi însoţesc, sunt acoperite cu întinse suprafeţe de pajişti, în care nardetele sunt bine reprezentate. Prezenţa nardetelor este semnalată numai unde precipitaţiile medii anuale depăşesc 600 mm până la 1 200 (1400) mm cu temperaturi medii anuale sub 6-8 0C până la 0-20C., în zona pădurilor de stejar, continuând cu etajele gorunului, fagului, molidului până la cel al jneapănului, pe terenuri plane până la o înclinaţie a pantelor de 450.
Nardetele indică covor ierbos degradat, producţie şi calitate furajeră necorespunzătoare, soluri acide, oligobazice, conţinut scăzut în elemente fertilizante, exces de umiditate, păşunat permanent şi selectiv cu intensitate moderată etc. Diferenţierea şi descrierea tipurilor de nardete s-a făcut după anumite criterii în care compoziţia floristică, condiţiile staţionale şi posibilităţile de îmbunătăţire a acestor pajişti degradate, au avut un rol preponderent în elaborarea unei clasificări unitare (MARUŞCA, 1982).

281
În judeţul Brasov, nardetele sunt bine reprezentate, începând cu 480 m altitudine în Depresiunea Făgăraşului până la 2 210 m altitudine în Masivul Bucegi, ceea ce a permis ca pe un spaţiu relativ restrâns să fie studiate principalele tipuri de pajişti cu Nardus stricta existente şi în alte zone din ţara noastră. Din cele şapte tipuri de nardete descrise, două aparţin etajului subalpin, trei zonei montane şi două zonei colinare depresionare.
Pentru stabilirea unor variante tehnologice de îmbunătăţire a fondului funciar s-au conturat subtipurile : a = soluri profunde ; b = soluri scheletice ; c = cu vegetaţie lemnoasă (arborescentă) şi d = cu muşuroaie înţelenite la care se adaugă numeroase combinaţii între acestea, întâlnite frecvent pe teren. Sub aspectul gradului de mecanizare a lucrărilor de îmbunătăţire, determinat de înclinaţia pantelor, tipul 1 si 4 sunt fără posibilităţi de mecanizare terestră cu mijloacele actuale din dotare, tipul 3 si 6 sunt parţial mecanizabile şi tipul 2, 5 si 7 sunt mecanizabile în totalitate. Lucrările biopedoameliorative generale constau din: defrişarea vegetaţiei lemnoase, nivelarea terenului, adunarea pietrelor, amendarea calcică, desecare dacă este cazul şi alte lucrări prezentate în Tabelul 6.5. După încheierea acestor lucrări obligatorii se trece la faza următoare de îmbunătăţire a covorului ierbos degradat.
6.2.2. Lucrări de îmbunătăţire şi valorificarea producţiei
În procesul de îmbunătăţire a nardetelor există în momentul de fată o gamă largă de variante tehnologice (Tabelul 6.6.).
Un prim grup de variante se referă la posibilităţile oferite de fertilizarea cu îngrăşăminte organice şi chimice, prin care se stimulează speciile valoroase furajere din flora spontană cum sunt : Festuca rubra, Agrostis tenuis, Poa pratensis, Festuca pratensis, Poa trivialis, Trifolium repens şi multe altele, care pot substitui specia dominantă, Nardus stricta, până la dispariţie.
Dintre acestea, târlirea 3-5 nopţi o oaie/mp, se poate aplica practic pe toate tipurile de nardete. Târlirea, deşi este una din cele mai valoroase metode de îmbunătăţire a nardetelor, poate fi aplicată în zona montană numai până la 25 % din suprafaţa de nardete (PAVEL 1962, PUŞCARU 1956, RESMERIŢĂ 1969).
Suprafeţele îmbunătăţite prin târlire pot fi simţitor mărite, dacă în apropierea nardetelor exista pajişti îmbunătăţite sau alte surse de furaje, care permit întreţinerea unui număr mai mare de animale cu care să se facă târlirea. Cu toate acestea în etapa actuală, numai prin târlire nu se poate cuprinde întreaga suprafaţă de nardete ce urmează a fi îmbunătăţită.
Aplicarea gunoiului de grajd (30-40 t/ha) este posibilă din punct de vedere economic numai în zonele unde există în apropiere surse de dejecţii de la animalele aflate în stabulaţie, cu posibilităţi de transport şi împrăştiere pe teren.
Având în vedere suprafeţele destul de restrânse de nardete care pot beneficia de îngrăşăminte organice, principala măsură de îmbunătăţire, într-o primă etapă, o constituie fertilizarea cu îngrăşăminte chimice în doze mari, 150-200 kg/ha N, 50-100 kg/ha P2O5 şi uneori 50 kg/ha K2O (PAVEL 1962).
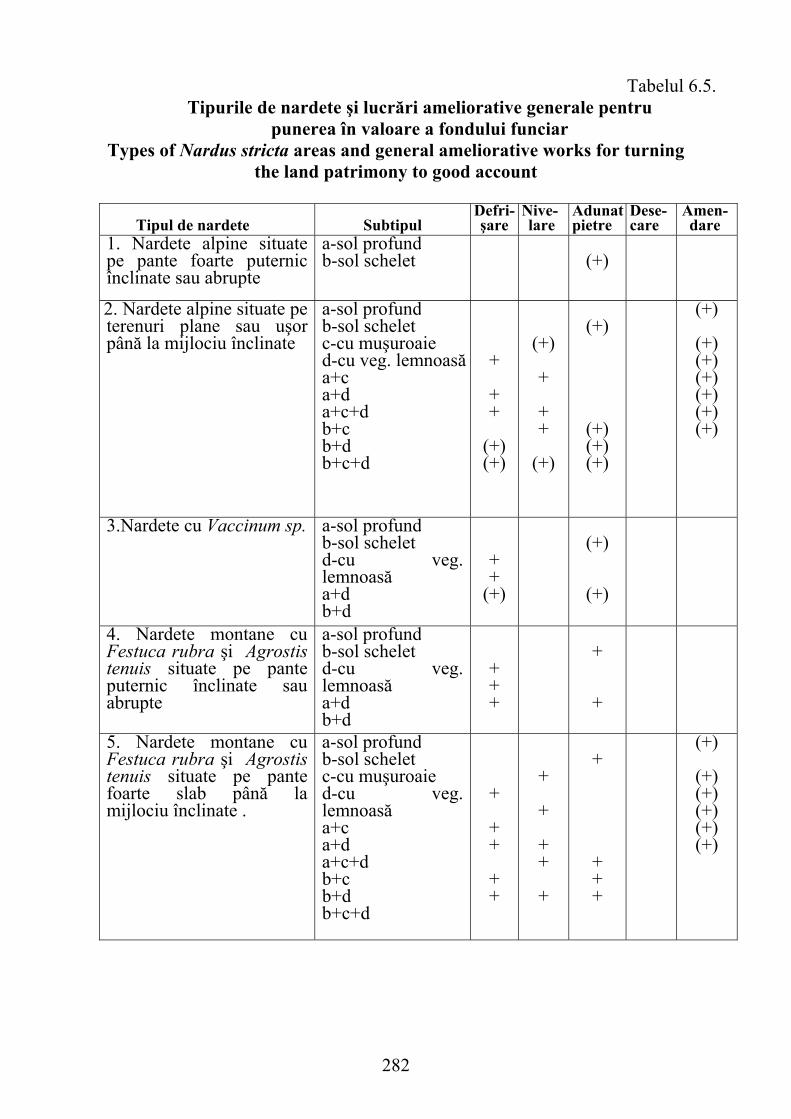
282
Tabelul 6.5. Tipurile de nardete şi lucrări ameliorative generale pentru
punerea în valoare a fondului funciar Types of Nardus stricta areas and general ameliorative works for turning
the land patrimony to good account
Tipul de nardete Subtipul
Defri-şare
Nive-lare
Adunat pietre
Dese-care
Amen-dare
1. Nardete alpine situate pe pante foarte puternic înclinate sau abrupte
a-sol profundb-sol schelet
(+)
2. Nardete alpine situate pe terenuri plane sau uşor până la mijlociu înclinate
a-sol profundb-sol schelet c-cu muşuroaie d-cu veg. lemnoasăa+c a+d a+c+d b+c b+d b+c+d
+
+ +
(+) (+)
(+)
+
+ +
(+)
(+)
(+) (+) (+)
(+)
(+) (+) (+) (+) (+) (+)
3.Nardete cu Vaccinum sp. a-sol profundb-sol schelet d-cu veg. lemnoasă a+d b+d
+ +
(+)
(+)
(+)
4. Nardete montane cu Festuca rubra şi Agrostis tenuis situate pe pante puternic înclinate sau abrupte
a-sol profundb-sol schelet d-cu veg. lemnoasă a+d b+d
+ + +
+
+
5. Nardete montane cu Festuca rubra şi Agrostis tenuis situate pe pante foarte slab până la mijlociu înclinate .
a-sol profundb-sol schelet c-cu muşuroaie d-cu veg. lemnoasă a+c a+d a+c+d b+c b+d b+c+d
+
+ +
+ +
+
+
+ +
+
+
+ + +
(+)
(+) (+) (+) (+) (+)

283
Tipul de nardete Subtipul
Defri-şare
Nive-lare
Adunat pietre
Dese-care
Amen-dare
6. Nardete cu diferite specii mezofile şi xeromezofile situate pe pante mijlociu până la foarte puternic înclinate .
a-sol profundb-sol schelet d-cu veg. lemnoasă a+d b+d
+ + +
(+)
(+)
7. Nardete din depresiuni cu diferite specii mezohigrofite şi hifrofite, situate pe terenuri plana sau uşor înclinate.
a-sol profundb-sol schelet c-cu muşuroaie d-cu veg. lemnoasă a+c a+d a+c+d b+c b+d b+c+d
+
+ +
+ +
+
(+) +
(+) + +
+
+
+ + +
+(+) (+) (+) + + +
(+) (+) (+)
+(+) + + + + +
(+) (+) (+)
Menţiune: + - lucrări obligatorii; (+) - lucrări necesare în unele cazuri
Pentru a evita apariţia şi proliferarea unor specii nitrofile nevaloroase, cum este
Deschampsia caespitosa, pe nardetele colinare şi din depresiuni, doza de azot se reduce la 100 kg/ha (MARUŞCA 1982)
Aplicarea îngrăşămintelor chimice cu mijloace terestre sau aviaţia utilitară, trebuie făcută cu mult discernământ, pe suprafeţele unde covorul ierbos are în componenţă şi specii valoroase furajere care după ameliorarea condiţiilor trofice ale solului, în prezenta unei folosiri raţionale, pot înlocui treptat până la dispariţie specia dominanta Nardus stricta. Al doilea grup de variante de îmbunătăţire a nardetelor fac apel la introducerea de specii furajere cultivate, prin supraînsămânţare şi reînsămânţare, după distrugerea prin diferite mijloace, mecanice şi chimice, a covorului ierbos degradat.
Supraînsămânţarea suprafeţelor intens târlite (BĂRBULESCU 1975), sau a golurilor rezultate după nivelarea muşuroaielor înţelenite, scoaterea cioatelor, arderea martoanelor, adunatul pietrelor etc. reprezintă o măsura complementară de introducere a unor specii cultivate valoroase, mai receptive la noile condiţii create pentru sporirea şi valorificarea producţiei.Cu toate că suprafeţele unde se efectuează supraînsămânţarea sunt relativ restrânse ca arie, totuşi această măsura prezintă importanţă practică deoarece se grăbeşte instalarea speciilor valoroase.
Una din principalele măsuri de îmbunătăţire a nardetelor o reprezintă reînsămânţarea ( LAZA 1979, MARUŞCA 1982, PUIA şi POPESCU 1971).
Prin reînsămânţare se înfiinţează o pajişte semănata, după distrugerea integrală a covorului ierbos degradat prin lucrări mecanice (arat, prelucrarea cu freza) chimice (erbicidare totală cu 5 1/ha ,,Gramoxone”) şi combinaţia dintre acestea (erbicidare + târlire, erbicidare + mobilizare superficială) şi altele.
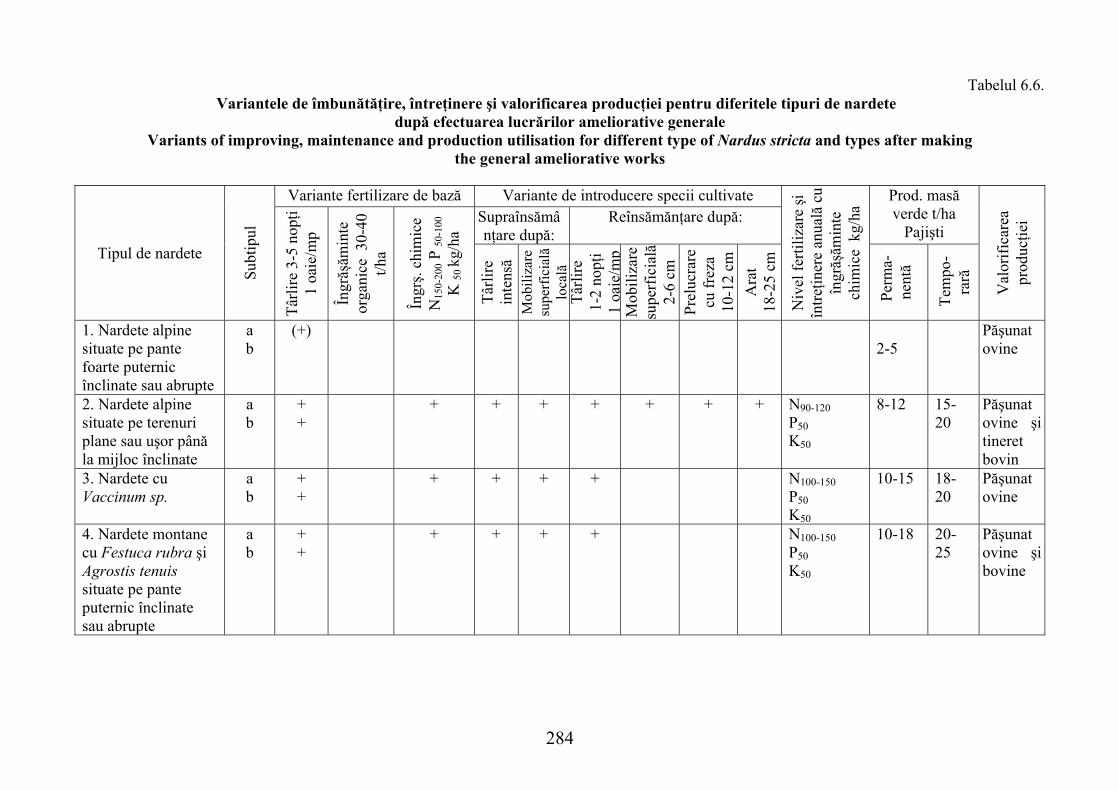
284
Tabelul 6.6. Variantele de îmbunătăţire, întreţinere şi valorificarea producţiei pentru diferitele tipuri de nardete
după efectuarea lucrărilor ameliorative generale Variants of improving, maintenance and production utilisation for different type of Nardus stricta and types after making
the general ameliorative works
Tipul de nardete S
ubti
pul
Variante fertilizare de bază Variante de introducere specii cultivate
Niv
el f
erti
liza
re ş
i în
treț
iner
e an
uală
cu
îngrășăm
inte
ch
imic
e k
g/ha
Prod. masă verde t/ha
Pajişti
Val
orif
icar
ea
prod
ucţie
i
Târ
lire
3-5
nopţi
1
oaie
/mp
Îngrășăm
inte
or
gani
ce 3
0-40
t/
ha
Îngrş.
chi
mic
e N
150-
200
P 5
0-10
0 K
50 kg
/ha
Supraînsămânţare după:
Reînsămănţare după:
Târ
lire
in
tensă
Mob
iliz
are
su
perf
icia
lă
loca
lă
Târ
lire
1-
2 no
pţi
1
oaie
/mp
Mob
iliz
are
supe
rfic
ială
2-
6 cm
P
relu
crar
e
cu f
reza
10
-12
cm
Ara
t 18
-25
cm
Per
ma-
ne
ntă
Tem
po-
rară
1. Nardete alpine situate pe pante foarte puternic înclinate sau abrupte
a b
(+) 2-5
Păşunat ovine
2. Nardete alpine situate pe terenuri plane sau uşor până la mijloc înclinate
a b
+ +
+ + + + + + + N90-120 P50 K50
8-12 15-20
Păşunat ovine şi tineret bovin
3. Nardete cu Vaccinum sp.
a b
+ +
+ + + + N100-150 P50 K50
10-15 18-20
Păşunat ovine
4. Nardete montane cu Festuca rubra şi Agrostis tenuis situate pe pante puternic înclinate sau abrupte
a b
+ +
+ + + + N100-150 P50 K50
10-18 20-25
Păşunat ovine şi bovine

285
Tipul de nardete
Sub
tipu
l
Variante fertilizare de bază Variante de introducere specii cultivate
Niv
el f
erti
liza
re ş
i în
treț
iner
e an
uală
cu
îngrășăm
inte
ch
imic
e k
g/ha
Prod. masă verde t/ha
Pajişti
Val
orif
icar
ea
prod
ucţie
i
Târ
lire
3-5
nopţi
1
oaie
/mp
Îngrășăm
inte
or
gani
ce 3
0-40
t/
ha
Îngrş.
chi
mic
e N
150-
200
P 5
0-10
0 K
50 kg
/ha
Supraînsămăn- ţare după:
Reînsămănţare după:
Târ
lire
in
tensă
Mob
iliz
are
supe
rfic
ială
lo
cală
T
ârli
re
1-2
nopţ
i
1 oa
ie/m
p M
obil
izar
e su
perf
icia
lă
2-6
cm
Pre
lucr
are
cu
fre
za
10-1
2 cm
Ara
t 18
-25
cm
Per
ma-
ne
ntă
Tem
po-
rară
5. Nardete montane cu Festuca rubra şi Agrostis tenuis situate pe pante foarte slab până la mijlociu înclinate .
a b
+ +
+ +
+ +
+ +
+ +
+ +
+ + + N100-150 P50 K50
20-25 25-30
Păşunat bovine şi ovine Mixt Fâneaţă
6. Nardete cu diferite specii mezofile şi xeromezofile situate pe pante mijlociu până la foarte puternic înclinate .
a b
+ +
+ +
+ +
+ +
+ +
+ +
+ N120-180 P50-100 K50
20-30 25-35
Păşunat bovine şi ovine
7. Nardete din depresiuni cu diferite specii mezohigrofite şi hifrofite, situate pe terenuri plana sau uşor înclinate.
a b
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ + +
N150-200 P100 K50
20-35 30-45
Mixt Păşunat bovine Fâneaţă
Menţiune: - Se alege numai una din fiecare variantă de fertilizare de bază sau de introducere specii cultivate prin supra sau reînsămănţare; - Subtipul "a" se referă la nardete situate pe soluri profunde iar "b" sunt nardetele situate pe soluri scheletice cu roci dure la suprafaţă sau la
mică adâncime

286
Dintre acestea, erbicidarea cu 5 1/ha ,,Gramoxone” după 14 zile, semănatul târlirea 1-2 nopţi poate fi aplicată practic pe aproape toate tipurile de nardete, inclusiv pe cele cu restricţii sub aspectul mecanizării. Întrucât această metodă nu face apel la mobilizarea solului se pune problema dacă trebuie sau nu inclusă în categoria lucrărilor de reînsămânţare. Deoarece prin erbicidare se distruge aproape integral covorul ierbos degradat, iar prin semănat şi târlire, ca rezultat final apare o pajişte nouă, cu specii cultivate, considerăm a fi inclusă în categoria lucrărilor de reînsămânţare.
De altfel şi la metoda clasica de reînsămânţare prin mobilizarea ţelinei cu freza sau plugul nu se realizează o distrugere integrală a tuturor plantelor din vechiul covor ierbos, la care dacă se adaugă şi rezerva de sămânţa viabilă din sol, nu se poate afirma că rezultă şi mai ales se menţine, o pajişte pură, formată numai din componentele amestecului semănat.
Faţă de metoda tradiţionala de târlire (3-5 nopţi), pe aceeaşi pajişte degradată şi cu acelaşi efectiv de animale, prin erbicidare totală, semănat şi târlire de încorporare (1-2 nopţi), suprafeţele îmbunătăţite cresc de două, trei ori, rezultând pajişti semănate foarte valoroase. Pe aceste pajişti se pot întreţine tot mai multe animale cu care se continuă şi se multiplică suprafeţele erbicidate şi târlite, astfel că vor fi suficiente animale într-o zonă mai greu accesibilă că pe parcursul a mai mulţi ani, să se îmbunătăţească integral o suprafaţă de nardete, numai prin această metodă. Evident, ca la oricare pajişte semănată, este necesară în continuare fertilizarea anuală de întreţinere cu îngrăşăminte chimice ce se pot aplica şi cu aviaţia utilitară pe suprafeţele cu restricţii de mecanizare terestră.
Pe suprafeţele de nardete mecanizabile, o metodă de reînsămânţare constă în erbicidare cu 5 1/ha ,,Gramoxone" şi prelucrarea superficială pe rânduri sau totală a ţelinei cu freza-semănătoare.
In absenţa acestui utilaj, se poate mobiliza ţelina rămasă după erbicidare, cu ajutorul grapei cu discuri prin 2-3 treceri pe direcţii perpendiculare urmată de tasare, semănat şi din nou tasare cu un tăvălug.
Prelucrarea ţelinei cu freza la 10-12 cm adâncime pe solurile lipsite de fragmente de roci dure la suprafaţă, reprezintă de asemenea una din metodele cele mai bune de pregătire a terenului în vederea însămânţării. După prelucrarea, la o săptămâna mai însorită, se tasează cu un tăvălug inelar, se seamănă şi imediat se tasează din nou cu un tăvălug neted. . .
Pe nardetele cu strat gros de ţelina, pe terenuri denivelate cu muşuroaie şi resturi de rădăcini rezultate după defrişarea vegetaţiei lemnoase se recomandă desţelenirea în toamnă cu plugul la adâncimea de 18-25 cm, precedată de premărunţirea ţelinei cu grapa cu discuri. În primăvară, terenul se pregăteşte normal cu grapa cu discuri, se tasează, se seamănă şi din nou se tasează cu tăvălugul.
Epocile optime de supraînsămânţare şi reînsămânţare, indiferent prin ce metodă se execută, variază în funcţie de zona în care se efectuează lucrările. În toate situaţiile epoca optima este primăvara cât mai devreme posibil, la fel şi vara, întrucât nardetele sunt situate în areale cu precipitaţii suficiente şi destul de bine eşalonate.

287
Restricţii apar doar spre toamnă, când plantele semănate trebuie să aibă suficient timp pentru a se dezvolta şi înfrăţi, înainte de venirea brumelor şi îngheţurilor la sol.
Amestecurile de ierburi şi cantităţile de sămânţa utilizate la hectar sunt cele cunoscute, adaptate de asemenea condiţiilor pedoclimatice, nivel de fertilizare, mod de folosire etc.
După alegerea şi aplicarea uneia din variante de îmbunătăţire descrise până acum, care au avut ca principal scop, înlocuirea speciei Nardus stricta cu specii valoroase furajere problema principală rămâne aplicarea măsurilor de întreţinere prin fertilizare şi alte lucrări.
Fertilizarea diferenţiată de întreţinere în funcţie de tipul de nardete, face posibilă menţinerea unei productivităţi ridicate, cât şi a speciilor valoroase din covorul ierbos.
Dozele de îngrăşăminte azotoase care depăşesc 100 kg ha N, se recomandă să fie fracţionate şi aplicate în reprize a câte 50 kg/ha, în vederea eşalonării producţiei de iarbă de-a lungul perioadei de păşunat. În zona montană inferioară şi cea colinară-depresionară, unde se pot realiza 20-45 t ha masă verde, sunt necesare lucrări de tarlalizare a suprafeţelor îmbunătăţite, în vederea folosirii lor prin păşunat raţional.
Valorificarea producţiei se face la altitudini mai înalte prin păşunat cu ovine şi tineret taurin, la cele medii în zone permanent locuibile prin păşunat cu bovine adulte, mixt şi fâneaţă iar pe cele de joasă altitudine prin toate metodele de folosire în funcţie de necesităţi şi condiţii economice.
6.2.3. Producţia de lapte pe nardetele îmbunătăţite
Cele mai degradate pajişti din zona montană sunt cele invadate şi dominate de specia Nardus stricta numită popular ţăpoşică, părul porcului, năgară şi altele. Locul de origine al acestei specii slab furajere a fost la marginea turbăriilor oligotrofe din zona montană înaltă. După defrişarea vegetaţiei lemnoase, înţelenirea terenului şi valorificarea prin păşunat neraţional cu animale, specia Nardus stricta s-a instalat şi a copleşit până la urmă pajişti mai valoroase de Festuca rubra, Agrostis capillaris, Festuca rupicola şi altele, formând nardete. Suprafaţa de nardete din ţara noastră ajunge la cca. 1 milion de hectare, fiind una din cele mai răspândite habitate de pajişti din zona montană. Pentru aceste considerente la Baza de cercetări pajişti montane Blana – Bucegi în anul 1995 s-a înfiinţat un lot demonstrativ cu cele mai bune variante de îmbunătăţire a nardetelor în suprafaţă de 0,75 ha fiecare, care s-au păşunat cu vaci de lapte din rasa Brună de Maramureş (Schwyz), o durată medie de 85 zile în condiţiile din etajul subalpin. Înainte de îmbunătăţire, pajiştea era dominată de Nardus stricta în proporţie de 40 – 60 % (MARUŞCA 2010) Variantele de îmbunătăţire a nardetelor au fost următoarele:
Grupa A, varianta martor, păşunat în afara câmpului pe nardete neîmbunătăţite între anii 1996 – 1999;

288
Grupa B, fertilizare cu îngrăşăminte chimice 3 ani consecutiv (1996 – 1998) cu 150 kg/ha N, 50 kg/ha P2O5 şi 50 kg/ha K2O, urmat de târlire (2004) concomitent cu fosfatare 100 kg/ha P2O5 respectiv cu „Eurobio – 26 P” fosfaţi măcinaţi pentru agricultura ecologică;
Grupa C, amendare calcică la 2/3 Ah (cca. 7 t/ha CaO – praf de var), în vara anului 1995, fertilizare NPK chimică (1996 – 1998) la fel ca în grupa B şi în continuare organică (2003 şi 2009 ) cu fosfatare;
Grupa D, erbicidare totală cu Roundup (glifosat) 5 L/ha în 150 litri apă în vara anului 1995, după două săptămâni amendare calcică identică cu varianta B, prelucrarea cu freza de pajişti la 10 – 12 cm adâncime şi după o săptămână semănat cu MSP – 2,5. În perioada 1996 – 1998 fertilizarea chimică NPK identică cu variantele B şi C şi în continuare organică prin târlire (2002 şi 2008 ) cu fosfatare.
Producţia medie de lapte la grupa A (martor), care a păşunat pe nardete neîmbunătăţite în medie pe 4 ani de măsurători a fost de 846 L/ha (923 litri în 1996, 847 în 1997, 844 în 1998 şi 771 în 1999) la o încărcare de cca. 1 vacă pe hectar şi o producţie a pajiştii de 1,2 t/ha SU (substanţă uscată) de slabă calitate.
După îmbunătăţirea nardetelor producţia de substanţă uscată şi de lapte este de câteva ori mai mare faţă de cele neîmbunătăţite (Tabelul 6.7.).
Tabelul 6.7. Date comparative medii privind productivitatea nardetelor subalpine
îmbunătăţite prin diferite metode 1995 – 2009 (Blana 1800 m alt.) Comparative data on the average productivity of subalpine nardions improved through various methods 1995 – 2009, (Blana 1800 m alt.)
Variante (lotul de vaci)
Producţia de substanţă uscată
(SU)
Producţia de lapte de vacă
t/ha % L/ha % A – martor, Nardus stricta 40 – 60 % 1,20 100 846 100 B – fertilizare NPK + târlire + P 3,38 282 3.248 384 C – amendare CaO + NPK + târlire + P 4,16 347 3.941 466 D – semănat + CaO + NPK + târlire + P 6,20 517 4.620 546
Astfel, producţia de substanţă uscată creşte de 3 – 5 ori şi cea de lapte la hectar de 4 – 5,5 ori pe o perioadă de 14 ani, ceea ce este extrem de economic. În plus, prin îmbunătăţirea nardetelor în porţiunile mecanizabile şi intensivizarea producţiei se măreşte şi se redistribuie încărcarea cu animale existentă, creându-se posibilităţi de protecţie şi conservare a biodiversităţii în zonele limitrofe, cu habitate mai fragile cum sunt stâncăriile, terenurile în pantă accentuată şi cele cu exces de umiditate.

289
Realizarea de produse animaliere ecologice pe pajiştile montane degradate nu este posibilă decât după ameliorarea lor. În ţările alpine (Elveţia, Franţa, Italia, Austria, Germania şi Slovenia), conversia la agricultura ecologică (biologică) se face după ce pe pajişti s-au aplicat decenii la rând doze mari de îngrăşăminte chimice (N 150 – 300 kg/ha), având un covor ierbos format din specii foarte bune furajere din flora spontană sau semănate, receptive la fertilizarea organică şi alte mijloace naturiste.
La noi există o tendinţă greşită de trecere directă de la pajişti degradate nefertilizate la agricultura „bio”, care cu certitudine nu va avea sorţi de izbândă, datorită solului foarte acid şi slab aprovizionat în elemente fertilizante şi covorului ierbos cu specii slab furajere cum este specia Nardus stricta (ţăpoşică) în zona montană mai umedă sau Botriochloa ischaemum (bărboasă) în zona de dealuri mai uscate erodate, de exemplu.
În aceste cazuri de pajişti degradate, obligatoriu este necesară îmbunătăţirea prin orice mijloace cunoscute a covorului ierbos (erbicidare, fertilizare chimică, amendare, supraînsămânţare, reînsămânţare) după care la 2 ani după încetarea fertilizării chimice sau al aplicării pesticidelor şi trecerea la fertilizare organică, se pot realiza pe pajişti produse animaliere „bio”.
Este iluzorie ideea că pajiştile noastre nefertilizate chimic de ani de zile pot fi deja considerate ca producătoare de furaje pentru hrănirea animalelor în sistem biologic. Cei care fac aceste afirmaţii şi le difuzează, nu se gândesc la performanţă şi aspectele economice care cântăresc greu în promovarea şi realizarea produselor animaliere ecologice.
Experienţa cu vaci de lapte din Platoul Bucegilor răspunde întrutotul acestui demers şi sistem enunţat mai înainte.
În primii 5 ani (1996 – 2000) producţia de SU şi de lapte s-a realizat în sistem convenţional (3 ani) şi de conversie la producţie ecologică (2 ani). În următorii 9 ani (2001 – 2009) se realizează deja o producţie ecologică, întrucât se fertilizează organic prin târlire şi se aplică fosforite măcinate, fiind prima experienţă cu animale cu produse „bio” pe păşune din ţara noastră (Tabelul 6.8.).
Producţia de lapte convenţional obţinută în primii 5 ani este mai mare decât producţia ecologică din următorii 9 ani. La varianta B (NPK), producţia ecologică ajunge la 82 %, la varianta C (CaO + NPK) la 81 % şi la varianta D (semănat + CaO + NPK) la 97 % faţă de producţia convenţională, ceea ce explică şi mai clar superioritatea pajiştii semănate în obţinerea laptelui ecologic faţă de covorul ierbos cu specii din flora spontană.
Din aceste date rezultă o extrem de mare diferenţă între ani şi variante. În cel mai secetos an, 2000, s-au obţinut cele mai slabe rezultate şi în primul an 1996 şi 2003 cele mai bune rezultate la producţia de iarbă şi lapte.
În ceea ce priveşte efectul amendării calcice asupra producţiei medii de lapte pe 14 ani în varianta C se constată că producţia este mai ridicată cu 21 % faţă de varianta B, cu tendinţă de menţinere a efectivului cel puţin 20 de ani, fapt necunoscut în literatura ştiinţifică de specialitate.

290
Tabelul 6.8. Producţia de substanţă uscată (SU) şi de lapte pe nardetele subalpine din
Masivul Bucegi, îmbunătăţite prin diferite metode convenţionale şi ecologice Dry matter (DM) and milk production on the subalpine nardions in Bucegi
Massif improved by various conventional and organic methods Fertilizări de
îmbunătăţire şi întreţinere
Anul Producţia de SU (t/ha) Producţia de lapte (L/ha) B C D B C D
Îmbunătăţire pedoameliorativă şi covor ierbos
1995
* * *
* Amendat
*
ErbicidatAmendatSemănat
* * *
* Amendat
*
Erbicidat Amendat Semănat
1. Producţie convenţională Fertilizare anuală, trei ani consecutiv cu 150 kg/ha N,50 kg/ha P2O5
şi 50 kg/haK2O
1996 6,95 7,61 8,68 4.180 5.690 7.020 1997 4,77 5,22 6,86 4.784 5.803 5.335 1998 1,84 2,15 2,58 3.356 4.272 4.673
Conversie la producţia ecologică
1999 1,56 2,09 2,68 4.216 4.364 4.368 2000 1,28 2,02 2,77 1.725 1.960 2.150
MEDIE convenţional * 3,28 3,82 4,71 3.652 4.418 4.709 Efect amendare * 100 116 144 100 121 129 Efect semănat * * 100 123 * 100 107
2. Producţie ecologică
Târlit 5 nopţi, o vacă pe 6 mp + 100 kg/ha P2O5
produs ecologic în diferiţi ani (T)
2001 3,22 4,62 5,54 2.070 2.460 2.690 2002 2,28 3,50 T 1.927 2.280 T 2003 4,12 T 10,65 2.111 T 5.899 2004 T 5,81 9,22 T 4.946 5.221 2005 6,00 6,73 6,38 3.056 3.350 4.658 2006 2,92 3,55 4,68 3.570 3.910 4.080 2007 3,02 3,67 8,00 3.689 4.072 4.976 2008 2,07 2,95 T 3.740 4.182 T 2009 3,97 T 6,34 3.798 T 4.372
MEDIE ecologic * 3,45 4,40 7,26 2.995 3.600 4.557 Efect amendare * 100 128 210 100 120 152 Efect semănat * * 100 165 * 100 127
Diferenţă 2-1, (+, -) * + 0,17 + 0,58 + 2,55 - 657 - 818 - 152 Efect relativ 2-1 (%) * 105 115 154 82 82 97
MEDIE GENERALĂ * 3,38 4,16 6,20 3.248 3.941 4.620 Efect amendare * 100 123 183 100 121 142 Efect semănat * * 100 149 * 100 117
La fel, efectul înfiinţării pajiştii din varianta D este cu 42 % mai mare în
producţia de lapte ecologic faţă de varianta C cu covor ierbos natural, efect care va dura minim 30 de ani.
Din cele prezentate rezultă clar necesitatea îmbunătăţirii covorului ierbos degradat în cazul de faţă a unei păşuni subalpine dominate de Nardus stricta şi

291
acceptarea unei perioade normale de conversie înainte de trecerea la producţie de lapte ecologic.
Efectul economic este evidenţiat prin producţiile de 3.000 – 4.600 litri pe hectarul de pajişte îmbunătăţită care este asigurator pentru cel puţin 20 – 30 de ani, indiferent de costurile măsurilor de îmbunătăţire prin fertilizare, amendare calcică şi înfiinţare pajişte semănată.
Facem în final o menţiune foarte importantă, care constă din împrejmuirea cu garduri fixe (stâlpi de beton cu 4 rânduri de sârmă ghimpată) a câmpului experimental de cca. 27 ha, în care din anul 1975 se păşunează raţional cu încărcătură adecvată de animale, la început de ovine şi în prezent de bovine.
Fără împrejmuirea suprafeţelor supuse îmbunătăţirii cu folosirea raţională a lor, rezultatele ar fi fost sortite din start eşecului, datorită imixtiunilor cu animale din afară şi deci al încărcării necorespunzătoare cu animale, care puteau degrada în final covorul ierbos al pajiştilor.
Vedere generală asupra Bazei de Cercetări Pajişti Montane Blana Bucegi 1800 m alt.

292
VII. GRADIENŢI DE PRODUCTIVITATE A PAJIŞTILOR PE ALTITUDINE Termenul de gradient, după DICŢIONARUL ENCICLOPEDIC din 1996,
provine din latină (gradiens = care înaintează) şi este folosit în matematică, fizică şi alte ştiinţe pentru exprimarea unor mărimi şi variaţii în funcţie de un anumit parametru, de obicei distanţă pe orizontală sau verticală (înălţime, adâncime) etc. În meteorologie, de exemplu, se utilizează gradientul baric care exprimă mărimea variabilă a presiunii barometrice, care se stabileşte între centru şi zona periferică a unui ciclon. În mediul marin se cunoaşte presiunea hidrostatică care creşte cu un gradient de cca 1 atmosferă cu fiecare 10 m adâncime (TÂRZIU, 1994). Dar cei mai utilizaţi gradienţi de până acum au fost cel termic, care indică scăderea medie a temperaturii atmosferei pe verticală pentru fiecare 100 m diferenţă de nivel cu 0,6°C, şi cel geotermic care indică creşterea medie a temperaturii spre interiorul pământului pentru fiecare 100 m adâncime, cu cca. 3°C în Europa şi cca. 2,5°C în America.
În paralel se foloseşte şi termenul de treaptă geotermică, care este distanţa exprimată în metri, cu care trebuie să se coboare pe verticală în adâncul pământului pentru a constata o creştere a temperaturii cu 1°C, distanţă ce variază între 5-150 m în funcţie de grosimea scoarţei terestre. Aceşti doi parametri, gradient şi treaptă, utilizaţi pentru acelaşi factor geotermic, diferă doar ca mod de exprimare. Astfel gradientul exprimă mărimea factorului la o echidistanţă fixă de 100 m, iar treapta exprimă atingerea unei unităţi de măsură a factorului studiat pe o distanţă variabilă.
Pentru condiţiile ţării noastre au fost stabiliţi gradientul termic vertical din troposferă, care este 0,5°... 0,6°C/100 m altitudine (STOENESCU, ŢÂŞTEA, 1962) şi gradientul hidric 40-50 mm/100 m (MARCU, 1983). O primă contribuţie la diversificarea gradienţilor climatici a avut-o N. TOPOR (1956), care a stabilit că întârzierea topirii zăpezii este de 4 zile (gradient disparitic) pentru fiecare 100 m altitudine.
Dispariţia zăpezii se face treptat în raport cu altitudinea: sub 200 m în jurul datei de 22 martie
200 - 500 m în jurul datei de 3 aprilie 500 - 1000 m în jurul datei de 23 aprilie
1000 - 1500 m în jurul datei de 12 mai 1500 - 2000 m în jurul datei de 1 iunie peste 2000 m în jurul datei de 21 iunie
Acest gradient este în funcţie de expoziţia versanţilor, el având cele mai mari valori în Munţii Apuseni pe versantul estic şi cele mai mici în Munţii Banatului (TOPOR, 1956, citat de GIURCĂNEANU, 1988). NIEDERMAIER (1962) constată că „pornirea în vegetaţie a întârziat în medie pentru 100 m diferenţă de altitudine cu 2,69 zile, sau – cu alte cuvinte a înaintat zilnic de la poalele munţilor cercetaţi spre vârful Cindrelului, în medie cu 37,2 m diferenţă de altitudine”. Pentru Masivul Bucegi, STOENESCU (1951) arată că … „primăvara

293
înaintează în înălţime cu 31 m pe zi”. Din aceste date putem observa că în Munţii Cindrelului situaţi mai spre vest se desprimăvărează în medie mai repede cu cca 6 m pe zi decât în Bucegi. Având în vedere modul sintetic şi evolutiv de exprimare al acestor factori (presiune atmosferică, temperatură aer, precipitaţii, topirea zăpezii, desprimăvărare, etc.) s-a pus problema dacă acest model poate fi extins şi la alţi factori climatici sau de altă natură specifici zonei montane, unde sunt cunoscute variaţiile mari ale acestora pe altitudine. Mai recent s-a analizat evoluţia pe altitudine a duratei intervalelor cu temperaturi medii zilnice mai mari sau egale cu 10°C, care după datele din CLIMATOLOGIA ROMÂNIEI (1966) sunt de -7,5 zile /100 m altitudine, parametru identic cu durata optimă a sezonului de păşunat în zona montană (MARUŞCA, 1992). În continuare au fost luaţi în studiu numeroşi alţi factori climatici, pedologici, productivitatea vegetaţiei, activitatea umană în aer liber, confortul termic în locuinţe şi multe altele care se prezintă în lucrarea de faţă. Exprimarea prin gradienţi a factorilor naturali şi socio-economici din zona montană prezintă numeroase avantaje din care enumerăm: - sintetizarea unui volum imens de date din diferite domenii ale cunoaşterii pentru un nivel sau niveluri altitudinale. - posibilitatea determinării cu o oarecare exactitate a datelor lipsă la un anumit nivel sau ecart altitudinal. - uşurinţa cu care se stabilesc legăturile între diferiţi factori şi explicarea unor fenomene şi interacţiuni mai complexe, cu evoluţia lor pe altitudine. - crearea unui limbaj mai accesibil de comunicare între specialişti din diferite domenii de activitate, fiind util şi în procesul instructiv - formativ. - fundamentarea pe baze ştiinţifice a legislaţiei montane cu regulamentele ei de aplicare specifice şi multe altele. Această nouă direcţie de sintetizare a numeroaselor acumulări de cunoştinţe intra- inter- şi trans-disciplinare pentru zona montană este la început de drum, existând multiple posibilităţi de dezvoltare şi îmbunătăţire a ei în continuare.
7.1. GRADIENŢI CLIMATICI Din cele prezentate mai înainte, primii gradienţi studiaţi au fost cei climatici cu deosebire temperatura medie anuală a aerului şi precipitaţiile atmosferice. În continuare se prezintă încercările efectuate asupra altor factori climatici, cu evoluţia lor pe altitudine.
7.1.1. Temperatura aerului şi solului Datele privind temperatura aerului din ţara noastră, prezentate în diferite lucrări de specialitate pentru perioadele 1896-1955 (STOENESCU, TÂŞTEA, 1962); 1928-1945 (STOENESCU 1951); 1946-1969 (TÂŞTEA, NEACŞA şi colab., 1974); 1961-1980
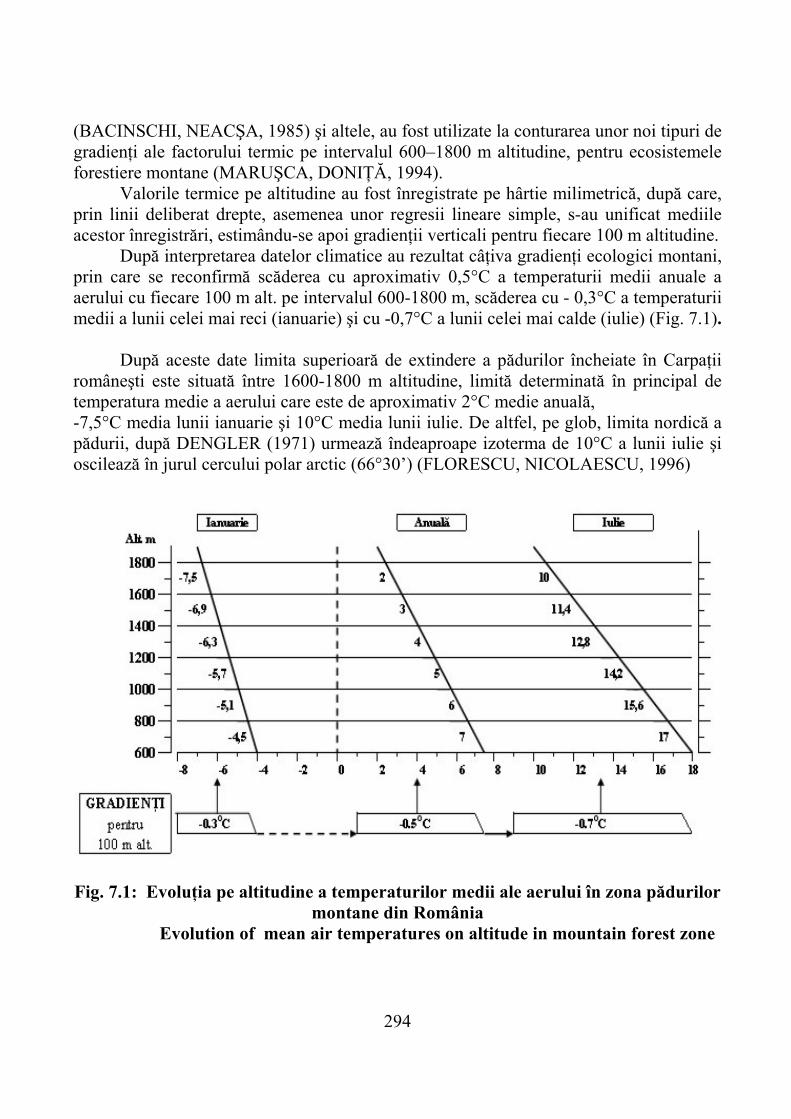
294
(BACINSCHI, NEACŞA, 1985) şi altele, au fost utilizate la conturarea unor noi tipuri de gradienţi ale factorului termic pe intervalul 600–1800 m altitudine, pentru ecosistemele forestiere montane (MARUŞCA, DONIŢĂ, 1994). Valorile termice pe altitudine au fost înregistrate pe hârtie milimetrică, după care, prin linii deliberat drepte, asemenea unor regresii lineare simple, s-au unificat mediile acestor înregistrări, estimându-se apoi gradienţii verticali pentru fiecare 100 m altitudine. După interpretarea datelor climatice au rezultat câţiva gradienţi ecologici montani, prin care se reconfirmă scăderea cu aproximativ 0,5°C a temperaturii medii anuale a aerului cu fiecare 100 m alt. pe intervalul 600-1800 m, scăderea cu - 0,3°C a temperaturii medii a lunii celei mai reci (ianuarie) şi cu -0,7°C a lunii celei mai calde (iulie) (Fig. 7.1).
După aceste date limita superioară de extindere a pădurilor încheiate în Carpaţii româneşti este situată între 1600-1800 m altitudine, limită determinată în principal de temperatura medie a aerului care este de aproximativ 2°C medie anuală, -7,5°C media lunii ianuarie şi 10°C media lunii iulie. De altfel, pe glob, limita nordică a pădurii, după DENGLER (1971) urmează îndeaproape izoterma de 10°C a lunii iulie şi oscilează în jurul cercului polar arctic (66°30’) (FLORESCU, NICOLAESCU, 1996)
Fig. 7.1: Evoluţia pe altitudine a temperaturilor medii ale aerului în zona pădurilor montane din România
Evolution of mean air temperatures on altitude in mountain forest zone
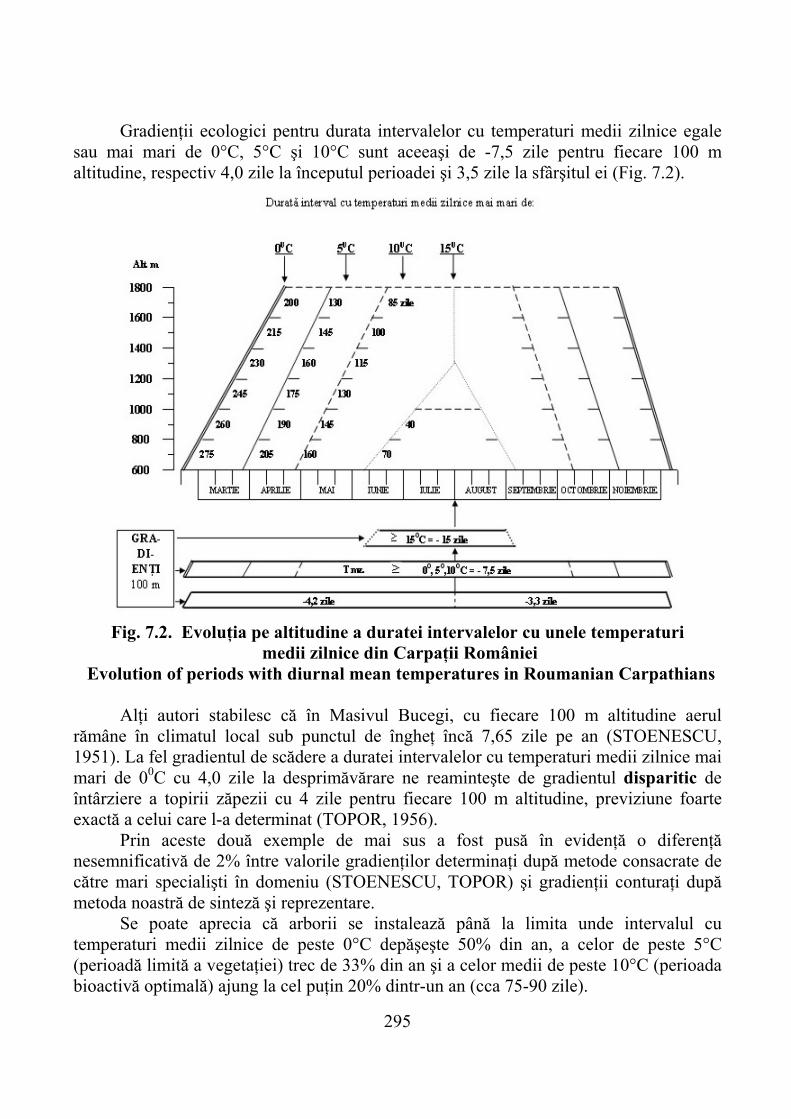
295
Gradienţii ecologici pentru durata intervalelor cu temperaturi medii zilnice egale sau mai mari de 0°C, 5°C şi 10°C sunt aceeaşi de -7,5 zile pentru fiecare 100 m altitudine, respectiv 4,0 zile la începutul perioadei şi 3,5 zile la sfârşitul ei (Fig. 7.2).
Fig. 7.2. Evoluţia pe altitudine a duratei intervalelor cu unele temperaturi
medii zilnice din Carpaţii României Evolution of periods with diurnal mean temperatures in Roumanian Carpathians Alţi autori stabilesc că în Masivul Bucegi, cu fiecare 100 m altitudine aerul rămâne în climatul local sub punctul de îngheţ încă 7,65 zile pe an (STOENESCU, 1951). La fel gradientul de scădere a duratei intervalelor cu temperaturi medii zilnice mai mari de 00C cu 4,0 zile la desprimăvărare ne reaminteşte de gradientul disparitic de întârziere a topirii zăpezii cu 4 zile pentru fiecare 100 m altitudine, previziune foarte exactă a celui care l-a determinat (TOPOR, 1956). Prin aceste două exemple de mai sus a fost pusă în evidenţă o diferenţă nesemnificativă de 2% între valorile gradienţilor determinaţi după metode consacrate de către mari specialişti în domeniu (STOENESCU, TOPOR) şi gradienţii conturaţi după metoda noastră de sinteză şi reprezentare. Se poate aprecia că arborii se instalează până la limita unde intervalul cu temperaturi medii zilnice de peste 0°C depăşeşte 50% din an, a celor de peste 5°C (perioadă limită a vegetaţiei) trec de 33% din an şi a celor medii de peste 10°C (perioada bioactivă optimală) ajung la cel puţin 20% dintr-un an (cca 75-90 zile).
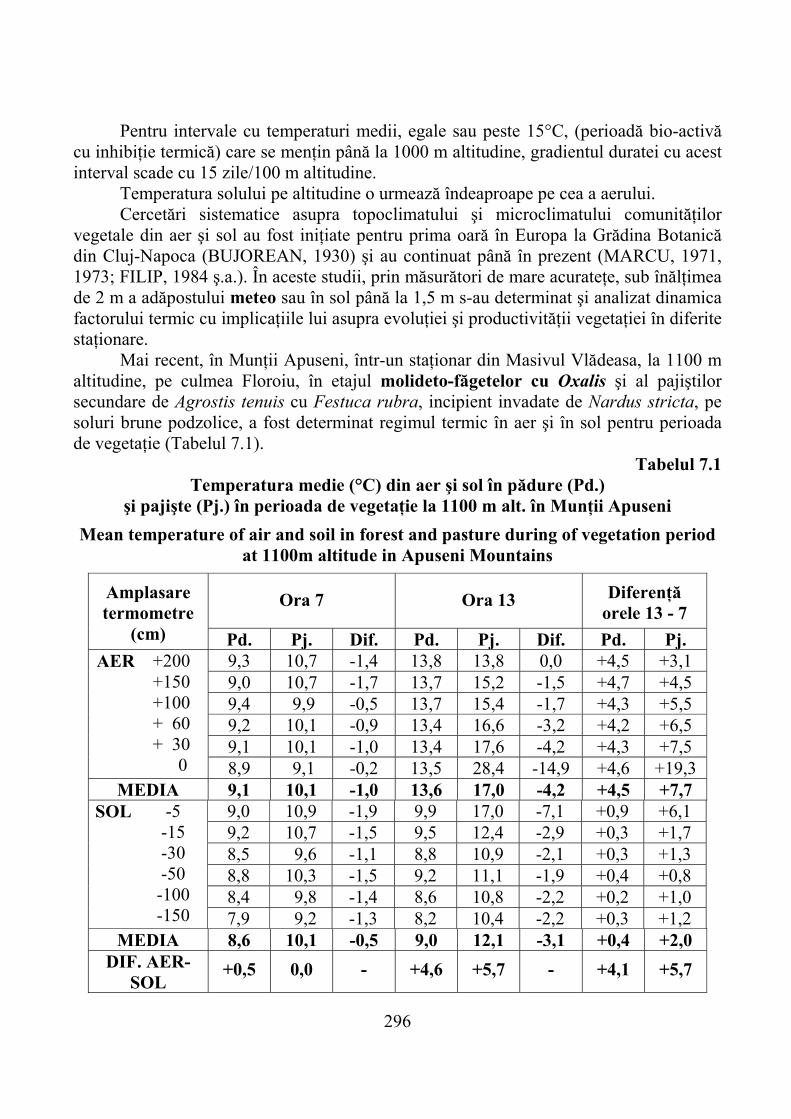
296
Pentru intervale cu temperaturi medii, egale sau peste 15°C, (perioadă bio-activă cu inhibiţie termică) care se menţin până la 1000 m altitudine, gradientul duratei cu acest interval scade cu 15 zile/100 m altitudine. Temperatura solului pe altitudine o urmează îndeaproape pe cea a aerului. Cercetări sistematice asupra topoclimatului şi microclimatului comunităţilor vegetale din aer şi sol au fost iniţiate pentru prima oară în Europa la Grădina Botanică din Cluj-Napoca (BUJOREAN, 1930) şi au continuat până în prezent (MARCU, 1971, 1973; FILIP, 1984 ş.a.). În aceste studii, prin măsurători de mare acurateţe, sub înălţimea de 2 m a adăpostului meteo sau în sol până la 1,5 m s-au determinat şi analizat dinamica factorului termic cu implicaţiile lui asupra evoluţiei şi productivităţii vegetaţiei în diferite staţionare. Mai recent, în Munţii Apuseni, într-un staţionar din Masivul Vlădeasa, la 1100 m altitudine, pe culmea Floroiu, în etajul molideto-făgetelor cu Oxalis şi al pajiştilor secundare de Agrostis tenuis cu Festuca rubra, incipient invadate de Nardus stricta, pe soluri brune podzolice, a fost determinat regimul termic în aer şi în sol pentru perioada de vegetaţie (Tabelul 7.1).
Tabelul 7.1 Temperatura medie (°C) din aer şi sol în pădure (Pd.)
şi pajişte (Pj.) în perioada de vegetaţie la 1100 m alt. în Munţii Apuseni
Mean temperature of air and soil in forest and pasture during of vegetation period at 1100m altitude in Apuseni Mountains
Amplasare termometre
(cm)
Ora 7 Ora 13 Diferenţă orele 13 - 7
Pd. Pj. Dif. Pd. Pj. Dif. Pd. Pj.AER +200
+150 +100 + 60 + 30 0
9,3 10,7 -1,4 13,8 13,8 0,0 +4,5 +3,1 9,0 10,7 -1,7 13,7 15,2 -1,5 +4,7 +4,59,4 9,9 -0,5 13,7 15,4 -1,7 +4,3 +5,5 9,2 10,1 -0,9 13,4 16,6 -3,2 +4,2 +6,5 9,1 10,1 -1,0 13,4 17,6 -4,2 +4,3 +7,58,9 9,1 -0,2 13,5 28,4 -14,9 +4,6 +19,3
MEDIA 9,1 10,1 -1,0 13,6 17,0 -4,2 +4,5 +7,7 SOL -5
-15 -30 -50 -100 -150
9,0 10,9 -1,9 9,9 17,0 -7,1 +0,9 +6,1 9,2 10,7 -1,5 9,5 12,4 -2,9 +0,3 +1,7 8,5 9,6 -1,1 8,8 10,9 -2,1 +0,3 +1,38,8 10,3 -1,5 9,2 11,1 -1,9 +0,4 +0,8 8,4 9,8 -1,4 8,6 10,8 -2,2 +0,2 +1,0 7,9 9,2 -1,3 8,2 10,4 -2,2 +0,3 +1,2
MEDIA 8,6 10,1 -0,5 9,0 12,1 -3,1 +0,4 +2,0 DIF. AER-
SOL +0,5 0,0 - +4,6 +5,7 - +4,1 +5,7

297
Aceste date au fost înregistrate în perioada mai - septembrie, în 3 etape a 10-12 zile, cu climat mediu, pe timp însorit, înnourat, ploios, atmosferă liniştită şi vânt, etc. Au fost efectuate determinări concomitente (în paralel), în pădure şi pajişte din 20 în 20 de minute asupra temperaturii de la suprafaţa solului până la 2 m înălţime şi în sol până la 1,5 m. Determinările pentru temperatura aerului s-au efectuat cu termometre obişnuite, fixate pe o stadie, având rezervoarele cu mercur umbrite cu conuri de hârtie, iar termometrele pentru sol au fost prevăzute cu carcasă metalică. Rezultatele din tabel pun în evidenţă diferenţele nete dintre temperaturile din pădure şi pajişte, la fel între cele din sol şi aer. Astfel, la 7 dimineaţa, diferenţa medie între temperatura din pădure şi pajişte este de 1°C în aer şi 0,5°C în sol, crescând la ora 1300, până la 4,2°C în aer şi 3,1°C în sol pe pajişte faţă de pădure. Cu ajutorul unor microgradienţi termici a fost posibilă conturarea evoluţiei temperaturii din 10 în 10 cm înălţime în aer şi adâncime în sol (MARUŞCA şi FILIP, 1995) (Fig. 7.3).
Schimbările cele mai mari ale microgradienţilor termici s-au înregistrat pe intervale 0–40 cm în aer şi sol îndeosebi pe pajişte unde sunt de -1,6°/10 cm (17,2-14,0°C) în aer şi -2,8°C (16,0-10,4°C) în sol. Sub pădure temperaturile în aer şi sol sunt mai constante, înregistrându-se valori subunitare pentru microgradienţi, confirmând rolul protector al vegetaţiei lemnoase în comparaţie cu alte moduri de folosinţă al terenurilor. Regimul termic al aerului (0-200 cm înălţime) şi al solului (0-150 cm adâncime) din vegetaţia lemnoasă este mai scăzut şi mult mai constant în comparaţie cu cel din vegetaţia ierboasă în perioada de vegetaţie (mai-septembrie) la nivelul de 1100 m altitudine. Microgradienţii termici medii din aer sunt absenţi în pădure pe intervalul 50-200 cm înălţime de la sol (11,50C) şi au o uşoară scădere pe pajişte de -0,04°C/10 cm (13,1-12,70C) la aceeaşi înălţime. În sol, pe adâncimea 50-150 cm microgradienţii sunt egali, respectiv de -0,08°C/10 cm, dar cu limitele medii de 8,7-8,30 sub pădure şi 10,3-9,9 0C sub pajişte. Aceste date atestă încă o dată, dacă mai este nevoie, rolul termoregulator al pădurilor în climatul zonei montane.
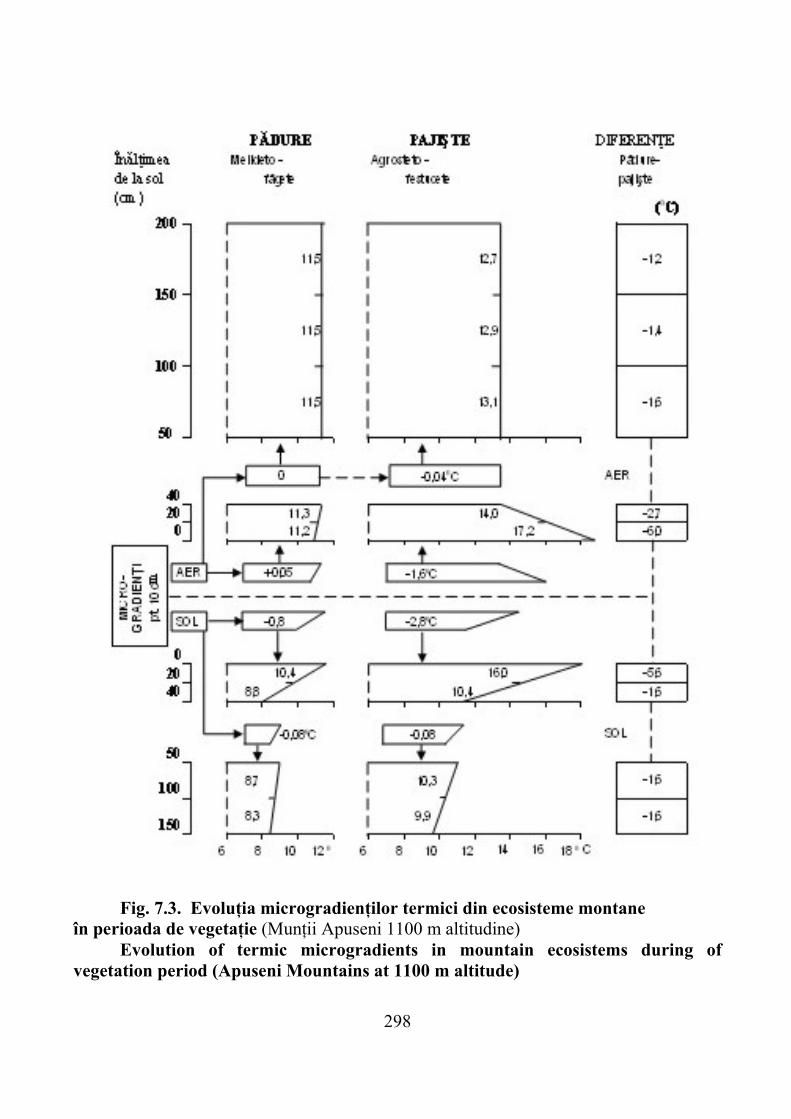
298
Fig. 7.3. Evoluţia microgradienţilor termici din ecosisteme montane în perioada de vegetaţie (Munţii Apuseni 1100 m altitudine) Evolution of termic microgradients in mountain ecosistems during of vegetation period (Apuseni Mountains at 1100 m altitude)
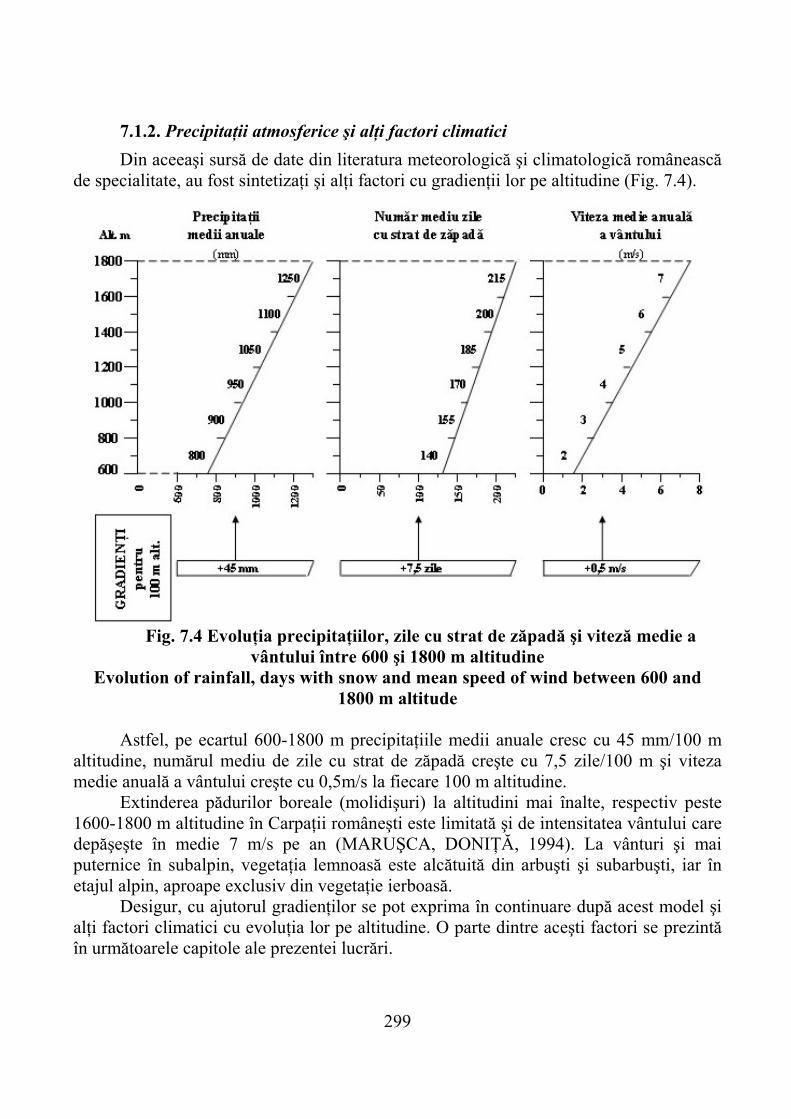
299
7.1.2. Precipitaţii atmosferice şi alţi factori climatici
Din aceeaşi sursă de date din literatura meteorologică şi climatologică românească de specialitate, au fost sintetizaţi şi alţi factori cu gradienţii lor pe altitudine (Fig. 7.4).
Fig. 7.4 Evoluţia precipitaţiilor, zile cu strat de zăpadă şi viteză medie a
vântului între 600 şi 1800 m altitudine Evolution of rainfall, days with snow and mean speed of wind between 600 and
1800 m altitude
Astfel, pe ecartul 600-1800 m precipitaţiile medii anuale cresc cu 45 mm/100 m altitudine, numărul mediu de zile cu strat de zăpadă creşte cu 7,5 zile/100 m şi viteza medie anuală a vântului creşte cu 0,5m/s la fiecare 100 m altitudine. Extinderea pădurilor boreale (molidişuri) la altitudini mai înalte, respectiv peste 1600-1800 m altitudine în Carpaţii româneşti este limitată şi de intensitatea vântului care depăşeşte în medie 7 m/s pe an (MARUŞCA, DONIŢĂ, 1994). La vânturi şi mai puternice în subalpin, vegetaţia lemnoasă este alcătuită din arbuşti şi subarbuşti, iar în etajul alpin, aproape exclusiv din vegetaţie ierboasă. Desigur, cu ajutorul gradienţilor se pot exprima în continuare după acest model şi alţi factori climatici cu evoluţia lor pe altitudine. O parte dintre aceşti factori se prezintă în următoarele capitole ale prezentei lucrări.

300
7.2. DISTRIBUŢIA ALTITUDINALĂ A ECOSISTEMELOR FORESTIERE ŞI TIPURI DE PAJIŞTI În lucrările de sinteză asupra vegetaţiei lemnoase şi ierboase din Carpaţii României s-a determinat conceptul de etajare verticală pe altitudine faţă de zonalitatea orizontală pe latitudine, caracteristică vegetaţiei planare şi colinare (ANGHEL, DONIŢĂ, 1980; BĂRBULESCU, MOTCĂ, 1983, 1987; DONIŢĂ, IVAN, 1985 şi alţii). Începând cu BRÂNZĂ (1879) până în prezent (IVAN, 1979; VELCEA, BADEA, 1983) sunt redate mai mult arealul sau limitele inferioare şi superioare de răspândire altitudinală a unor specii, fitocenoze, ecosisteme, etc., fără menţionarea suprafeţelor sau distribuţia lor procentuală pe nivele altitudinale, obiectiv propus a se lămuri pe cât posibil în continuare. Acumularea de date noi privind tipurile de ecosisteme forestiere cu înveliş biocenetic natural (DONIŢĂ şi colab., 1990) şi a tipurilor de pajişti anturale (ŢUCRA şi colab., 1987) au permis elaborarea unui sistem mai sugestiv de reprezentare paralelă a vegetaţiei primare şi secundare precum şi a relaţiei pădure-pajişte la diferite nivele altitudinale. Pentru fitocenozele şi tipurile de pajişti naturale dispunem de date incomplete sub aspectul suprafeţelor şi limitelor altitudinale de răspândire a lor. Au fost luate în studiu principalele fitocenoze montane de pajişti naturale (PUŞCARU-SOROCEANU, 1963) şi cca 20 tipuri de pajişti situate între 600-1800 m altitudine (ŢUCRA, 1987) la care se cunosc în linii generale suprafeţele şi ecartul altitudinal. În completare au fost utilizate suprafeţele de pajişti între diferite limite altitudinale (TEACI, 1980). După această metodologie de lucru şi surse de date din literatura noastră de specialitate, a fost posibilă întocmirea unei sinteze grafice în care se prezintă concomitent răspândirea altitudinală, distribuţia la nivel altitudinal şi structură spaţială (suprafaţa) a pădurilor şi pajiştilor naturale montane (MARUŞCA, DONIŢĂ, 1994a) (Fig.7.5.).
Prezentarea grafică faţă în faţă a vegetaţiei primare şi secundare, confirmă şi explicitează datele obţinute de ANGHEL şi DONIŢĂ (1980) prin care, în etajul pădurilor de molid se instalează pajişti dominante de Festuca rubra ssp. commutata (F. nigrescens), în subetajul pădurilor de fag şi de amestec de fag cu răşinoase domină Agrostis tenuis (A. capillaris) cu Festuca rubra, iar în subetajul pădurilor de gorun sau amestec de gorun, vegetaţia secundară este dominată de Festuca rupicola cu Agrostis tenuis şi alte specii. Acest grafic se poate considera un abac simplu prin care se determină pe nivele altitudinale sau etaje de vegetaţie mari (subalpin, boreal, nemoral) corespondenţa între tipurile de ecosisteme forestiere şi tipurile de pajişti care se succed după înlăturarea prin defrişare a vegetaţiei lemnoase.

301
Fig.7.5. Răspândirea, structura şi distribuţia pe nivele altitudinale a pădurilor şi
pajiştilor naturale din Carpaţii României (MARUSCA, 2001) Spreading, structure and distribution of forests and natural grasslands on
altitudinal levels in Roumanian Carpathians
Datele obţinute au permis stabilirea raportului pădure-pajişte la diferite nivele altitudinale (Tabelul 7.2).

302
Tabelul 7.2 Raportul între păduri şi pajişti naturale la diferite altitudini
Share between forests and natural grasslands at different altitudes
Altitudine Suprafaţa de păduri
Suprafaţa de pajişti
Raportul pădure/ pajişte
Direcţii impact
antropic
Cauzele despăduririi
mii ha % mii ha %1600-1800 1400-1600 1200-1400 1000-1200 800-1000 600- 800
90310660
1130700310
2,8 9,7
20,6 35,3 21,9 9,7
70105120160500645
4,46,67,5
10,031,240,3
1:0,78*) 1:0,34 1:0,18 1:0,14 1:0,71 1:2,08
păduri
Extindere păşuni animale Gospodării Fâneţe Arabil, etc.
*) Raport influenţat şi de limita superioară naturală de răspândire a pădurilor 7.3. PRODUCTIVITATEA PRIMARĂ A ECOSISTEMELOR MONTANE PE NIVELURI ALTITUDINALE
În extrasul privind principalele tipuri de ecosisteme forestiere pe lângă ecart altitudinal, suprafaţă şi biometrie se prezintă în final volumul mediu (m3/ha) de masă lemnoasă la vârsta de 100 de ani, date care s-au utilizat la calculul productivităţii lor (DONIŢĂ, CHIRIŢĂ, STĂNESCU şi alţi, 1990). La o primă analiză, productivitatea medie a ecosistemelor forestiere montane este de aproximativ 450 m3/ha, cele mai productive fiind brădetele (500 m3/ha) care sunt şi cele mai complexe având pe lângă brad, în diferite proporţii, molid, fag şi alte specii (Tabelul 7.3).
Tabelul 7.3 Date generale privind productivitatea ecosistemelor forestiere montane
din România General date regarding to the productivity of mountain forestry ecosistems in
Romania Masă lemnoasă la 100 ani
Tipul de formaţie ecosistemică
Nr. total tipuri
Limite altitudinale
(m)
Suprafaţa Volum mii ha % m3/ha %
Molidişuri Brădete Făgete montane Făgete colinare Gorunete Zăvoaie
14 20 13 11 5 2
800 -1850 600 -1450 700 -1500
(600)-1000 (600)- 750
(600)-1200
88510201040210
45+
28 32 33 6 1 +
450 500 425 370 270
*
100111948260*
TOTAL 65 (600)-1850 3200 100 450 100
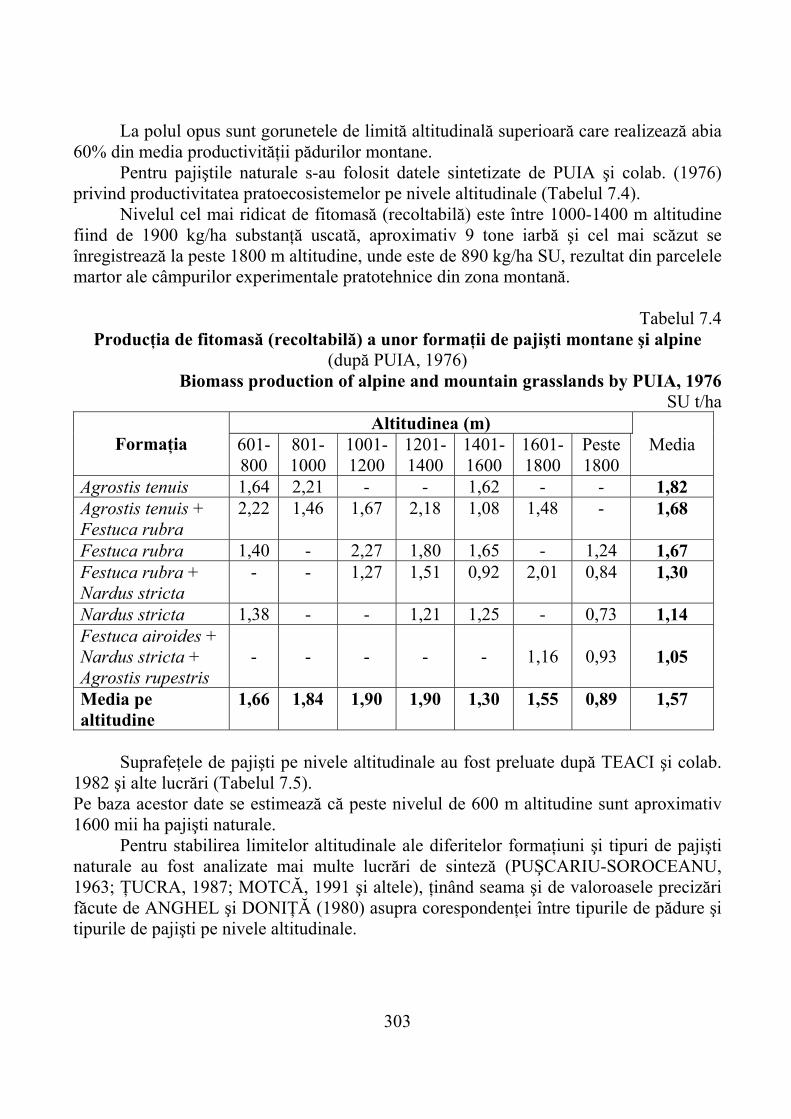
303
La polul opus sunt gorunetele de limită altitudinală superioară care realizează abia 60% din media productivităţii pădurilor montane.
Pentru pajiştile naturale s-au folosit datele sintetizate de PUIA şi colab. (1976) privind productivitatea pratoecosistemelor pe nivele altitudinale (Tabelul 7.4).
Nivelul cel mai ridicat de fitomasă (recoltabilă) este între 1000-1400 m altitudine fiind de 1900 kg/ha substanţă uscată, aproximativ 9 tone iarbă şi cel mai scăzut se înregistrează la peste 1800 m altitudine, unde este de 890 kg/ha SU, rezultat din parcelele martor ale câmpurilor experimentale pratotehnice din zona montană.
Tabelul 7.4
Producţia de fitomasă (recoltabilă) a unor formaţii de pajişti montane şi alpine (după PUIA, 1976)
Biomass production of alpine and mountain grasslands by PUIA, 1976 SU t/ha
Formaţia
Altitudinea (m)601-800
801-1000
1001-1200
1201-1400
1401-1600
1601-1800
Peste 1800
Media
Agrostis tenuis 1,64 2,21 - - 1,62 - - 1,82 Agrostis tenuis + Festuca rubra
2,22 1,46 1,67 2,18 1,08 1,48 - 1,68
Festuca rubra 1,40 - 2,27 1,80 1,65 - 1,24 1,67 Festuca rubra + Nardus stricta
- - 1,27 1,51 0,92 2,01 0,84 1,30
Nardus stricta 1,38 - - 1,21 1,25 - 0,73 1,14 Festuca airoides + Nardus stricta + Agrostis rupestris
-
-
-
-
-
1,16
0,93
1,05
Media pe altitudine
1,66 1,84 1,90 1,90 1,30 1,55 0,89 1,57
Suprafeţele de pajişti pe nivele altitudinale au fost preluate după TEACI şi colab.
1982 şi alte lucrări (Tabelul 7.5). Pe baza acestor date se estimează că peste nivelul de 600 m altitudine sunt aproximativ 1600 mii ha pajişti naturale.
Pentru stabilirea limitelor altitudinale ale diferitelor formaţiuni şi tipuri de pajişti naturale au fost analizate mai multe lucrări de sinteză (PUŞCARIU-SOROCEANU, 1963; ŢUCRA, 1987; MOTCĂ, 1991 şi altele), ţinând seama şi de valoroasele precizări făcute de ANGHEL şi DONIŢĂ (1980) asupra corespondenţei între tipurile de pădure şi tipurile de pajişti pe nivele altitudinale.

304
Tabelul 7.5 Repartiţia pajiştilor pe altitudini absolute
(după TEACI, 1980) Disposition of the grasslands on absolute altitudes by TEACI, 1980
mii hectare
Altitudine (m) Păşuni Fâneţe Total %
peste 2000 1501-2000 1001-1500 751-1000 501- 750 251- 500 101- 250 sub 100
44 154 227 331 488 819 606 384
2 21 131 291 376 387 168 38
46 176 358 602 863
1206 774 422
1,0 7,4 8,1
13,6 19,4 27,1 17,4 9,5
TOTAL 3033 1413 4446 100,0

305
Având la dispoziţie acest volum mare de date din diferite domenii după aplicarea metodelor specifice de determinare a distribuţiei ecosistemelor forestiere şi praticole s-au conturat gradienţii productivităţii pădurilor şi pajiştilor naturale (MARUŞCA, DONIŢĂ, 1994) (Fig. 7.6.).
Fig. 7.6. Evoluţia pe altitudine a distribuţiei şi productivităţii pădurilor şi pajiştilor naturale (după MARUSCA şi DONIŢĂ, 1994)
Evolution of distribution and productivity of forests and natural grasslands by MARUSCA and DONITA, 1994
Productivitatea ecosistemelor forestiere montane la vârsta de 100 ani, de la limita de 600 m altitudine, creşte în medie cu 15 m3 pe ha (430-490 m3) volum masă lemnoasă până la 1000-1200 m altitudine, după care scade cu 30 m3/ha (490-310 m3) pentru fiecare 100 m altitudine, până la limita superioară de răspândire a lor (cca 1800 m altitudine).
Productivitatea protoecosistemelor creşte în medie cu 0,5 t/ha/an (7-9 t) masă verde până la acelaş nivel de 1000-1200 m alt. după care scade cu 1,0 t/ha/an (9-3 t) pentru fiecare 100 m până la acelaş nivel de 1800 m altitudine.

306
Productivitatea maximă a vegetaţiei în Carpaţii României în jurul altitudinii de 1000 m, a fost prezentată şi în alte lucrări de specialitate (ANGHEL, DONIŢĂ, 1980; DONIŢĂ; IVAN, 1982; s.a.), în lucrarea de faţă se aduc câteva elemente care completează şi întăresc această concluzie.
Valorile maxime ale productivităţii pădurilor şi pajiştilor înregistrate între 1000-1200 m altitudine sunt condiţionate climatic prin 4-60C temperatură medie anuală, 13-150C, temperatura medie a lunii celei mai calde, 150-180 zile cu temperaturi medii de peste 50C şi 110-140 zile durată cu temperaturi peste 100C, la precipitaţii anuale de 900-1050 mm pe soluri cu o reacţie de 5,0-5,5 pH în apă şi 35-45% grad de saturaţie în baze (V%), unde se întâlnesc 53% brădete, 30% făgete montane, 17% molidişuri, după a căror defrişare au rezultat şi s-au menţinut 65% agrosteto-festucete, 15% festucete şi 20% nardete din suprafaţa ecosistemelor situate la acest nivel altitudinal. Din această înşiruire de condiţii climatice, pedochimice, învelişuri vegetale cu productivitatea lor, rezidă importanţa aparte care merită să fie acordată gradienticii montane ca metodologie şi instrument de caracterizare complexă şi mai explicită a unui interval altitudinal din spaţiul carpatic românesc. Reflectarea influenţei factorilor ecologici montani prezentaţi mai înainte, împletiţii cu tehnologiile aplicate de către om, este redată în final de performanţele de productivitate a aminalelor domestice în special a celor care valorifică pajiştile prin păşunat. Experimentările privind sporirea producţiei pajiştilor prin fertilizare cu îngrăşăminte chimice sau organice începute cu mai bine de 75 de ani în urmă (SAFTA, 1936) au continuat în paralel cu exprimarea randamentului în spor greutate vie la tineretul taurin sau lapte de vacă (MARUŞCA, 1974, 2000, 2008) până în zilele noastre, acumulându-se un volum mare de date îndeosebi în Carpaţii Meridionali mai accesibili şi intens cercetaţi din toate punctele de vedere. 7.4. GRADIENŢI DE PRODUCTIVITATE A PAJIŞTILOR MONTANE ÎMBUNĂTĂŢITE
7.4.1. Munţii Cindrel
7.4.1.1. Evoluţia pe altitudine a unor caracteristici pedoclimatice Munţii Cindrelului, denumiţi şi Munţii Cibinului sau ai Mărginimii Sibiului sunt o foarte veche vatră de vieţuire păstorească de unde au difuzat în timp, pe întreg lanţul carpatic şi alte meleaguri creşterea animalelor, în special al oilor.
Aceşti munţi cu culmi domoale care au permis un grad ridicat de umanizare, au atras de-a lungul vremii pe mulţi specialişti din diferite domenii de activitate, care au dezlegat o bună parte din complexitatea tainelor care le ascunde clima, solul, vegetaţia primară şi secundară cu productivitatea animalelor şi altele, în scopul unei mai bune gospodăriri a resurselor montane.
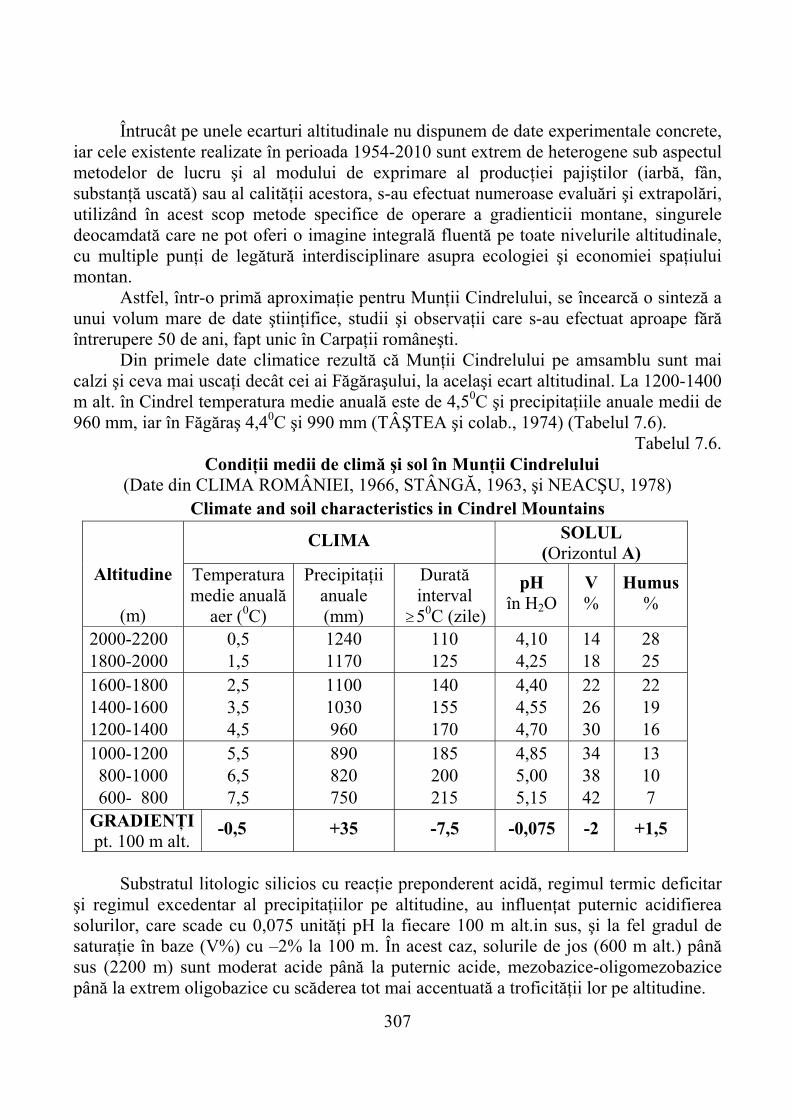
307
Întrucât pe unele ecarturi altitudinale nu dispunem de date experimentale concrete, iar cele existente realizate în perioada 1954-2010 sunt extrem de heterogene sub aspectul metodelor de lucru şi al modului de exprimare al producţiei pajiştilor (iarbă, fân, substanţă uscată) sau al calităţii acestora, s-au efectuat numeroase evaluări şi extrapolări, utilizând în acest scop metode specifice de operare a gradienticii montane, singurele deocamdată care ne pot oferi o imagine integrală fluentă pe toate nivelurile altitudinale, cu multiple punţi de legătură interdisciplinare asupra ecologiei şi economiei spaţiului montan. Astfel, într-o primă aproximaţie pentru Munţii Cindrelului, se încearcă o sinteză a unui volum mare de date ştiinţifice, studii şi observaţii care s-au efectuat aproape fără întrerupere 50 de ani, fapt unic în Carpaţii româneşti. Din primele date climatice rezultă că Munţii Cindrelului pe amsamblu sunt mai calzi şi ceva mai uscaţi decât cei ai Făgăraşului, la acelaşi ecart altitudinal. La 1200-1400 m alt. în Cindrel temperatura medie anuală este de 4,50C şi precipitaţiile anuale medii de 960 mm, iar în Făgăraş 4,40C şi 990 mm (TÂŞTEA şi colab., 1974) (Tabelul 7.6).
Tabelul 7.6. Condiţii medii de climă şi sol în Munţii Cindrelului
(Date din CLIMA ROMÂNIEI, 1966, STÂNGĂ, 1963, şi NEACŞU, 1978) Climate and soil characteristics in Cindrel Mountains
Altitudine
(m)
CLIMA SOLUL (Orizontul A)
Temperatura medie anuală
aer (0C)
Precipitaţii anuale (mm)
Durată interval
50C (zile)
pH în H2O
V %
Humus %
2000-2200 1800-2000
0,5 1,5
1240 1170
110 125
4,10 4,25
14 18
28 25
1600-1800 1400-1600 1200-1400
2,5 3,5 4,5
1100 1030 960
140 155 170
4,40 4,55 4,70
22 26 30
22 19 16
1000-1200 800-1000 600- 800
5,5 6,5 7,5
890 820 750
185 200 215
4,85 5,00 5,15
34 38 42
13 10 7
GRADIENŢI pt. 100 m alt.
-0,5 +35 -7,5 -0,075 -2 +1,5
Substratul litologic silicios cu reacţie preponderent acidă, regimul termic deficitar şi regimul excedentar al precipitaţiilor pe altitudine, au influenţat puternic acidifierea solurilor, care scade cu 0,075 unităţi pH la fiecare 100 m alt.in sus, şi la fel gradul de saturaţie în baze (V%) cu –2% la 100 m. În acest caz, solurile de jos (600 m alt.) până sus (2200 m) sunt moderat acide până la puternic acide, mezobazice-oligomezobazice până la extrem oligobazice cu scăderea tot mai accentuată a troficităţii lor pe altitudine.

308
Ca urmare a acestor procese pedogenetice nefavorabile creşterii plantelor, se acumulează progresiv cu altitudinea o cantitate tot mai mare de humus în solul pajiştilor, respectiv cu +1,5%/100 m, de la 7% (600-800 m) până la 28% (2000-2200 m altitudine) de la mull până la moder şi humus brut foarte acid.
7.4.1.2. Etajarea vegetaţiei primare şi secundare Asupra paralelismului şi în parte a echivalenţei între vegetaţia lemnoasă (primară)
şi ierboasă (secundară şi primară) sunt puţine lucrări în literatura noastră de specialitate (ANGHEL, DONIŢĂ, 1980). Pentru teritoriul carpatic românesc au fost conturate şi propuse următoarele etaje de vegetaţie ierboasă:
- Etajul ierbii vântului – corespunzător Etajului foioaselor; - Etajul păiuşului roşu – corespunzător Etajului molidişurilor; - Etajul păiuşului oilor – corespunzător Etajului jnepenişurilor; - Etajul coarnei şi pipiriguţului – corespunzător Etajului alpin. Având ca punct de plecare aceste constatări cu caracter general şi date din Munţii
Cindrelului (ANGHEL şi colab., 1965, 1982; BURCEA, 1965; NIEDERMAIER, 1968; BUZA, 1978 şi alţii) ne-am propus să prezentăm grafic corespondenţa între etajele de vegetaţie lemnoasă şi ierboasă (Fig.7.7).
Fig. 7.7. Etajele de vegetaţie primară şi răspândirea pajiştilor naturale (31,3
mii ha) din Munţii Cindrelului (după ANGHEL, 1982, modificat) Primary vegetation levels and spreading of natural grasslands (31.3 thausand
ha) in Cindrel Mountains by ANGHEL, 1982, modified

309
Rezultatele obţinute sunt convingătoare şi pun în evidenţă extinderea maximă a pajiştilor de 61% între 600-1200 m cu cele mai valoroase agrostete nemorale (etajul ierbii vântului), unde sunt gospodăriile şi odăile de vară, urmată de o secvenţă mai îngustă de 19% pe ecartul 1200-1800 m, cu festucete boreale (etajului păiuşului roşu) şi continuată peste 1800 m de 16% festucete oreale (etajul păiuşului oilor) şi 0,3% caricete alpine, care constituie păşuni de vară pentru ovine, la care se adaugă 3,7% vegetaţie ierboasă de lunci, mlaştini şi stâncării, însumând în total peste 31,3 mii hectare pajişti naturale.
7.4.1.3. Producţia şi calitatea pajiştilor Debutul cercetărilor privind îmbunătăţirea prin fertilizare şi calitatea pajiştilor din
Munţii Cindrelului a fost făcut de către NIEDERMAIER (1954). În paragraful a fost prezentată etajarea vegetaţiei primare şi secundare din aceşti munţi, consideraţi unul din principalele leagăne ale păstoritului românesc. Pe toate aceste etaje de pajişti au fost efectuate experimentări diverse, în special sporirea producţiei prin îngrăşare (fertilizare organică sau chimică) mai ales a celor degradate de Nardus stricta, evaluarea calităţii nutreţurilor de pajişti şi experimentări pe animale, după metodele uzuale. În lucrare s-au folosit numeroase date publicate în literatura de specialitate cu referire strictă la Munţii Cindrelului sau condiţii naturale apropiate cât şi unele rezultate pe pajiştile din Păltiniş sau de la poalele muntelui parţial publicate sau nepublicate (MARUŞCA, 1968-1970; RUSU, 1985-1997; CIORTEA, 2000). Pentru început s-au adus la un numitor comun de exprimare în substanţă uscată (SU) a fitomasei recoltabile a pajiştilor naturale prin utilizarea unor indicatori de transformare şi anume 0,20 pentru masa verde (iarbă) şi 0,84 pentru fân şi otavă (PUIA şi colab., 1976). La fel s-au recalculat compoziţia chimică, coeficienţii de digestibilitate, echivalenţii amidon (EA) şi unităţile nutritive (UN ovăz = EA x 1,66), care s-au raportat şi ele la SU. Evoluţia proceselor climatice, solurile şi gospodărirea terenurilor au influenţat producţia pajiştilor naturale primare (alpine) şi secundare (din zona forestieră) (Tabelul 7.7). Spre deosebire de ansamblul Carpaţilor Meridionali unde în jurul altitudinii de 1200 m se înregistrează un maxim al productivităţii pădurilor şi pajiştilor (MARUŞCA, 1993) în Munţii Cindrelului, intens şi bine gospodăriţi, producţia maximă o întâlnim la poale (600-800 m), asemănător pajiştilor din Alpi, după care scade uniform spre vârf cu 0,15 t/ha SU, pe pajiştile naturale nefertilizate şi cu 0,35 t/ha SU pe cele fertilizate la nivel de 50 N kg/ha şi aceeaşi proporţie PK (P2O5 şi K2O). Reducerea eficienţei fertilizării pe altitudine este evidentă, respectiv cu 1,35 kg SU/kg NPK cu fiecare 100 m în cifre absolute sau 5,5% în cifre relative.

310
Tabelul 7.7 Producţia pajiştilor naturale şi eficienţa fertilizării
cu îngrăşăminte chimice în Munţii Cindrelului (Date prelucrate după NIEDERMAIER: 1958, 1960, 1968; BURCEA: 1965; CARDAŞOL: 1975, 1979; RUSU: 1996; ROTAR: 1997; CIORTEA: 2000)
Production of natural grasslands and fertilisation efficiency with mineral fertilizers in Cindrel Mountains
Altitudine (m)
Producţia de SU (t/ha) Spor SUt/ha
Realizat kg SU
kg NPK
Eficienţă % Neferti-
lizate N50P50K50
kg/ha %
2000-2200 1800-2000
0,9 1,2
1,6 2,3
178 192
0,7 1,1
4,6 7,3
20 31
1600-1800 1400-1600 1200-1400
1,5 1,8 2,1
3,0 3,7 4,4
200 206 210
1,5 1,9 2,3
10,0 12,7 15,4
43 54 66
1000-1200 800-1000 600- 800
2,4 2,7 3,0
5,1 5,8 6,5
213 215 217
2,7 3,1 3,5
18,1 20,8 23,5
77 88
100
GRADI-ENŢI
pt. 100 m
-0,15 t
-0,35 t -0,2 t -1,35
-5,5
Astfel, dacă am considera eficienţa fertilizării pajiştilor de 100% la 600-800 m,
aceasta se reduce de până la 5 ori (20%) la nivelul de 2000-2200 m altitudine. Limita economică a aplicării îngrăşămintelor chimice în Munţii Cindrelului s-ar situa undeva la 1200-1400 m, nivel unde eficienţa îngrăşării scade până la 2/3 (66%) faţă de nivelul de bază (600-800 m).

311
În ceea ce priveşte compoziţia chimică a nutreţurilor de pe pajiştile naturale neîngrăşate se constată o destul de mare creştere pe altitudine (+ 0,35% din SU/100 m) a conţinutului în substanţe organice (SO) şi o scădere în aceeaşi proporţie a cenuşii brute (Cen. B) (Tabelul 7.8).
Tabelul 7.8 Compoziţia chimică medie a nutreţului de pe pajiştile naturale
neîngrăşate din Munţii Cindrelului (Datele recalculate după NIEDERMAIER şi SLUŞANSCHI, 1964)
Mean chemical composition of forage on unfertilised natural grasslands in Cindrel Mountains
% din SU Altitudine (m) SO PB GB SEN Cel B Cen B
1800-2000 95,5 10,4 2,4 52,3 30,4 4,51600-1800 1400-1600 1200-1400
94,8 94,1 93,4
10,711,0 11,3
2,62,8 3,0
51,550,7 49,9
30,0 29,6 21,6
5,25,9 6,6
1000-1200 800-1000 600- 800
92,7 92,0 91,3
11,611,9 12,2
3,23,4 3,6
49,148,3 47,5
28,8 28,4 28,0
7,38,0 8,7
GRADIENŢI pt. 100 m alt.
+0,35 -0,15 -0,10 +0,40 +0,20 -0,35
Dintre SO se remarcă creşterea substanţelor extractive neazotate (SEN) şi a
celulozei brute (Cel. B) direct proporţional cu altitudinea şi o oarecare scădere a conţinutului de proteină brută (PB = N x 6,25) şi grăsime brută (GB) din nutreţul acestor pajişti nefertilizate.Prin îngrăşarea pajiştilor naturale (târlire, gunoire, fertilizare chimică, etc.) conţinuturile de SO şi Cen, B din nutreţ (iarbă, fân, otavă) sunt constante pe ecartul 600- 1400 m alt. (Tabelul 7.9).
Tabelul 7.9. Compoziţia chimică medie a nutreţului de pe pajiştile îngrăşate
(târlite sau gunoite) din Munţii Cindrelului (Datele recalculate după NIEDERMAIER şi SLUŞANSCHI, 1964)
Mean chemical composition of forage on fertilised natural grasslands (paddocked or manured) in Cindrel Mountains
% din SU
Altitudine (m) SO PB GB SEN Cel B Cen B 1200-1400 92,0 17,0 3,9 46,6 24,5 8,01000-1200 800-1000 600- 800
92,0 92,0 92,0
16,516,0 15,5
3,53,1 2,7
46,045,4 44,8
26,0 27,5 29,0
8,08,0 8,0
GRADIENŢI pt. 100 m alt.
0 +0,25 +0,20 +0,30 -0,75 0
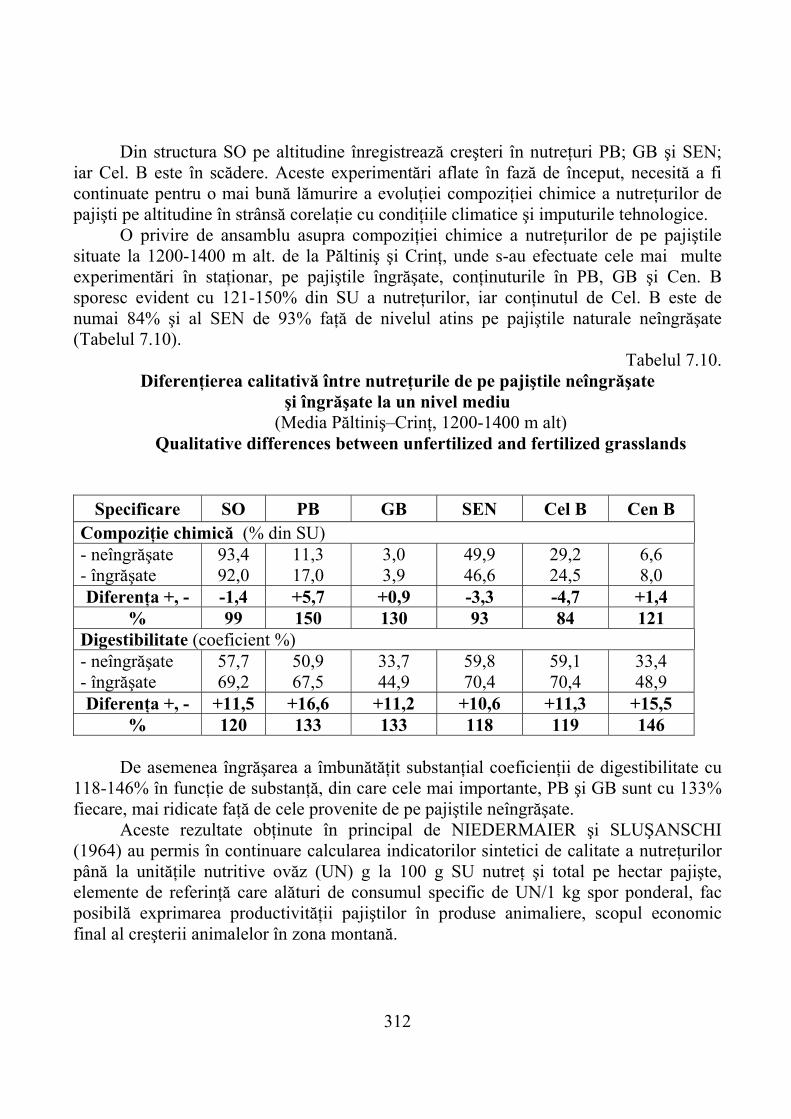
312
Din structura SO pe altitudine înregistrează creşteri în nutreţuri PB; GB şi SEN;
iar Cel. B este în scădere. Aceste experimentări aflate în fază de început, necesită a fi continuate pentru o mai bună lămurire a evoluţiei compoziţiei chimice a nutreţurilor de pajişti pe altitudine în strânsă corelaţie cu condiţiile climatice şi imputurile tehnologice.
O privire de ansamblu asupra compoziţiei chimice a nutreţurilor de pe pajiştile situate la 1200-1400 m alt. de la Păltiniş şi Crinţ, unde s-au efectuate cele mai multe experimentări în staţionar, pe pajiştile îngrăşate, conţinuturile în PB, GB şi Cen. B sporesc evident cu 121-150% din SU a nutreţurilor, iar conţinutul de Cel. B este de numai 84% şi al SEN de 93% faţă de nivelul atins pe pajiştile naturale neîngrăşate (Tabelul 7.10).
Tabelul 7.10. Diferenţierea calitativă între nutreţurile de pe pajiştile neîngrăşate
şi îngrăşate la un nivel mediu (Media Păltiniş–Crinţ, 1200-1400 m alt)
Qualitative differences between unfertilized and fertilized grasslands
Specificare SO PB GB SEN Cel B Cen B Compoziţie chimică (% din SU) - neîngrăşate - îngrăşate
93,4 92,0
11,3 17,0
3,0 3,9
49,9 46,6
29,2 24,5
6,6 8,0
Diferenţa +, - -1,4 +5,7 +0,9 -3,3 -4,7 +1,4 % 99 150 130 93 84 121
Digestibilitate (coeficient %) - neîngrăşate - îngrăşate
57,7 69,2
50,9 67,5
33,7 44,9
59,8 70,4
59,1 70,4
33,4 48,9
Diferenţa +, - +11,5 +16,6 +11,2 +10,6 +11,3 +15,5 % 120 133 133 118 119 146
De asemenea îngrăşarea a îmbunătăţit substanţial coeficienţii de digestibilitate cu
118-146% în funcţie de substanţă, din care cele mai importante, PB şi GB sunt cu 133% fiecare, mai ridicate faţă de cele provenite de pe pajiştile neîngrăşate.
Aceste rezultate obţinute în principal de NIEDERMAIER şi SLUŞANSCHI (1964) au permis în continuare calcularea indicatorilor sintetici de calitate a nutreţurilor până la unităţile nutritive ovăz (UN) g la 100 g SU nutreţ şi total pe hectar pajişte, elemente de referinţă care alături de consumul specific de UN/1 kg spor ponderal, fac posibilă exprimarea productivităţii pajiştilor în produse animaliere, scopul economic final al creşterii animalelor în zona montană.

313
7.4.1.4. Randamentul în spor greutate vie la tineretul taurin şi ovin în perioada de păşunat După cum este bine cunoscut cel mai fidel parametru al productivităţii unei pajişti sunt produsele animaliere (spor greutate, lapte, lână, etc.) realizate în special prin păşunat în zona montană.
Experimentările cu animale pe păşune sunt deosebit de dificile, astfel că avem puţine date în acest sens pe niveluri altudinale. Prin aplicarea unor metode specifice gradienticii montane a fost posibilă conturarea unor consumuri specifice de unităţi nutritive la un kilogram de spor pe altitudine, ceea ce a permis recalcularea producţiei animale pe niveluri altitudinale.
Experimentări mai recente (ROTAR, PRICA, SIMA, 1997) au permis calcularea randamentului în spor ponderal la hectar pentru tineretul ovin la Păltiniş, care este foarte asemănător cu cel stabilit în Munţii Făgăraşului la tineretul taurin şi anume un consum specific calculat de 23 kg SU pentru 1 kg spor ponderal pe ecartul 1200-1400 m altitudine (MARUŞCA, 1997).
Având la dispoziţie date riguroase privind fertilizarea pajiştilor şi stabilirea coeficienţilor de digestibilitate a producţiei (iarbă, fân, otavă) de pe suprafeţele îngrăşate şi neîngrăşate obţinute direct pe batali după metoda in vivo de către NIEDERMAIER şi colaboratorii (1954-1972), urmate de alte experimentări legate de sporirea producţiei efectuate de BURCEA (1965), CARDAŞOL (1979, 1985) şi alţii a fost posibilă elaborarea într-o primă aproximaţie a gradienţilor altitudinali, începând cu cei uzuali de climă şi continuând cu cei de sol, producţia de fitomasă până la produsele animaliere a pajiştilor naturale, care sunt rezultanta interacţiunii tuturor factorilor ecologici şi economici.
La calcularea diferiţilor indicatori s-a căutat a se respecta pe cât posibil datele exacte reieşite din calcul cu o abatere care în majoritate este de 0,1-3%, excepţional 6-7% în plus sau în minus faţă de medie, ajustări care servesc conturării unor gradienţi cu cifre mai rotunde, care nu influenţează pe amsamblu evoluţia rezultatelor şi legităţile unor factori pe niveluri de altitudine.
Pe pajiştile naturale neîngrăşate (nefertilizate) calitatea nutreţurilor respectiv conţinutul în UN pe unitate de produs (SU) este în creştere, ceea ce exprimă şi o calitate a produselor animaliere în special lapte şi carne, insuficient analizate şi apreciate organoleptic, sanogen şi economic în condiţiile ţării noastre până în prezent (Tabelul 7.11.).
Cu toate că se remarcă o creştere generală a calităţii nutreţului pe altitudine, pe ansamblu se înregistrază o scădere a producţiei de UN/ha cu -90 UN/100 m din cauza scăderii pronunţate a producţiei totale de SU/ha pe altitudine. Consumul specific creşte cu 1 UN/1 kg spor ponderal/100 m alt., datorită în principal necesarului suplimentar energetic pentru reglarea temperaturii corpului animalelor într-un climat din ce în ce mai aspru, cu disconfort termic, eolian, pluvial, stres produs de unele animale răpitoare (lupi, urşi, râşi, etc), efort mai mare pentru
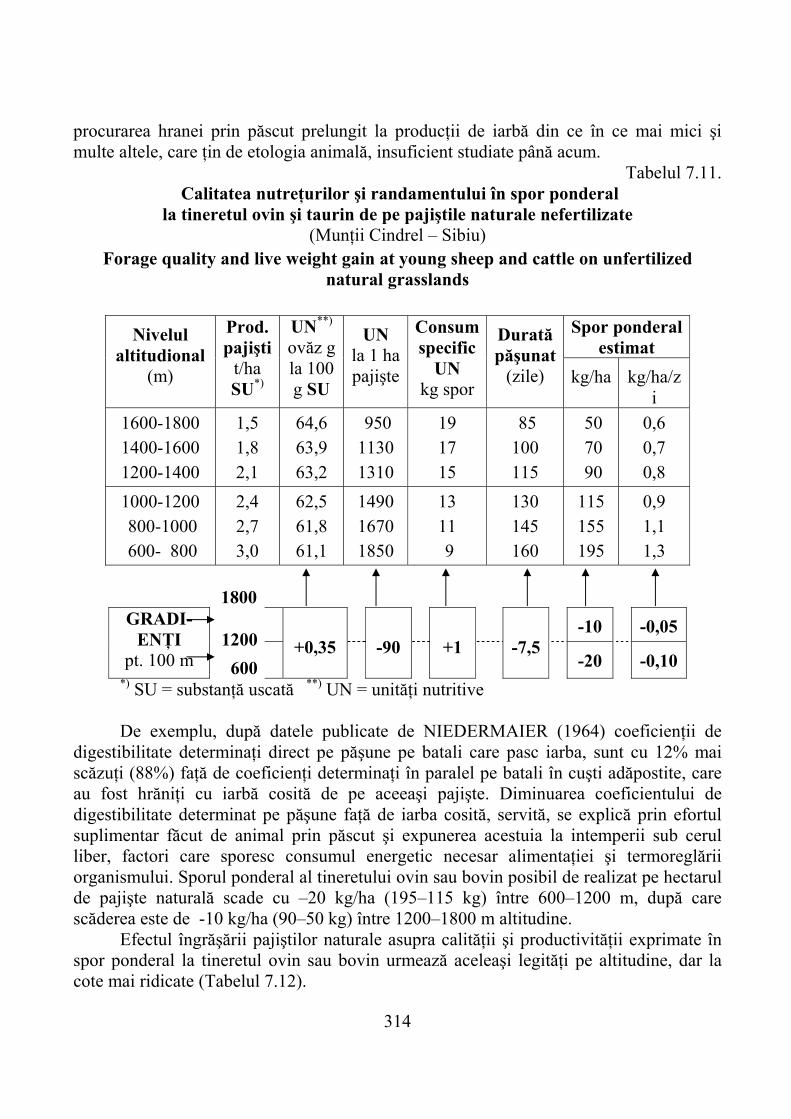
314
procurarea hranei prin păscut prelungit la producţii de iarbă din ce în ce mai mici şi multe altele, care ţin de etologia animală, insuficient studiate până acum.
Tabelul 7.11. Calitatea nutreţurilor şi randamentului în spor ponderal
la tineretul ovin şi taurin de pe pajiştile naturale nefertilizate (Munţii Cindrel – Sibiu)
Forage quality and live weight gain at young sheep and cattle on unfertilized natural grasslands
Nivelul altitudional
(m)
Prod. pajişti
t/ha SU*)
UN**)
ovăz g la 100 g SU
UN la 1 ha pajişte
Consum specific
UN kg spor
Durată păşunat
(zile)
Spor ponderal estimat
kg/ha kg/ha/zi
1600-1800 1400-1600 1200-1400
1,5 1,8 2,1
64,6 63,9 63,2
950 1130 1310
19 17 15
85 100 115
50 70 90
0,6 0,7 0,8
1000-1200 800-1000 600- 800
2,4 2,7 3,0
62,5 61,8 61,1
1490 1670 1850
13 11 9
130 145 160
115 155 195
0,9 1,1 1,3
1800
GRADI-ENŢI
pt. 100 m
1200
600
+0,35
-90
+1
-7,5
-10 -0,05
-20 -0,10*) SU = substanţă uscată **) UN = unităţi nutritive
De exemplu, după datele publicate de NIEDERMAIER (1964) coeficienţii de digestibilitate determinaţi direct pe păşune pe batali care pasc iarba, sunt cu 12% mai scăzuţi (88%) faţă de coeficienţi determinaţi în paralel pe batali în cuşti adăpostite, care au fost hrăniţi cu iarbă cosită de pe aceeaşi pajişte. Diminuarea coeficientului de digestibilitate determinat pe păşune faţă de iarba cosită, servită, se explică prin efortul suplimentar făcut de animal prin păscut şi expunerea acestuia la intemperii sub cerul liber, factori care sporesc consumul energetic necesar alimentaţiei şi termoreglării organismului. Sporul ponderal al tineretului ovin sau bovin posibil de realizat pe hectarul de pajişte naturală scade cu –20 kg/ha (195–115 kg) între 600–1200 m, după care scăderea este de -10 kg/ha (90–50 kg) între 1200–1800 m altitudine. Efectul îngrăşării pajiştilor naturale asupra calităţii şi productivităţii exprimate în spor ponderal la tineretul ovin sau bovin urmează aceleaşi legităţi pe altitudine, dar la cote mai ridicate (Tabelul 7.12).
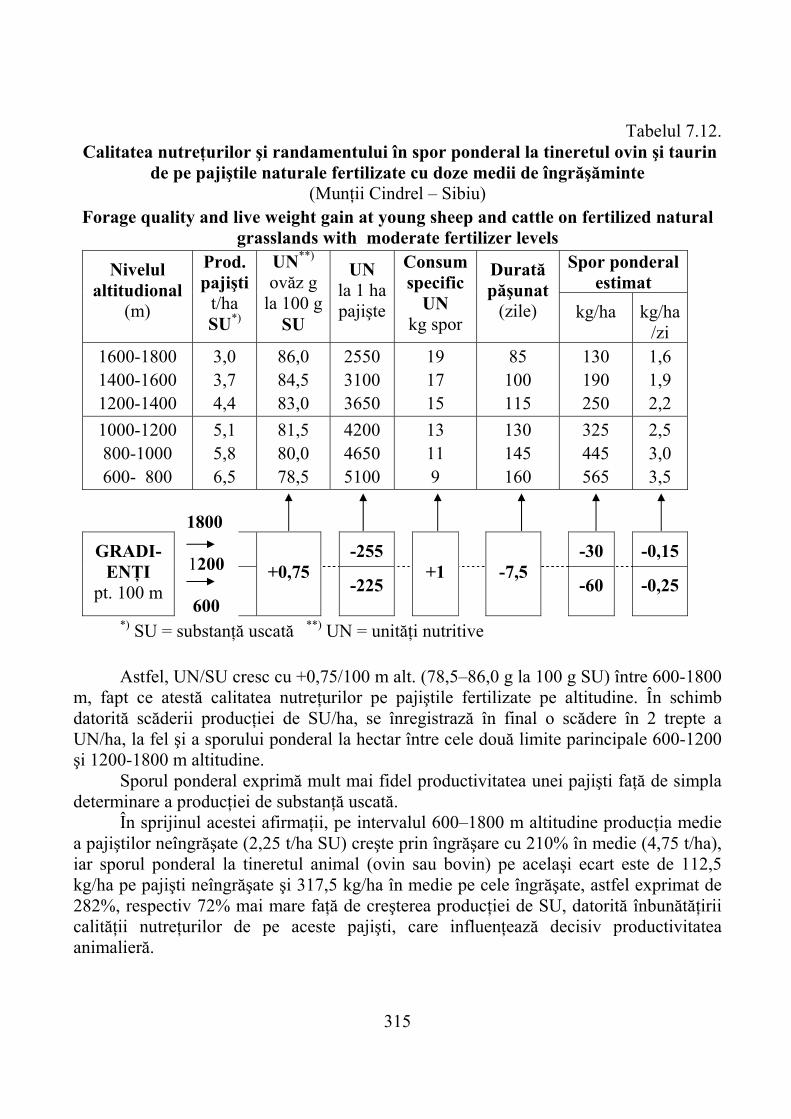
315
Tabelul 7.12. Calitatea nutreţurilor şi randamentului în spor ponderal la tineretul ovin şi taurin
de pe pajiştile naturale fertilizate cu doze medii de îngrăşăminte (Munţii Cindrel – Sibiu)
Forage quality and live weight gain at young sheep and cattle on fertilized natural grasslands with moderate fertilizer levels
Nivelul altitudional
(m)
Prod. pajişti
t/ha SU*)
UN**) ovăz g
la 100 g SU
UN la 1 ha pajişte
Consum specific
UN kg spor
Durată păşunat
(zile)
Spor ponderal estimat
kg/ha kg/ha/zi
1600-1800 1400-1600 1200-1400
3,0 3,7 4,4
86,0 84,5 83,0
2550 3100 3650
19 17 15
85 100 115
130 190 250
1,6 1,9 2,2
1000-1200 800-1000 600- 800
5,1 5,8 6,5
81,5 80,0 78,5
4200 4650 5100
13 11 9
130 145 160
325 445 565
2,5 3,0 3,5
1800
GRADI-ENŢI
pt. 100 m
1200
600
+0,75
-255 +1
-7,5
-30 -0,15
-225 -60 -0,25
*) SU = substanţă uscată **) UN = unităţi nutritive
Astfel, UN/SU cresc cu +0,75/100 m alt. (78,5–86,0 g la 100 g SU) între 600-1800 m, fapt ce atestă calitatea nutreţurilor pe pajiştile fertilizate pe altitudine. În schimb datorită scăderii producţiei de SU/ha, se înregistrază în final o scădere în 2 trepte a UN/ha, la fel şi a sporului ponderal la hectar între cele două limite parincipale 600-1200 şi 1200-1800 m altitudine. Sporul ponderal exprimă mult mai fidel productivitatea unei pajişti faţă de simpla determinare a producţiei de substanţă uscată.
În sprijinul acestei afirmaţii, pe intervalul 600–1800 m altitudine producţia medie a pajiştilor neîngrăşate (2,25 t/ha SU) creşte prin îngrăşare cu 210% în medie (4,75 t/ha), iar sporul ponderal la tineretul animal (ovin sau bovin) pe acelaşi ecart este de 112,5 kg/ha pe pajişti neîngrăşate şi 317,5 kg/ha în medie pe cele îngrăşate, astfel exprimat de 282%, respectiv 72% mai mare faţă de creşterea producţiei de SU, datorită înbunătăţirii calităţii nutreţurilor de pe aceste pajişti, care influenţează decisiv productivitatea animalieră.

316
Cu toate dificultăţile care au apărut în ordonarea şi sintetizarea datelor, în final se poate constata oportunitatea determinării gradienţilor altitudinali, valorificând superior datele mai vechi sau mai noi, în scopul unei analize pe ansamblu şi pe niveluri altitudinale a evoluţiei factorilor ecologici, tehnologici şi economici cu toată complexitatea lor într-un spaţiu montan bine conturat ca Munţii Cindrelului. Această viziune globală de abordare a montanologiei pe lângă caracterul strict descriptiv sau de sinteză, este generatoare de noi direcţii de studiu şi cercetare a unei zone montane de către echipe interdisciplinare în spaţii şi domenii mai puţin abordate. Pentru a ajunge la aceste deziderate, într-o primă fază este necesară valorificarea unui volum imens de date, obţinute cu multă trudă şi dăruire de înaintaşii noştri.
7.4.2. Munţii Făgăraş 7.4.2.1. Evoluţia pe altitudine a vegetaţiei primare în funcţie de climă şi
expoziţia versanţilor Munţii Făgăraşului sunt cei mai masivi şi mai înalţi din tot lanţul carpatic
românesc, având 6 vârfuri ce trec de 2500 m altitudine din care domină Moldoveanu cu 2544 m fiind denumiţi de Em. De MARTONNE, Alpii Transilvaniei. Din punct de vedere geologic sunt formaţi pe sisturi cristaline. Clima este caracteristică zonelor montane cu temperaturi medii anuale de 6-70C la poale până la cca –20C (-2,50C) la vârf şi precipitaţii de 700-850 până la 1300-1400 mm anual în părţile cele mai înalte.
Învelişul de soluri poartă în principal amprenta naturii substratului litologic, care este în general acid, a climatului rece şi umed a vegetaţiei sub care s-a format (MOTCĂ, ROŞU, BELU, 1985).
Vegetaţia primară forestieră este formată aproape în exclusivitate din fag, brad şi molid, care singure sau în amestec alcătuiesc etaje de vegatţie ce se desfăşoară în altitudine, începând cu 600-700 m până la 1650 m pe versantul nordic şi până la 1750 m pe versantul sudic, după care urmează celelalte etaje, subalpin şi aplin (Tabelul 7.13).
Tabelul 7.13. Delimitarea etajelor de vegetaţie primară în Munţii Făgăraşului
(După MOTCĂ, ROŞU, BELU, 1985 Delimitation of primary vegetation levels in Fagaras Mountains
Etajul de vegetaţie Versantul nordic m alt. Versantul sudic m alt.
Limita inferioară
Limita superioară
Limita inferioară
Limita superioară
Subetajul făgetelor Etajul molidişurilor Etajul jnepenişului (subalpin) Etajul alpin
600 - 7001200-12501600-1650
2200
1200-12501600-1650
22002544
650- 900 1350-1450 1700-1750
2300
1350-14501700-1750
23002544

317
Acest model de reprezentare sub formă de tabel este cu mult mai explicită decât prezentarea simplă a acestor date în cuprinsul unui text. Prin reprezentare grafică aceleaşi limite altitudinale ale vegetaţiei completate cu date climatice (ŢÂŞTEA, NEACŞA, 1974) câştigă şi mai mult în valoare prin posibilitatea stabilirii unor corelaţii între etajele de vegetaţie şi climă (Fig. 7.8.)
Fig. 7.8. Condiţii climatice şi etaje de vegetaţie primară din Munţii Făgăraş
Climate and primary vegetation levels in Fagaras Mountains
De asemenea s-a stabilit gradientul hidric pentru Munţii Făgăraşului care este de +30 mm/100 m, iar gradientul termic este cel cunoscut de –0,50C/100 m altitudine. Toate aceste date se folosesc în continuare la caracterizarea generală a condiţiilor naturale, a formaţiilor de pajişti şi productivităţii animalelor.

318
7.4.2.2.Îmbunătăţirea prin fertilizare cu îngrăşăminte chimice În urma unor experimentări riguroase în staţionare, pe durate diferite, dar de
minim 3 ani, a fost posibilă conturarea evoluţiei pe altitudine a producţiei şi calităţii pajiştilor fertilizate cu diferite doze de îngrăşăminte chimice (PUŞCARU-SOROCEANU EVDOCHIA şi colab.: 1963, 1965, 1969; PUŞCARU şi colab.: 1977; MOTCĂ: 1972; BELU: 1982) (Tabelul 7.14).
Tabelul 7.14. Producţia de substanţă uscată (SU) şi de proteină digestibilă (PD)
la principalele tipuri de pajişti din Munţii Făgăraşului (după MOTCĂ, ROŞU, BELU: 1985)
Dry matter yield (DM) and digestibile protein ( DP) for main types grasslands in Fagaras Mountains
Tipul de pajişte
Locul determinării
(muntele, altitudinea)
Nefertilizat
100 N*) 50 P2O5
50 K2O
300 N*) 100 P2O5
100 K2O SUq /ha
PDkg/ha
SUq /ha
PDkg/ha
SUq /ha
PDkg/ha
Agrostis tenuis
Recea 650 m Valea Cumpăna 800 m Mormântul 1100 m Florea 1350 m
36,9 21,6 26,0 38,1
-100180370
82,0 50,0 61,1 75,3
- 840 910 950
90,9 77,6 83,7 94,5
-176017001020
Festuca rubra ssp. commutata
Marginea 1500 m 19,4 170 67,5 800 87,1 930
Nardus stricta Marginea 1600 m Maliţa 1600 m Maliţa 1700 m Comarnic 1800 m Maliţa 1820 m Maliţa 1900 m Maliţa 2000 m Podeanu 2000 m Maliţa 2050 m
19,6 27,8 24,5 13,8 27,2 12,2 8,6 14,5 4,3
90--
130---
110-
53,6 47,2 35,8 49,5 39,0 31,8 19,9 22,8 18,4
420 - -
950 - - -
220 -
88,1 56,7 43,0 64,6 41,9 39,4 30,3 25,4 25,1
930--
1430---
400-
Festuca ovina ssp. sudetica
Podeanu 2150 m 11,8 60 30,5 220 27,7 250
Agrostis rupestris
Podeanu 2200 m 6,5 40 11,5 150 18,5 340
Juncus trifidus Lespezi 2300 m 6,6 50 - - - -Carex curvula Lespezi 2350 m 9,7 80 12,0 130 7,1 80
*)La Recea: 70 P2O5, 80 K2O o dată la 2 ani; La Maliţa: fără K2O

319
Din acest volum impresionant de date privind producţia de SU şi PB al diferitelor tipuri de pajişti situate pe un ecart de 1700 m (650-2350 m alt.) se conturează concluzia diminuării pe altitudine a acestor doi importanţi parametri pratotehnici, chiar şi la doze mari de îngrăşăminte chimice.
Această evoluţie este completată şi întărită după stabilirea unor gradienţi ai producţiei pajiştilor la cele 3 niveluri principale de fertilizare chimică (Fig.7.9.)
Fig. 7.9. Evoluţia pe altitudine a producţiei pajiştilor permanente, fertilizate cu diferite doze de îngrăşăminte chimice în Munţii Făgăraşului Evolution of permanent grasland production fertilized with different mineral levels
in Fagaras Mountains
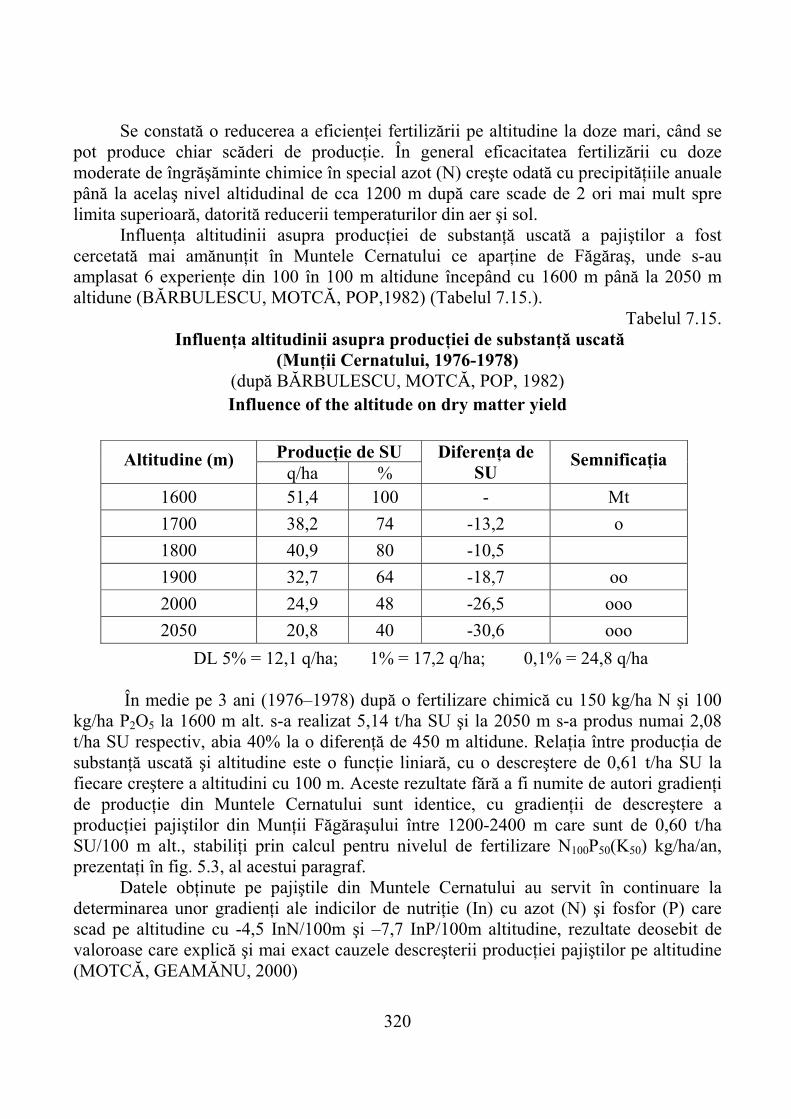
320
Se constată o reducerea a eficienţei fertilizării pe altitudine la doze mari, când se pot produce chiar scăderi de producţie. În general eficacitatea fertilizării cu doze moderate de îngrăşăminte chimice în special azot (N) creşte odată cu precipităţiile anuale până la acelaş nivel altidudinal de cca 1200 m după care scade de 2 ori mai mult spre limita superioară, datorită reducerii temperaturilor din aer şi sol.
Influenţa altitudinii asupra producţiei de substanţă uscată a pajiştilor a fost cercetată mai amănunţit în Muntele Cernatului ce aparţine de Făgăraş, unde s-au amplasat 6 experienţe din 100 în 100 m altidune începând cu 1600 m până la 2050 m altidune (BĂRBULESCU, MOTCĂ, POP,1982) (Tabelul 7.15.).
Tabelul 7.15. Influenţa altitudinii asupra producţiei de substanţă uscată
(Munţii Cernatului, 1976-1978) (după BĂRBULESCU, MOTCĂ, POP, 1982) Influence of the altitude on dry matter yield
Altitudine (m) Producţie de SU Diferenţa de SU
Semnificaţia q/ha %
1600 51,4 100 - Mt
1700 38,2 74 -13,2 o
1800 40,9 80 -10,5
1900 32,7 64 -18,7 oo
2000 24,9 48 -26,5 ooo
2050 20,8 40 -30,6 ooo
DL 5% = 12,1 q/ha; 1% = 17,2 q/ha; 0,1% = 24,8 q/ha
În medie pe 3 ani (1976–1978) după o fertilizare chimică cu 150 kg/ha N şi 100 kg/ha P2O5 la 1600 m alt. s-a realizat 5,14 t/ha SU şi la 2050 m s-a produs numai 2,08 t/ha SU respectiv, abia 40% la o diferenţă de 450 m altidune. Relaţia între producţia de substanţă uscată şi altitudine este o funcţie liniară, cu o descreştere de 0,61 t/ha SU la fiecare creştere a altitudini cu 100 m. Aceste rezultate fără a fi numite de autori gradienţi de producţie din Muntele Cernatului sunt identice, cu gradienţii de descreştere a producţiei pajiştilor din Munţii Făgăraşului între 1200-2400 m care sunt de 0,60 t/ha SU/100 m alt., stabiliţi prin calcul pentru nivelul de fertilizare N100P50(K50) kg/ha/an, prezentaţi în fig. 5.3, al acestui paragraf.
Datele obţinute pe pajiştile din Muntele Cernatului au servit în continuare la determinarea unor gradienţi ale indicilor de nutriţie (In) cu azot (N) şi fosfor (P) care scad pe altitudine cu -4,5 InN/100m şi –7,7 InP/100m altitudine, rezultate deosebit de valoroase care explică şi mai exact cauzele descreşterii producţiei pajiştilor pe altitudine (MOTCĂ, GEAMĂNU, 2000)
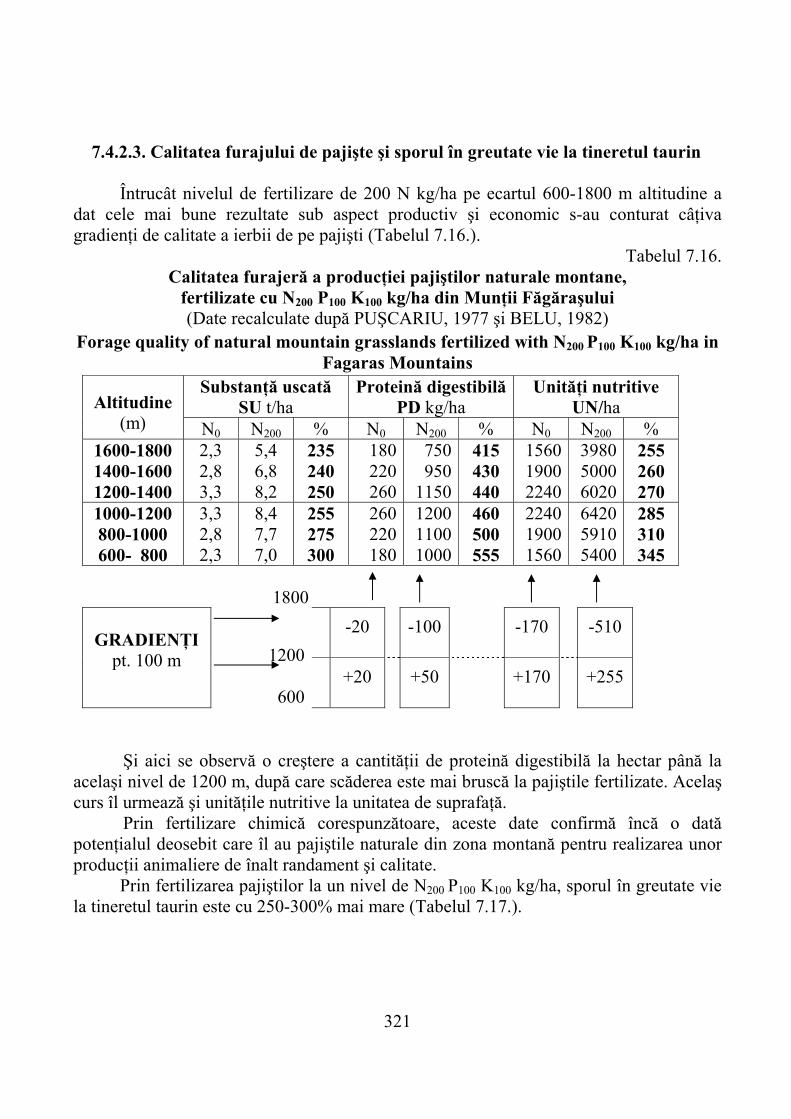
321
7.4.2.3. Calitatea furajului de pajişte şi sporul în greutate vie la tineretul taurin
Întrucât nivelul de fertilizare de 200 N kg/ha pe ecartul 600-1800 m altitudine a
dat cele mai bune rezultate sub aspect productiv şi economic s-au conturat câţiva gradienţi de calitate a ierbii de pe pajişti (Tabelul 7.16.).
Tabelul 7.16. Calitatea furajeră a producţiei pajiştilor naturale montane,
fertilizate cu N200 P100 K100 kg/ha din Munţii Făgăraşului (Date recalculate după PUŞCARIU, 1977 şi BELU, 1982)
Forage quality of natural mountain grasslands fertilized with N200 P100 K100 kg/ha in Fagaras Mountains
Altitudine (m)
Substanţă uscată SU t/ha
Proteină digestibilăPD kg/ha
Unităţi nutritive UN/ha
N0 N200 % N0 N200 % N0 N200 %1600-1800 1400-1600 1200-1400
2,3 2,8 3,3
5,4 6,8 8,2
235 240 250
180220260
750950
1150
415 430 440
1560 1900 2240
3980 5000 6020
255 260 270
1000-1200 800-1000 600- 800
3,3 2,8 2,3
8,4 7,7 7,0
255 275 300
260220180
120011001000
460 500 555
2240 1900 1560
6420 5910 5400
285 310 345
1800
GRADIENŢI
pt. 100 m
1200
600
-20
-100 -170 -510
+20
+50 +170 +255
Şi aici se observă o creştere a cantităţii de proteină digestibilă la hectar până la acelaşi nivel de 1200 m, după care scăderea este mai bruscă la pajiştile fertilizate. Acelaş curs îl urmează şi unităţile nutritive la unitatea de suprafaţă. Prin fertilizare chimică corespunzătoare, aceste date confirmă încă o dată potenţialul deosebit care îl au pajiştile naturale din zona montană pentru realizarea unor producţii animaliere de înalt randament şi calitate.
Prin fertilizarea pajiştilor la un nivel de N200 P100 K100 kg/ha, sporul în greutate vie la tineretul taurin este cu 250-300% mai mare (Tabelul 7.17.).
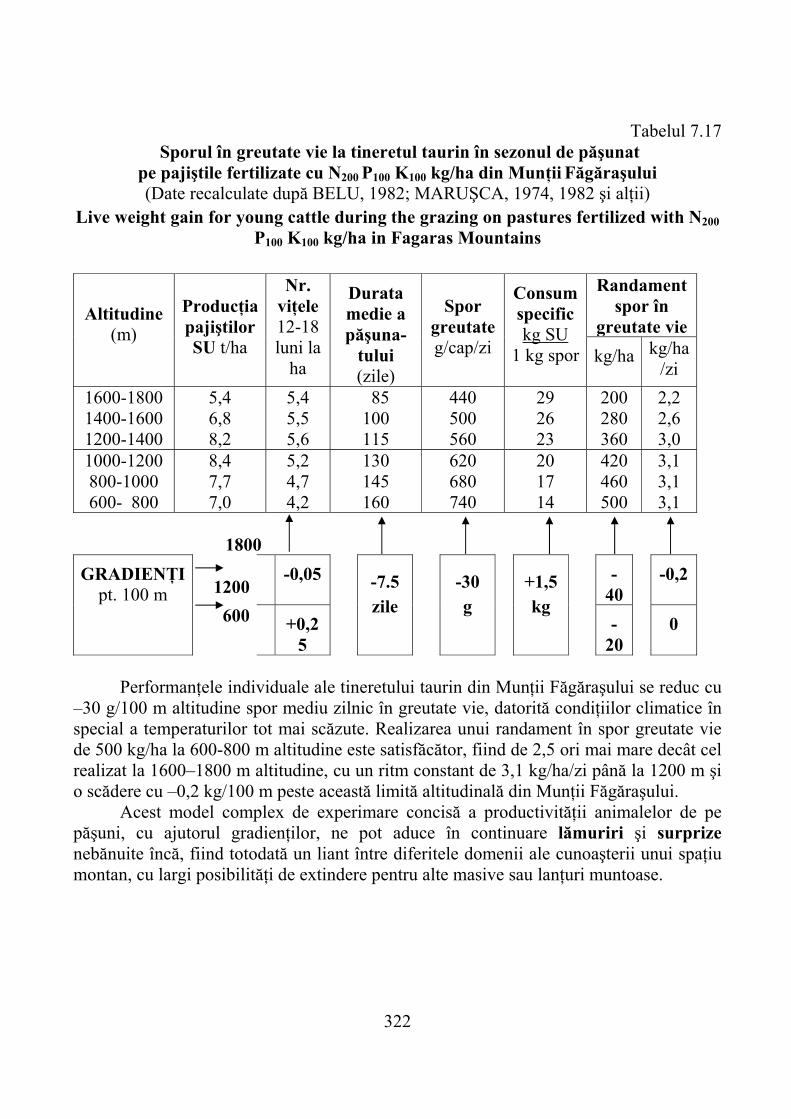
322
Tabelul 7.17 Sporul în greutate vie la tineretul taurin în sezonul de păşunat
pe pajiştile fertilizate cu N200 P100 K100 kg/ha din Munţii Făgăraşului (Date recalculate după BELU, 1982; MARUŞCA, 1974, 1982 şi alţii)
Live weight gain for young cattle during the grazing on pastures fertilized with N200
P100 K100 kg/ha in Fagaras Mountains
Altitudine (m)
Producţia pajiştilor SU t/ha
Nr. viţele 12-18 luni la
ha
Durata medie a păşuna-
tului (zile)
Spor
greutate g/cap/zi
Consum specific kg SU
1 kg spor
Randament spor în
greutate vie
kg/ha kg/ha/zi
1600-1800 1400-1600 1200-1400
5,4 6,8 8,2
5,4 5,5 5,6
85 100 115
440 500 560
29 26 23
200 280 360
2,2 2,6 3,0
1000-1200 800-1000 600- 800
8,4 7,7 7,0
5,2 4,7 4,2
130 145 160
620 680 740
20 17 14
420 460 500
3,1 3,1 3,1
1800
GRADIENŢI pt. 100 m
1200
600
-0,05 -7.5 zile
-30 g
+1,5 kg
-40
-0,2
+0,25
-20
0
Performanţele individuale ale tineretului taurin din Munţii Făgăraşului se reduc cu
–30 g/100 m altitudine spor mediu zilnic în greutate vie, datorită condiţiilor climatice în special a temperaturilor tot mai scăzute. Realizarea unui randament în spor greutate vie de 500 kg/ha la 600-800 m altitudine este satisfăcător, fiind de 2,5 ori mai mare decât cel realizat la 1600–1800 m altitudine, cu un ritm constant de 3,1 kg/ha/zi până la 1200 m şi o scădere cu –0,2 kg/100 m peste această limită altitudinală din Munţii Făgăraşului.
Acest model complex de experimare concisă a productivităţii animalelor de pe păşuni, cu ajutorul gradienţilor, ne pot aduce în continuare lămuriri şi surprize nebănuite încă, fiind totodată un liant între diferitele domenii ale cunoaşterii unui spaţiu montan, cu largi posibilităţi de extindere pentru alte masive sau lanţuri muntoase.

323
7.4.3. Munţii Bucegi 7.4.3.1. Evoluţia pe altitudine a taliei unor specii de graminee perene din
etajele subalpin - alpin
Condiţiile climatice şi de vegetaţie pentru creşterea şi dezvoltarea plantelor sunt progresiv mai nefavorabile pe altitudine, fapt ce se reflectă în schimbările ce au loc în structura învelişului vegetal şi talia speciilor componente (MARUŞCA, SPIRESCU, 1994). Pentru a contura mai exact amploarea acestor fenomene au fost efectuate mai multe observaţii şi măsurători biometrice la plantele mature ce aparţin la 7 specii de graminee perene mai răspândite în golul montan între 1400-2400 m altitudine (Fig. 7.10).
Fig. 7.10. Gradienţii creşterii în înălţime (G.h.) a unor graminee de pajişti din
„golul montan” românesc Gradients of height increasing of some grasses in roumanian „mountain hole”
În condiţiile biotermice prezentate în capitole anterioare, gradienţii creşterii în înălţime a gramineelor sunt în scădere cu 1,5 cm/100 m alt. (20-8 cm) la Agrostis rupestris între 1400-2400 m şi până la 5 cm/100 m (45-25 cm) pentru Deschampsia flexuosa pe ecartul 1400-2000 m altitudine. Restul speciilor luate în studiu descresc cu 2-3 cm/100 m până la nivelul altitudinal unde vieţuiesc în covorul ierbos.

324
Prin descreşterea progresivă a taliei gramineelor perene peste 1400 m altitudine se reduce evident şi producţia pajiştilor având în vedere că specile din această familie (Graminee) domină în covorul ierbos. Ca urmare a înăspririi condiţiilor de vegetaţie pe altitudine pe lângă micşorarea taliei gramineelor, se diminuiază şi proporţia lăstarilor generativi până la un prag de dispariţie periodică sau totală a lor, lăsând locul altor ecotipuri sau specii cu înmulţire preponderent vegetativă în etajul alpin superior.
7.4.3.2. Evoluţia biodiversităţii nardetelor pe altitudine După o analiză mai atentă a compoziţiei floristice a releveurilor pe nivele
altitudinale s-a stabilit o reducere a numărului cu 6 specii pentru fiecare 100 m altitudine (Fig. 7.11.).
Această tendinţă binecunoscută de reducere a biodiversităţii pe altitudine în zona montană este în strânsă corelaţie cu diminuarea treptată a condiţiilor climatice de vieţuire a plantelor şi animalelor.
Fig. 7.11. Reducerea pe altitudine a biodiversităţii nardetelor
din Carpaţii Meridionali Declin of biodiversity nardion on altitude in Meridional Carpathians
7.4.3.3. Producţia de lapte de vacă în perioada de păşunat
Cu toate dificultăţile mari care le presupune experimentările pe vaci de lapte în scopul determinării productivităţii pajiştilor, la Institutul de Cercetare şi Producţie pentru Cultura Pajiştilor, după 1995 au fost începute astfel de cercetări la Măgurele (600 m alt.)

325
şi Blana (1800 m alt.), câmpuri reprezentative pentru seria litologică bazică Bucegi – Piatra Craiului. Pentru exemplificare se prezintă rezultatele obţinute la Baza de cercetări pentru Pajişti Montane de la Blana-Bucegi unde în medie pe 3 ani pe nardetele îmbunătăţite s-a depăşit 5000 l/ha lapte de vacă de la rasa Schwitz (Brună de Maramureş) adaptată mai bine condiţiilor climatice montane (Tabelul 7.18). După lucrările de îmbunătăţire a nardetelor s-au ameliorat substanţial caracteristicile agrochimice ale solului, compoziţia floristică, producţia de substanţă uscată şi proteină, care au permis întreţinerea a peste 5 vaci/ha o perioadă medie de 83 zile de păşunat (MARUŞCA, POP, NEAGU, MOCANU, BLAJ, OPREA, 2000). Calitatea furajeră a ierbii a redus consumul de substanţă uscată, proteină şi durata de păscut pentru realizarea unui litru de lapte la nivelul de 64-86% din consumul şi timpul necesar pentru obţinerea aceluiaşi produs pe păşunea neîmbunătăţită. Numai prin caracterizarea complexă a factorilor de sol-plantă-animal produs şi comportament animalier într-un concept unitar se pot contura agroecosistemele de pajişti sau pratoecosistemele.
Pe baza acestor date proprii realizate la institutul de profil s-au stabilit gradienţii producţiei de lapte pe altitudine (Tabelul 7.19).
Rezultatele obţinute pe pajişti fertilizate şi raţional folosite cu vaci de lapte pot fi destul de mari şi mai ales profitabile pentru crescătorii de animale din zona montană. Şi în acest caz producţia individuală de lapte scade cu 0,4 l/100 m altitudine, datorită în primul rând a condiţiilor climatice, iar consumul specific pentru realizarea unui litru de lapte creşte cu 0,05 kg SU/l pentru fiecare 100 m altitudine, astfel de la cca. 10000 l/ha lapte, care se produce la 600-800 m, se ajunge la numai 5000 l/ha la 1600-1800 m altitudine, bine înţeles într-o perioadă mai scăzută de timp de numai 85 zile faţă de 160 zile de păşunat.
Evident pentru susţinerea producţiei de lapte de vacă în zona montană sunt necesare acordarea unor preţuri diferenţiate pe altitudine, aşa cum se practică în economia agricolă a unor ţări dezvoltate din jurul Alpilor.
Gradienţii producţiilor animaliere se pot utiliza la fundamentarea ştiinţifică a noilor preţuri pentru producţia de carne, lapte şi alte produse din zona montană, care pot avea un rol decisiv pentru stabilizarea populaţiei în acest spaţiu defavorizat pentru agricultură şi în final, cu efecte benefice pe ansamblul economiei.

326
Tabelul 7.18. Date generale privind solul, producţia de substanţă uscată, lapte şi comportamentul
vacilor pe păşunile subalpine de Nardus stricta din Platoul Bucegilor (1996-1998) General data regarding to the soil, dry matter yield, milk and cows behaviour on
subalpine pastures of Nardus stricta in Bucegi Plateau Muntele Blana, 1800 m altitudine
Specificare UM Păşune Diferenţă +, -
% Neameliorată Îmbunătăţită*
Caracteristicile agrochimice ale solui (0-15 cm adc.) - pH în H2O - grad saturaţie baze (V) - conţinut humus - conţinut P-AL - conţinut K-AL - conţinut Al3+
Ind.% %
ppm ppm me
4,824,2 16,1 19,4 96,0 4,1
5,251,2 15,5 52,0
108,0 1,3
+0,4 +27,0 -0,6
+32,6 +12,0 -2,8
108212 96
268 113 32
Compoziţia floristică specii cu valoare redusă** specii cu valoare bună***
%%
8515
1585
-70 +70
**
Valoarea pastorală 10/15 50/65 +45 450 Producţii (83 zile păşunat) - substanţă uscată - proteină brută - încărcare păşune - lapte vacă (3,5%G) - lapte vacă (3,5%G)
t/hakg/ha
vaci/hal/cap l/ha
2,48468 1,48 10,8 1324
6,001519 5,12 11,8 5012
+3,52 +1051 +3,64 +1,0 3688
242325 346 109 379
Comportament animal (24 ore) - păscut - rumegat - odihnă din care: somn
h minh min h min (h min)
10,006,08 7,52
(0,42)
8,306,56 8,34
(0,55)
-1,30 +0,48 +0,42
(+0,13)
85113 109 130
Necesar pentru un litru lapte - substanţă uscată (SU) - proteină brută furaj (PB) - durată păscut
kgkg
min
1,870,35 56
1,200,30 43
-0,67 -0,05 -13
6486 77
* Păşunea neameliorată având în compoziţie peste 70% participare Nardus stricta a fost îmbunătăţită prin fertilizare cu N150 P50 K50 kg/ha 3 ani consecutiv, toate cele 3 variante, pe fondul amendat cu CaO la 2/3 Ah (2 variante) şi neamendat (1 variantă) cât şi înfiinţarea unei pajişti semănate pe fond amendat (1 variantă); ** Nardus stricta, Festuca nigrescens, Festuca ovina, Phleum alpinum, Deschampsia flexuosa, Potentilla aurea, Viola declinata, Campanula napuligera, etc.; *** Semănate: Phleum pratense, Festuca pratensis, Lolium perenne, Poa pratensis; Spontane: Festuca rubra, Agrostis capillaris, A. rupestris, Poa media, Trifolium repens, Ligusticum mutellina, etc.
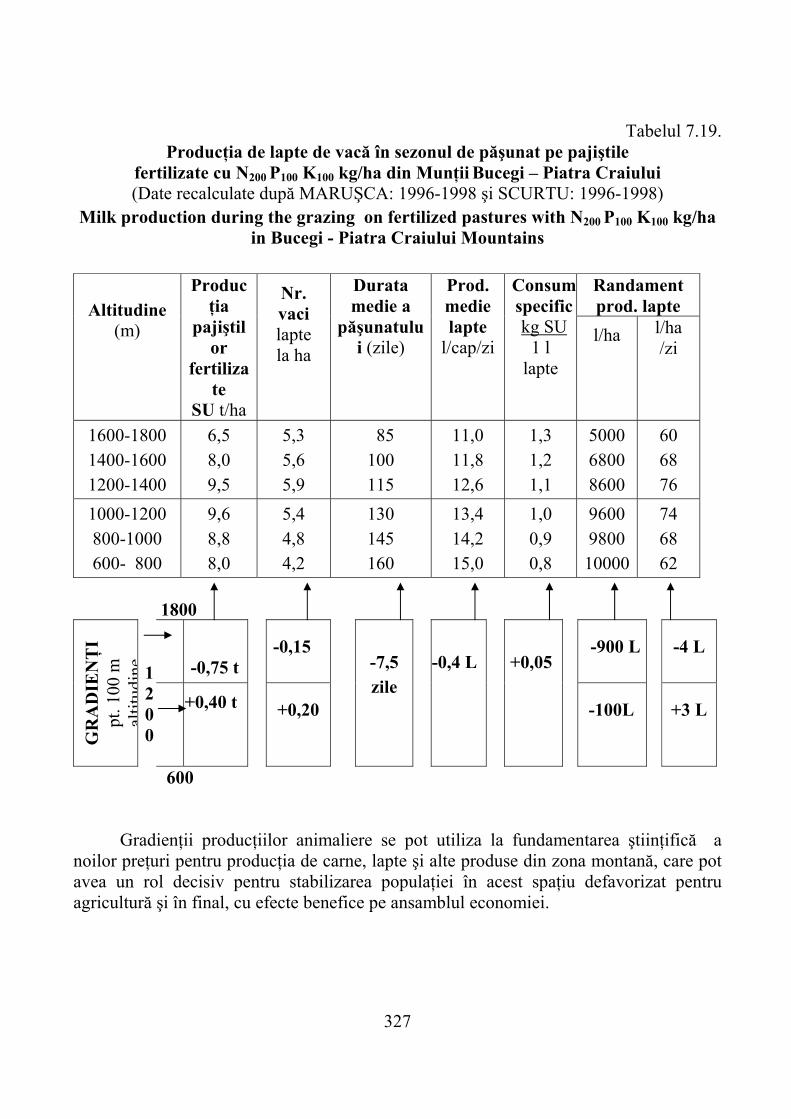
327
Tabelul 7.19. Producţia de lapte de vacă în sezonul de păşunat pe pajiştile
fertilizate cu N200 P100 K100 kg/ha din Munţii Bucegi – Piatra Craiului (Date recalculate după MARUŞCA: 1996-1998 şi SCURTU: 1996-1998)
Milk production during the grazing on fertilized pastures with N200 P100 K100 kg/ha in Bucegi - Piatra Craiului Mountains
Altitudine (m)
Producţia
pajiştilor
fertilizate
SU t/ha
Nr. vaci lapte la ha
Durata medie a
păşunatului (zile)
Prod. medie lapte
l/cap/zi
Consum specific kg SU
1 l lapte
Randament prod. lapte
l/ha l/ha /zi
1600-1800 1400-1600 1200-1400
6,5 8,0 9,5
5,3 5,6 5,9
85 100 115
11,0 11,8 12,6
1,3 1,2 1,1
5000 6800 8600
60 68 76
1000-1200 800-1000 600- 800
9,6 8,8 8,0
5,4 4,8 4,2
130 145 160
13,4 14,2 15,0
1,0 0,9 0,8
9600 9800 10000
74 68 62
1800
GR
AD
IENŢ
I pt
. 100
m
alti
tudi
ne
1200
-0,75 t
-0,15
-7,5 zile
-0,4 L +0,05 -900 L -4 L
+0,40 t +0,20
-100L +3 L
600 Gradienţii producţiilor animaliere se pot utiliza la fundamentarea ştiinţifică a noilor preţuri pentru producţia de carne, lapte şi alte produse din zona montană, care pot avea un rol decisiv pentru stabilizarea populaţiei în acest spaţiu defavorizat pentru agricultură şi în final, cu efecte benefice pe ansamblul economiei.

328
7.5. PROGNOZA PRODUCTIVITĂŢII PAJIŞTILOR MONTANE ÎN CONTEXTUL ÎNCĂLZIRII GLOBALE A CLIMEI
7.5.1. Evoluţia climatică şi consecinţele ei Încălzirea globală a climei va redistribui pe niveluri altitudinale parametrii de
temperatură şi umiditate împreună cu învelişul vegetal şi productivitatea lui. Cu ajutorul gradienţilor altitudinali se pot stabili în timp evoluţia acestor parametrii climatici şi de vegetaţie care vor influenţa la rândul lor performanţele realizate din creşterea animalelor şi practicarea agroturismului. În curând populaţia va fi nevoită să recucerească zonele montane abandonate după ce va fi obligată să părăsească zonele costiere inundate de nivelul tot mai crescut al oceanului planetar şi extinderea accentuată a suprafeţelor aride improprii pentru practicarea agriculturii.
Zona montană mai înaltă poate deveni astfel un spaţiu de refugiu, supravieţuire şi petrecerea mai plăcută a timpului liber în actualul context climatic nefavorabil pentru zonele mai joase. Pentru aceasta sunt necesare luarea din timp a unor măsuri specifice de creştere a productivităţii pajiştilor, drumuri de acces şi construcţii, plantaţii forestiere, acumulări de apă, etc., care să satisfacă nevoile noilor veniţi în spaţiul montan.
Conform ultimelor previziuni ale evoluţiei climei pe glob, fenomenul de încălzire generală datorită activităţilor umane (defrişare, industrializare, transporturi, etc.) cu sporirea emisiilor de dioxid de carbon, topirea calotelor glaciare şi a gheţarilor montani, creşterea nivelului oceanelor, inundaţii, aridizare şi deşertificare, intensificarea proceselor orajoase (uragane, taifunuri, tornade, cicloane, etc.) vor avea un impact major asupra întregii umanităţi cu consecinţe imprevizibile dintre cele mai negative (AL GORE, 2007).
Nu întâmplător de Ziua Internaţională a Muntelui din anul 2007, s-a ales ca obiectiv tematic schimbările climatice globale pentru care s-au făcut câteva previziuni cu exemplificări din Carpaţii României.
Din datele OMM (Organizaţia Meteorologică Mondială) cu sediul la Geneva, temperatura medie a globului a crescut în perioada 1901 – 2000 cu 0,6 0C ceea ce este extrem de mult. Pentru România, conform INMH – Bucureşti, această creştere este de 0,3 0C, mai mare în regiunile de sud şi est (0,8 0C) şi mai mică în regiunile intracarpatice (0,10C). Încălzirea climei este mai pronunţată după anii 1961 şi cu deosebire după anul 2000 (2003, 2005) când frecvenţa zilelor tropicale (maxima zilnică > 30 0C) a crescut îngrijorător de mult şi zilele de iarnă (maxima zilnică < 0 0C ) a scăzut substanţial. Drept urmare mai multe zone din ţara noastră prezintă un risc ridicat de secetă şi deşertificare în special cele unde temperatura medie anuală este mai mare de, 10 0 C; suma precipitaţiilor atmosferice anuale este sub 350 – 550 mm; precipitaţii aprilie – octombrie sunt sub200 – 350 mm iar rezerva apă din sol 0 – 100 cm la 31 martie este mai mică de 950 –1500 mc /ha
Conform Convenţiei Naţiunilor Unite pentru Combaterea Deşertificării (UNCDD) indicele de ariditate (cantitatea anuală de precipitaţii / evapotranspiraţia potenţială –
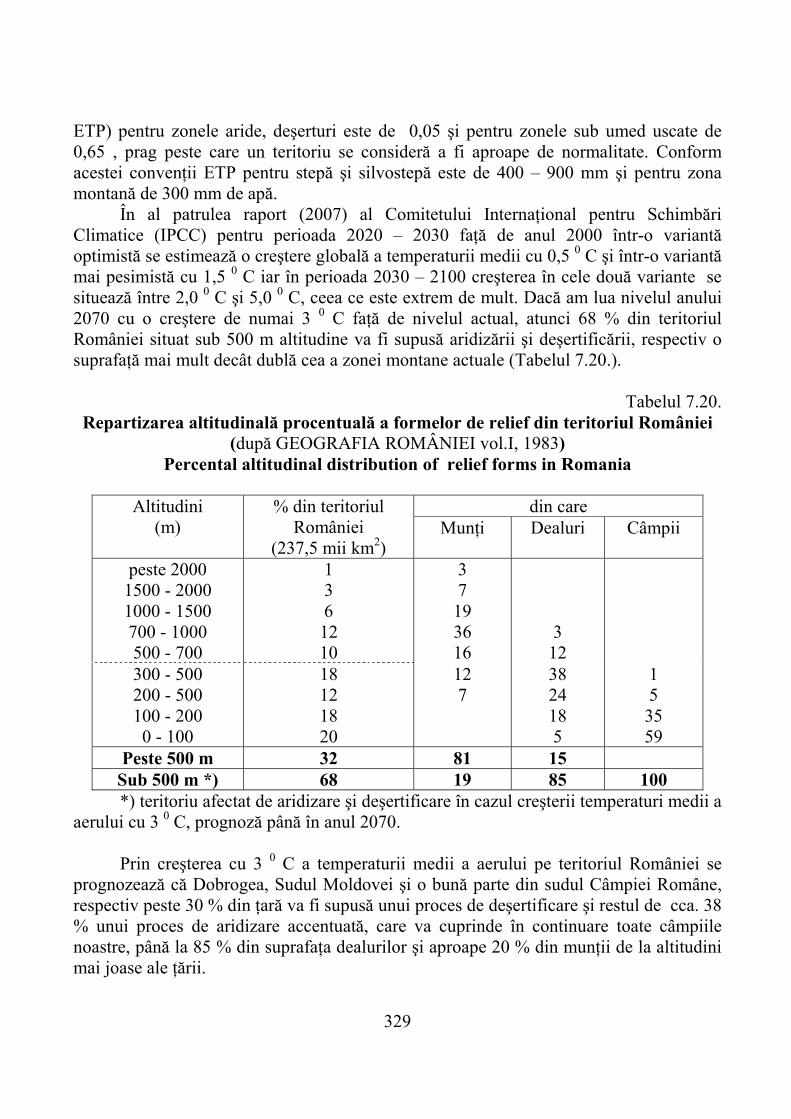
329
ETP) pentru zonele aride, deşerturi este de 0,05 şi pentru zonele sub umed uscate de 0,65 , prag peste care un teritoriu se consideră a fi aproape de normalitate. Conform acestei convenţii ETP pentru stepă şi silvostepă este de 400 – 900 mm şi pentru zona montană de 300 mm de apă.
În al patrulea raport (2007) al Comitetului Internaţional pentru Schimbări Climatice (IPCC) pentru perioada 2020 – 2030 faţă de anul 2000 într-o variantă optimistă se estimează o creştere globală a temperaturii medii cu 0,5 0 C şi într-o variantă mai pesimistă cu 1,5 0 C iar în perioada 2030 – 2100 creşterea în cele două variante se situează între 2,0 0 C şi 5,0 0 C, ceea ce este extrem de mult. Dacă am lua nivelul anului 2070 cu o creştere de numai 3 0 C faţă de nivelul actual, atunci 68 % din teritoriul României situat sub 500 m altitudine va fi supusă aridizării şi deşertificării, respectiv o suprafaţă mai mult decât dublă cea a zonei montane actuale (Tabelul 7.20.).
Tabelul 7.20. Repartizarea altitudinală procentuală a formelor de relief din teritoriul României
(după GEOGRAFIA ROMÂNIEI vol.I, 1983) Percental altitudinal distribution of relief forms in Romania
Altitudini
(m) % din teritoriul
României (237,5 mii km2)
din care Munţi Dealuri Câmpii
peste 2000 1 3 1500 - 2000 3 7 1000 - 1500 6 19 700 - 1000 12 36 3 500 - 700 10 16 12 300 - 500 18 12 38 1200 - 500 12 7 24 5 100 - 200 18 18 350 - 100 20 5 59
Peste 500 m 32 81 15 Sub 500 m *) 68 19 85 100 *) teritoriu afectat de aridizare şi deşertificare în cazul creşterii temperaturi medii a
aerului cu 3 0 C, prognoză până în anul 2070. Prin creşterea cu 3 0 C a temperaturii medii a aerului pe teritoriul României se
prognozează că Dobrogea, Sudul Moldovei şi o bună parte din sudul Câmpiei Române, respectiv peste 30 % din ţară va fi supusă unui proces de deşertificare şi restul de cca. 38 % unui proces de aridizare accentuată, care va cuprinde în continuare toate câmpiile noastre, până la 85 % din suprafaţa dealurilor şi aproape 20 % din munţii de la altitudini mai joase ale ţării.
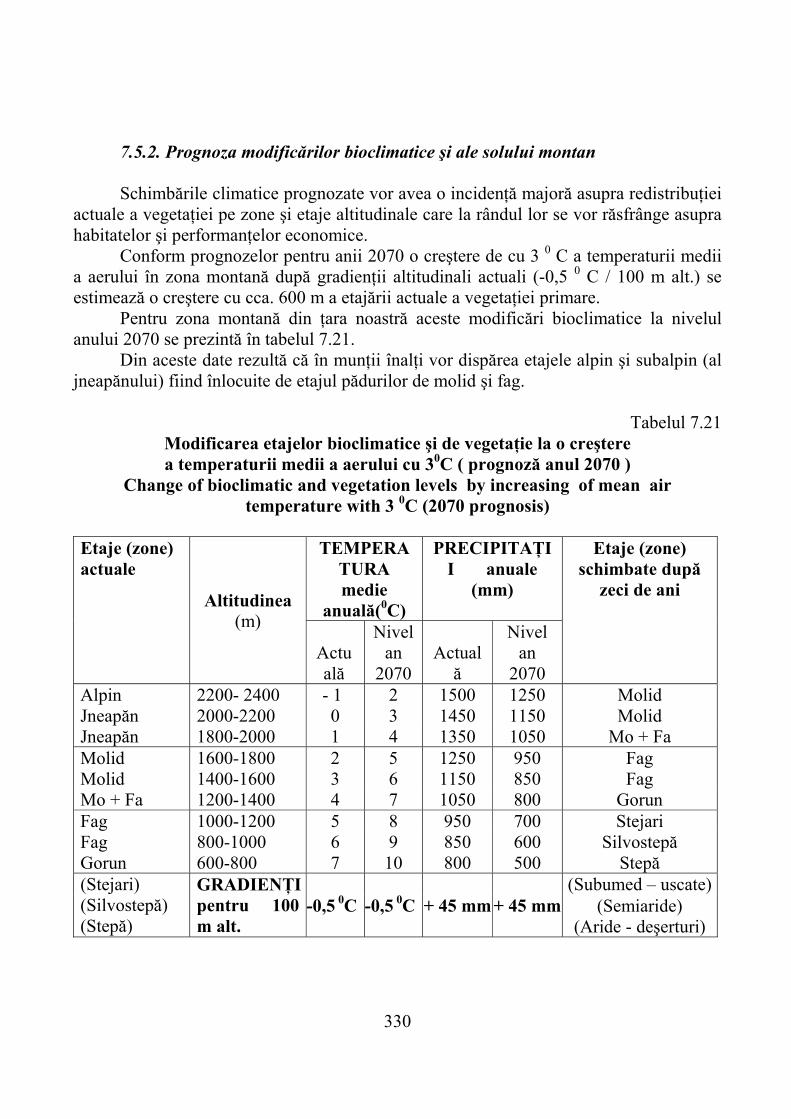
330
7.5.2. Prognoza modificărilor bioclimatice şi ale solului montan Schimbările climatice prognozate vor avea o incidenţă majoră asupra redistribuţiei
actuale a vegetaţiei pe zone şi etaje altitudinale care la rândul lor se vor răsfrânge asupra habitatelor şi performanţelor economice.
Conform prognozelor pentru anii 2070 o creştere de cu 3 0 C a temperaturii medii a aerului în zona montană după gradienţii altitudinali actuali (-0,5 0 C / 100 m alt.) se estimează o creştere cu cca. 600 m a etajării actuale a vegetaţiei primare.
Pentru zona montană din ţara noastră aceste modificări bioclimatice la nivelul anului 2070 se prezintă în tabelul 7.21.
Din aceste date rezultă că în munţii înalţi vor dispărea etajele alpin şi subalpin (al jneapănului) fiind înlocuite de etajul pădurilor de molid şi fag.
Tabelul 7.21 Modificarea etajelor bioclimatice şi de vegetaţie la o creştere a temperaturii medii a aerului cu 30C ( prognoză anul 2070 )
Change of bioclimatic and vegetation levels by increasing of mean air temperature with 3 0C (2070 prognosis)
Etaje (zone) actuale
Altitudinea (m)
TEMPERATURA medie
anuală(0C)
PRECIPITAŢII anuale
(mm)
Etaje (zone) schimbate după
zeci de ani
Actu ală
Nivel an
2070
Actuală
Nivel an
2070Alpin 2200- 2400 - 1 2 1500 1250 Molid Jneapăn 2000-2200 0 3 1450 1150 MolidJneapăn 1800-2000 1 4 1350 1050 Mo + Fa Molid 1600-1800 2 5 1250 950 Fag Molid 1400-1600 3 6 1150 850 Fag Mo + Fa 1200-1400 4 7 1050 800 Gorun Fag 1000-1200 5 8 950 700 Stejari Fag 800-1000 6 9 850 600 Silvostepă Gorun 600-800 7 10 800 500 Stepă(Stejari) (Silvostepă) (Stepă)
GRADIENŢI pentru 100 m alt.
-0,5 0C -0,5 0C + 45 mm+ 45 mm (Subumed – uscate)
(Semiaride) (Aride - deşerturi)
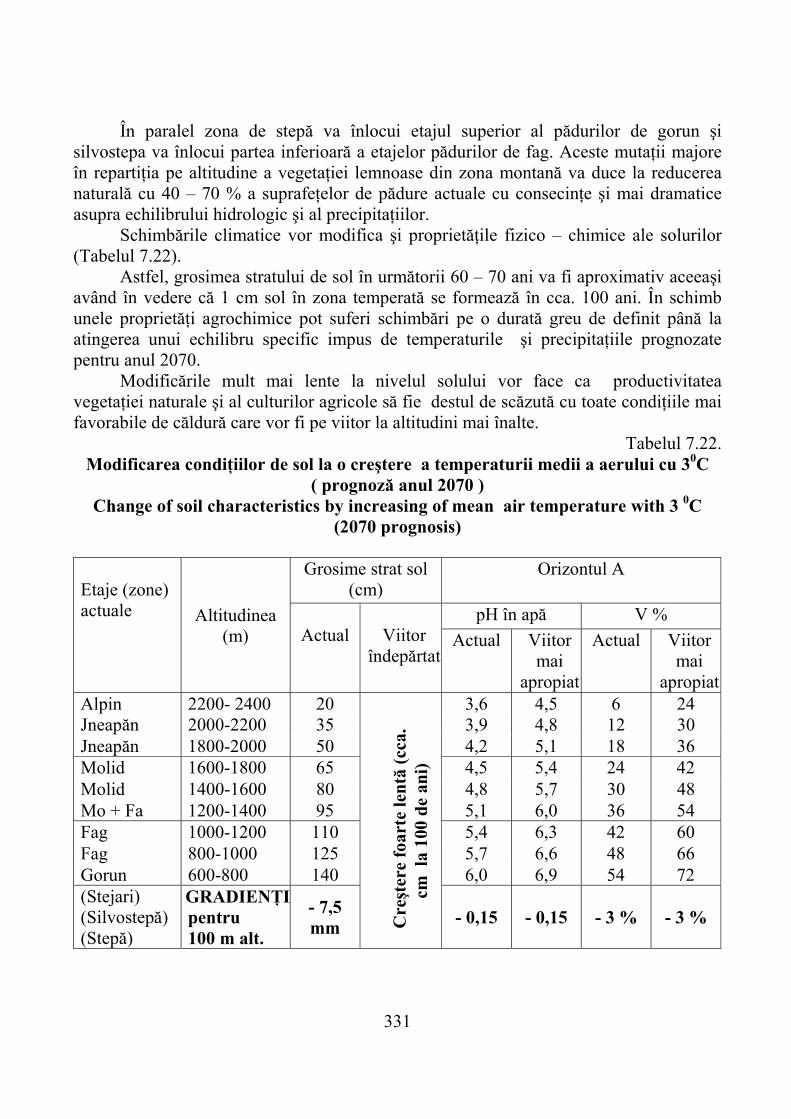
331
În paralel zona de stepă va înlocui etajul superior al pădurilor de gorun şi silvostepa va înlocui partea inferioară a etajelor pădurilor de fag. Aceste mutaţii majore în repartiţia pe altitudine a vegetaţiei lemnoase din zona montană va duce la reducerea naturală cu 40 – 70 % a suprafeţelor de pădure actuale cu consecinţe şi mai dramatice asupra echilibrului hidrologic şi al precipitaţiilor.
Schimbările climatice vor modifica şi proprietăţile fizico – chimice ale solurilor (Tabelul 7.22).
Astfel, grosimea stratului de sol în următorii 60 – 70 ani va fi aproximativ aceeaşi având în vedere că 1 cm sol în zona temperată se formează în cca. 100 ani. În schimb unele proprietăţi agrochimice pot suferi schimbări pe o durată greu de definit până la atingerea unui echilibru specific impus de temperaturile şi precipitaţiile prognozate pentru anul 2070.
Modificările mult mai lente la nivelul solului vor face ca productivitatea vegetaţiei naturale şi al culturilor agricole să fie destul de scăzută cu toate condiţiile mai favorabile de căldură care vor fi pe viitor la altitudini mai înalte.
Tabelul 7.22. Modificarea condiţiilor de sol la o creştere a temperaturii medii a aerului cu 30C
( prognoză anul 2070 ) Change of soil characteristics by increasing of mean air temperature with 3 0C
(2070 prognosis)
Etaje (zone) actuale Altitudinea
(m)
Grosime strat sol (cm)
Orizontul A
Actual
Viitor
îndepărtat
pH în apă V %
Actual Viitor mai
apropiat
Actual Viitor mai
apropiatAlpin 2200- 2400 20 3,6 4,5 6 24 Jneapăn 2000-2200 35
C
reşt
ere
foar
te le
ntă
(cc
a.
cm l
a 10
0 d
e an
i)
3,9 4,8 12 30Jneapăn 1800-2000 50 4,2 5,1 18 36 Molid 1600-1800 65 4,5 5,4 24 42 Molid 1400-1600 80 4,8 5,7 30 48Mo + Fa 1200-1400 95 5,1 6,0 36 54 Fag 1000-1200 110 5,4 6,3 42 60 Fag 800-1000 125 5,7 6,6 48 66 Gorun 600-800 140 6,0 6,9 54 72 (Stejari) (Silvostepă) (Stepă)
GRADIENŢIpentru 100 m alt.
- 7,5 mm
- 0,15 - 0,15 - 3 % - 3 %
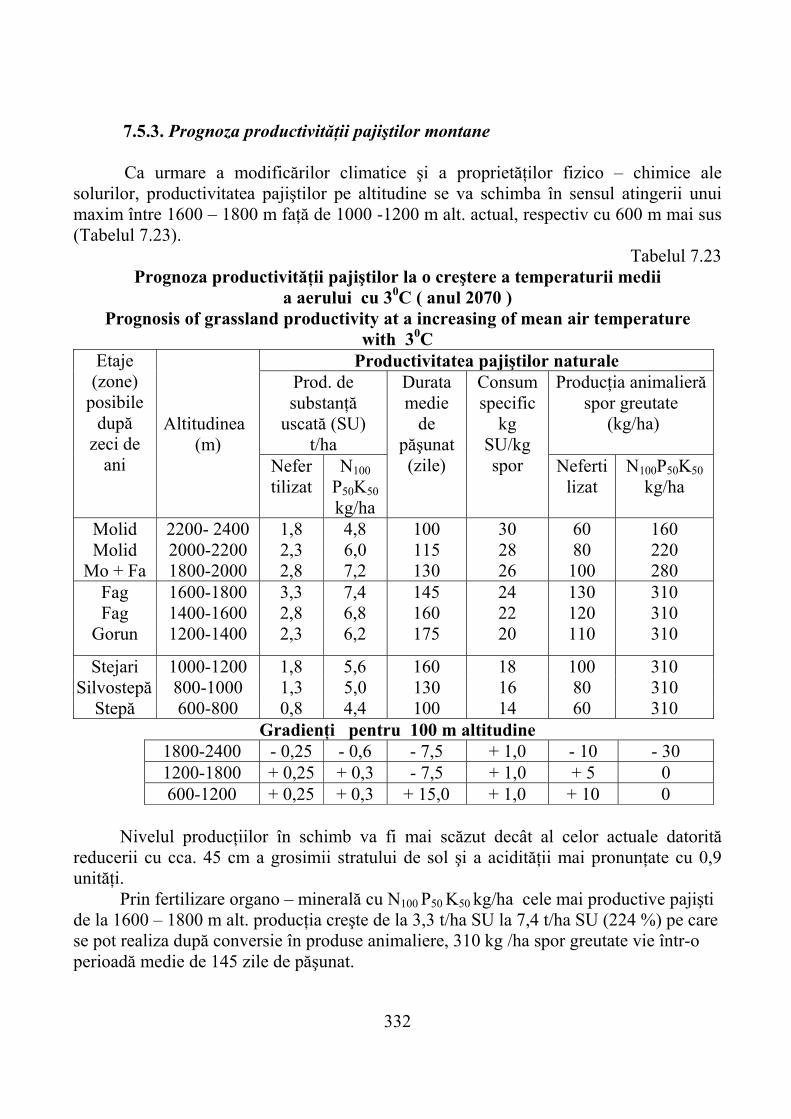
332
7.5.3. Prognoza productivităţii pajiştilor montane
Ca urmare a modificărilor climatice şi a proprietăţilor fizico – chimice ale solurilor, productivitatea pajiştilor pe altitudine se va schimba în sensul atingerii unui maxim între 1600 – 1800 m faţă de 1000 -1200 m alt. actual, respectiv cu 600 m mai sus (Tabelul 7.23).
Tabelul 7.23 Prognoza productivităţii pajiştilor la o creştere a temperaturii medii
a aerului cu 30C ( anul 2070 ) Prognosis of grassland productivity at a increasing of mean air temperature
with 30C Etaje (zone)
posibile după
zeci de ani
Altitudinea (m)
Productivitatea pajiştilor naturale Prod. de substanţă
uscată (SU) t/ha
Durata medie
de păşunat (zile)
Consum specific
kg SU/kg spor
Producţia animalieră spor greutate
(kg/ha)
Nefer tilizat
N100
P50K50 kg/ha
Nefertilizat
N100P50K50 kg/ha
Molid 2200- 2400 1,8 4,8 100 30 60 160 Molid 2000-2200 2,3 6,0 115 28 80 220
Mo + Fa 1800-2000 2,8 7,2 130 26 100 280 Fag 1600-1800 3,3 7,4 145 24 130 310 Fag 1400-1600 2,8 6,8 160 22 120 310
Gorun 1200-1400 2,3 6,2 175 20 110 310
Stejari 1000-1200 1,8 5,6 160 18 100 310 Silvostepă 800-1000 1,3 5,0 130 16 80 310
Stepă 600-800 0,8 4,4 100 14 60 310 Gradienţi pentru 100 m altitudine
1800-2400 - 0,25 - 0,6 - 7,5 + 1,0 - 10 - 30 1200-1800 + 0,25 + 0,3 - 7,5 + 1,0 + 5 0 600-1200 + 0,25 + 0,3 + 15,0 + 1,0 + 10 0
Nivelul producţiilor în schimb va fi mai scăzut decât al celor actuale datorită
reducerii cu cca. 45 cm a grosimii stratului de sol şi a acidităţii mai pronunţate cu 0,9 unităţi.
Prin fertilizare organo – minerală cu N100 P50 K50 kg/ha cele mai productive pajişti de la 1600 – 1800 m alt. producţia creşte de la 3,3 t/ha SU la 7,4 t/ha SU (224 %) pe care se pot realiza după conversie în produse animaliere, 310 kg /ha spor greutate vie într-o perioadă medie de 145 zile de păşunat.

333
Durata perioadei de păşunat variază între 100 zile la 600 - 800 m alt. datorită perioadei de uscăciune, la fel şi pe ecartul 2200 – 2400 m alt. unde intervine o perioadă de vegetaţie mai scurtă. Durata maximă de păşunat de 175 zile se atinge la 1200-1400 m altitudine.
Datorită altitudinii, consumul specific pentru 1 kg spor creşte cu 1 kg SU pentru fiecare 100 m alt., respectiv de la 14 kg la 600 – 800 m la 30 kg la 2200 – 2400 m alt.
Ca urmare a scăderii temperaturilor active pe altitudine şi al creşterii cantităţii de precipitaţii se creează un echilibru căldură – umiditate între 600 – 1800 m alt., interval între care productivitatea pajiştilor exprimată în spor greutate vie rămâne aproape constantă fiind de 310 kg /ha pe suprafeţele fertilizate la un nivel mediu.
Condiţiile de sol şi climă din zona montană şi mai nefavorabile pe altitudine pentru culturile tradiţionale agricole, impune dezvoltarea creşterii animalelor erbivore pe pajiştile naturale mai performante şi practicarea pe scară mai largă a agroturismului, asemănător ţărilor alpine.
Datorită aridizării climatului este necesară implementarea şi extinderea sistemului agrosilvopastoral prin care au de câştigat atât covorul ierbos cât şi animalele de pe păşune.

334
BIBLIOGRAFIE 1. Aerts R., Caluwe H., Beltman B., 2003, Plant community mediated vs. nutritional
controls on litter decomposition rates in grasslands, Ecology, 84, 3198-3208
2. Amundson R.G., Chadwick O.A., Sowers J.M., 1989, A comparision of soil climate and biological activity along an elevational gradient in the eastern Mojave Desert, Oecologia, 80, 395-400
3. Anghel Gh., 1960, Geobotanica aplicată în studiul pajiştilor şi al terenurilor cultivate, Comunicări de botanică, Soc. Şt. Nat. Geogr., Bucureşti
4. Anghel Gh. ş.a., 1965, Studiul şi cartarea păşunilor din corpul Crinţ - Munţii Cibinului, Comunicări de botanică, vol. III
5. Anghel Gh.şi colab., 1967, Cultura pajiştilor, Ed. Agrosilvică, Bucureşti
6. Anghel Gh., Răvăruţ M., Turcu Gh., 1971, Geobotanica, Editura Ceres, Bucureşti
7. Anghel Gh., Motcă Gh., 1975, Principii de bază în elaborarea tipologiei pajiştilor din R.S. România, Lucrări ştiinţifice S.C.C.P. Măgurele-Braşov, vol.I
8. Anghel Gh., C. Bărbulescu, A. Bârsan, 1977: Pajiştile din Platforma Bran, Casa Agronomului, Braşov
9. Anghel Gh., Doniţă N.,1980, Etajarea vegetaţiei lemnoase şi ierboase în teritoriul carpatic, Staţiunea centrală de cercetări pentru cultura pajiştilor Măgurele-Braşov-10 ani de activitate, Casa agronomului-Braşov
10. Anghel Gh., 1981, Scheme pentru clasificarea vegetaţiei pajiştilor permanente (naturale, seminaturale) din R.S.România, Lucrări ştiinţifice, S.C.C.P. Măgurele-Braşov, vol. VII
11. Anghel Gh., Neacşu Marcela, Vasu Alexandra, 1982, Etajarea vegetaţiei ierboase din Munţii Cibinului (Munţii Cindrel), Lucr. şt. ICPCP Braşov, vol. VIII, pag. 3-17, Buc.
12. Anghel Gh., 1984, Pajişti intensive. Ed. Ceres, Bucureşti
13. Atkin O.K.,Cummins W.R, 1994, The effect of nitrogen source on growth, nitrogen economy and respiration of two high arctic plant species differing in relative growth rate, Functional Ecology, 8, 389-399
14. Austin M.P., 2005, Vegetation and environment: discontinuities and continuities, Pages 52-84 in E. van der Maarel, editor. Vegetation ecology. Blackwell Publishing, Oxford

335
15. Bacinschi D., Neacşa O., 1985, Principalele caracteristici climatice ale Carpaţilor româneşti, Lucr. Şt. ale ICPCP Măgurele-Braşov, vol. X, pag. 33-74, Red. prop. tehn. Agr. Bucureşti
16. Badea L., 1967, Subcarpaţii dintre Cerna Olteţului şi Gilort, Studiu de geomorfologie, Ed. Acad. Bucureşti
17. Balsberg-Pahlsson A.M.,1995, Growth, radicle and root hair development of Deschampsia flexuosa (L) Trin seedlings in relation to soil acidity, Plant and Soil, 175, 125-132
18. Banaticla M.C.N., Buot I.E., 2005, Altitudinal zona on of pteridophytes on Mt. Banahaw de Lucban, Luzon Island, Philippines, Plant Ecology, 180,135-151
19. Barbu N., 1976, Obcinile Bucovinei, Ed. Ştiinţifică şi Enciclopedică, Bucureşti
20. Bartsch M. şi al., 1940, Vegetationskunde des Schwarzwaldes Pflanzensoziologie, Band 4, Verlag Gustav Fischer, Jena
21. Băcăinţan N., 1999, Munţii Baraolt- Studiu Geomorfologic, Ed. Acad. Rom. Bucureşti, 160 p.
22. Bălăceanu V., Marian Elisabeta, 1985, Soils of the Romanian Carpathians, Lucrările ştiinţifice ale Institutului de Cercetare şi Producţie pentru Cultura Pajiştilor Măgurele-Braşov vol. X, Bucureşti
23. Bărbos M.I., 2007, Studii privind cenologia şi ecologia pajiştilor montane din judeţul Maramureş, Universitatea "Babeş-Bolyai", Cluj Napoca
24. Bărbulescu C., 1961, Păşunile alpine din Munţii Iezer- Păpuşa, Natura, nr.3
25. Bărbulescu C., Burcea ., 1970, Determinator pentru flora pajiştilor, Ed. Ceres, Bucureşti
26. Bărbulescu Viorica, 1975, Cercetări privind relaţia dintre solul şi vegetaţia pajiştilor naturale din Platforma Bran, I.A.N.B.- Bucureşti
27. Bărbulescu C., Burcea P., Bărbulescu Viorica, Motcă Gh., Ştefănescu Al., 1978, Pajiştile permanente, în “Pajiştile din Platforma Bran”, Casa agronomului- Braşov
28. Bărbulescu C., Motcă Gh., Pop M., Capşa Stela, Belu C., Neagu M., 1982, Efectul îngrăşămintelor chimice pe păşunile munţilor înalţi în funcţie de altitudine, Lucr. şt. ale ICPCP Braşov, vol. VIII, pag. 19-36, Red. prop. tehn. agr. Bucureşti
29. Bărbulescu C., Motcă Gh., 1983, Păşunile munţilor înalţi, Ed. Ceres, Bucureşti
30. Bărbulescu, C., Motcă Gh.,1987, Pajiştile de deal din România, Ed. “Ceres” Bucureşti;
31. Bărbulescu C. şi colab., 1991, Cultura pajiştilor şi a plantelor furajere, Ed. Didactică şi Pedagogică, Bucureşti

336
32. Bâlteanu D., 1983, Experimentul de teren în geomorfologie. Aplicaţii la Subcarpaţii Buzăului, Ed. Acad., Bucureşti
33. Begon M., Townsend C.R., Harper J.L., 2006, Ecology: from individuals to ecosystems, Blackwell Publishing Ltd.
34. Beldie Al., 1967, Flora şi vegetaţia munţilor Bucegi, Editura Academiei R.S.R.
35. Beldie Al., 1977, 1979 Flora României. Determinator ilustrat al plantelor vasculare, vol I şi II. Editura Academiei R.S.R
36. Belu C., 1982, Valorificarea păşunilor alpine din Munţii Cernatului-Masivul Făgăraş, Teză de doctorat, Institutul agronomic „N. Bălcescu”, Bucureşti
37. Berendse F., Lammerts E.J., Olff H., 1998, Soil organic matter accumulation and its implications for nitrogen mineralization and plant species composition during succesion in coastal dune slacks, Plant Ecology, 137, 71-78
38. Biala K., Zyszkowska M., 2004, Effects of changes in grassland management on biodiversity and landscape in the Sudeten Mountains in Poland, Grassland Science in Europe, vol.9, pg. 210-212
39. Blaga G., Filipov F., Udrescu S., Rusu I., Vasile D., 2005, Pedologie, Editura AcademicPres, Cluj Napoca
40. Bleahu M., Rusu T., 1965, Carstul din România, Lucrările Instit. de speol. E. Racoviţă", tom IV
41. Bojoi I., 2000, România, Geografie fizică, Ed. Univ. Al. I. Cuza, Iaşi
42. Borza Al., 1934, Studii fitosociologice în Munţii Retezat. Bul. grăd. bot., Cluj, vol.XI
43. Borza Al., 1934, Buletinul Grădinii Botanicii şi Muzeul botanic, vol. XIV, Cluj
44. Borza Al., 1959, Flora şi vegetaţia Văii Sebeşului, Ed. Academiei RPR, Bucureşti
45. Borza. AL., 1960, Elementele florei şi răspândirea lor, Monografia geografică a R.P.R., vol. I Geografia fizică", Bucureşti
46. Borza Al., Boşcaiu M., 1965, Introducere în studiul covorului vegetal. Editura Academiei R.S.R., Bucureşti
47. Bosy J. L., Reader R.J., 1995, Mechanisms underlying the suppression of forb seedling emergence by grass (Poa pratensis) litter, Functional Ecology, 9, 635-639
48. Boşcaiu N., 1971, Flora şi vegetaţia Munţilor Ţarcu, Godeanu şi Cernei. Editura Academiei R.S.R., Bucureşti
49. Boşcaiu N., 1985, Criterii pentru constituirea şi gestionarea ecologică a rezervaţiilor botanice, Ocrotirea naturii şi mediului înconjurător, 28, 2, 126-135

337
50. Botnariuc N., Vădineanu A., 1982, Ecologie, Editura Didactică şi Pedagogică, Bucureşti
51. Bradshaw A.D., Lodge R.W., Jowett D., Chadwick M.J., 1958, Experimental Investigations Into the Mineral-Nutrition of Several Grass Species .1. Calcium Level, Journal of Ecology, 46, 749-757
52. 52.Braun – Blanquet J., 1964, Pflanzensoziologie, III, Aufl. Springer, Verlag, Wien, New York
53. Brânduş C., 1981, Subcarpaţii Tazlăului. Studiu Geomorfologic, Ed. Acad. R.S.R., Bucureşti
54. Bruun H.H., Moen J., Virtanen R., Grytnes J.A., Oksanen L., Angerbjorn A., 2006, Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities, Journal of Vegetation Science, 17, 37-46
55. Bujorean G., 1930, Contribuţii la cunoaşterea succesiunii şi întovărăşirii plantelor, Bul. Grădinii şi Muzeului Botanic, vol. X, Cluj-Napoca;
56. Buia Al., 1943, Contribuţii la studiul fitosociologic al păşunilor din Munţii Carpaţi, Bul. Fac. Agronom., Cluj, X
57. Buia Al., Păun M., Safta I., Pop M., 1959, Contribuţii geobotanice asupra păşunilor şi fîneţelor din Oltenia, Inst. Agr. “T. Vladimirescu” Craiova, Lucrările ştiinţifice, 90p.
58. Buia Al., Pavel C., Safta I., 1962, Pajiştile din Masivul Parîng şi îmbunătăţirea lor, Ed. Agrosilvică, Bucureşti
59. Buia Al., Păun M., Pavel C.,1962, Studiul geobotanic al pajiştilor. Pajiştile din Masivul Parîng şi îmbunătăţirea lor, Ed. Agro-Silvică, Bucureşti, 143-274
60. Buia Al., 1963, Les association de Nardus stricta de la R.P.R., Revista de biologie, seria Botanică, 8, 2, 119-137
61. Burcea P., 1965, Cercetări asupra fîneţelor naturale dintre rîul Cibin şi rîul Dobra, raionul Sibiu, Teză de doctorat, I.A.N.B., Bucureşti
62. Burduja C., Dobrescu C.,Grîneanu A., Răvăruţ M., Căzăceanu I., Bîrcă C., Raclaru P., Turenschi E.,1956, Contribuţii la cunoaşterea pajiştilor naturale din Moldova sub raport geobotanic şi agroproductiv, Studii şi cercetări şt. biol. agr., tom. VII, 1
63. Buza M., 1978, Munţii Cindrelului – Studii de geografie fizică, Teză de doctorat, Inst. de Geografie, Academia României
64. Buza M., 2000, Munţii Cindrelului, Studiu geoecologic, Ed. Univ. "Lucian Blaga" Sibiu, 135 p.

338
65. Candrea B., Candrea P., Niţă M.D., 2008, Harta unităţilor de relief din România, http://earth.unibuc.ro/
66. Cardaşol V., Maruşca T., 1975, Importanţa pajiştilor semănate pentru intensivizarea producţiei de furaje pe solurile cu exces de umiditate freatică, Revista de creşterea animalelor, Bucureşti, nr. 11, 31-38
67. Cardaşol, V., Daniliuc D., 1979, Contribuţii la punerea în valoare a pajiştilor din Munţii Cibin, Lucrări ştiinţifice ale SCCP Măgurele Braşov, vol. V, pag. 137-142
68. 68.Cardaşol V., Pop M., Razec I., Brezean, D., 1985, Pajiştile din Munţii Bucegi, Lucrări ştiinţifice ale In stitutului de Cercetare şi Producţie pentru Cultura Pajiştilor Măgurele – Braşov, vol.X, 181-202
69. Carson W. P.,Peterson C.J., 1990, The role of litter in an old-field community - impact of litter quantity in different seasons on plant-species richness et abundance, Oecologia, 85, 8-13
70. Călinescu R. şi colab., 1969, Biogeografia României, Ed. Ştiinţifică, Bucureşti
71. Cernelea I., 1974, Cercetări privind pajişitile de Agrostis tenuis din Depresiunea Haţegului şi posibilităţi de sporire a producţiei prin aplicarea îngrăşămintelor, Rezumat al tezei de doctorat, IANB, Bucureşti
72. Chadwick M. J., 1960, Biological Flora of the British Isles: Nardus stricta, L.J.E. col.48
73. Chiriţă C.D., 1955, Pedologie generală, Editura Agro-Silvică de Stat, Bucuresti
74. Chiriţă C. şi col., 1964, Fundamentele naturalistice şi metodologice ale tipologiei şi cartării staţionare forestiere, Ed. Academia RPR, Bucureşti
75. Chiriţă C., 1974, Ecopedologie cu baze de pedologie generală, Ed. Ceres, Bucureşti
76. Ciocârdel R., 1957, Hidrogeologie, Ed. Tehnică, Bucureşti
77. Ciortea G., 2000, Ameliorarea producţiei pajiştilor de munte prin îmbunătăţirea regimului de nutriţie, Teză de doctorat, USAMVB Timişoara
78. Ciubotaru C., 1974, Contribuţii la studiul florei, vegetaţiei şi îmbunătăţirii pajiştilor naturale din Depresiunea Liteni, jud. Suceava, Rezumat al tezei de doctorat, Inst. Agron.Iasi
79. Ciucă Maria, 1984, Vegetaţia pajiştilor din masivul Ciucaş, Ed. Academiei, Bucureşti
80. Coldea Gh., Pînzaru Gh., 1967, Vegetaţia rezervaţiei naturale “Omul” din Munţii Suhardului,St. şi Cerc. Biol., Seria Biol.Veget., Bucureşti, 39, 4, 109-118

339
81. Coldea Gh., 1972, Flora şi vegetaţia Munţilor Plopiş, Teză de doctorat, Cluj
82. Coldea Gh., 1984, L, etude des paturages basiphyles sousalpins des monts Rodnei (Les Carpates Orientales), Contrib. Bot., Cluj-Napoca, 137-154
83. Coldea Gh.,1990, Munţii Rodnei. Studiu geobotanic, Ed. Academiei, Bucureşti
84. Coldea Gh., 1991, Documents phytosociologiques. Prodrome des associations vegetales des Carpates du sud-est (Carpates Roumaines), Camerino Universita degli Studi
85. Coldea Gh., Contribuţii la studiul vegetaţiei României (I), Contrib. Bot., Cluj-Napoca, 1-8, 1995-1996
86. Coldea Gh., Sanda V., Popescu A.,Ştefan N., 1997, Les associations vegetales de Roumanie, Tome 1, Les associations herbacees naturelles, Presses Universitaires de Cluj, 261 p.
87. Common T.G., Hunter E.A., Floate M.J.S., Eadie J., Hodgson J., 1991, The long-term effects of a range of pasture treatments applied to three semi-natural hill grassland communities. 1. Pasture production and botanical composition, Grass and Forage Science, 46, 239-251
88. Cornish P.S., Raison R.J., 1977, Effects of phosphorus and plants on nitrogen mineralization in three grassland soils, Plant and Soil, 47, 289-295
89. Coteţ P., 1973, Geomorfologia României, Ed. Tehnică, Bucureşti
90. Crăciun C., 2000, Mineralele argiloase din sol- Implicaţii în agricultură, Ed. G.N.P. Minischool, Bucureşti
91. Cristea V., 1991, Fitocenologie şi vegetaţia României; Îndrumător de lucrări practice, Universitatea Babeş-Bolyai, Cluj Napoca
92. Cristea V., Denaeyer S., 2004, De la biodiversitate la OGM-uri?, Editura Eikon, Cluj Napoca
93. Csuros St., Csuros-Kaptalan Margareta, 1966, Caracterizarea unor asociaţii de plante din Transilvania pe baza indicilor ecologici, Contribuţii botanice, Cluj, 2, 163-179
94. Curtis C. J., Emmett B. A., Grant H., Kernan M., Reynolds B., Shilland E., 2005, Nitrogen saturation in UK moorlands: the critical role of bryophytes et lichens in determining retention of atmospheric N deposition, Journal of Applied Ecology, 42, 507-517
95. Daget Ph., Poissonet J., 1969, Publication, CEPE Montpellier
96. Daget Ph., Poissonet J., 1971, Annuaire Agronomique, nr. 22, Paris
97. Dihoru Gh., 1975, Învelişul vegetal din muntele Siriu, Editura Academiei R.S.R.

340
98. Dix R.L., 1958, Some slope-plant relationships in the grasslands of the Little Missouri Badlands of Noth Dakota. Journal of Range Management, 11, 88-92
99. Dolezal J., Srutek M., 2002, Altitudinal changes in composition and structure of mountain-temperate vegetation: a case study from the Western Carpathians, Plant Ecology, 158, 201-221
100. Donisă I., 1968, Geomorfologia văii Bistriţei, Ed. Acad. R.S.R., Bucureşti
101. Doniţă, N., Doina Ivan, 1982, Producţia primară a vegetaţiei terestre din R. S. România, Probleme moderne de ecologie, pag. 294-305, Ed. Ştiinţ. şi enciclopedică, Buc.
102. Doniţă N., Ivan D., 1985, Vegetaţia Carpaţilor, Pajiştile din Carpaţii României, Lucr. şt. ale Inst. de Cerc. şi Producţie pentru Cultura Pajiştilor Măgurele – Braşov, Vol.X, 109-140
103. Doniţă N., Chiriţă C., Stănescu V.ş. a., 1990, Tipuri de ecosisteme forestiere din România, ICAS, Red. de prop. tehn. agr. Bucureşti
104. Doniţă N., Popescu A., Paucă-Comănescu M., Mihăilescu S., Biriş I.A., 2005, Habitatele din România, Editura Tehnică Silvică, Bucureşti
105. Dragomir N., 2005, Pajişti şi plante furajere, Tehnologii de cultivare, Editura Eurobit, ISBN 973-620-115-5
106. Dumitru M., 1980, Cercetări asupra florei şi vegetaţiei Subcarpaţilor dintre Ialomiţa şi Dîmboviţa, Rezumat al tezei de doctorat, Univ. Bucureşti
107. Eckstein R. L., Donath T. W., 2005, Interactions between litter et water availability affect seedling emergence in four familial pairs of floodplain species, Journal of Ecology, 93, 807-816
108. Ellenberg H., 1963, Vegetation Mitteleuropas Mit Den Alpen, Eugen Ulmer, Stuttgart
109. Evans G.C.m 1969, Spectral composition of light in field . Its measurement and ecological importance, Journal Of Ecology, 57, 109-&
110. Facelli J. M., Pickett S.T.A., 1991, Plant litter - light interception et effects on an old-field plant community, Ecology, 72, 1024-1031
111. Facelli J. M.,Carson W.P., 1991, Heterogeneity of plant litter accumulation in successional communities, Bulletin of the Torrey Botanical Club, 118, 62-66
112. Fageria N.K., 1974, Absorption of magnesium and its influence on uptake of phosphorus, potassium, and calcium by intact groundnut plants, Plant and Soil, 40, 313-320

341
113. Fageria N.K., Baligar V.C., Wright R.J., 1989, Growth and nutrient concentrations of alfalfa and common bean as influenced by soil acidity, Plant and Soil, 119, 331-333
114. Filip, N. F., 1984, Cercetări privind solurile şi staţiunile forestiere din Podişul Târnavelor, Teză de doctorat, Universitatea Braşov
115. Fitter A.H., Graves J.D., Self G.K., Brown T.K., Bogie D.S., Taylor K., 1998, Root production, turnover and respiration under two grassland types along an altitudinal gradient: influence of temperature and solar radiation, Oecologia, 114, 20-30
116. Florea N., Munteanu I, Rapaport C., Chiţu C., Opriş M.,1968, Geografia solurilor României, Ed. Ştiinţifică, Bucureşti
117. Florea N., Munteanu I., 2003, Sistemul român de taxonomie a solurilor (SRTS), Ed, Estfalia, Bucureşti, 182 p.
118. Florea N., Gheorghe Maria 2008, Unităţile pedoagrotehnologice de teren. Cerinţe specifice, Ştiinţa solului nr. 1, vol XLII
119. Florea N. şi colab., 2009, Gruparea terenurilor din România după specificul cerinţelor tehnologice de management, Lucrările celei de a XIX-a Conferinţe Naţionale pentru Ştiinţa Solului, Iaşi
120. Florescu, I., Nicolaescu N.V., 1996, Silvicultura, vol. I, Studiul pădurii Editura Lux Libris-Braşov;
121. Gârda N., 2010, Studiul elementelor de landşaft montan (cu privire specială asupra ecosistemelor de pajişti din comuna Gârda de Sus, Munţii Apuseni), Teză de doctorat, U.S.A.M.V. Cluj-Napoca
122. Foster B.L., Gross K.L.,1998, Species richness in a successional grassland: effects of nitrogen enrichment et plant litter, Ecology, 79, 2593-2602
123. Gârbacea V., 1956, Piemontul Călimanilor, Studii şi cercetări de geologie geogr., VII, 1-4, Acad.R.P.R., Fil. Cluj
124. Gheorghe Maria, Florea N., 2006, Harta unităţilor pedo-agrotehnologice din Câmpia Română de Est, a 18-a Conferinţă Naţ. Rom. de Şt. Solului, Cluj Napoca
125. Gheorghe Maria, 2007, Gruparea solurilor agricole în funcţie de specificul lor care determină diferenţierea lucrărilor agrotehnice cu aplicaţie în partea de est a Câmpiei Române, Ed. Ceres, Bucureşti, 226 pag. (teză de doctorat, USAMV Bucureşti, 2006)
126. Gibson D. J., Ely J. S., Collins S.L., 1999, The core-satellite species hypothesis provides a theoretical basis for Grime's classification of dominant, subordinate, et transient species, Journal of Ecology, 87, 1064-1067

342
127. Gigon A., Rorison I.H., 1972, Response of some ecologically distinct plant species to nitrate-nitrogen and to ammonium-nitrogen, Journal of Ecology, 60, 93-&
128. Giurcăneanu C., 1988, Populaţia şi aşezările din Carpaţii româneşti, Ed. ştiinţifică şi enciclopedică, Bucureşti
129. Granger J.E.,Schulze R.E. ,1977, Incoming solar-radiation patterns and vegetation response - examples from Natal Drakensberg, Vegetatio, 35, 47-54
130. Greszta J., Gruszka A., Wachalewski T., 1992, Humus degradation under the influence of simulated "acid rain", Water, Air, and Soil Pollution, 63, 51-66
131. Grigoraş C., 1999, Solurile României, Ed. Universitatea din Craiova Facultatea de Ştiinţe, Secţia Geografie
132. Grime J. P., 1984, Dominant et subordinate components of plant communities: implications for succesion, stability et diversity, Pages 413-428 in AJ Gray, MJ Crawley, et PJ Edwards, editors. Colonization, Succesion et Stability. Blackwell Scientific Publications, Oxford
133. Grime J. P., 1998, Benefits of plant diversity to ecosystems: immediate, filter et founder effects, Journal of Ecology, 86, 902-910
134. Grime J.P., Hodgson J.G., Hunt R., 1996, Comparative plant ecology. A functional approach to common British specie, Chapman and Hall, London
135. Grime J.P., 2001, Plant strategies, vegetation processes, and ecosystem properties, John Wiley & Sons, Ltd.
136. Grîneanu A.,Popovici D., Pleşa D., 1967, Zece ani de activitate în sprijinul producţiei agricole în Câmpia Moldovei, Revista agricolă, Bucureşti
137. Grytnes J.A., 2003, Species-richness patterns of vascular plants along seven altitudinal transects in Norway, Ecography, 26, 291-300
138. Gusewell S., 2004, N : P ratios in terrestrial plants: variation and functional significance, New Phytologist, 164, 243-266
139. Guş P., 1976, Contribuţii la cunoaşterea vegetaţiei şi a solurilor cu exces de umiditate din Valea Sieului, Rezumat al tezei de doctorat, Inst. Agric., Cluj
140. Hector A., Beale A. J., Minns A., Otway S. J., J. H. Lawton J.H., 2000, Consequences of the reduction of plant diversity for litter decomposition: effects through litter quality et microenvironment, Oikos, 90, 357-371
141. Hejcman M., Pavlusi V., Gaisler J., 2004, Vegetation structure under unmanaged, grassed and cut grasslands in the Giant Mountains, Czech Republic, Grassland Science in Europe, vol.9, pg. 213-215

343
142. Hejcman M., Klaudisova M., Schellberg J., Honsova D., 2007, The regen grassland experiment: Plant species composition after 64 years of fertilizer application Agriculture, Ecosystems & Environment, 122, 259-266
143. Hoorens B., Aerts R., Stroetenga M., 2002, Litter quality et interactive effects in litter mixtures: more negative interactions under elevated CO2? Journal of Ecology, 90, 1009-1016
144. Hoorens B., Aerts R., Stroetenga M., 2003, Does initial litter chemistry explain litter mixture effects on decomposition? Oecologia, 137, 578-586
145. Hopkins D.W., Ibrahim D.M., Odonnell A.G., Shiel R.S., 1990, Decomposition of cellulose, soil organic-matter and plant litter in a temperate grassland soil, Plant and Soil, 124, 79-85
146. Hund R., 1964, Die Bergwisen des Harzes des Thuringer Waldes und des Erzgebirges Pflanzensoziologie, 14, Jena
147. Hund R., 1966, Ekologisch – geobotanische Untersuchungen an Pflanzen der mittel europaischen, Wiesen, Jena
148. Huntjens J.L.M.,Albers R.A.J.M., 1978, A model experiment to study influence of living plants on accumulation of soil organic-matter in pastures, Plant and Soil, 50, 411-418
149. Iacob T., Vîntu V., Samuil C., 2000, Tehnologia producerii şi conservării furajelor, Ed. „Ion Ionescu de la Brad”, Iaşi
150. Iancu M. şi col., 1971, Judeţul Braşov, Ed. Academiei RSR, Bucureşti
151. 152. Ionescu I., şi colab., 2001, Pajiştile permanente din Nordul Olteniei, Ed. Universitatea Craiova
152. Ivan Doina, 1979, Fitocenologie şi vegetaţia R.S.R., Editura tehnică şi pedagogică, Bucureşti
153. Jeangros B.,Thomet P., 2004, Multifunctionality of grassland systems in Switzerland, Grass and Science in Europe, vol.9, pg. 11-23
154. Keizer P.J., Vantooren B.F., During H.J., 1985, Effects of bryophytes on seedling emergence and establishment of short-lived forbs in chalk grassland, Journal of Ecology, 73, 493-504
155. King I., Nicholson I. A., 1964, The vegetation of Scotland, Olivier and Bayd, Edinburg
156. Kirkpatrick J.B.,Nunez M., 1980, Vegetation-radiation relationships in mountainous terrain - eucalypt-dominated vegetation in the Risdon Hills,. Tasmania, Journal of Biogeography, 7, 197-208

344
157. Kirkpatrick J.B., Fensham R.J., Nunez M., Bowman D.M.J.S., 1988, Vegetation-radiation relationships in the wet-dry tropics - Granite Hills in Northern Australia. Vegetatio, 76, 103-112
158. Klapp E., 1965, Grunland Vegetation und Standort, Paul- Parey, Berlin, Hamburg
159. Klapp E., 1971, Wiesen und Weidwn, Ed. IV, Berlin-Hamburg
160. Klimek S., Kemmermann A.R., Hofmann M., Isselstein J., 2007, Plant species richness and composition in managed grasslands: The relative importance of field management and environmental factors, Biological Conservation, 134, 559-570
161. Kormondy E., 1996, Concepts of ecology, Prentice Hall
162. Kovacs At., 1978, Bonitarea pajiştilor, Biblioteca agricolă A.S.A.S., Bucureşti
163. Kovacs At., 1979, Flora şi vegetaţia Munţilor Bodoc (jud.Covasna, Studiu taxonomic, geobotanic şi economic, Teză de doctorat, Cluj-Napoca
164. Kovacs At., 1979, Indicatorii biologici, ecologici şi economici ai florei pajiştilor, Redacţia de propagandă tehnică agricolă, Bucureşti, 50 p.
165. Larin I.V. şi col., 1951, Kormovîie rastenia senocosov I pastbisci SSSR, Vol.I, II, Moskva-Leningrad
166. Lauer C., 1974, Studiul geobotanic şi topologic al pajiştilor naturale din carstul bănăţean, zona Craşova (judeţul Caraş- Severin) ,Teză de doctorat I.A.N.B., Bucureşti
167. Lombardi G., 2001, Gestione delle risorse proto-pascolive alpine, Contributi alla conoscenza scientifica, MIPAF, Italy
168. Lupaşcu Gh., Rusu C., Andreiaşi N., Seceleanu I., 1985, Consideraţii pedogeografice asupra teritoriului jud. Suceava, Lucrările Seminarului Geografic Dimitrie Cantemir, nr. 6 Iaşi
169. Lupaşcu Gh., 1996, Depresiunea Cracău-Bistriţa. Studiu Geomorfologic, Ed. Corson, Iaşi
170. Magid J., 1993, Vegetation effects on phosphorus fractions in set-aside soils, Plant and Soil, 149, 111-119
171. Maloş C., 1977, Flora şi vegetaţia din Bazinul Superior al Motrului, Rezumat al tezei de doctorat Universitatea Bucureşti
172. Mamolos.P., Vasilikos, C.V., Veresoglou D.S., 2005, Vegetation in contrasting soil water sites of upland herbaceous grasslands and N : P ratios as indicators of nutrient limitation, Plant and Soil, 270, 355-369
173. Marcu M., 1971, Cercetări topoclimatice şi fenologice în Masivul Postăvarul, Teză de doctorat, Braşov;

345
174. Maret M. P., Wilson M.V., 2005, Fire et litter effects on seedling establishment in western Oregon upland prairies, Restoration Ecology, 13, 562-568
175. Marcu M., 1983, Meteorologie şi Climatologie Forestieră, Ed. “Ceres” Bucureşti
176. Maruşca, T., 1974, Influenţa măsurilor de îmbunătăţire a pajiştilor de Nardus stricta L. asupra randamentului în producţie animală, Revista de zootehnie şi medicină veterinară nr.3, pag. 49-57, Bucureşti
177. Marusca T. 1976, Observatii privind comportamentul tineretului taurin de pasune. Revista de cresterea animalelor nr.3, pag. 66-73, Bucuresti
178. Marusca T. 1977, Sisteme de infiintare a pajistilor temporare pe suprafetele dominate de Nardus stricta L. Lucrari stiintifice ale SCCP Magurele-Brasov, vol. III, pag. 35-49, Re-dactia materiale de propaganda agricola, Bucuresti.
179. Marusca T. 1977, Influenta modului de folosire, fertilizare si amendare asupra pajistilor dominate de specia Nardus stricta. L. din zona de deal. Lucrari stiintifice ale SCCP Magurele-Brasov, vol. III, pag. 5l-6l, Redactia materiale de propaganda agricola, Bucuresti.
180. Marusca T. 1980, Randamentul in produse animale (spor greutate vie) a conveierelor de pajisti.Lucrari stiintifice ale SCCP Magurele-Brasov, vol.VI, pag. 207-218, Redactia materiale de propaganda agricola, Bucuresti.
181. Maruşca T., 1982, Studiul geobotanic şi topologic al nardetelor din judeţul Braşov, Teză de doctorat, Institutul agronomic “N. Bălcescu”, Bucureşti
182. Maruşca T., 1982, Suprainsamantarea pajistilor, mijloc eficient de sporire a productiei si calitatii furajelor. Revista de cresterea animalelor nr. 2, pag. 18-22, Bucuresti.
183. Maruşca T., 1982, Comportamentul pe pasune a tineretului ovin si bovin. Revista de cresterea animalelor nr. 5 pag. 7-14, Bucuresti.
184. Maruşca T., 1983, Tipuri de pajisti cu Nardus stricta L. si posibilitati de imbunatatire a lor.Revista de cresterea animalelor nr. 3 pag. 14-21, Bucuresti.
185. Maruşca T., 1992, Contributii privind cauzele degradarii unor agro-ecosisteme de pajisti naturale prin expansiunea speciei Nardus stricta L. in covorul vegetal A V-a Conferinta Nationala de Ecologie, pag. 188, Cluj Napoca
186. Maruşca T., 1993, Bioproductivitatea pădurilor şi pajiştilor naturale din Carpaţii Meridionali în sprijinul ipotezei privind amplasarea capitalei statului dac – Sarmisegetuza Regia la 1200 m alt., Lucrările Sesiunii anuale de comunicări ştiinţifice, ICAS-Staţiunea Braşov, pag. 351-354, Braşov

346
187. Maruşca T., 1993, Studii privind exigenţele nardetelor faţă de înclinaţia şi expoziţia terenului pe altitudine în Carpaţii Meridionali, Sesiunea ştiinţifică Universitea “Babeş Bolyai”, Secţia de biologie, pag. 41-42, Cluj-Napoca
188. Maruşca T., Doniţă N., 1994, Studiul privind distribuţia şi structura pe altitudine a pădurilor şi pajiştilor naturale din Carpaţii României, Sesiunea jubiliară “45 de ani de cercetare şi proiectare silvică” în Staţiunea Braşov (ICAS), 22-23 iunie
189. Maruşca T., Doniţă N.,1994, Aspecte de gradientică ecologică montană din Carpaţii Românie, Meteorologie, Studii şi cercetări, nr. 9 (I), pag. 75-83, INMH Buc.
190. Maruşca T., Spirescu N., 1994, Studii asupra evoluţiei pe altitudine a înălţimii unor graminee perene de pajişti din Platoul Bucegilor (Carpaţii Meridionali), Conferinţa ştiinţifică a botaniştilor Ocrotirea, reproducerea şi utilizarea plantelor, pag. 31-32, Chişinău, Moldova
191. Maruşca T., 1995, Stabilirea unor gradienţi orografici de răspândire a pajiştilor cu Nardus stricta din Carpaţii Meridionali, A VI-a Conferinţă naţională de ecologie, pag. 247-248, Arad
192. Maruşca T., 1995, Gradientica ecologică - Baza strategiilor de dezvoltare durabilă în zona montană, A VI-a Conferinţă naţională de ecologie, Universitatea de Vest, Arad, 237-245
193. Maruşca T., 1995, Cercetări asupra ecologiei pajiştilor dominate de Nardus stricta L. din judeţul Braşov, Lucrări ştiinţifice, Simpozion omagial „George Bujorean”, USAB, Timişoara, vol. XXVIII, 487-492
194. Maruşca T., 1995, Contributii la stabilirea unor gradienti pedoclimatici verticali caracteristici conurilor vulcanice din Muntii Harghitei. Factori si procese pedogenetice din zona temperata, vol. II, pag.119-122, Ed. Universitatii “A.I. Cuza” Iasi
195. Maruşca T., Oprea Georgeta, Chiper Constantina, 1996, Studies concerning to impact of rational grazing with youngh cattle on some sown grasslands, Proceding of the 3nd International Scientific Conference, pag. 114-117, USAMV-Bucureşti
196. Maruşca T., Oprea Georgeta, 1996, Caracteristicile unor tipuri de telina din zona montana inalta si rolul lor in protectia antierozionala. A 3-a conferinta nationala pentru protectia mediului prin metode si mijloace biologice si biotehnice Brasov, pag. 483-486, Universitatea Transilvania Brasov
197. Maruşca T., Gh. Dologa, 1996,Cercetari asupra mijloacelor biotehnice de inierbare a suprafetelor fara vegetatie din etajul subalpin al platoului Bucegilor. A 3-a Conferinta nationala pentru protectia mediului, Brasov, pag.487-491, Universitatea Transilvania Brasov

347
198. Maruşca T., Mocanu V., Hermenean I. 1997, Erbicidele paraquat si glifosat in sprijinul reconstructiei ecologice a covorului ierbos de gradat Nardus stricta L. di n pajistile montane Proplant 97, pag. 118, Calimanest
199. Maruşca T., 1997, Studii asupra gradienţilor ecologici şi economici din spaţiul carpatic românesc, Lucrările primului Congres al Academiei Oamenilor de Ştiinţă din România “Ştiinţa la sfârşit de mileniu”, pag. 176-178, Bucureşti
200. Marusca T., 1998, Utilisation des gradients altitudinaux pour le completement et le transfer des connaissances sur le fond pastoral mantagnard de Roumanie FAO- CIHEAM, 10 eme reunion du grupe de travail des herbages de montagne, 10-13 septembrie, Braşov – România
201. Maruşca T., Neagu M., Mocanu V.1999, Strategies of sustainable agropastoral development in Plateau Bucegi (Meridional Carpathians), Journal of Mountain Agriculture on the Balkans, vol. 2, nr.1 pag.67-73, IUSBA-Troyan, Bulgaria
202. Maruşca T., 1999, Genetic resources of grasses and legumes in Romania IPGRI, Seventh Meeting of the ECP/GR Working Group on Forages, Pp. 132-136, Elvas, Portugalia
203. Maruşca, T., 2000, The productivity of different types of sown grasslands expossed in weight gain, 18th General Meeting of the European Grassland Federation, Aalborg – Denmark, Balancing enviromental and economic demands, EGF, Volume 5, Pp. 479-581, British Grassland Society, U.K.
204. Marusca T., Mariana Rusu G. Ciortea, 2000, Determinarea unor gradienti altitudinali de productivitate a pajistilor naturale din Muntii Cindrelului-Sibiu Buletin stiintific, Seria Agrisib, nr. 1, pag.175-180, Universitatea “Lucian Blaga” Sibiu
205. Maruşca, T., 2000,The productivity of different types of sown grassland expreseed in weight gain Grassland Farming, Balancing enviromental and economic demands, Grassland Sciences in Europe, vol. 5, Pp 579-581, British Grassland Society, United Kingdom
206. Maruşca T., Pop M., Neagu M., Mocanu V., Blaj A.V., Oprea Georgeta, 2001, Efectul tehnologiilor de îmbunătăţire a păşunilor subalpine asupra producţiei vacilor de lapte, Simpozionul “Cercetarea ştiinţifică şi Agricultura durabilă”, Academia Oamenilor de Ştiinţă din România, Bucureşti, 93-102
207. Marusca T. Blaj V.A. Stejaru Nicoleta, 2001, Gradientică montană, o posibilitate nouă de valorificare a rezultatelor cercetării ştiinţifice din spaţiul agrosilvopastoral Lucrările celui de al XIX-lea Simpozion Naţional al Societăţii de Istorie şi Retrologie Agrară din România, pag.293-294, Braşov

348
208. Maruşca T., 2001, Elemente de gradientică şi ecologie montană, Editura Universităţii „Transilvania”, Braşov, 130 p.
209. Maruşca T., 2002, Utilizarea gradienţilor pentru aprecierea evoluţiei pe altitudine a resurselor naturale şi socioeconomice din spaţiul montan Lucrările Clocviului Naţional de Geografie a Populaţiei şi Aşezărilor Umane, Ediţia a XIII-a, “Habitatul rural şi urban în spaţiul montan”, pag. 38, Universitatea “Valahia”, Târgovişte – Căprioara, 25-27 octombrie.
210. Maruşca T., Mocanu V., Blaj A.V., 2002, Effect of improvement tehnologies on subalpine pastures in milk production of dairy cows Proceedings of the 19th General Meeting of the European Grassland Federation, Multi – Function Grasslands, Grassland Science in Europe, vol. 7, Pp 1052-1053, La Rochelle, France.
211. Maruşca T., Lăcătuşu R., Kovacsovics B., Chiper C., Blaj V.A., Stejaru N., 2002, Cercetări privin durata optimă de târlire cu ovinele pe păşunile subalpine de Nardus stricta L. şi miloace de atenuare a efectelor supratârlirii Lucrări ştinţifice ale Instiutului de Cercetare Dezvoltare pentru Pajişti Braşov, pag. 172 – 179, ed. Universitâţii “Transilvania” din Braşov
212. Maruşca T., Frame J., 2003., Pasture improvement strategies in the Carpathians pacage with dairy cows EGF – 2003, Bulgaria, Optimal Forage Systems for Animal Production and the Environement, Grassland Science in Europe , Vol.8, Pp.219-221
213. Maruşca T., .Pop O., Vodă Al.,Vergheleţ M., 2003, Evaluation of grazing presure on the Piatra Craiului National Parks pastures an management issues Research in Piatra Craiului National Park, Vol.I, Pp. 130-141 Ed. Ph
214. Marusca T.,2003, The productivity of different types of sown grasslands expressed in weight gain, FAO, “HERBA” nr.13, Pp. 53-55, Nyon, Suisse
215. Maruşca T., Mocanu V., Neagu M., Blaj V.A., 2005, Posibilităţi de ameliorare a efectelor negative asupra supratârlirii cu oile pe nardetele subalpine A 7-a Conferinţă Naţională pentru Protecţia Mediului, Universitatea Transilvania, Braşov
216. Maruşca T., Mocanu V., 2006, Utilization of altitudinal gradients for interdisciplinary knowlwdge on mountain grasslands productivity Proceedings of the 21st General Meeting EGEF, Badajos, Spain, Pp.802-804
217. Maruşca T.,2006, Contribuţii la reconstrucţia ecologică a pajiştilor degradate de supratârlirea cu animale. MEC, AMCSIT, Cercetarea de excelenţă – premiză favorabilă pentru dezvoltarea spaţiului românesc de cercetare, vol.I, Braşov, Editura PRINTECH

349
218. Maruşca T., Mocanu V., Blaj V.A., 2007, Improvement systems of mountain grasslands depending of the altitudinal productivity in Romanian Carpathians, Journal of mountain agriculture on the Balkans, 14 th Meeting of the FAO - CIHEAM Mountain Pastures Network, RIMSA, Troyan, Bulgaria Pp. 187 – 193
219. Maruşca T., Mocanu V., Blaj V.A.Hermenean I.,2007, Systems for improvement of Nardus stricta subalpine grasslands from Carpathian Mountains Permanent and Temporary Grassland Plant, Environment and Economy , Volume 12, Grassland Science in Europe, European Grassland Federation EGF 3-5 september 2007 Ghent, Belgium, Pp. 94-97
220. Maruşca T., Mocanu V., Blaj V.A., 2007, Sisteme de reconstrucţie ecologică a pajiştilor dominate de Nardus stricta L din Munţii Bucegi AMCSIT Politehnica, CEEX, Conference, Braşov, CD –Rom, ISSN 1843-5904, Editura Tehnică, Bucureşti
221. Maruşca T., 2007, Creşterea performanţei târlirii cu animalele pe pajiştile montane invadate de Nardus stricta (nardete) Oferta cercetării ştiinţifice pentru transfer Tehnologic în agricultură, industria alimentară şi silvicultură,vol.IX,2005-2006,Bucureşti pp189-190
222. Maruşca T., Mocanu V., Blaj V.A., 2008,Research regarding to obtaining of milk cow for agri-tourism ecological methods in Bucegi Plateau, Journal of Eco-AgriTourism, Proceeding of BIOATLAS, Conference vol 1, Transilvania University of Brasov, pp.80-84
223. Maruşca T., Mocanu V., Blaj V.A.Hermenean I.,2008, Organic farming system utilization of improved subalpine pastures in the Carpathian Mountains, Biodiversity and Animal Feed, 22nd General Meeting of the European Grassland Federation, Uppsala, Sweden, pp.111
224. Maruşca T., Mocanu V., Blaj V.A. Hermenean I., Scurtu I., 2008, Biological milk production on improved sub alpine pastures in Carpathian Mountains, International Pasture Conference, Luxembourg, pp. 25-26
225. Marusca T., 2008, Reconstructia ecologica a pajistilor degradate, Editura Universităţii Transilvania din Brasov, 108 pagini
226. Marusca T., 2010, Îmbunătăţirea nardetelor montane şi conversia la producţia ecologică de lapte, Revista Profitul Agricol, nr. 36 şi 37 septembrie, Bucureşti
227. Marusca T.,Mocanu V., Cardaşol V., Hermenean I., Blaj A. V., Oprea Georgeta, Tod A. Monica, 2010, Ghid de producere ecologică a furajelor de pajişti montane, Editura Universităţii Transilvania, Braşov
228. McAlpine K.G., Drake D.R., 2003, The effects of small-scale environmental heterogeneity on seed germination in experimental treefall gaps in New Zealand, Plant Ecology, 165, 207-215

350
229. Meentemeyer R.K., Moody A., 2000, Rapid sampling of plant species composition for assessing vegetation patterns in rugged terrain, Landscape Ecology, 15, 697-711
230. Miller J.,Franklin J., 2002, Modeling the distribution of four vegetation alliances using generalized linear models and classification trees with spatial dependence, Ecological Modelling, 157, 227-247
231. Moisuc, Al., Samfira, I., Carrere, P., 2001 – Pajişti naturale şi exploataţii ecologice, Ed. Agroprint, Timişoara
232. Moisuc Al,, Dukic D., 2002, Cultura plantelor furajere, Ed. Orizonturi Universitare,Timişoara
233. Moldovan I., 1970, Flora şi vegetaţia Muntelui Gutîi, Teză de doctorat, Cluj
234. Monk C. D., Gabrielson F. C., 1985, Effects of shade, litter et root competition on old-field vegetation in South-Carolina, Bulletin of the Torrey Botanical Club, 112, 383-392
235. Montard F.m Gachon L., 1976, Les paturages d’altitude des Mont Dore, Station de Agronomie, Clermont Ferrand
236. Morariu I., 1970, Un aspect de vegetaţie naturală din Dobrogea, Acta Bot. Horti., Bucureşti, 193-199
237. Morariu I., Danciu M., 1977, Studii şi cercetări de biologie, Seria Biologie vegetală, Tom 29, nr. 2, Ed. Academiei RSR
238. Moravec J., 1965, Wiesen im mittleren Teil des Bohmer-waldes-Sumava, Praha
239. Morea A., 2008, Cercetări asupra evoluţiei biodiversităţii şi productivităţii în pajiştile de Festuca rubra din etajul boreal (cu privire specială asupra Munţilor Apuseni ), Teză de doctorat, U.S.A.M.V. Cluj-Napoca
240. Motcă Gh., 1972: Cercetări privind metodele de îmbunătăţire a pajiştilor din Depresiunea Făgăraşului, Teză de doctorat, IANB-Bucureşti
241. Motcă Gh., Roşu, C., Belu C., 1985, Pajiştile din Munţii Făgăraşului, Lucrări ştiinţifice a ICPCP Braşov, vol. X, pag. 247-264, Red. prop. tehn. agr. Bucureşti
242. Motcă Gh., I. Oancea, Lidia Geamănu, 1994: Pajiştile României, Tipologie şi Tehnologie, Editura tehnică agricolă, Bucureşti
243. Munteanu I., Florea N., Seceleanu I., 1988, The principles of soil classification în Romania, International Scientific Conference on Soil Classification, Alma-Ata, URSS

351
244. Munteanu I., Sorina Dumitru, Victoria Mocanu, Irina Moise, 1998, Tipurile de terenuri din România la sc. 1:2.500.000 după metodologia SOTER şi utilizarea lor pentru fundamentarea strategiei de conservare şi protecţie a fondului funciar, Lucrările Simpozionului "Protecţia mediului în agricultură", 29 septembrie, 103-122
245. Nakamura T., 1996, Ecological relationships between seedling emergence et litter cover in theearliest stage of plant succession on sandy soil, Ecological Research, 11, 105-110
246. Näsholm T., Huss-Danell K., Högberg P., 2000, Uptake of organic nitrogen in the field by four agriculturally important plant species, Ecology, 81, 1155-1161
247. Neacşu Marcela, Vasu Alexandra, Zanelli Sofia Cardaşol V., Kovacs A., 1978, Caracterizarea condiţiilor staţionale ale pajiştilor din perimetrul Păltiniş-Cibin şi aprecierea capacităţii de producţie, Lucrările şt. ale SCCP Măgurele Braşov, vol. IV, Bucureşti
248. Neacşu Marcela, Mielcescu Anca, 1979, Aspecte privind influenţa condiţiilor staţionale asupra vegetaţiei pajiştilor permanente, Ştiinţa solului, vol. 5-6, Bucureşti
249. Neacşu Marcela,. Kovács A., 1979, Aspecte privind cercetarea vegetaţiei şi staţiunilor de pajişti din perimetrul Sebeş- Braşov, Lucrările ştiinţifice ale Staţiunii Centrale de Cercetări pentru Cultura Pajiştilor Măgurele Braşov, vol. V, pag. 127-136, Bucureşti
250. Neacşu Marcela, Dragu I., Ţucra I., Cardaşol V., Vasu Alexandra., Dulvara Eufrosina, Tudor Ana., 1986, Caracterizarea câmpurilor experimentale în pajişti în cadrul grupelor de tipuri de pajişti din R.S.România, Lucrările Ştiinţifice ale Institutului de Cercetare şi Producţie pentru Cultura Pajiştilor Măgurele-Braşov, vol. XI, Bucureşti
251. Newell C.L., Peet R.K., 1998, Vegetation of Linville George Wilderness, North Carolina, Castanea 63, 275-322
252. Niedermaier, K., 1958, Experienţe de îngrăşare a pajiştilor de munte, Analele ICA, vol. XXVI, Seria A, pag. 121-139, Editura Academiei Române, Bucureşti
253. Niedermaier, K., 1960, Contribuţii la cunoaşterea efectului îngrăşămintelor pe pajiştile din Munţii Cibinului, Analele ICA, seria A, vol. XXVIII, pag. 163-177, Editura Academie Române, Bucureşti
254. Niedermaier K., 1962, Cercetări asupra dinamicii producţiei unor pajişti situate la diferite nivele altitudinale. Comunicări ale Academiei Române, Tom. XII, nr. 12, pag. 1333-1340, Bucureşti

352
255. Niedermaier K., Sluşanschi H., 1964, Cotribuţii la cunoaşterea valorii nutritive a pajiştilor naturale din Munţii Cibinului, Analele ICCPT-Fundulea, vol. XXII, Seria B, pag. 269-386, Centrul de Documentare Agricolă, Bucureşti
256. Niedermaier K., 1967, Rev. Probleme agricole, nr. 5, Bucureşti
257. Niedermaier K., 1968, Productivitatea pajiştilor dintre Rîul Mare şi Valea Sadului-Munţii Cibinului, Teză de doctorat, I.A.N.B., Bucureşti
258. Niinemets U., Kull K., 2005, Co-limitation of plant primary productivity by nitrogen and phosphorus in a species-rich wooded meadow on calcareous soils, Acta Oecologica-International Journal of Ecology, 28, 345-356
259. Nogués-Bravo D., Araujo M.B., Romdal T., Rahbek C., 2008, Scale effects and human impact on the elevational species richness gradients, Nature, 453, 216-220
260. Nyarady A. şi col., 1974, Lucrări ştiinţifice ale Inst. Agr. “Dr. P. Groza”, Cluj
261. Oberdorfer E., 1957, Suddetsche Pflanzengesellschaften, Gustav Fischer, Verlag, Jena
262. Obrejanu Gr., 1941, Rev. Agricultura nouă, nr.6, Bucureşti
263. Obrejanu Gr., 1957, Studii şi cercetări de agronomie, Tom VIII, nr.1-3, Filiala Cluj, Ed. Acad.RPR
264. Olff H., Huisman J., Vantooren B.F., 1993, Species dynamics and nutrient accumulation during early primary succession in coastal sand dunes, Journal of Ecology, 81, 693-706
265. Otsus M., Zobel M., 2004, Moisture conditions et the presence of bryophytes determine fescue species abundance in a dry calcareous grassland, Oecologia, 138, 293-299
266. Panait V., 1969, Contribuţii la studiul vegetaţiei şi îmbunătăţirea pajiştilor naturale din Bazinul Moldoviţei, judeţul Suceava, Rezumat al tezei de doctorat. Institutul agronomic “Ion Neculce”,Iaşi
267. Paraschiv D., 1965, Piemontul Cândeşti, Ştiinţă şi tehnică geologică, Bucureşti
268. Parichi M., Seceleanu I., 1989, Contribuţii la cunoaşterea perioadelor caracteristice de geneză şi evoluţie a unor soluri din Câmpia Română şi Piemontul Getic, Studii şi cercetări de geografie, Tomul XXXIV Acad. RSR.
269. 236.Parker A.J., 1982, The topographic relative moisture index: an approach to moisture assessment in mountain terrain, Physical Geography, 3, 160-168
270. Parton W.J., Stewart J.W.B., Cole C.V., 1988, Dynamics of C, N, P and S in grassland soils - a model,, Biogeochemistry, 5, 109-131
271. Pascal P., 1973, Flora şi vegetaţia din bazinul Bistriţa Aurie (jud. Suceava), Teză de doctorat, Bucureşti, 374 p.

353
272. Pascovschi S., Leandru V., 1958, Tipuri de pădure din RPR, Ed. Agro-Silvică, Bucureşti
273. Pastor J., Stillwell M.A., Tilman D., 1987, Nitrogen mineralization and nitrification in four Minnesota old fields, Oecologia, 71, 481-485
274. Pavel C. şi col., 1962, Pajiştile din Masivul Parâng şi îmbunătăţirea lor, Inst. Agr. Craiova, Ed. Agro-Silvică
275. Pavel C., 1973, Pajiştile din zona subcarpatică a Olteniei, Ed. Scrisul Românesc, Craiova
276. Păcurar F., Rotar I., Gârda Nicoleta, 2008, Elemente de dezvoltare durabilă în managementul tradiţional al pajiştilor cu Arnica montana, Hameiul şi plantele medicinale, Anul XVI, nr. 1-2 (31-32), Cluj-Napoca, ISSN 1454-7805 pp. 251-254
277. Păcurar F., Rotar I., Morea Adriana, Garda Nicoleta, 2008. Productivity and biodiversity evolution of Festuca rubra grasslands during a seven years period, Bulletin of University of Agricultural Sciences and Veterinary Medicine, seria Agricultura Print ISSN1843-5246 Electronic ISSN 1843-5386, pp. 186-190
278. Păltineanu Cr., Seceleanu I, Crăciun C., 2003, Vertisolurile şi solurile cu caractere vertice din România, Ed. Estfalia, Bucureşti
279. Păucă A., Puşcaru-Soroceanu Evd., Ciucă M., 1960, Contribuţii la studiul pajiştilor din masivul Ciucaş, Bul. Soc. de ştiinţe nat. şi geografie, 1
280. Păun M., Popescu Gh., Zaharia I., Vegetaţia. Pajiştile din Zona Subcarpatică a Olteniei, Ed. Scrisul Românesc, Craiova, 1973, 98-172
281. Piper F.I., Cavieres L.A., Reyes-Diaz M., Corcuera L.J., 2006, Carbon sink limitation and frost tolerance control performance of the tree Kageneckia angustifolia D. Don (Rosaceae) at the treeline in central Chile, Plant Ecology, 185, 29-39
282. Pîrvu I. şi col., 1980, Ecosistemele din Romînia, Ed. Ceres Bucureşti
283. Perkins F.D., 1965, Ecology of Nardus stricta L. , Journal of Ecology, 54
284. Pop E. ,1960, Mlaştinile de turbă din R.P.R, Ed. Academiei R.P.R. Bucureşti
285. Pop I., Hodişan I., 1969, Consideraţii asupra florei şi vegetaţiei masivelor calcaroase de pe Valea Sighişel (Munţii Bihor), Studia Univ. Babeş-Bolyai, Seria Biol., Cluj, 1, 33-43
286. Pop I., 1977, Biogeografie ecologică, Editura Dacia, Cluj Napoca
287. Pop I., Cristea V., Hodisan I., 2000, Vegetatia judetului Cluj (Studiu fitocenologic, ecologic, bioeconomic si eco-protectiv), Contribuţii botanice, XXXV, 5-254
288. Popovici D., Chifu T., Ciubotariu C., Mititelu D., Lupaşcu G., Davidescu G., Pascal P., 1996, Pajiştile din Bucovina, Ed. Helios, 340 p.

354
289. Puia, I.., Popescu V., Erdely Şt., Vorica Târău, Heinke Klemm, 1976, Productivitatea unor ecosisteme de pajişti montane şi alpine din România, Lucrări şt. ale SCCP Măgurele-Braşov, vol. II. Red. rev. de prop. agr., Bucureşti
290. Puia I., Soran V., 1979, Alimentaţia şi agricultura în următoarele trei decenii, Editura Academiei RSR, Bucureşti
291. Puia I. şi colab., 1984, Producerea şi păstrarea furajelor, Ed. Didactică şi pedagogică Bucureşti.
292. Purchase B.S., 1974, Influence of phosphate deficiency on nitrification, Plant and Soil, 41, 541-547
293. Puşcaru D., Puşcaru- Soroceanu Evdochia, Păucă A.,Şerbănescu I.,Beldie Al., Ştefureac Fr., Cernescu N., Saghin F., Creţu V., Lupan L., Taşcenco V., 1956, Păşunile alpine din Munţii Bucegi, Edit. Acad. R.P.R., Bucureşti
294. Puşcaru- Soroceanu Evd. ş.a., 1963, Păşunile şi fâneţele din R.P.R., Editura Academiei R.P.R., Bucureşti
295. Puşcaru- Soroceanu Evd., Doniţă N.,1963, Inventarierea şi cartarea vegetaţiei din R.P.R., S.S.N.G. Comunicări de botanică, vol.II, P.I
296. Puşcaru – Soroceanu Evdochia ş.a., 1965, Fertilizarea pajiştilor de Nardus stricta din Masivul Făgăraş, Analele I.C.C.P.T. Fundulea, seria B, vol. XXXIII
297. Puşcaru - Soroceanu Evd., Puşcaru D., 1969, Asociaţiile pajiştilor alpine din Munţii Făgăraş sub aspect fitogeografic şi al valorii lor productive, Comunicări de botanică, vol.XI
298. Quested H.,Eriksson O., 2006, Litter species composition influences the performance of seedlings of grassland herbs, Functional Ecology, 20, 522-532
299. Raţiu O., 1970, Consideraţii ecologice asupra principalelor grupări de plante din Bazinul hidrografic al Văii Colobiţa (Munţii Călimani), Contribuţii botanice, Cluj
300. Raţiu O., Moldovan I., 1974, Consideraţii cenologice asupra vegetaţiei muntelui Ignis, Contrib. Bot., Cluj-Napoca, 85-94
301. Raţiu O., 1980, Fitocenoze ierboase specifice bazinului Someşului Cald şi acţiunea lor antierozională, Contrib.Bot., Cluj-Napoca, 143-159
302. Raubuch M., Beese F., 2005, Influence of soil acidity on depth gradients of microbial biomass in beech fores soils, European Journal of Forest Research, 124, 87-93
303. Resmeriţă I.,1963, Studii de vegetaţie pe pajiştile din Maramureş ,St. şi Cerc.Biol., Cluj, 14, 2, 223-255
304. Resmeriţă I., 1969, Pajiştile Masivului Vlădeasa. Flora, vegetaţia şi potenţialul productiv, Teză de doctorat, Inst. Agr., Timişoara

355
305. Resmeriţă I., 1970, Flora, vegetaţia şi potenţialul productiv pe Masivul Vlădeasa. Editura Academiei R.S.R.
306. Resmeriţă I., 1971, Cercetări experimentale pe nardete de joasă altitudine, Inst. Agr. Cluj
307. Roşu Al., 1973, Geografia fizică a României, Ed. Didactică şi Pedagogică, Bucureşti
308. Rotar I., Prica Fl., Sima N., 1997, Valorificarea cu ovine a pajiştilor de Festuca rubra–Agrostis tenui, Simpozionul Naţional “Realizări şi perspective în zooterhnie”, vol. XXVIII, pag. 408-417, Tipo Agronomia, Cluj-Napoca
309. Rotar I., Păcurar F., Stoie A., Gârda Nicoleta, Dale Laura, 2010, The evolution of Arnica montana L. grasslands depending on the performed management (Apuseni Mountains, România), USAMV - Iasi, in Revista Lucrari stiintifice vol. 53/2010, Seria Agronomie, ISSN 1454-7414, (CD)
310. Rothmaler W., 1970, Exkursionflora von Deutcshland, Wolk und Wisen Volkseigener, Verlag, Berlin
311. Rotundo J. L., Aguiar M.R., 2005, Litter effects on plant regeneration in arid lands: a complex balance between seed retention, seed longevity et soil-seed contac, Journal of Ecology, 93, 829-838
312. Sabatini S., Albertosi A., Argenti G., Bianchetto E., Stagliano N., 2003, Improvement of pastures encroached by shrubs through animal grazing in an Italian Alpine environment, Grassland Science in Europe, vol.8, pg. 74-76
313. Safta I., 1936, Contribuţiuni la studiul fitosocial-agricol al fâneţelor din jud. Cluj, Analele Inst. de Cercet. Agron. ale Romîniei 8, Bucureşti
314. Safta I., 1938, Păşunile din judeţul Timiş, Analele I.C.A.R.
315. Safta I., 1943, Cercetări geobotanice asupra păşunilor din Transilvania, Buletinul Facultăţii de Agronomie Cluj -Timişoara, vol. X
316. Safta I. Şi col., 1979, Pajiştile din Banat, Red. de prop. tehn. Agric. , Bucureşti
317. Samoilă Z., 1971, Cercetări experimentale privind reacţia la fertilizareşi amendare cu calciu a unor tipuri de pajişti din Banat, Rezumat al tezei de doctorat, Institutul Agronomic Timişoara
318. Samoilă Z., şi colab., 1979, Pajiştile din Banat, M.A.I.A.-Redacţia de propagandă tehnică agricolă
319. Sanda V., Popescu A., Peicea I., 1972, Contribuţii la cunoaşterea vegetaţiei din judeţul Hunedoara, St. şi Cerc. Biol., Seria Bot., Bucureşti, 24, 4, 295-317
320. Sanda V., Popescu A., Doltu M.I., 1980, Cenotaxonomia şi corologia grupărilor vegetale din România, Studii şi comunicări, Ştiinţe naturale, 24, 11-171

356
321. Sanda V., Popescu A., Barabaş N., 1998, Cenotaxonomia şi caracterizarea gruparilor vegetale din România, Studii şi Comunicări - Biologie Vegetală, 14, 9-366
322. Sanda V., Popescu A., Stancu D., 2001, Structura cenotică şi caracterizarea ecologică a fitocenozelor din România, Ed. Conphis, Piteşti, 359 p.
323. Sanda V.,Ollerer K., Burescu P., 2008, Fitocenozele din România, Sintaxonomie, structură, dinamică şi evoluţie, Editura Ars Docendi, Universitatea Bucureşti
324. Sanda V., Vicol I., Ştefănuţ S., 2008, Biodiversitatea cenostructurală a învelişului vegetal din România, Editura Ars Docendi, Universitatea Bucureşti
325. Sârbu Anca, Coldea G., Negrean G., Cristea V., Hanganu J., Veen P., 2004, Grasslands of Romania, Ed.Alo, Bucureşti, 67 p.
326. Schellberg J., Moseler B.M., Kuhbauch W., Rademacher I.F., 1999, Long-term effect of fertilizer on soil nutrient concentration, yield, forage quality and floristic composition of a hay meadow in the Eifel mountains, Germany, Grass and Forage Science, 54,195-207
327. Schinner F., 1982, Soil microbial activities and litter decomposition related to altitude, Plant and Soil, 65, 87-94
328. Schulze E.D., Beck E., Müller-Hohenstein K., 2005, Plant ecology, Springer, Berlin
329. 294.Sedia E. G., Ehrenfeld J. G., 2003, Lichens et mosses promote alternate stable plant communities in the New Jersey Pinelands, Oikos, 100, 447-458
330. Seceleanu I., Andreiaşi N., 1975, Argumente pedogeografice în sprijinul originii şi limitelor pajiştilor din golul subalpin al Masivului Gârbova, Lucrările staţiunii de cercetări biologice, geologice şi geografice "Stejarul", seria Geologie-Geografie, jud. Neamţ
331. Seceleanu I., 1982, The Romanian Land rating system în comparison with the Dutch system of soil survey interpretation , Soil Survey Institute, Wageningen, The Netherlands
332. Seceleanu I., Vasu Alexandra., Filip Fl. N., Crăciun C., Matei Gabi Mirela, Matei S., Popescu I., Taină Ioana., Chivulete S., 1998, The study of two Mediteranean ecosistems for the assessement of the desertification risk, Soil Science nr. 1-2, Bucureşti
333. Seceleanu I., Vasu Alexandra., Filip Fl. N., Crăciun C., Matei Gabi .Mirela, Matei S., Popescu I., Taină Ioana., Chivulete S., Seceleanu I. Al., 1999, Cercetări ecopedologice în vederea evaluării riscului deşertificării şi planificării folosirii terenurilor într-o regiune de coastă mediteraniană, vol. Simp. "Protecţia mediului înconjurător în agricultură" ASAS-ICPA, Bucureşti

357
334. Seceleanu I., Mocanu Victoria, Ispas Şt., 2003, Cercetări ecopedoclimatice asupra luvisolurilor albice din Piemontul Cândeşti, Analele Univ. Valahia-Târgovişte, Seria Geografie Tom 3
335. Seceleanu I., 2004, Geografia fizică a României, Ed. Venus, Bucureşti
336. Sival F.P.,Grootjans A.P., 1996, Dynamics of seasonal bicarbonate supply in a dune slack: effects on organic matter, nitrogen pool and vegetation succession, Vegetatio, 126, 39-50
337. Sîrcu I., 1971, Geografia fizică a Republicii Socialiste România, Ed. Didactică şi Pedagogică, Bucureşti
338. Smith R.L., 1996, Ecology and field biology, HarperCollins College Publishers, New York
339. Someşan L., 1948, Consideraţiuni geomorfologice asupra Munţilor Călimani, Lucr. Inst. Geogr, Univ. Cluj, vol. VII
340. Spirescu M., MehedinţI V., Latiş L., 1985, Pajiştile masivului Gârbova (Munţii Baiului), Lucrările Ştiinţifice ale Institutului de Cercetare şi Producţie pentru Cultura Pajiştilor Măgurele-Braşov, vol. X, Buc.
341. Stănescu V., 1973, Tipologia pădurilor , Silvicultura, Ed. Ceres, Bucureşti
342. Stewart J.W.B.,Cole C.V., 1989, Influences of elemental interactions and pedogenic processes in organic-matter dynamics, Plant and Soil, 115, 199-209
343. Stîngă N. ş.a., 1963, Solurile de pajişti din Munţii Cibinului, Analele secţiei de pedologie, vol. 31
344. Stoenescu Şt. M., 1951, Clima Bucegilor, Mem. şi st. Institut. meteorologic central, Bucureşti, vol.IV
345. Stoenescu Şt.M., Ţîştea D., 1962, Clima României, vol. I, Institutul Meteorologic Bucureşti
346. Stugren B., 1982, Bazele ecologiei generale, Editura Stiintifica si Enciclopedica, Bucuresti
347. Susan F., Ziliotto U:, 2004, Effects of arboreal and shrubby components on some features of pastures in a dolomitic area, Grassland Science in Europe, vol.9, pg. 314-316
348. Şerbănescu I., 1964, Anuarul Comitetului Geologic, vol. XXXIV, partea II-a, Bucureşti
349. Ştefan N., 1980, Cercetarea florei şi vegetaţiei din Bazinul Superior şi Mijlociu al râului Râmnicu- Sărat, Rezumat al tezei de doctorat, Universitatea Al.I. Cuza, Iaşi
350. Ştefănescu Al., 1970, Contribuţi la studiul vegetaţiei şi îmbunătăţirea pajiştilor naturale de deal dintre Olt şi Argeş, Teză de doctorat, IANB, Bucureşti

358
351. Şuteu St., 1970, Flora şi vegetaţia bazinului inferior al Văii Rîmeţului, Rez. Tezei de doctorat, Cluj
352. Tîrziu D., 1994, Ecologie, Universitatea “Transilvania” Braşov
353. Teaci D., 1970, Bonitarea terenurilor agricole, Ed. Ceres, Bucureşti
354. Teaci, D. şi colab., 1980, Resursele de terenuri de pajişti din România şi problemele principale ale productivităţii actuale şi perspectivă, Lucr. şt. SCCP Măgurele-Braşov, vol. VI. Red. prop. tehn. Agr. Bucureşti
355. Teaci D., 1980, Bonitarea ternurilor agricole, Ed. Ceres, Bucureşti
356. Tufescu V., 1966, Subcarpaţii, Ed. Ştiinţifică, Bucureşti
357. Turcu Gh., 1970, Flora şi vegetaţia regiunii deluroase dintre Argeş şi râul Doamnei, Rezumat al tezei de doctorat, Universitatea Bucureşti
358. Tyler G., Olsson T., 2001, Plant uptake of major and minor mineral elements as influenced by soil acidity and liming, Plant and Soil, 230, 307-321
359. Ţîştea D. şi colab., 1974, Studii de climatologie, vol. I, pag. 105-206, IMH Bucureşti;
360. Ţucra I., Kovacs A., Cardaşol V., Neacşu Marcela., 1980, Caracterizarea şi bonitarea pajiştilor, S.C.C.P. Măgurele –Braşov, 10 ani de activitate, Casa agronomului Braşov
361. Ţucra I., Neacşu Marcela, 1984, Conspectul tipurilor de pajişti din Dobrogea, Lucrări ştiinţifice I.C.P.C.P. Măgurele – Braşov, vol.IX
362. Ţucra I., Neacşu Marcela, 1985, Aspecte privind caracterizarea cartarea şi bonitarea pajiştilor permanente (naturale), Lucrările Ştiinţifice ale Institutului de Cercetare şi Producţie pentru Cultura Pajiştilor Măgurele-Braşov, vol. X, Bucureşti
363. Ţucra I., Kovacs A., Roşu C., Ciubotaru C., Chifu T., Neacşu M., Bărbulescu C., Cardaşol V., Popovici D., Simtea N., Motcă GH., Dragu I., Spirescu M., 1987, Principalele tipuri de pajişti din R.S.România, Redacţia de propagandă tehnică agricolă
364. Ularu P., 1972, Cercetări asupra cormofitelor din Munţii Perşani, Teză de doctorat, Universitatea Bucureşti
365. Vasu Alexandra, Neacşu Marcela, Daniliuc D., 1986, Caracterizarea ecopedologică a unităţilor reprezentative de pajişti din regiunea de munte în vederea stabilirii necesarului de măsuri de tehnologii de imbunătăţire, Bucureşti
366. Vîntu V., Moisuc Al., Motca Gh., Rotar I., 2004, Cultura pajştilor şi a plantelor furajere, Ed.Ion Ionescu de la Brad, Iasi, 34

359
367. Velcea Valeria, 2001, Geografia fizică a României, Ed. Univ. "Lucian Blaga", Sibiu
368. Vespremeanu E., 1998, Pedimente, piemonturi şi glacisuri în Depresiunea Mureşului de Jos, Ed. Univ. Bucureşti
369. Walsh R.P.D., Voigt P.J.,1977, Vegetation litter-underestimated variable in hydrology et geomorphology, Journal of Biogeography, 4, 253-274
370. Weaver J.E.,Clements F.E., 1938, Plant ecology, McGraw-Hill Book Company, Inc., New York, London
371. White P.S., Miller R.I., 1988, Topographic Models of Vascular Plant Richness in the Southern Appalachian High Peaks, Journal of Ecology, 76,192-199
372. Williams B.L., Shand C.A., Sellers S., Young M.E., 1999, Impact of synthetic sheep urine on N and P in two pastures in the Scottish uplands, Plant and Soil, 214, 93-103
373. Xiong S. J.,Nilsson C., 1999, The effects of plant litter on vegetation: a meta-analysis, Journal of Ecology, 87, 984-994
374. Xiong S. J., Johansson M. E., Hughes F. M. R., Hayes A., Richards K. S., Nilsson C., 2003, Interactive effects of soil moisture, vegetation canopy, plant litter et seed addition on plant diversity in a wetland community, Journal of Ecology, 91, 976-986
375. Zamfir M., 2000, Effects of bryophytes et lichens on seedling emergence of alvar plants: evidence from greenhouse experiments, Oikos, 88, 603-61
376. Zanoschi V., 1971, Contribuţii la cunoaşterea vegetaţiei masivului Ceahlău, Stud. şi Cerc.de Biol., Seria Bot., Bucureşti, 23(4), 221-226
377. *** Clima R.S.R., vol. II, Date climatologice, Comitetul de stat al Apelor de pe lângă Consiliul de Miniştri, Institutul Meteorologic, 1966, Bucureşti.
378. *** Metodologia elaborării studiilor pedologice, 1987,vol.I,II şi III, I.C.P.A (redactori coord.: Florea N., Bălăceanu V., Răuţă C., Canarache A.) Ed. Redacţia de propagandă şi Tehnică Agricolă, Bucureşti
379. *** Monografia geografică R.P.R., 1960,vol.I, Geografia fizică. Editura Academiei R.P.R., Bucureşti
380. *** Raionarea pedoclimatică, bonitarea şi caracterizarea tehnologică a terenurilor agricole în vederea zonării producţiei agricole,1975, Arhiva I.C.P.A. Bucureşti

ISBN 978-973-598-787-9

















