Page 1
1
ROMÂNIA
MINISTERUL EDUCAŢIEI, CERCETĂRII, TINERETULUI ŞI SPORTULUI
UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINĂ VETERINARĂ
CLUJ-NAPOCA
DEPARTAMENTUL ÎNVĂŢĂMÂNT LA DISTANŢĂ
ŞI FRECVENŢĂ REDUSĂ
Str. Mănăştur Nr.3-5, 400372 Cluj-Napoca, România
tel.+ 40-264-596.384; fax + 40-264-593.792
FACULTATEA DE ZOOTEHNIE ŞI BIOTEHNOLOGII
SPECIALIZAREA: ZOOTEHNIE
ANUL II
CENTRUL UNIVERSITAR ŞI CULTURAL ROMÂN
VITERBO - ITALIA
GENETICĂ ANIMALĂ 1
Prof.univ.dr. AUGUSTIN VLAIC
SEMESTRUL I
ACADEMICPRES
CLUJ-NAPOCA
2011/2012
Page 2
2
CUPRINS
pag.
CAPITOLUL 1 - DEFINIŢIA, OBIECTIVUL ŞI IMPORTANŢA
GENETICII ..............................................................................................
3
CAPITOLUL 2- STRUCTURI CELULARE CU ROL GENETIC .... 4
CAPITOLUL 3 - DIVIZIUNILE CELULARE .................................... 5
3.1. Mitoza .......................................................................................... 5
3.2. Meioza .......................................................................................... 7
CAPITOLUL 4 - TRANSMITEREA CARACTERELOR DE
LA O GENERAŢIE LA ALTA ..............................................................
9
4.1. Interacţiunile genelor alele ........................................................... 9
4.2. Interacţiunea genelor nealele ....................................................... 13
CAPITOLUL 5 - TRANSMITEREA CARACTERELOR
DETERMINATE DE GENE SITUATE PE ACELAŞI
CROMOZOM ..........................................................................................
20
CAPITOLUL 6 - TRANSMITEREA ÎNLĂNŢUITĂ CU SEXUL,
SAU FENOMENUL DE SEX – LINKAGE ..........................................
24
6.1. Transmiterea înlănţuită cu sexul la tipul Abraxas ........................ 25
6.2. Transmiterea înlănţuită cu sexul la tipul Drosophila ................... 26
CAPITOLUL 7 - ACIZII NUCLEICI: STRUCTURĂ ŞI FUNCŢII . 27
CAPITOLUL 8 - FUNCŢIONAREA ADN-ULUI ŞI
EXPRESIA GENELOR ..........................................................................
31
CAPITOLUL 9 - STRUCTURA MOLECULARĂ A
CROMOZOMILOR LA PROCARIOTE ŞI EUCARIOTE ...............
33
9.1. Structura moleculară a cromozomului bacterian ......................... 34
9.2. Structura moleculară a cromozomului la eucariote ..................... 34
9.3. Organizarea secvenţelor de nucleotide în ADN-ul cromozomal
al eucariotelor ...............................................................................
36
Capitolul 10 - MUTAŢIILE ŞI MUTAGENEZA ................................. 37
10.1. Mutaţiile genice ........................................................................... 38
10.2. Mutaţiile genomice (de număr) .................................................... 40
10.3. Mutaţiile cromozomiale sau de structură ..................................... 42
CAPITOLUL 11 - NOŢIUNI DE EREDOPATOLOGIE .................... 44
11.1. Boli ereditare la taurine ................................................................ 44
11.2. Boli ereditare la porcine ............................................................... 45
11.3. Boli ereditare la păsări ................................................................. 46
CAPITOLUL 12 - INGINERIA GENETICĂ ŞI APLICAŢIILE
SALE .........................................................................................................
47
12.1. Tehnologia ADN recombinant ..................................................... 47
12.2. Hibridarea celulară ....................................................................... 50
13.3 Clonarea animalelor ..................................................................... 50
BIBLIOGRAFIE ...................................................................................... 61
Page 3
3
CAPITOLUL 1
DEFINIŢIA, OBIECTIVUL ŞI IMPORTANŢA
GENETICII
G e n e t i c a este ştiinţa biologică ce studiază fenomenele de ereditae şi
variabilitate la toate organismele vii.
Ereditatea (hereditas = moştenire). Este o însuşire fundamentală proprie
tuturor organismelor vii.
Ereditatea, ca însuşire a organismelor vii, asigură continuitatea formelor
de viaţă în succesiunea generaţiilor, sau poate fi definită ca un proces prin care se
realizează asemănarea dintre părinţi şi descendenţi.
Ereditatea se moşteneşte, deci se transmite din generaţie în generaţie,
dar poate suferi în decursul generaţiilor şi unele modificări care conduc la
deosebiri între indivizii componenţi ai diferitelor generaţii, sau chiar între indivizii
aceleiaşi generaţii.
Aceasta înseamnă că organismelor vii le este proprie încă o însuşire
fundamentală – variabilitatea (prima fiind ereditatea).
Variabilitatea fenotipică (VP) manifestată în cadrul generaţiilor
succesive sau în cadrul aceleiaşi generaţii se datorează diferenţelor dintre bazele
ereditare ale indivizilor (VG) precum şi diferenţelor provocate de condiţiile de
mediu (VE).
VP VG + VE
O b i e c t i v u l cursului de genetică - îl constituie cunoaşterea
mecanismului de transmitere ereditară a diferitelor caractere şi însuşiri, adică în ce
mod informaţia genetică transmisă de la o generaţie la alta determină deosebiri şi
asemănări între indivizi.
I m p o r t a n ţ a cursului de genetică. Cunoscând şi înţelegând legile
care guvernează formarea, dezvoltarea şi evoluţia organismelor vii, specialistul în
creşterea animalelor are posibilitatea să acţioneze în direcţia formării unor
organisme care să satisfacă din ce în ce mai mult cerinţele omului.
Complexitatea aspectelor studiate de genetică a determinat concretizarea
unor ramuri ale acesteia, cum sunt: citogenetica, radiogenetica, imunogenetica,
Page 4
4
genetica moleculară, ingineria genetică, genetica populaţiilor şi genetica
cantitativă.
Genetica populaţiilor şi genetica cantitativă sunt ramuri care
fundamentează una dintre cele mai importante ştiinţe aplicative şi anume
ameliorarea plantelor şi animalelor.
Genetica moleculară şi ingineria genetică fundamentează biotehnologiile.
Genetica a revoluţionat întregi domenii ale ştiinţelor, având implicaţii în
biotehnologii, agricultură, horticultură, zootehnie, medicină umană, medicină
veterinară, industria farmaceutică, industria alimentară etc.
CAPITOLUL 2
STRUCTURI CELULARE CU ROL GENETIC
Cele mai importante structuri celulare cu rol genetic sunt cromozomii, care
se găsesc în nucleul celulelor la organismele eucariote. Pe cromozomi sunt aşezate
genele care determină diferitele caractere ale indivizilor.
În metafază, un cromozom este alcătuit din două cromatide surori,
centromer, constricţie secundară, satelit şi telomere (fig.1). Centromerul leagă
cele două cromatide surori la nivelul unei constricţii primare şi cu ajutorul lui,
cromozomii se fixează de fibrele fusului de diviziune. La nivelul constricţiei
secundare se află aşezată regiunea organizatorului nucleolar. Satelitul este prezent
numai la cromozomii care prezintă constricţie secundară. Telomerele sunt situate
la capetele cromozomului, conţin ADN noninformaţional şi au rolul de a menţine
stabilitatea cromozomilor în timpul diviziunilor celulare.
În unele faze ale mitozei (anafază, telofază) cromozomii au structură
monocromatidică, datorită duplicării lor în metafază.
Setul de cromozomi din celulele plantelor şi animalelor prezintă trei
însuşiri importante:
1. Nucleul fiecărei celule somatice conţine un număr fix de cromozomi,
caracteristic fiecărei specii (om = 46, taurine= 60, ovine = 54, porcine = 38,
cabaline= 64, grâu= 42, cartof= 48, mazăre= 14 etc);
2. Celulele somatice conţin cromozomi care sunt dispuşi în perechi (2n);
Page 5
5
3. Celulele sexuale (gameţii) conţin în nucleul lor un singur set de cromozomi
(n), ele fiind celule haploide (cu numărul de cromozomi redus la jumătate).
Fig.1 Structura fizică a cromozomului metafazic
CAPITOLUL 3
DIVIZIUNILE CELULARE
Diviziunile celulare sunt reprezentate de:
- mitoză, care menţine constant numărul cromozomilor din celulele somatice;
- meioză, care înjumătăţeşte numărul cromozomilor din gameţi.
3.1. Mitoza reprezintă tipul de diviziune celulară specific celulelor somatice.
În urma ei, dintr-o celulă cu număr diploid de cromozomi (2n) rezultă două celule
fiice cu acelaşi număr de cromozomi (2n) ca şi celula din care provin.
Mitoza cuprinde două etape: interfaza şi mitoza propriu-zisă (profază,
metafază, anafază şi telofază) care împreună alcătuiesc ciclul celular sau ciclul
de viaţă al unei celule.
Interfaza poate fi subdivizată în 3 perioade: presintetică (G1), sintetică
(S) şi postsintetică (G2).
Page 6
6
Cele mai importante fenomene de interes genetic care au loc în faza
sintetică (S) sunt: replicarea ADN-ului după modelul semiconservativ şi
replicarea cromozomilor. La sfârşitul acestei faze, fiecare cromozom va fi
alcătuit din două cromatide surori.
Fig.2 Fazele mitozei (ciclul celular)
Mitoza propriu-zisă cuprinde fazele: profaza, metafaza, anafaza şi
telofaza.
Cel mai important fenomen genetic are loc în metafază: duplicarea
cromozomilor, fenomen în urma căruia cromozomii devin monocromatidici.
La sfârşitul mitozei (telofază), dintr-o celulă cu 2n cromozomi rezultă
două celule fiice cu acelaşi număr de cromozomi (2n). În celulele fiice,
cromozomii sunt cu structură monocromatidică. Aceste celule fiice cu
cromozomi monocromatidici vor intra şi ele într-un nou ciclu celular şi la
sfârşitul fazei S a interfazei vor rezulta cromozomi cu structură bicromatidică şi
ciclul continuă.
Rezumat: Datorită replicării cromozomilor în interfază şi duplicării
cromozomilor în metafază, mitoza asigură menţinerea constantă a numărului de
cromozomi de la o celulă la alta şi implicit asemănarea celulelor dintr-un
organism.
3.2. Meioza este un tip special de diviziune care are loc în cursul
gametogenezei (spermatogeneză şi ovogeneză) în urma căreia rezultă celulele
sexuale (gameţii) care au un număr de cromozomi redus la jumătate (fig.3).
Page 7
7
Meioza constă în două diviziuni succesive: meioza I şi meioza II.
Meioza I este o diviziune reducţională în urma căreia dintr-o celulă
diploidă (2n) specializată (spermatocitul şi ovocitul de ordinul I), rezultă două
celule fiice haploide (n), cu numărul de cromozomi redus la jumătate
(spermatociţii şi ovociţii de ordinul II). Meioza I conţine următoarele faze:
profaza I, care este o fază mai lungă ce cuprinde mai multe stadii (leptonem,
zigonem, pachinem, diplonem şi diachineză) metafaza I, anafaza I şi telofaza I.
Cele mai importante fenomene genetice care au loc în meioza I sunt:
schimbul reciproc de gene între cromozomii omologi sau crossingoverul (în
diplonem) şi dansul cromozomilor sau repartizarea la întâmplare a cromozomilor
de origine maternă şi paternă în spermatociţii şi ovociţii de ordinul I (metafaza I –
telofaza I).
Celulele haploide rezultate după meioza I conţin cromozomi cu structură
bicromatidică, deoarece în metafaza I nu a avut loc fenomenul de duplicare a
cromozomilor.
Meioza II este o diviziune de multiplicaţie asemănătoare mitozei. Ea
conţine următoarele faze: profaza II, metafaza II, anafaza II şi telofaza II.
În această diviziune, cel mai important fenomen care are loc este
duplicarea cromozomilor, care are loc la sfârşitul metafazei II. În urma duplicării
rezultă cromozomi cu structură monocromatidică. La sfârşitul telofazei II vor
rezulta patru celule haploide (patru spermatozoizi în spermatogeneză şi un ovul şi
trei globuli polari în ovogeneză) în care cromozomii sunt monocromatidici.
Fig.3 Schema spermatogenezei
Page 8
8
Crossingoverul şi dansul cromozomilor, fenomene care au loc în meioză,
reprezintă sursele prin care se realizează variabilitatea genetică în populaţiile de
animale.
Teme de control
1. Răspundeţi cu „DA” sau „NU” în funcţie de corectitudinea sau de falsitatea
afirmaţiei. Justificaţi răspunsurile:
a.) Celulele musculare şi gameţii aceluiaşi animal au număr egal de cromozomi?
Exemplificaţi la taurine.
b.) Cromozomii sexuali se găsesc numai în celulele sexuale (spermatozoid şi
ovulă)?
c.) Celulele sexuale (spermatozoid şi ovulă) conţin şi autozomi?
d.) Din cei 30 cromozomi ai spermatozoidului de taur 20 ar putea fi de
provenienţă paternă, dar 34?
e.) Celulele epidermului, celulele musculare şi cele hepatice ale aceluiaşi animal
au număr diferit de cromozomi, conţin informaţie genetică diferită şi
îndeplinesc funcţii diferite în organism?
2. Câţi autozomi conţin celulele somatice şi spermatozoizii unui taur?
3. Câţi cromozomi de sex (heterozomi) conţin celulele somatice şi spermatozoizii
unui taur?
4. Câţi autozomi şi câţi cromozomi de sex conţin celulele somatice la om?
5. Câţi autozomi şi câţi cromozomi de sex conţin celulele sexuale
(spermatozoidul şi ovula) la om?
6. În care faze ale mitozei cromozomul are structură monocromatidică şi în care
are structură bicromatidică?
7. Câte celule rezultă în urma mitozei şi câţi cromozomi conţin aceste celule la
taurine?
8. Câte celule rezultă în urma meiozei I şi meiozei II şi câţi cromozomi conţin
acestea la om?
9. Ce structură (monocromatidică sau bicromatidică) au cromozomii rezultaţi în
urma mitozei, meiozei I şi meiozei II?
10. Cum se numesc celulele rezultate în urma meiozei (spermatogenezei şi
ovogenezei) la masculi şi femele?
11. Care sunt cele două procese care au loc în cursul ciclului celular care asigură
menţinerea constantă a numărului de cromozomi în celulele somatice?
CAPITOLUL 4
TRANSMITEREA CARACTERELOR DE LA O
GENERAŢIE LA ALTA
Page 9
9
Interacţiunile genelor alele, interacţiunile genelor nealele, linkage-ul şi
crossingoverul reprezintă câteva modele de transmitere a unor caractere de la o
generaţie la alta.
4.1. Interacţiunile genelor alele sunt relaţii care se stabilesc între
genele alele (genele care ocupă aceeaşi poziţie pe o pereche de cromozomi
omologi).
Aceste interacţiuni pot fi de: dominanţă completă, dominanţă incompletă,
codominanţă şi supradominanţă.
Dominanţa completă este o interacţiune în care o genă de la un locus (A)
este complet dominantă asupra alelei sale (a). Din această cauză, indivizii cu
genotip heterozigot (Aa sau AaBb) vor manifesta un fenotip asemănător
genotipului homozigot dominant (AA sau AABB).
Monohibridarea, dihibridarea, trihibridarea etc, sunt cazuri de interacţiuni
ale genelor alele, în care la fiecare locus există o genă complet dominantă asupra
alelei sale.
Monohibridarea presupune încrucişarea între indivizi care se deosebesc
printr-o pereche de caractere (pereche de gene) (fig.4). De exemplu, din
încrucişarea unor tauri din rasa Friză bălţată cu negru, cu vaci din rasa Friză
bălţată cu roşu, s-au obţinut în prima generaţie hibrizi bălţaţi cu negru, iar în
generaţia a doua, 75%indivizi bălţaţi cu negru şi 25% indivizi bălţaţi cu roşu.
Datorită dominanţei complete dintre cele două alele (A şi a), indivizii
heterozigoţi (Aa) din F1 şi F2 manifestă caracterul dominant (culoarea bălţată cu
negru).
Friză Friză
bălţată cu negru bălţată cu roşu
P ♂ AA x aa ♀
Gameţi A A a a
F1 ♂ Aa x Aa ♀ Bălţată cu negru 100%
Gameţi A a A a
Page 10
10
F2 AA Aa Aa aa
Bălţată cu negru Bălţată cu roşu
Rf = 3 : 1; Rg = 1 : 2 : 1
Fig.4 Schema monohibridării care demonstrează dominanţa completă
Concluzie: gena determinantă a culorii bălţate cu negru (A) este complet
dominantă asupra alelei sale care determină culoarea bălţată cu roşu (a).
Dihibridarea presupune încrucişarea între indivizi care se deosebesc prin
două perechi de caractere (două perechi de gene), la fiecare pereche de gene
existând interacţiune de dominanţă completă (fig.5).
taurine taurine
negre fără coarne roşii cu coarne
P ♂ AABB x aabb ♀
Gameţi AB AB ab ab
F1 AaBb x AaBb negre fără coarne 100%
F2 A – B – negre fără coarne (9/16)
A – bb negre cu coarne (3/16)
aaB – roşii fără coarne (3/16)
aabb roşii cu coarne (1/16)
Fig. 5 Schema dihibridării care demonstrează dominanţa completă
Hibrizii heterozigoţi din generaţia F1 (AaBb) produc fiecare câte 4 gameţi
(AB, Ab, aB, ab), din combinarea cărora, cu şanse egale, rezultă 16 genotipuri în
generaţia F2 şi un raport de segregare fenotipică egal cu 9:3:3:1.
Concluzie: În acest tip de interacţiune, gena care determină culoarea neagră (A)
este complet dominantă asupra alelei sale care determină culoarea roşie (a) iar
gena care determină lipsa coarnelor (B) este complet dominantă asupra alelei sale
care determină prezenţa coarnelor (b).
Dominanţa incompletă (semidominanţa) este interacţiunea dintre două
gene alele, în care o genă de la un locus (A1) este incomplet dominantă asupra
alelei sale (A2). Din această cauză, indivizii heterozigăţi (A1A2) vor avea un
fenotip intermediar formelor parentale homozigote (fig. 6 ).
Page 11
11
După dominanţa incompletă se transmite culoarea robei (părului) la rasa
de taurine Shorthorn, culoarea penajului la găinile de Andaluzia, culoarea florilor
la Barba împăratului, garoafe etc.
penaj negru penaj alb
P ♂ A1A1 x A2A2 ♀
F1 A1A2 – penaj gri-albăstrui
F2 A1A1 A1A2 A1A2 A2A2
negru gri-albăstrui alb
Rf = 1 : 2 : 1; Rg = 1 : 2 : 1
Fig. 6 Schema dominanţei incomplete
Codominanţa este interacţiunea în care o genă de la un locus nu prezintă
nici un fel de dominanţă asupra alelei sale (codominanţa = lipsă de dominanţă
între alele).
Codominant se transmit grupele sangvine la om şi animale, proteinele din
sânge (hemoglobinele, transferinele), proteinele din lapte (k-cazeina, β-
lactoglobulina) etc.
Grupele sangvine din sistemul AB0 la om (A, B, AB şi 0) sunt controlate
de trei gene alele: LA, L
B şi l. Numai între alelele L
A şi L
B există interacţiune de
codominanţă (lipsă de dominanţă) astfel că indivizi cu genotip heterozigot (
LAL
B) vor avea grupa sangvină AB, adică vor manifesta fenotipul determinat de
ambele alele (fig. 7).
grupa A grupa B
P ♂ LAL
A x L
BL
B ♀
F1 LAL
B – 100% grupa AB
F2 LAL
A L
AL
B L
AL
B L
BL
B
grupa A grupa AB grupa B
Fig. 7 Schema codominanţei
Page 12
12
În mod asemănător, din împerecherea unor tauri cu tipul A de
hemoglobină (genotip HbAHb
A) cu vaci cu tipul B (genotip Hb
BHb
B) vor rezulta
în generaţia F1 descendenţi cu tipul AB de hemoglobină (genotip HbAHb
B), iar în
generaţia F2: 1/4 cu hemoglobină A, 2/4 cu hemoglobină AB şi 1/4 cu
hemoglobină de tip B. Din acest exemplu, rezultă că între cele două alele HbA
şi
HbB
există interacţiuni de codominanţă (lipsă de dominanţă).
Supradominanţa este o interacţiune dintre genele alele care aflate în stare
heterozigotă se completează reciproc. În acest fel, organismele heterozigote
prezintă un fenotip superior formelor parentale homozigote.
Interacţiunea de supradominanţă implică mai mulţi loci aflaţi în stare
heterozigotă care determină caracterele cantitative (producţia de lapte, producţia
de lână, ouă etc). Supradominanţa, împreună cu dominanţa completă şi cu
epistazia, determină apariţia efectului heterozis, manifestat de hibrizii rezultaţi
din încrucişarea a două sau mai multe rase, soiuri sau linii. De exemplu, hibrizii
de carne la porcine, hibrizii de carne şi ouă la păsări realizează însuşiri de
producţie superioare formelor parentale (raselor sau liniilor), datorită efectului
heterozis.
Genele letale sunt gene care în stare homozigotă produc moartea unor
genotipuri în diferite stadii ale dezvoltării ontogenetice (zigot, embrion, făt sau
postnatal).
Genele letale pot fi: dominante (A+) sau recesive (a
+) şi ele modifică
raporturile normale de segregare.
Letalitatea determinată de gene letale dominante se întâlneşte la ovinele
din rasa Caracul brumăriu, Ţurcană brumărie, vulpile platinate, crapul oglindă,
şoarecii galbeni etc.
De exemplu, din încrucişarea unor berbeci Caracul brumăriu (A+a) cu oi
din aceeaşi rasă (A+a) vor rezulta în prima generaţie: 25% miei brumării (A
+A
+)
care mor postnatal de boala numită timpanism; 50% miei brumării viabili (A+a) şi
25% miei de culoare neagră viabili (aa). Deci, ovine adulte homozigote (A+A
+)
de culoare brumărie nu există, ci numai ovine purtătoare a genei letale (A+a).
Letalitatea determinată de gene letale recesive se întâlneşte la taurinele
din rasa Dexter cu membre scurte, la unele rase de găini numite "târâtoare" etc.
De exemplu, rasa de taurine Dexter cu membre scurte este purtătoare a
unei gene letale recesive în stare heterozigotă (Aa+). Din încrucişarea unor tauri
cu vaci din rasa Dexter (Aa +
) vor rezulta în descendenţă: 25% viţei cu membre
Page 13
13
normale (AA), 50% viţei cu membre scurte (Aa+) şi 25% viţei care mor (a
+a
+) de
o boală a sistemului osos, numită acondroplazie (lipsa câtorva vertebre).
4.2. Interacţiunea genelor nealele
Genele nealele sunt gene situate la loci diferiţi pe o pereche de cromozomi
sau pe perechi de cromozomi diferite. Genele nealele se notează cu litere diferite,
de exemplu A şi B; A şi b; a şi B sau a şi b.
Dintre interacţiunile genelor nealele fac parte: interacţiunea
complementară, interacţiunea de epistazie şi interacţiunea aditivă a genelor. În
toate cazurile, două sau mai multe perechi de gene situate la loci diferiţi
interacţionează pentru determinarea unui singur caracter (fenotip).
Interacţiunea complementară a genelor este interacţiunea în care două
perechi de gene nealele se completează reciproc determinând fenotipuri diferite.
Această interacţiune poate fi de două feluri:
a) Interacţiunea complementară între gene nealele (A şi B) care singure
determină fenotipuri asemănătoare. În acest tip de interacţiune, două perechi de
gene de la loci diferiţi interacţionează complementar pentru determinarea unui
singur caracter (fenotip). De exemplu, din încrucişarea unei rase de găini cu
penajul alb (AAbb) cu altă rasă tot cu penajul alb (aaBB), în generaţia F1 rezultă
descendenţi cu penajul de culoare roşcată, iar în F2 se obţin 9/16 păsări cu penajul
roşcat şi 7/16 păsări cu penajul alb.
Negerhün alb Dorking alb
P ♂ AAbb x aaBB ♀
F1 AaBb – penaj roşcat
F2 A – B – 9/16 penaj roşcat
A – bb 3/16
aaB – 3/16 7/16 cu penaj alb
aabb 1/16
În F1 şi F2 genele nealele A şi B se completează reciproc şi determină
apariţia unui nou fenotip (penajul roşcat).
b) Interacţiunea complementară între gene nealele (A şi B) care singure
determină fenotipuri diferite
Din încrucişarea unor rase de găini cu creasta bătută (AAbb) cu alte
Page 14
14
rase care au creasta în formă de rozetă (aaBB) rezultă în F1 găini cu creasta
nuciformă, iar în F2: 9/16 cu creasta nuciformă; 3/16 cu creasta bătută; 3/16 cu
creasta rozetă şi 1/16 cu creasta dinţată.
creastă bătută creastă rozetă
P ♂ AAbb x aaBB ♀
F1 AaBb – creastă nuciformă
F2 A – B – 9/16 creastă nuciformă
A – bb 3/16 creastă bătută
aaB – 3/16 creastă rozetă
aabb 1/16 creastă dinţată
Creasta nuciformă rezultă din interacţiunea complementară a genelor
nealele dominante A şi B, iar creasta dinţată din interacţiunea genelor nealele
recesive a şi b.
Interacţiunea de epistazie este interacţiunea în care o genă de la un
anumit locus inhibă efectul genelor de la alt locus.
Gena care inhibă se numeşte genă epistatică iar genele inhibate se numesc
gene hipostatice.
Genele epistatice pot fi dominante (A) sau recesive, dar în stare
homozigotă (aa). Genele inhibate (B şi b) sunt de obicei gene cromogene. În
funcţie de natura genei epistatice, epistazia poate fi de două feluri: de dominanţă
şi de recesivitate.
Epistazia de dominanţă este un tip de interacţiune între gene nealele în
care o genă dominantă de la un locus (A) inhibă genele nealele (B şi b).
inhibă
Gena A genele B şi b
B = neagră gene cromogene
b = brună
câini albi câini bruni
P ♂ AABB x aabb ♀
F1 AaBb – albi
F2 A – B – 9/16 12/16 albi
A – bb 3/16
aaB – 3/16 negri
Page 15
15
aabb 1/16 bruni
Rf = 12 : 3 : 1
Concluzie: Toate genotipurile care conţin gena epistatică (A) vor manifesta
culoarea albă, deoarece genele cromogene (B şi b) sunt inhibate.
Epistazia de recesivitate este interacţiunea între gene nealele în care o
genă recesivă de la un locus, în stare homozigotă inhibă genele nealele B şi b.
inhibă
Gena aa genele B şi b
B = gri
b = negri gene cromogene
cobai gri cobai albi
P ♂ AABB x aabb ♀
F1 AaBb – gri
F2 A – B – 9/16 gri
A – bb 3/16 negri
aaB – 3/16
aabb 1/16 4/16 albi
Rf = 9 : 3 : 4
Concluzie: Toate genotipurile care conţin gena epistatică (aa) vor manifesta
culoarea albă, deoarece genele B şi b sunt inhibate. Culoarea gri şi neagră se
manifestă în lipsa genei epistatice (aa).
Interacţiunea aditivă a genelor (poligenia) reprezintă tipul de
interacţiune după care se transmit toate caracterele cantitative (producţia de lapte,
de lână, talie, spor mediu zilnic etc). Caracterele cantitative sunt determinate de
mai multe perechi de gene nealele numite gene aditive, poligene sau gene minore.
Aceste gene îşi adiţionează (cumulează) efectul pentru determinarea valorii unui
caracter cantitativ. Fiecare poligenă are un efect minor asupra valorii caracterului
cantitativ. Când un caracter este determinat exclusiv de poligene, valoarea medie a
descendenţilor în generaţia F1 şi F2 se va situa pe media părinţilor.
Să presupunem în mod ipotetic că producţia de lână la ovine este
determinată numai de interacţiunea a două perechi de gene aditive: genele A şi B
determină fiecare câte 2 kg lână, iar genele a şi b determină fiecare câte 1 kg lână.
Merinos Ţigaie
8kg 4 kg
P ♂ AABB x aabb ♀
Page 16
16
F1 AaBb – 6 kg
F2 AABB (1/16) - 8 kg
AABb (2/16); AaBB (2/16) = 4/16 - 7 kg
AaBb (4/16); AAbb (1/16); aaBB (1/16) = 6/16 - 6 kg
Aabb (2/16); aaBb (2/16) = 4/16 - 5 kg
aabb (1/16) - 4 kg
Rf = 1 : 4 : 6 : 4 : 1
kg 8 7 6 5 4
Valoarea fenotipică (producţia de lână) manifestată de fiecare genotip este
rezultatul adiţionării efectelor pe care le au genele prezente în genotip (de
exemplu AABb = 2 + 2 + 2 + 1 kg = 7 kg sau aabb = 1 + 1 +1 + 1 kg = 4 kg).
Teme de control
1. Din încrucişarea unor taurine din rasa Brună fără coarne cu taurine cu coarne
din aceeaşi rasă s-a obţinut în F2 un număr de 62 viţei fără coarne şi 20 viţei cu
coarne.
- Stabiliţi genotipurile părinţilor şi a hibrizilor F1 şi F2.
- Stabiliţi raportul de segregare fenotipic şi genotipic în F2, exprimat în părţi şi
întocmiţi schema de hibridare.
- Precizaţi după ce acţiune a genelor s-a transmis caracterul.
2. La taurine, culoarea neagră este determinată de o genă dominantă (A) iar
culoarea roşie de alela sa recesivă (a).
Un taur negru este împerecheat cu trei vaci:
- cu vaca A care este roşie a produs un viţel de culoare roşie;
- cu vaca B care este roşie a produs un viţel de culoare neagră;
- cu vaca C care este neagră a produs un viţel de culoare roşie.
Care este genotipul taurului şi a celor trei vaci?
Întocmiţi schema de hibridare.
3. La om, ochii căprui sunt determinaţi de o genă dominantă (B) iar ochii albaştri
de alela sa recesivă (b).
Care este probabilitatea ca primul copil dintr-o familie în care ambii soţi au
ochii căprui să fie cu ochi albaştri? Care esta probabilitatea ca din această familie
să rezulte copii cu ochi căprui?
Întocmiţi schema de hibridare şi precizaţi după ce acţiune a genelor s-a
transmis caracterul.
Page 17
17
4. Din încrucişarea unor cocoşi de culoare neagră şi cu creasta bătută (AABB) cu
găini de culoare roşie şi creasta dinţată (aabb) rezultă în generaţia F2, 16
combinaţii genotipice. Între genele alele A şi a, pe de o parte şi B şi b pe de altă
parte, există interacţiune de dominanţă completă.
- Întocmiţi schema de hibridare, stabiliţi fenotipurile şi raportul de segregare
fenotipică în F2;
- Stabiliţi raportul de segregare fenotipică rezultat în urma retroîncrucişării
unui diheterozigot şi a unui homozigot dominant din F2 cu un genotip
homozigot recesiv;
5. La om, gena B = ochi căprui, gena b = ochi albaştri, gena R = dreptaci şi gena r
= stângaci.
Un bărbat cu ochii căprui şi dreptaci se căsătoreşte cu o femeie cu ochii albaştri şi
dreptace. Primul copil are ochii albaştri şi este stângaci. Ce caractere şi cu ce
probabilităţi vor apare ceilalţi copii care se vor naşte din această familie?
Întocmiţi schema hibridării şi precizaţi despre ce tip de acţiune a genelor este
vorba.
6. Taurinele din rasa Shorthorn de culoare piersicie provin din încrucişarea unei
rase de taurine cu roba de culoare roşie (A1A1) cu o altă rasă cu roba de
culoare albă (A2A2).
Stabiliţi care este genotipul taurinelor de culoare piersicie şi care va fi raportul
fenotipic rezultat din următoarele încrucişări, ştiind că între cele două alele A1 şi
A2 există o interacţiune de dominanţă incompletă:
a) tauri piersicii x vaci piersicii
b) tauri piersicii x vaci roşii.
c) tauri roşii x vaci albe
7. Din încrucişarea unei varietăţi de garoafe cu flori roşii (A1A1) cu o altă
varietate cu flori albe (A2A2), în generaţia F1 rezultă garoafe cu flori roz
(A1A2), iar în generaţia F2 se produce segregarea în raport de 1 : 2 : 1.
Întocmiţi schema de hibridare până în generaţia F2 şi precizaţi după ce
interacţiune a genelor s-a transmis culoarea la garoafe.
8. Tipurile de hemoglobine din sange şi tipurile de transferine serice la rasele de
taurine europene se transmit după interacţiunea codominantă a genelor.
Stabiliţi raportul de segregare fenotipică rezultat din următoarele încrucişări:
a) Tauri cu tipul “A” de transferine cu vaci cu tipul “D” de transferine;
b) Tauri cu tipul “AD” de transferine cu vaci cu tipul “AE” de transferine;
c) Tauri cu tipul “AB” de hemoglobină cu vaci cu tipul “AB” de hemoglobină.
9. Din încrucişarea unor taurine între ele s-au obţinut în generaţia F1 100%
descendenţi cu tipul AB de hemoglobină. Din aceşti descendenţi, 50% aveau
tipul D de transferină şi 50% aveau tipul DE.
Care sunt genotipurile părinţilor şi a descendenţilor?
Întocmiţi schema de hibridare şi precizaţi după ce acţiune a genelor s-au
transmis cele două caractere.
10. a) Precizaţi dacă paternitatea se confirmă sau nu se confirmă în următorul caz:
- copilul, grupa 0;
- mama grupa B;
- tatăl prezumtiv grupa AB.
Page 18
18
b) Un copil are grupa sangvină A, mama are grupa 0. Ce grupe posibile poate
avea tatăl?
Întocmiţi schemele de hibridare.
11. Starea de fecunditate a cocoşilor din rasa Wyandotte alb este determinată de
una din interacţiunile genelor alele B şi b. Cocoşii cu genotip BB au creasta
bătută şi fecunditate scăzută, cocoşii cu genotip Bb au creasta bătută şi
fecunditate normală şi cei cu genotip bb au creasta dinţată şi fecunditate
normală. Acest fenomen nu s-a observat la femele. Despre ce interacţiune a
genelor alele este vorba şi la care genotip al cocoşilor este prezentă?
12. Daţi un exemplu de hibrid de carne trirasial la porcine, precizaţi ce efect
manifestă acest hibrid şi care sunt interacţiunile genelor care determină acest
efect.
13. Vulpile platinate sunt purtătoare a unei gene letale în stare heterozigotă (A+a).
a) Care va fi raportul de segregare rezultat din încrucişarea unor masculi şi
femele de culoare platinată?
b) Care va fi raportul de segregare rezultat din încrucişarea unor femele
palatinate cu masculi cu argintii (aa) ?
Precizaţi despre ce tip de interacţiune a genelor este vorba.
14. Găinile având caracterul “creeper” (membre foarte scurte) sunt heterozigote
pentru o genă letală recesivă (Cc+).
Care va fi raportul de segregare rezultat din încrucişarea unor cocoşi şi găini
cu caracterul “creeper”?
Precizaţi despre ce interacţiune a genelor este vorba.
15. Acondroplazia este un tip de piticism la om care afectează o persoană din
10.000 şi este determinată de o alelă a unei gene. Două personae cu
acondroplazie (genotipul Aa) se căsătoresc şi au dat naştere la un copil pitic şi
unul normal.
Precizaţi dacă acondroplazia este determinată de către o alelă recesivă sau de o
alelă dominantă.
Stabiliţi genotipurile părinţilor, a descendenţilor şi întocmiţi schema de
hibridare până în generaţia F1.
16. Să presupunem că, culoarea albă a părului la două rase de porcine este
determinată de interacţiunea a două perechi de gene nealele.
Precizaţi care va fi fenotipul hibrizilor F1 şi F2 rezultaţi din încrucişarea unor
vieri din rasa Marele alb (AAbb) cu scroafe din rasa Landrace alb (aaBB), ştiind
că în F1 hibrizii vor fi de culoare neagră, iar în F2 se produce segregarea, rezultând
descendenţi de culoare albă şi de culoare neagră.
Întocmiţi schema de hibridare, stabiliţi raportul fenotipic de segregare în F2 şi
precizaţi despre ce tip de interacţiune este vorba.
17. Concepeţi o problemă de interacţiune complementară între gene nealele care
singure determină fenotipuri asemănătoare, la specia ovine, luând în
considerare culoarea lânii, cu menţiunea că rasa Merinos alb are genotipul
AAbb, rasa Ţigaie albă are genotipul aaBB, iar hibrizii rezultaţi în generaţia F1
au lâna de culoare neagră. Întocmiţi schema de hibridare până în generaţia F2
şi stabiliţi fenotipurile descendenţilor rezultaţi.
Page 19
19
18. Concepeţi o problemă de interacţiune complementară între gene nealele care
singure determină fenotipuri diferite la iepuri, luând în considerare culoarea
blăniţelor (gri, alb cu extremităţi negre, agouti şi albinotic).
19. Se încrucişează găini dintr-o rasă cu penajul alb (AABB) cu cocoşi dintr-o
rasă cu penajul roşu (aabb).
epist.
Gena A B şi b
B = culoarea porumbacă
b = culoarea roşie
Întocmiţi schema de încrucişare şi stabiliţi raportul de segregare fenotipică
în F2.
Precizaţi după ce tip de interacţiune a genelor se transmite acest caracter.
20. Se încrucişează porcine de culoare neagră (AABB) cu porcine de culoare albă
(aabb).
epist.
Genele aa B şi b
B = culoarea neagră
b = culoarea roşie
Cele două culori se vor manifesta numai în prezenţa genei A care
controlează sinteza unei enzime cu rol în formarea pigmentului melanic.
Precizaţi despre ce tip de interacţiune este vorba.
21. Se încrucişează berbeci cu lâna albă din rasa Merinos (AABB) cu oi din rasa
Ţurcană cu lâna albă (aabb).
epist.
A B şi b
epist.
aa bb
B – culoare neagră; b – culoare brumărie
Întocmiţi schema de hibridare până în F2 şi stabiliţi raportul de segregare
fenotipic.
Precizaţi despre ce tip de interacţiune a genelor este vorba.
22. Din încrucişarea unor rase de taurine cu producţia de lapte de 6000 kg
(AABB) cu rase a căror producţie medie este de 4000 kg (aabb) se obţin în F1
descendenţi cu producţii intermediare părinţilor, iar în generaţia F2 16
genotipuri repartizate în 5 clase fenotipice.
Să se întocmească schema de hibridare şi să se stabilească raportul de
segregare fenotipic în F2.
Precizaţi după ce tip de interacţiune a genelor s-a transmis caracterul.
23. La ovine, gena A determină 2 kg lână, gena B determină 2 kg lână, gena a şi b
câte 1 kg fiecare.
- Stabiliţi ce producţie de lână vor avea descendenţii rezultaţi în F1 din
încrucişarea unor berbeci cu 8 kg lână cu oi cu 4 kg lână;
- Ce producţie vor avea descendenţii obţinuţi din încrucişarea hibrizilor F1
(masculi) cu oi cu producţia de 4 kg?
- Ce producţie vor avea descendenţii obţinuţi din încrucişarea hibrizilor F1 (6
kg) cu oi cu producţie de 7 kg lână?
Page 20
20
Cum pot fi explicate aceste rezultate?
CAPITOLUL 5
TRANSMITEREA CARACTERELOR
DETERMINATE DE GENE SITUATE PE
ACELAŞI CROMOZOM
În urma experienţelor de dihibridare efectuate la musculiţa de oţet
(Drosophila melanogaster), Morgan elaborează teoria cromozomială a eredităţii cu
cele două teze: transmiterea înlănţuită a genelor situate pe acelaşi cromozom
(linkage-ul) şi schimbul reciproc de gene între cromozomii omologi
(crossingover-ul).
Linkage-ul
Morgan obţine într-o experienţă de dihibridare raportul de segregare de 3 :
1 în loc de 9 : 3 : 3 : 1, datorită aşezării genelor nealele (A şi B) pe acelaşi
cromozom şi transmiterii lor înlănţuite.
corp cenuşiu corp negru
aripi normale aripi vestigiale
P ♂ AABB x aabb ♀
F1 AaBb – corp cenuşiu, aripi normale
F2 AABB AaBb AaBb aabb
corp cenuşiu corp negru
aripi normale aripi vestigiale
Rf = 3 : 1 diferit de 9 : 3 : 3 : 1
Concluzie: La hibrizii F1, genele dominante A şi B sunt situate pe un cromozom,
iar alelele lor recesive a şi b pe cromozomul omolog A B
a b .
Datorită linkageului, hibrizii heterozigoţi din F1 (AaBb) vor produce numai doi
gameţi (AB şi ab) şi nu patru gameţi (AB, Ab, aB şi ab) ca şi în dihibridare, când
genele erau aşezate pe cromozomi omologi diferiţi şi segregau independent.
Page 21
21
Crossingover–ul sau schimbul reciproc de gene între cromozomii
omologi, are loc în cursul meiozei şi în urma lui se obţin de la un organism dublu
heterozigot (AaBb) 4 tipuri de gameţi: gameţi nerecombinaţi (AB şi ab) şi gameţi
recombinaţi (Ab şi aB). Întotdeauna, gameţii recombinaţi sunt într-o proporţie mai
mică.
Crossingoverul se produce între genele nealele (A şi B) situate pe acelaşi
cromozom (înlănţuite).
corp cenuşiu corp negru
aripi normale aripi vestigiale
P ♂ AABB x aabb ♀
corp cenuşiu
aripi normale
F1 ♀ AaBb x aabb ♂ retroîncrucişare
Gameţi AB ab Ab aB ab
A B a b A b a B
R1 a b a b a b a b
cenuşiu negru cenuşiu negru
ar. normale ar. vestigiale ar. vestigiale ar. normale
40% 40% 10% 10%
20% organisme recombinate
Concluzie: Raportul de segregare obţinut în urma retroîncrucişării fiind diferit de
1:1:1:1 indică faptul că la hibrizii F1(AaBb) genele dominante sunt situate pe un
cromozom, iar cele recesive pe cromozomul omolog A B
a b ,
dar între ele s-a produs crossingoverul, rezultând la femelă 4 tipuri de gameţi.
Frecvenţa cu care apar organismele recombinate indică frecvenţa crossingoverului
dintre genele A şi B (20%).
Hărţile genetice cromozomiale sunt reprezentări grafice ale
locilor în care se găsesc plasate genele pe cromozomii unei specii. Etapele
întocmirii unei hărţi genetice cromozomiale sunt:
Page 22
22
a) Identificarea genelor situate pe acelaşi cromozom (a grupelor de linkage) se
face în funcţie de raportul de segregare obţinut în urma unei retroîncrucişări
dintre un organism dublu heterozigot (AaBb) şi un organism dublu homozigot
recesiv (aabb). Dacă raportul obţinut este diferit de 1 : 1: 1 : 1, înseamnă că
genele A şi B sunt situate pe acelaşi cromozom. Dacă raportul este egal cu 1 :
1 : 1 : 1 înseamnă că genele A şi B sunt situate pe cromozomi omologi diferiţi.
b) Stabilirea distanţei dintre genele situate pe acelaşi cromozom. Distanţa dintre
gene se măsoară în centimorgani (CM). Un centimorgan este egal cu 1% din
frecvenţa crossingoverului (1% din frecvenţa organismelor recombinate).
c) Stabilirea succesiunii genelor situate pe acelaşi cromozom se face în funcţie
de distanţa dintre gene, respectiv de procentul organismelor recombinate care
apar în urma retroîncrucişării.
De exemplu, dacă există trei gene situate pe acelaşi cromozom (A, B şi C) şi
între genele A şi B s-au obţinut organisme recombinate în proporţie de 20%, între
B şi C 10%, iar între A şi C 30%, distanţele dintre cele trei gene şi succesiunea
genelor pe cromozom va fi ABC.
A 20 CM B 10CM C
30 CM
Dacă între genele A şi C s-ar fi obţinut organisme recombinate în proporţie
de 10%, distanţele şi succesiunea genelor pe cromozom ar fi ACB.
A 10 CM C 10 CM B
20 CM
Teme de control
1. La unele rase de găini gena F = penaj frizat; f = penaj normal; I = penaj alb şi i
= penaj colorat.
Din încrucişarea unor cocoşi cu penajul frizat şi alb cu găini cu penajul normal
şi colorat s-au obţinut în F1 descendenţi cu caracterele dominante. Din
retroîncrucişarea găinilor hibride F1 cu cocoşi dublu homozigoţi recesivi s-au
obţinut în R1 două fenotipuri cu următorul număr de indivizi: 21 capete cu penajul
frizat şi de culoare albă şi 20 capete cu penajul normal şi colorat. Întocmiţi
schema de hibridare, explicaţi fenomenul şi stabiliţi modul în care au fost plasate
genele pe cromozomi (pe aceeaşi pereche de cromozomi sau pe perechi diferite de
cromozomi).
2. Din încrucişarea Drosophilelor cu ochii roşii şi membre nepăroase (caractere
domiannte) cu Drosophile cu ochii purpurii şi membre păroase (caractere
recesive) se obţin în F1 hibrizi cu caractere dominante. Prin retroîncrucişarea
Page 23
23
hibrizilor cu forma homozigotă recesivă s-a obţinut în generaţia de
retroîncrucişare 42 musculiţe cu ambele caractere recesive.
Întocmiţi schema de hibridare, stabiliţi raportul exprimat în părţi şi explicaţi
fenomenul ştiind că cele două gene nealele se află pe acelaşi cromozom.
3. Din încrucişarea unor cocoşi cu penaj frizat (zburlit) şi colorat (FFii) cu găini
cu penajul normal şi alb (ffII) în F1 s-au obţinut descendenţi cu penajul frizat şi
alb. Prin retroîncrucişarea hibrizilor dublu heterozigoţi din F1 cu indivizi dublu
heterozigoţi recesivi s-au obţinut în R1 patru fenotipuri cu următorul număr de
indivizi: 63 capete, 63 capete, 18 capete, 13 capete.
Intocmiţi schema de analiză genetică şi explicaţi fenomenul care duce la
obţinerea celor 4 fenotipuri în proporţii diferite, ştiind că genele F şi i, pe de o
parte şi genele f şi I, pe de altă parte, sunt situate pe acelaşi cromozom.
4. La Drosophila, genele D = picioare lunfi, d = picioare scurte, B = corp gri, b =
corp negru, P = ochi roşii, p = ochi purpurii, E = ochi normali şi e = ochi barr.
Din încrucişarea unor femele heterozigote pe câte două perechi de gene cu
masculi dublu recesivi, s-au obţinut următoarele rezultate: a) picioare lungi şi
corp gri
40%
picioare lungi şi
corp negru
10%
picioare scurte şi
corp gri
8%
picioare scurte şi
corp negru
42%
b) corp gri şi
ochi roşii
9,5%
corp gri şi
ochi purpurii
40,5%
corp negru şi
ochi roşii
42%
corp negru şi
ochi purpurii
8%
c) picioare lungi şi
ochi normali
25%
picioare lungi şi
ochi barr
25%
picioare scurte şi
ochi normali
25%
picioare scurte şi
ochi barr
25%
d) picioare lungi şi
ochi roşii
18%
picioare lungi şi
ochi purpurii
32%
picioare scurte şi
ochi roşii
32,5%
picioare scurte şi
ochi purpurii
17,5%
Întocmiţi harta genetică, stabilind care din cele patru gene nealele sunt
situate pe acelaşi cromozom, distanţa dintre gene şi poziţia (cis şi trans) a genelor
situate pe acelaşi cromozom.
CAPITOLUL 6
TRANSMITEREA ÎNLĂNŢUITĂ CU SEXUL,
SAU FENOMENUL DE SEX - LINKAGE
Unele caractere somatice cum sunt: culoarea penajului la unele rase de
găini, gradul de îmbrăcare cu penaj, piticismul la găini sunt determinate de gene
situate pe cromozomul sexului sau pe heterozomul Z. De asemenea, unele boli la
om, cum sunt daltonismul şi hemofilia sunt determinate de gene recesive situate pe
heterozomul X. Genele situate pe heterozomii Z sau X se transmit odată cu
aceştia, fenomen denumit sex – linkage.
Heterozomii W de la păsări şi Y de la mamifere nu conţin astfel de gene.
Page 24
24
Caracterele determinate de gene situate pe heterozomi se transmit la
descendenţi altfel decât caracterele determinate de gene situate pe autozomi.
6.1. Transmiterea înlănţuită cu sexul la tipul Abraxas
Păsările prezintă un determinism al sexelor de tip Abraxas, caracterizat
prin prezenţa a doi heterozomi identici (ZZ) la mascul şi a doi heterozomi diferiţi
(ZW) la femelă.
Fenomenul de sex – linkage are aplicaţii practice în avicultură servind la
stabilirea sexului, în prima zi de la ecloziune, pe baza culorii penajului sau a
gradului de îmbrăcare cu penaj. Procedeul de stabilire a sexului după aceste
caractere determinate de gene heterozomale poartă numele de autosexare.
Autosexarea este posibilă numai în generaţia F1 şi numai atunci când se
încrucişează cocoşi cu caracterul recesiv cu găini având caracterul dominat. Gena
S (silver) determină culoarea argintie, iar gena s determină culoarea roşie a
penajului. Aceste gene sunt gene heterozomale situate pe cromozomul Z.
Astfel, din încrucişarea unor cocoşi din rasa Cornish cu penajul de culoare
roşie (ZsZ
s) cu găini din rasa Rock cu penajul de culoare argintie (Z
SW) se obţin
în prima generaţie F1 pui de carne autosexabili după culoarea penajului. Astfel,
toţi cocoşeii vor fi de culoare argintie şi toate puicuţele vor fi de culoare roşie.
Continuându-se împerecherile între hibrizii obţinuţi în prima generaţie, în
generaţia a doua F2 se obţine următorul raport de segregare: 50% cocoşi argintii,
50% cocoşi roşii; 50% puicuţe argintii, 50% puicuţe roşii. Se constată că în F2 nu
este posibilă autosexarea deoarece apar şi cocoşei şi puicuţe de aceeaşi culoare.
Din hibridarea inversă dintre cocoşi aparţinând rasei Rock argintiu cu găini
din rasa Cornish roşu în F1 toţi cocoşeii şi toate puicuţele manifestă culoarea
dominantă argintie, nefiind posibilă stabilirea sexului după culoarea penajului.
Nici în generaţia F2 nu este posibilă autosexarea, deoarece se obţin cocoşei şi
puicuţe de culoare argintie.
Îmbrăcarea rapidă şi lentă cu penaj la unele rase de găini este determinată
de asemenea, de gene heterozomale situate pe heterozomul Z. Gena L determină
îmbrăcarea lentă cu penaj, iar gena l determină îmbrăcarea rapidă cu penaj. Din
încrucişarea unor cocoşi cu îmbrăcare rapidă (ZlZ
l) cu găini cu îmbrăcare lentă
Page 25
25
(ZLW) se obţin în generaţia F1 descendenţi autosexabili, respectiv cocoşi cu
îmbrăcare lentă şi puicuţe cu îmbrăcare rapidă. În F2 nu mai este posibilă
autosexarea.
De asemenea, piticismul la găini este determinat de o genă recesivă
heterozomală situată pe cromozomul Z.
6.2. Transmiterea înlănţuită cu sexul la tipul Drosophila
Determinismul sexelor la tipul Drosophila (musculiţa de oţet, mamifere) se
caracterizează prin prezenţa a doi heterozomi identici la femele (XX) şi a doi
heterozomi diferiţi la masculi (XY).
La Drosophila melanogaster culoarea roşie şi albă a ochilor este
determinată de două gene alele situate pe heterozomul X. Gena A determină
culoarea roşie a ochilor, iar gena a determină culoarea albă. De exemplu, din
încrucişarea unor masculi cu ochii roşii (XAY) cu femele cu ochi albi (XaXa), în
generaţia F1 toate femelele vor avea ochii roşii şi toţi masculii vor avea ochi albi.
În generaţia F2, 50% din masculi au ochii roşii şi 50% ochii albi, de asemenea, la
femele, 50% au ochii roşii şi 50% ochi albi. Aceste rezultate se obţin datorită
transmiterii înlănţuite a genelor care determină culoarea roşie şi albă a ochilor,
odată cu heterozomul X (fenomenul de sex-linkage).
Hemofilia şi daltonismul la om sunt boli determinate de gene recesive
situate pe heterozomul X (gene sex – linkate). La masculi, aceste gene se
manifestă în stare hemizigotă (XhY sau X
dY) determinând apariţia bolii, iar la
femele ele se manifestă în stare homozigotă (XhX
h sau X
dX
d). Rezultă că
frecvenţa de apariţie a acestor boli la bărbaţi este mai mare decât la femei.
S-a constatat că fetele hemofilice (XhX
h) nu supravieţuiesc deoarece gena
h în stare homozigotă determină moartea acestora, ceea ce însemnă că nu pot
exista femei hemofilice mature.
Aplicaţie: Stabiliţi ce descendenţi vor rezulta în urma unor căsătorii între
un bărbat sănătos şi o femeie purtătoare; între un bărbat hemofilic şi o femeie
sănătoasă şi nepurtătoare a genei pentru hemofilie; între un bărbat hemofilic şi o
femeie purtătoare.
Teme de control
1. La om, daltonismul este determiant de o genă recesivă heterozomală (d), iar
vederea normală de alela sa dominantă (D).
Un bărbat cu vederea normală căsătorit cu o femeie cu vederea normală au
trei copii:
Page 26
26
- un fiu daltonic, care la rândul său căsătorindu-se are o fată normală;
- o fată normală, care căsătorindu-se are un fiu daltonic şi unul normal;
- o fată normală, care căsătorindu-se are cinci fii normali.
Cum se poate explica această situaţie?
2. Să se stabilească raportul se segregare în F1 şi F2 rezultat din încrucişarea unor
cocoşi din rasa Rhode-Island roşu (ZsZ
s) cu găini din rasa Rock porumbac
(ZSW).
Precizaţi despre ce fenomen este vorba şi în care generaţie se poate
determina sexul după culoarea penajului (autosexarea).
3. La găini, îmbrăcarea lentă cu penaj se datoreşte unei gene dominante
heterozomale (L), iar îmbrăcarea rapidă, acţiunii alelei sale recesive (l).
Să se alcătuiască o schemă de împerecheri pentru a se crea un hibrid
autosexabil, folosind caracterele amintite.
4. La găini, piticismul este determinat de acţiunea unei gene recesive
heterozomale (dw), iar creşterea normală de alela sa dominantă (dw+).
Din încrucişarea unui cocoş cu dezvoltare normală cu găini pitice, s-au
obţinut în F1 50% din femele pitice şi 50% din cocoşi normali.
Ce fel de descendenţi şi cu ce probabilităţi pot să mai rezulte din această
încrucişare?
Stabiliţi genotipurile părinţilor şi întocmiţi schema de încrucişare până în
F1. Explicaţi fenomenul.
5. La rasa de găini Cornish roşu, culoarea roşie este determinată de o genă
recesivă (s) plasată pe cromozomul de sex Z. De asemenea, culoarea argintie a
rasei Rock este determinată de o genă dominantă (S) plasată pe cromozomul
de sex Z.
a) Întocmiţi schema de încrucişare dintre un cocoş Cornish roşu şi o găină Rock
argintie până în F1 şi stabiliţi culorile pe sexe.
b) Stabiliţi culorile la descendenţii rezultaţi din încrucişarea hibrizilor F1 între ei.
Precizaţi ce aplicaţii practice are acest fenomen în avicultură.
CAPITOLUL 7
ACIZII NUCLEICI: STRUCTURĂ ŞI FUNCŢII
Acizii nucleici, purtătorii informaţiei genetice, sunt reprezentaţi de acidul
dezoxiribonucleic (ADN) şi acidul ribonucleic (ARN). Ei sunt asociaţi cu
proteinele histonice şi nonhistonice, alcătuind nucleoproteinele care intră în
structura cromozomilor. Gena reprezintă un fragment mai mare sau mai mic din
macromolecula de ADN sau ARN care deţine informaţia necesară pentru sinteza
unei proteine, enzime etc.
Page 27
27
ADN-ul este purtătorul informaţiei genetice la organismele eucariote şi la
unele organisme procariote (bacterii, dezoxiribovirusuri). ADN-ul are o structură
primară şi una secundară.
Structura primară a ADN se referă la elementele din care este alcătuită o
singură catenă. În structura ADN intră bazele azotate purinice (A-adenina, G-
guanina), bazele azotate pirimidinice (C-citozina, T-timina), o pentoză
(dezoxiriboza) şi un radical fosforic.
O catenă polinucleotidică este alcătuită din unităţi mai simple numite
nucleotide. O nucleotidă este alcătuită dintr-o bază azotată purinică sau
pirimidinică, o moleculă de dezoxiriboză şi un radical fosforic. Prin legarea unei
baze azotate cu dezoxiriboza rezultă un nucleosid. Legarea unui radical fosforic
de nucleosid dă naştere la un nucleotid. Polimerizarea nucleotidelor de pe o
catenă se face deci prin intermediul radicalului fosforic, care leagă două
nucleoside învecinate prin carbonii 3' şi 5
' din pentoze.
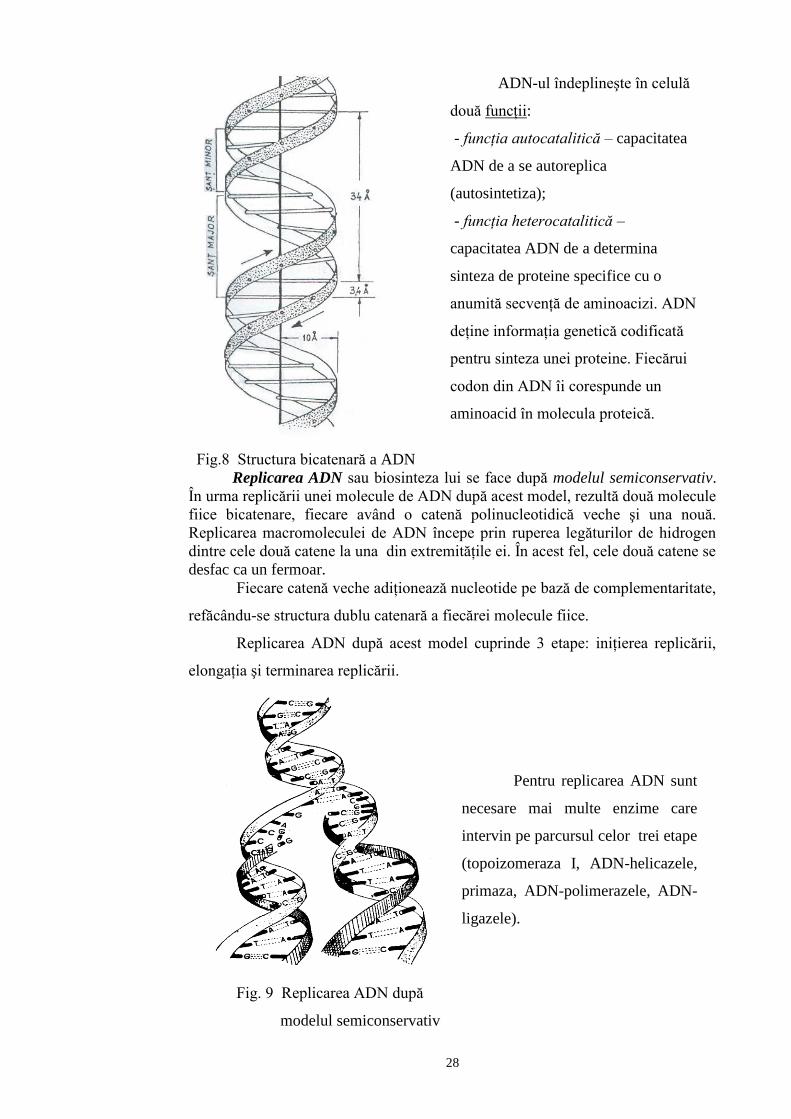
Structura secundară a ADN se referă la structura dublu catenară (dublu
helix) a macromoleculei de ADN. Cele două catene ale ADN sunt înfăşurate
helicoidal în jurul unui ax longitudinal imaginar. Una din catene are direcţie
ascendentă ( 3'
5' ) iar cealaltă catenă are direcţie descendentă ( 5
' 3
' ).
Cele două catene sunt legate între ele prin legături de hidrogen, care se realizează
între o bază azotată purinică de pe o catenă şi o bază azotată pirimidinică de pe
cealaltă catenă. Între adenină şi timină există legături de hidrogen duble ( A = T)
iar între guanină şi citozină legături triple ( G = C).
În cadrul fiecărei specii, cantitatea de A este egală cu cantitatea de T, iar
cantitatea de G este egală cu cea de C.
Page 28
28
ADN-ul îndeplineşte în celulă
două funcţii:
- funcţia autocatalitică – capacitatea
ADN de a se autoreplica
(autosintetiza);
- funcţia heterocatalitică –
capacitatea ADN de a determina
sinteza de proteine specifice cu o
anumită secvenţă de aminoacizi. ADN
deţine informaţia genetică codificată
pentru sinteza unei proteine. Fiecărui
codon din ADN îi corespunde un
aminoacid în molecula proteică.
Fig.8 Structura bicatenară a ADN
Replicarea ADN sau biosinteza lui se face după modelul semiconservativ.
În urma replicării unei molecule de ADN după acest model, rezultă două molecule
fiice bicatenare, fiecare având o catenă polinucleotidică veche şi una nouă.
Replicarea macromoleculei de ADN începe prin ruperea legăturilor de hidrogen
dintre cele două catene la una din extremităţile ei. În acest fel, cele două catene se
desfac ca un fermoar.
Fiecare catenă veche adiţionează nucleotide pe bază de complementaritate,
refăcându-se structura dublu catenară a fiecărei molecule fiice.
Replicarea ADN după acest model cuprinde 3 etape: iniţierea replicării,
elongaţia şi terminarea replicării.
Pentru replicarea ADN sunt
necesare mai multe enzime care
intervin pe parcursul celor trei etape
(topoizomeraza I, ADN-helicazele,
primaza, ADN-polimerazele, ADN-
ligazele).
Fig. 9 Replicarea ADN după
modelul semiconservativ
Page 29
29
Topoizomeraza I este o enzimă de restricţie care produce tăieturi (breşe) la
nivelul unei singure catene a ADN-ului.
ADN-helicazele pătrund prin breşele făcute de către topoizomeraza I şi rup
legăturile de hidrogen dintre cele două catene, dând naştere unui ochi de sinteză.
Primaza este o ARN-polimerază care sintetizează o secvenţă de
ribonucleotide numite primer la capetele 3' ale celor două catene ale ADN-ului.
ADN-polimerazele (ADN-polimeraza III la procariote şi ADN-polimeraza
delta-δ la eucariote) sunt enzimele cu rol principal în elongaţie. Această enzimă
catalizează reacţia de formare a unor legături fosfodiesterice dintre gruparea OH-
3' liberă a primerului şi atomul de fosfor 5' ascuns al nucleozidului trifosfat ce este
încorporat la noul capăt al primerului.
Denaturarea şi renaturarea ADN
Prin încălzirea unei soluţii în care se află ADN la 90-94oC se rup legăturile
de hidrogen dintre cele două catene. Dacă răcirea soluţiei se face brusc, rezultă
ADN denaturat (monocatenar), deci nu se refac punţile de hidrogen. Dacă răcirea
soluţiei se face lent, se refac punţile de hidrogen dintre cele două catene şi rezultă
ADN renaturat.
Fenomenul de denaturare - renaturare serveşte la formarea hibrizilor
moleculari de tip ADN – ADN de la specii diferite sau ADN – ARN. Aceşti
hibrizi moleculari servesc la studierea filogeniei speciilor, localizarea mutaţiilor
apărute prin deleţie, sau pentru identificarea prezenţei unor gene în unele celule
(genele care determină sinteza ARNm, ARNr şi ARNt).
ARN–ul este purtătorul informaţiei genetice la unele ribovirusuri (ARN-
viral) sau îndeplineşte alte funcţii în sinteza proteinelor la eucariote şi procariote.
Cea mai mare cantitate de ARN se găseşte în citoplasmă (mitocondrii şi
ribozomi).
Are structură monocatenară, spre deosebire de ADN care este bicatenar.
Catena de ARN rezultă prin polimerizarea mai multor ribonucleotide. O
ribonucleotidă este alcătuită dintr-o bază azotată purinică (A sau G) sau o bază
azotată pirimidinică (C sau U), o pentoză (riboza) şi un radical fosforic. Deci, în
ARN uracilul înlocuieşte timina, iar riboza înlocuieşte dezoxiriboza. După
funcţia, structura şi locul pe care îl ocupă în celulă, există mai multe tipuri de
ARN: ARN mesager, ARN de transport şi ARN ribozomal.
Page 30
30
ARN-mesager (ARNm) se sintetizează în nucleu şi reprezintă 2 – 5% din
cantitatea totală de ARN din celulă. Are rol în transcripţie şi reprezintă, din punct
de vedere al informaţiei genetice, imaginea complementară a catenei de ADN
matriţă (3'
5') după modelul căreia a fost sintetizat. În ARN-m direcţia
bazelor azotate este 5'
3'. El conţine informaţia genetică sub formă
codificată, necesară sintezei unei proteine.
La eucariote, ARNm precursor conţine şi intronii şi exonii în timp ce
ARNm matur conţine numai exonii.
El duce informaţia genetică la nivelul ribozomilor, în citoplasmă.
ARN de transport (ARNt) reprezintă 10 – 15% din cantitatea totală de
ARN din celulă şi are un rol important în procesul de sinteză a proteinelor,
transportând aminoacizii activaţi la nivelul poliribozomilor. Are greutate
molecualră mică, fiind alcătuit din 70-90 nucleotide. Este monocatenar, dar
prezintă şi porţiuni bicatenare (formă de trifoi). La eucariote există peste 40 tipuri
de ARNt care transportă pe cei cca 20 aminoacizi esenţiali.
ARN-ribozomal (ARNr) reprezintă 80-85% din totalul ARN din celulă. Se
găseşte localizat, în principal, în ribozomi. Este alcătuit din 120-1000 nucleotide.
Participă la procesul de sinteză a proteinelor (translaţie) la nivelul
poliribozomilor. Are o compoziţie în nucleotide foarte asemănătoare la diferite
organisme.
Teme de control
1. Din ce este alcătuit un nucleosid şi un nucleotid?
2. Care sunt bazele azotate purinice şi bazele azotate pirimidinice care intră în
structura ADN-ului?
3. La ce se referă structura primară şi cea secundară a ADN-ului?
4. Între care baze azotate există legături duble şi între care există legături triple?
5. Câte molecule fiice de ADN rezultă în urma replicării unei molecule de ADN
după modelul semiconservativ?
6. Ce structură (monocatenară sau bicatenară) are ADN-ul denaturat şi ADN-ul
renaturat?
7. La ce serveşte fenomenul de denaturare-renaturare a ADN-ului?
8. Din punct de vedere structural, care este diferenţa între ADN şi ARN?
9. Care este diferenţa între o dezoxiribonucleotidă şi o ribonucleotidă?
10. Ce se înţelege prin transcripţia informaţiei genetice?
11. Ce sunt exonii şi intronii care intră în structura unei gene şi la care organisme
se întalnesc (eucariote sau procariote)?
12. La eucariote numărul tipurilor de ARNt este egal cu numărul de aminoacizi
esenţiali (20) sau este diferit de acesta?
Page 31
31
CAPITOLUL 8
FUNCŢIONAREA ADN-ULUI ŞI
EXPRESIA GENELOR
Structura genelor la procariote şi eucariote
La procariote, genele au o structură continuă fiind alcătuite numai din
secvenţe informaţionale (exoni). La eucariote, genele au structură discontinuă
fiind alcătuite din secvenţe informaţionale (exoni) şi din secvenţe
noninformaţionale (introni). Exonii sunt secvenţe de nucleotide din ADN care
conţin informaţia genetică pentru sinteza unei proteine cu o anumită secvenţă de
aminoacizi. La fiecare aminoacid din proteină îi corespunde în ADN un codon sau
triplet (secvenţă de trei nucleotide). La procariote, de pe mai multe gene se
sintetizează o singură moleculă de ARNm numit ARN policistronic, în timp ce la
eucariote, de pe fiecare genă se sintetizează o moleculă de ARNm. Intronii sunt
secvenţe de ADN noninformaţional (ADN repetitiv).
La eucariote, în urma transcripţiei se sintetizează de pe fiecare genă o
moleculă de ARNm precursor ce conţine şi exonii şi intronii, apoi intronii sunt
eliminaţi, rezultând ARNm matur, care migrează în citoplasmă la nivelul
ribozomilor, unde duce informaţia genetică pentru sinteza proteinelor.
Codul genetic este asemănător unui alfabet în care cu ajutorul celor 4 baze
azotate (A, G, C, U) se pot forma 43 = 64 cuvinte de cod (codoni). Deci, codul
genetic din ARNm cuprinde 64 codoni, din care numai 61 codoni specifică pe cei
20 aminoacizi esenţiali. Trei codoni (UAA, UAG şi UGA) sunt numiţi codoni
stop sau nonsens, deoarece nu specifică nici un aminoacid şi ei se găsesc la
sfârşitul unui mesaj genetic (gene).
Caracteristicile codului genetic:
1. Codul genetic este universal, deci aceeiaşi codoni codifică acelaşi aminoacid
la toate speciile;
2. Codul genetic este degenerat, adică mai mulţi codoni pot codifica acelaşi
aminoacid;
3. Codul genetic este nesuprapus, deci doi codoni vecini nu-şi împrumută unul
altuia nucleotide;
4. Codul genetic este lipsit de virgule, adică între codonii vecini nu există semne
de punctuaţie, nu există spaţii.
Page 32
32
Între succesiunea codonilor din catena polinucleotidică a ADN sau ARNm
şi succesiunea aminoacizilor dintr-o catenă polipeptidică (proteină) există o
corespondentă numită colinearitate.
Sinteza proteinelor se realizează la nivelul poliribozomilor din
citoplasmă şi cuprinde două etape:
1. Transcripţia, care constă în copierea informaţiei genetice (secvenţei
nucleotidelor) din catena matriţă a ADN-ului în molecula de ARN mesager.
Are loc în nucleu.
2. Translaţia, care constă în decodificarea informaţiei genetice din ARN mesager
şi sinteza unei proteine cu o anumită secvenţă de aminoacizi. La fiecare codon
din ARNm în proteină îi corespunde un aminoacid (din cei cca 20 aminoacizi
esenţiali).
Teme de control
1. La ce se referă structura continuă a genelor de la procariote şi structura
discontinuă a genelor de la eucariote?
2. Ce diferenţă există între ARNm matur şi ARNm precursor la eucariote?
3. Din cei 64 codoni ai codului genetic din ARNm, câţi codifică pe cei 20
aminoacizi esenţiali?
4. Ce se înţelege prin cod genetic universal şi prin cod genetic degenerat?
CAPITOLUL 9
STRUCTURA MOLECULARĂ A
CROMOZOMILOR LA PROCARIOTE ŞI
EUCARIOTE
Sistemele biologice cuprind două tipuri de organizare: organizarea
acelulară (virală) şi organizarea celulară, întâlnită la procariote şi eucariote.
Virusurile au organizare acelulară şi prezintă ca şi material genetic fie
ADN (dezoxiribovirusuri) fie ARN (ribovirusuri) care intră în structura
cromozomului viral.
Unele virusuri conţin o singură moleculă de ADN monocatenar (fagul ФX
174, fagul M 13) care intră în structura cromozomului viral. Majoritatea
virusurilor au cromozomul alcătuit dintr-o moleculă de ADN bicatenar (fagul T2,
Page 33
33
Fagul T4, SV 40, fagul lambda (λ) etc). Un număr mic de virusuri (ribovirusuri) au
cromozomul alcătuit din ARN monocatenar (virusul turbării, virusul gripei,
virusul poliomielitei, virusul MS2, virusul mozaicului tutunului etc).
Celulele procariote (bacterii, actinomicete, alge albastre-verzi,
microorganismele unicelulare etc) nu au nucleul organizat, materialul lor genetic
se găseşte în citoplasmă, fiind nedelimitat de o membrană nucleară şi se numeşte
nucleoid. De exemplu, bacteriile conţin un singur cromozom mare şi câteva
plasmide (ADN circular).
Celulele eucariote au nucleul înconjurat de membrană nucleară ce conţine
mai mulţi cromozomi, fiecare din aceştia conţinând o moleculă de ADN complicat
împăturită.
Cele mai mari diferenţe în structura şi organizarea moleculară există între
cromozomii eucariotelor şi cei ai procariotelor.
9.1. Structura moleculară a cromozomului bacterian
Cromozomul de la E. coli este o unitate condensată obţinută prin pliere şi
superspiralizare numită nucleoid sau cromozom pliat, care conţine o singură
moleculă de ADN bicatenar de formă circulară.
Cromozomul bacterian nu poate exista decât sub formă superspiralizată
(superpliată), deoarece lungimea lui este de 1300 μm (1,3 mm), în timp ce celula
are un diametru de 1-2 μm.
Cromozomul pliat al bacteriei E. coli conţine 40-50 bucle, iar la
cromozomul superpliat fiecare buclă prezintă răsuciri secundare alcătuite din cca
400 perechi nucleotide.
Buclele sunt menţinute cu ajutorul unei ARN cu rol de linker. Pe lângă
ADN şi ARN, cromozomul bacterian conţine şi cantităţi mici de proteine (lipsesc
cele histonice), care sunt responsabile de aranjamentul cu bucle multiple a ADN.
Fig. 10 Structura moleculară a cromozomului bacterian
a) cromozom nepliat; b) cromozom pliat; c) cromozom superpliat
Page 34
34
9.2. Structura moleculară a cromozomului la eucariote
Fiecare cromozom la eucariote conţine o singură moleculă de ADN de
lungime enormă, care se găseşte sub formă pliată şi superpliată. Cromozomii sunt
alcătuiţi din cromatină, care conţine un agregat complex format din ADN şi
proteine histonice şi nonhistonice.
Histonele sunt proteine bazice ce conţin 100-200 aminoacizi şi sunt
responsabile de menţinerea structurii cromozomului. Există 5 tipuri de histone,
notate cu: H1, H2A, H2B, H3 şi H4.
Proteinele nonhistonice sunt reprezentate de unele polipeptide cum sunt:
ADN-polimerazele, ARN-polimerazele, ligazele, helicazele, unele proteine
reglatoare ale transcripţiei, proteinele structurale (actina, tubulina).
La microscopul electronic, cromatina din cromozomi arată ca un şirag de
perle, fiecare perlă reprezentând un nucleozom. Unitatea organizatorică a firului
de cromatină este nucleozomul. Un nucleozom este alcătuit dintr-un octamer de
histone (câte 2 molecule din histonele H2A, H2B, H3 şi H4) şi un segment de ADN
nucleozomic (alcătuit din cca 145 perechi de nucleotide) care înconjoară
octamerul de histone de aproximativ 1,75 ori. În structura nucleozomului intră şi
ADN internucleozomic (alcătuit din cca 55 perechi de nucleotide) care este asociat
cu histona H1.
Acest tip de ADN face legătura între 2 nucleozomi vecini.
Fig. 11 Structura nucleozomului
Fibrele de cromatină din cromozomul metafazic al eucariotelor prezintă
mai multe nivele de organizare: fibra nucleozomală, fibra solenoidală, cromatida.
Fibra nucleozomală are diametrul de 11 nm şi rezultă prin alăturarea
nucleozomilor unul după altul.
Page 35
35
Fibra solenoidală este mai scurtă şi mai groasă, având diametrul de 30 nm
şi apare prin împachetarea fibrei nucleozomale cu ajutorul histonei H1, cu câte 6
nucleozomi pe rând.
Cromatida cromozomului metafazic reprezintă ultimul nivel de organizare
a cromatinei. Ea se obţine prin condensarea (superplierea) fibrei solenoidale.
Această fibră are diametrul de cca 700 nm.
9.3. Organizarea secvenţelor de nucleotide în ADN-ul
cromozomal al eucariotelor
Aproximativ 60% din ADN-ul eucariot este reprezentat de ADN
informaţional (gene), iar restul de 40% din ADN noninformaţional (care nu
codifică aminoacizi). Secvenţele de ADN noninformaţional sunt secvenţe alcătuite
din câteva zeci, sute sau mii de perechi de nucleotide care se repetă.
ADN-ul din genomul eucariot conţine 3 fracţiuni: ADN cu secvenţe unice,
ADN cu secvenţe înalt repetate şi ADN cu secvenţe mediu repetate.
ADN cu secvenţe unice include secvenţele de nucleotide ale genelor
structurale şi secvenţele adiacente ale acestora (promotorul, operatorul,
terminatorul etc). Cromozomul viral şi cel al procariotelor conţine, cu mici
excepţii, numai secvenţe unice (informaţionale). La om, aceste secvenţe reprezintă
cca 50% din genom, iar la Drosophila melanogaster 70%.
ADN cu secvenţe înalt repetate reprezintă 5-45% din genom. Aceste
secvenţe, de obicei sunt foarte scurte şi sunt aranjate în blocuri care se repetă în
tandem. Ele sunt localizate mai ales în regiunea telomerelor şi a centromerului şi
constituie ADN satelit.
În funcţie de numărul perechilor de nucleotide care compun monomerii
din structura ADN înalt repetat putem avea: ADN satelit, cuprinzând monomerii
alcătuiţi din mii perechi de nucleotide; minisateliţii, care au monomerii formaţi
din câteva sute de perechi de nucleotide; microsateliţii, care cuprind 1-6 perechi
de nucleotide repetate sub formă de tandem.
Microsateliţii şi minisateliţii, având un polimorfism accentuat, servesc la
stabilirea amprentei genetice (amprenta ADN), la studiul biodiversităţii speciilor
şi raselor şi la cartarea genică.
Page 36
36
ADN cu secvenţe mediu repetate reprezintă 1-40% din genomul
eucariotelor. Acesta include secvenţe de nucleotide care se repetă de la câteva ori
până la 100.000 ori pe genom. În acest ADN, monomerii nu sunt asociaţi în
tandem, ci sunt dispersaţi în tot genomul.
Genele care controlează sinteza ARNr, ARNt, cele care controlează
sinteza histonelor, sunt alcătuite din secvenţe mediu repetate. Aceste gene se
găsesc într-un număr mare de copii per celulă (500-2000 copii).
Tot din această categorie de ADN fac parte şi elementele genetice mobile
(transpozomii), care sunt secvenţe de nucleotide care se pot mişca în cromozom
dintr-o poziţie în alta şi între cromozomi diferiţi.
Transpozomii pot determina ruperea şi rearanjarea cromozomului,
modificarea expresiei genelor, mutaţii stabile, inactivarea genelor. Ei pot fi
folosiţi ca şi vectori în ingineria genetică.
Teme de control
1. Câţi cromozomi conţine bacteria E. coli?
2. Din ce este alcătuită cromatina care intră în structura cromozomilor la
eucariote?
3. Din ce este alcătuit un nucleozom?
4. Cum rezultă fibra nucleozomală şi ce diametru are?
5. Ce este ADN-ul satelit, de câte feluri este şi la ce poate fi utilizat?
6. ADN-ul cu secvenţe înalt repetate şi mediu repetate intră şi în structura
genelor de la procariote?
CAPITOLUL 10
MUTAŢIILE ŞI MUTAGENEZA
Definiţia şi clasificarea mutaţiilor
Mutaţia reprezintă orice modificare în structura şi funcţia materialului genetic
(cromozomi şi gene) şi ca urmare, orice modificare apărută în fenotip.
O schimbare în structura sau în numărul cromozomilor reprezintă o aberaţie
cromozomială, iar o schimbare în structura chimică a ADN-ului dintr-o genă
reprezintă o mutaţie genică.
Mutaţiile pot fi clasificate după mai multe criterii, în următoarele categorii:
a) după natura materialului genetic implicat în mutaţii, mutaţiile pot fi: mutaţii
genomice (de număr), mutaţii cromozomiale (de structură), mutaţii genice;
Page 37
37
b) după natura celulelor în care au loc mutaţii, mutaţiile pot fi: mutaţii
somatice şi mutaţii germinale sau gametice.
Mutaţiile somatice apar în celulele corpului unui organism şi sunt transmise la
toate celulele care provin din celula mutantă originală. Niciodată mutaţiile somatice
nu se transmit la descendenţi.
Mutaţiile germinale sau gametice apar în celulele germinale ale unui organism
(ovogonii şi spermatogonii). Aceste mutaţii se regăsesc şi în gameţi şi se transmit la
descendenţi.
c) după modul de apariţie, mutaţiile pot fi spontane şi induse sau provocate cu
ajutorul agenţilor mutageni.
Mutaţiile spontane sunt produse de radiaţiile cosmice şi variaţiile bruşte de
temperatură şi apar cu o frecvenţă de 1/100000 – 1/1000000.
d) după direcţia de realizare, pot să existe mutaţii înainte (directe), când gena
normală (A) se transformă în genă mutantă (a) si mutaţii înapoi (inverse), când gena
mutantă se transformă în genă normală
e) după acţiunea genei mutante, mutaţiile pot fi dominante sau recesive.
f) după efectul fenotipic sau gradul de viabilitate,mutaţiile pot fi:dăunătoare,
letale si viabile.
10.1. Mutaţiile genice
Gena este un fragment din macromolecula de ADN şi intră în structura unui
cromozom. Fiecare genă este alcătuită de la 900 - 1500 nucleotide. Numărul şi
secvenţa nucleotidelor dintr-o genă reprezintă informaţia genetică sau mesajul
genetic.
Dacă din diferite cauze se produc modificări în numărul şi secvenţa
nucleotidelor dintr-o genă rezultă o genă mutantă. Aceste modificări se pot realiza
prin înlocuirea, pierderea sau adăugarea unui nucleotid sau grup de nucleotide sau
prin inversarea ordinii nucleotidelor din structura unei gene.
Astfel de modificări care se produc în numărul şi secvenţa nucleotidelor dintr-
o genă poartă numele de mutaţii genice sau punctiforme. Mutaţia la nivelul genei se
manifestă şi prin modificarea funcţiei acesteia. O genă mutantă va determina un
caracter diferit faţă de cel determinat de o genă normală.
Agenţii mutageni şi mecanismul de producere a mutaţiei genice
Page 38
38
Orice agent sau substanţă care prin acţiunea sa asupra materialului genetic
produce o mutaţie se numeşte agent mutagen. Agenţii mutageni pot fi de trei tipuri:
chimici, fizici şi biologici.
Agenţii mutageni chimici sunt reprezentaţi de substanţe chimice care
acţionează asupra genelor (acizilor nucleici), modificându-le structura normală.
Aceşti agenţi produc mutaţii în structura genelor prin următoarele mecanisme:
- substituţie, când o bază azotată este înlocuită cu altă bază.
- deleţie, când se pierde una sau mai multe nucleotide din structura ADN-ului sau
genei;
- adiţie, când se adaugă una sau mai multe nucleotide în structura unei gene;
- inversie, când se inversează ordinea nucleotidelor din structura unei gene.
Substanţele chimice mutagene pot fi de trei feluri:
- substanţe chimice mutagene care acţionează direct asupra ADN-ului în fază stabilă;
- substanţe chimice mutagene care acţionează în momentul replicării ADN-ului
şi a sintezei ARNm;
- substanţe chimice mutagene radiomimetice.
a) Substanţele chimice mutagene care acţionează direct asupra ADN-ului în faza
stabilă sunt reprezentate de acidul azotos, hidroxilamina.
Acidul azotos sau nitros (HNO2) are proprietatea de a dezamina citozina,
adenina şi guanina. Prin dezaminare, citozina este transformată în uracil, adenina în
hipoxantină, iar guanina în xantină. Prin dezaminarea citozinei şi transformarea ei în
uracil, în molecula de ADN, rezultă în loc de perechea CG, perechea de baze AT.
Acidul azotos produce substituţia de baze azotate prin tranziţie. Efectul mutagen al
acidului azotos constă în formarea unei molecule de ADN în care o catenă este
mutantă (A-T) iar cealaltă este normală (G-C).
b) Substanţe chimice mutagene care acţionează în momentul replicării ADN sau
sintezei ARNm – sunt cunoscute sub numele de analogi ai bazelor azotate, ei
având structură asemănătoare bazelor azotate.
Analogii bazelor azotate sunt reprezentaţi de: 5–bromuracil, 5–
bromdeoxiuridina, fluoruracilul, 2- aminopurina.
Dacă în cursul replicării ADN în loc de timină sunt încorporate aceste
substanţe analoage, atunci după câteva replicări ale ADN se vor produce modificări în
secvenţa nucleotidelor. Încorporarea 5–bromuracilului sau 5– bromdeoxiuridinei va
duce la apariţia în catena de ADN a perechii adenină - bromuracil în loc de perechea
Page 39
39
A-T. Bromuracilul se leagă de G. Aceasta are ca efect înlocuirea în noua moleculă de
ADN a perechii A-T cu perechea de baze
G-C.
c) Substanţe chimice mutagene radiomimetice - acţionează asemănător radiaţiilor
ionizante provocând rupturi şi restructurări ale acizilor nucleici.
Din această categorie fac parte substanţele alchilante: dietilsulfatul,
dimetilsulfatul, metilmetansulfonatul, etilmetansulfonatul, iperita. Aceste substanţe
posedă gruparea alchil.
Substanţele alchilante acţionează asupra moleculelor de ADN prin:
- alchilarea radicalului fosforic, ceea ce duce la ruperea legăturilor dintre
dezoxiriboză şi radicalul fosforic cu dezagregarea moleculei de ADN;
- alchilarea bazelor azotate şi transformarea lor în analogi ai acestora, ceea ce are ca
urmare erori de replicare a moleculei de ADN;
- eliminarea bazelor purinice si înlocuirea cu baze pirimidinice (depurinizarea ADN),
ceea ce duce la mutaţii prin substituţie de tipul transversiei.
Agenţi mutageni fizici
Radiogenetica este ramura geneticii, care studiază efectele mutagene şi
teratogene ale radiaţiilor naturale şi ale celor artificiale.
Agenţii mutageni fizici sunt reprezentaţi de două categorii de radiaţii:
ionizante şi neionizante.
Radiaţiile ionizante pot fi clasificate în:
- radiaţii electromagnetice - razele X şi Gamma, a căror lungime de undă este mică
şi au o putere de penetraţie foarte mare, producând mutaţii la toate organismele, prin
fenomenul de ionizare;
- radiaţii corpusculare - razele Alfa, Beta, protonii, neutronii lenţi şi rapizi şi
particule grele, care sunt particule cu energie mare dar cu putere de penetraţie mică.
Razele Alfa, Beta penetrează ţesuturile moi doar o fracţiune de mm sau câţiva mm
(razele Beta). Cantitatea de radiaţie primită de ţesuturile iradiate este numită doză de
iradiere şi se măsoară în razi. Un rad reprezintă cantitatea de energie absorbită de un
gram de ţesut.
Radiaţiile neionizante (ultraviolete) produc mutaţii atunci când acţionează
asupra unor celule aşezate în strat subţire, cum sunt culturile de microorganisme
(bacterii, mucegaiuri, alge, etc.). Cea mai mare absorbţie pentru razele ultraviolete pe
care o prezintă molecula de ADN se realizează la lungimea de undă de 2600 Å.
Page 40
40
10.2. Mutaţiile genomice (de număr)
Mutaţiile genomice (de număr) se pot clasifica în două categorii: euploidia,
care presupune existenţa unui set sau mai multor seturi de cromozomi în celele şi
aneuploidia, care presupune existenţa unuia sau mai multor cromozomi în plus sau în
minus faţă de numărul normal.
Euploidia poate fi de mai mute feluri: monoploidia (în nucleul celulelor
somatice există un singur set de cromozomi), diploidia (indică existenţa a două seturi
de cromozomi -2n), triploidia (3n), tetraploidia (4n), hexaploidia (6n).
Când în celule există mai mult de două seturi de cromozomi este vorba despre
poliploidie. Poliploidia este mai frecvent întâlnită la plante. Proporţia plantelor
poliploide din flora spontană creşte de la ecuator spre poli. Aceasta dovedeşte că
poliploidia este o modalitate de adaptare a plantelor la condiţiile aspre de climă.
Poliploidia este de dorit la plantele furajere, deoarece aceste plante poliploide au
partea vegetativă mai bogată şi în acest fel ele asigură producţii mai mari de furaje la
hectar. La animale, poliploidia este rar întâlnită. Se întâlneşte la organisme ce se
reproduc asexuat.
Poliploidia este de două feluri: autopoliploidia şi allopoliploidia.
Autopoliploidia – presupune existenţa în celulele somatice a unor seturi de
cromozomi identice. Formele autopoliploide rezultă în urma multiplicării garniturii
cromozomiale proprii în timpul diviziunii meiotice, fenomen numit endoreplicare
cromozomială.
Allopoliploidia – presupune existenţa în celulele somatice a unor seturi de
cromozomi care nu sunt identice. Allopoliploizii apar în urma unor încrucişări între
indivizi care aparţin unor specii diferite.
Aneuploidia presupune existenţa unuia sau mai multor cromozomi în plus
sau în minus faţă de starea normală (2n).
Aneuploidia poate fi de două feluri: hipoploidie şi hiperploidie.
Hipoploidii:
- monosomia, lipsa unui cromozom dintr-o pereche de cromozomi omologi (2n–1);
- nulisomia, lipsa unei perechi de cromozomi omologi (2n – 2);
- dubla monosomie, afectează mai multe perechi de cromozomi (2n – 1 – 1).
Hiperploidii:
- trisomia, apariţia unui cromozom în plus, identic cu cromozomii unei perechi
oarecare de omologi (2n + 1);
- dubla trisomie (2n + 1 + 1);
Page 41
41
- tetrasomia, prezenţa unei perechi de cromozomi în plus (2n +2).
Aneuploidia este frecvent întâlnită la plante şi rar la animale. La animale,
cazurile de aneupolidie determină stări anormale, duc la letalitate.
Aneuploidia poate avea trei cauze: nondisjuncţia unor perechi de cromozomi
în timpul meiozei, fenomenul de fuziune - fisiune centromerică şi întârzierea
anafazică a unor cromozomi.
În timpul diviziunii meiotice, dacă are loc fenomenul de nondisjuncţie
cromozomială sau nesegregare la nivelul unei perechi de cromozomi, vor rezulta la
sfârşitul meiozei gameţi care au un cromozom în plus, iar alţii care au un cromozom
în minus. Participarea la fecundare a unor gameţi neechilibraţi genetic conduce la
obţinerea de organisme aneuploide.
Prin fuziunea centromerică a doi cromozomi telocentrici rezultă un cromozom
metacentric (duce la micşorarea numărului de cromozomi din celulă).
Prin fisiunea unui cromozom metacentric vor rezulta doi cromozomi
telocentrici (mărirea numărului de cromozomi din celulă).
Unii cromozomi în timpul anafazei meiozei migrează cu întârziere spre polii
celulei şi nu vor mai intra în nucleul celulei sexuale, având ca efect reducerea
numărului de cromozomi din celulele rezultate în urma fecundaţiei.
10.3. Mutaţiile cromozomiale sau de structură
În timpul diviziunii celulare şi în special în timpul meiozei, anumiţi factori
externi pot duce la fragmentarea cromozomilor. Fragmentele rupte pot să se
realipească de unde s-au rupt şi atunci totul reintră în normal, dar altele pot să se
realpipească greşit, ceea ce duce la modificări ale însuşirilor pe care le determină
genele conţinute în fragmentele respective de cromozomi.
Mutaţiile cromozomiale care afectează structura cromozomilor, sunt: deleţia,
duplicaţia, inversia şi translocaţia.
Deleţia presupune ruperea şi pierderea unui segment intercalat dintr-un
cromozom. Deleţia pot avea loc în meioză şi mitoză, însă când se produce, la nivelul
celulelor germinale au repercursiuni grave şi duc la obţinerea de gameţi neviabili sau
nefuncţionali. Deleţia poate fi heterozigotă, când numai un cromozom din perechea
de omologi pierde fragmentul respectiv şi homozigotă, când ambii cromozomi
omologi pierd aceşlaşi fragment.
Page 42
42
În 1977 s-a constatat în Belgia, că fenomenul numit culard (crupă dublă), pe
care-l manifestă rasa de carne belgiană Blanc Blue Belge se datorează unei deleţii de
11 nucleotide din gena miostatinei (hormon tisular ce mediază reflexele muşchilor).
Duplicaţia presupune ruperea unui fragment dintr-un cromozom şi realipirea
lui la celălalt cromozom omolog. Cromozomul care primeşte fragmentul respectiv va
avea două seturi de gene de acelaşi fel. Deoarece genele de pe un cromozom se află în
doză dublă, însuşirile acestor indivizi se vor dezvolta altfel decât la organismele fără
duplicaţie.
Duplicaţiile pot provoca la indivizii care le posedă, accentuarea puternică a
însuşirilor determinate de genele duplicate. Duplicaţia genelor care determină
rezistenţa la unii dăunători sau la unele boli criptogamice (determinate de ciuperci) la
plante poate duce la obţinerea unor descendenţi deosebit de valoroşi pentru practică.
Inversia presupune ruperea unui fragment dintr-un cromozom şi realpirea lui
la locul de unde s-a rupt după ce a făcut o rotaţie de 180o.
Inversia poate fi homozigotă şi heterozigotă. Inversia homozigotă afectează
ambii cromozomi dintr-o pereche, iar cea heterozigotă doar un cromozom. Inversiile
homozigote nu au urmări grave în mersul meiozei. Din acest motiv, inversiile
homozigote constituie un factor de evoluţie, deoarece prin schimbarea poziţiei
genelor într-un cromozom, se modifică şi funcţia. Inversiile heterozigote care au loc
în cursul meiozei conduc la obţinerea de gameţi neviabili şi de aici la cazuri de
infecunditate.
Translocaţia constă în detaşarea unui fragment de la un cromozom dintr-o
pereche şi alipirea lui la un cromozom dintr-o altă pereche.
Translocaţia poate fi terminală, când segmentul de cromozom translocat se
ataşează la unul din capetele cromozomului neomolog şi intercalată, când
segmentul de cromozom translocat se ataşează la alt cromozom neomolog în zona
mijlocie a braţului.
Translocaţia poate fi nereciprocă, când transferarea unui segment de
cromozom este unidirecţională şi reciprocă, când au loc interschimburi de segmente
cromozomiale între cromozomi neomologi.
Translocaţia reciprocă poate fi homozigotă, dacă sunt afectaţi ambii
cromozomi dintr-o pereche de omologi sau heterozigotă, dacă este afectat numai
unul. În translocaţia homozigotă reciprocă sinapsa cromozomilor în meioză este
normală şi gametogeneza decurge normal. În cazul translocaţiei reciproce
heterozigote de obicei se obţin gameţi neviabili. Gameţii viabili se obţin atunci când
Page 43
43
într-un gamet ajung ambii cromozomi care au suferit translocaţia, iar în celălalt gamet
ajung cromozomii ce nu au suferit translocaţia. Translocaţia heterozigotă are ca efect
semisterilitatea.
Teme de control
1. Ce sunt mutaţiile genice sau punctiforme?
2. Prin ce mecanisme se pot realiza mutaţiile în structura genelor de către agenţii
mutageni chimici?
3. Dezaminarea unei nucleotide duce duce la substituţia de baze azotate sau la
deleţie?
4. Ce rol au analogii bazelor azotate în producerea mutaţiilor?
5. Care este diferenţa dintre poliploidie şi aneuploidie?
6. Care sunt avantajele poliploidiei la plantele furajere?
7. Care sunt cauzele aneuploidiei?
8. Care este diferenţa între mutaţiile genomice (de număr) şi cele cromozomiale
(de structură)?
9. Care sunt principalele efecte negative ale mutaţiilor de structură?
CAPITOLUL 11
NOŢIUNI DE EREDOPATOLOGIE
11.1. Boli ereditare la taurine
Brevignatismul inferior – malformaţie a sistemului osos, care se manifestă
prin scurtarea mandibulei, caracterul fiind letal. Este determinat de o genă
recesivă heterozomală (aşezată pe cromozomul X).
Brevignatismul superior – se manifestă prin scurtarea maxilarului
superior, viţeii au cap de Buldog sau ştiucă. Este determinat de o genă recesivă,
este o anomalie neletală.
Cataractul congenital – anomalie neletală, care se transmite recesiv, este
caracterizat prin opacitatea cristalinului, defect care se constată la naştere sau
după primele luni de viaţă.
Page 44
44
Hidrocefalia – anomalie letală, care se transmite recesiv, constă în
acumularea de lichid în cutia craniană, fapt care afectează ţesuturile cerebrale,
provocând în special atrofia creierului.
Achondroplazia – anomalie a scheletului, se poate prezenta sub mai multe
forme. O astfel de anomalie, la care moartea survine în lunile 6–8 de gestaţie,
urmată de expluzarea imediată a fătului, se caractetzează prin scurtarea coloanei
vertebrale, hernie inghinală, fruntea rotundă şi umflată, capul de buldog, vălul
palatin despicat şi membre scurte. Această stare anormală se transmite parţial
dominant, astfel de cazuri fiind întâlnite la rasele Jersey, Hereford şi Friză
britanică. În altă formă de achondroplazie, viţeii se nasc, însă mor la câteva zile
după naştere, datorită dificultăţilor de respiraţie. Anomalia este determinată de
gene letale recesive.
Hidropizia – constă în acumularea de lichid în cavitatea abdominală,
toracică şi subcutanat. Viţeii se nasc morţi sau mor imediat după naştere. Este o
anomalie letală, care se transmite recesiv, fiind determinată de o genă
autosomală.
Herniile – constau în deplasarea totală sau parţială a unui organ în afara
învelişului său normal sau a cavităţii în care se află în mod fiziologic. Sunt hernii
cervicale, inghinale, ombilicale, diafragmatice. La taurine se semnalează mai
frecvent hernia ombilicală, care apare la 8–10 zile după fătare şi este determinată
de gene recesive.
Hipotricoza – se manifestă prin lipsa părului de pe anumite regiuni
corporale sau de pe întregul corp al animalului. Este determinată de gene
recesive.
Polimastia şi politelia – constă în prezenţa uneia sau mai multor glande
mamare sau mameloane, frecvenţa este de 15 – 50%, tipul de transmitere fiind cel
dominant.
Epilepsia – este o boală ereditară a sistemului nervos. Se pare că este
determinată de o genă autosomală cu ereditate dominantă.
Nanismul – se manifestă sub forma unei creşteri reduse a tuturor părţilor
corpului, datorită unui defect de proliferare al celulelor la nivelul cartilajului de
creştere. Este determinat de o genă recesivă.
Polidactilismul – se manifestă prin apariţia unor ongloane supranumerare.
Anomalia este dată de o genă dominantă.
Page 45
45
Anchiloza – se caracterizează prin curbarea membrelor anterioare, datorită
sudării oaselor la nivelul articulaţiilor. Este o anomalie ereditară care se transmite
recesiv.
Paralizia – afectează în special membrele posterioare. Viţelul moare
imediat după naştere. Se transmite recesiv.
Sindactilia – se manifestă prin prezenţa unui singur onglon, de regulă la
membrele posterioare. Anomalia este determinată de o genă recesivă.
11.2. Boli ereditare la porcine
Atresia anală – lipsa orificiului anal. Moartea survine în câteva zile de la
naştere, întrucât animalele nu pot elimina fecalele. După unele date sa pare că
această anomalie este provocată de două perechi de gene alelice, cu interacţiune
epistatică.
Herniile – ombilicale şi ingvino-scrotale, apar evident imediat după fătare
sau până cel mult la 30 zile şi afectează mai mult partea stângă. Unii cercetători
consideră că sunt determinate de doi factori recesivi, iar herniile ombilicale se
datoresc tipului de transmitere dominantă.
Polidactilia – sunt afectate cel mai des membrele anterioare. Transmiterea
este dominantă.
Sindactilia – este o anomalie ereditară cu transmitere dominantă. Se
caracterizează prin prezenţa unui singur onglon cu poziţie verticală.
Membrele anterioare îngroşate – se datoresc unei infiltraţii gelatinoase de
ţesut conjunctiv în locul ţesutului muscular. Este o anomalie letală şi se transmite
recesiv.
Amelia – este o anomalie caracterizată prin absenţa tuturor membrelor. Se
transmite recesiv.
11.3. Boli ereditare la păsări
Ataxia puilor de găină – apare frecvent şi constituie o consecinţă a
tulburărilor funcţionale ale cerebelului. Se transmite recesiv şi este letală.
Alopecia – se caracterizează prin absenţa totală a penelor sau prin
prezenţa unor pene rudimentare pe cap, gât şi vârful cozii. Este o anomalie
semiletală recesivă.
Polidactilia – anomalie frecventă la păsări, se caracterizează prin prezenţa
unor falange în plus şi este determinată de factori letali recesivi.
Page 46
46
Malformaţii ale ciocului – ciocul de papagal, alungirea sau curbarea în
sus a ciocului.
Nanismul – este produs de o genă recesivă (dw) Z-lincată. Nomalia apare
evidentă la vârsta de 8-10 săptîmâni şi devine pregnantă la vârsta de 5 luni când
greutatea masculilor (ZdwZdw) este mai mică cu 40% şi a femelelor (ZdwW) cu
30% faţă de păsările normale de aceeaşi vârstă.
CAPITOLUL 12
INGINERIA GENETICĂ ŞI
APLICAŢIILE SALE
Definiţie: Ingineria genetică este un ansamblu de operaţii şi tehnologii
efectuate in vitro cu gene, cromozomi şi uneori cu celule întregi , în scopul
obţinerii unor organisme cu proprietăţii ereditare premeditate sau prestabilite.
Tehnologiile ingineriei genetice sunt:
1. Tehnologia ADN recombinat
2. Hibridarea şi cibridarea celulară
12.1. Tehnologia ADN recombinant presupune introducerea şi
funcţionarea unei gene numite pasager, într-o celulă sau organism numit
receptor, care suferă un proces de transformare. Pentru ca o genă străină
(pasagerul) să poată pătrunde într-o celulă receptoare şi să nu fie degradată
enzimatic, ea trebuie introdusă într-un vehicul adecvat (plasmid, virus), rezultând
o moleculă de ADN recombinat.
Pentru obţinerea pasagerului ce conţine informaţia genetică necesară
sintezei unei molecule specifice, se pot utiliza următoarele metode:
a) izolarea pasagerului cu ajutorul enzimelor de restricţie din celulele
specializate în care pasagerul este funcţional (gena insulinei umane poate fi
izolată din celulele pancreatice). Această metodă presupune extragerea ADN,
Page 47
47
scindarea ADN izolat cu ajutorul enzimelor de restricţie, identificarea, izolarea şi
pregătirea pasagerului pentru a fi inclus în vehicul.
b) metoda enzimatică de obţinere a pasagerului – în acest caz pasagerul sau
gena dorită se sintetizeată pornind de la ARNm care a transcris informaţia
genetică a genei respective. Transcripţia inversă se face cu ajutorul unei enzime,
revers transcriptază şi se obţine la început o moleculă de ADN complementar
monocatenar (ADNc). Cu ajutorul ADN polimerazei se sintetizează cea de-a
doua catenă a ADN, rezultând un ADNc dublu catenar (gena).
Prin această metodă a fost sintetizată gena insulinei umane şi gena
ovalbuminei de găină, folosind matriţe de ARNm extrase din celulele pancreatice
şi respectiv din celulele zonei albuminifere a oviductului de pasăre.
c) sinteza chimică a pasagerului se face cunoscând compoziţia şi secvenţa
aminoacizilor, ce alcătuiesc o proteină, un hormon sau o enzimă. În acest fel a
fost sintetizată gena somatostatinei umane. Somatostatina este alcătuită dintr-o
secvenţă de 14 aminoacizi. Gena somatostatinei umane conţine 42 nucleotide (14
x 3). Prin această metodă a fost sintetizată şi gena insulinei umane. Metode de
sinteză chimică: metoda fosfodiesterilor, metoda fosfotriesterilor, sinteza
chimică în fază solidă.
Ca vectori sau vehicule pentru transferul genelor, pot fi utilizate
plasmidele bacteriene sau anumite virusuri (fagi sau bacteriofagi).
Utilizând plasmide ca vectori pentru transferul genelor, s-a reuşit
transferul unor gene de origine umană sau animală în celulele bacteriene de
Escherichia coli, care devin capabile să sintetizeze proteine de origine umană sau
animală: insulina umană, somatostatina, hormonul de creştere, interferonul,
ovalbumina.
Prin utilizarea ca vectori a virusurilor bacteriene (bacteriofagi), s-a reuşit
transferul unor gene de la bacterii în celulele umane. Gena care permite
metabolizarea galactozei a fost transferată de la Escherichia coli în celulele
umane, provenite de la un bolnav ce prezenta galactozemie.
În tehnologia ADN recombinant se folosesc mai multe tipuri de enzime,
cum sunt:
- enzime de restricţie, care au rolul de a tăia moleculele de ADN ale vectorului şi
ale pasagerului la anumite situsuri sau locuri de restricţie.;
- ADN ligazele, care au rolul de a lega sau suda fragmentele de ADN ale
vectorului şi pasagerului;
Page 48
48
- revers transcriptaza, care copiază informaţia genetică din ARNm în ADN;
- terminal transferazele, care au rol în formarea unor cozi poli A la capetele
vehicolului şi pasagerului.
Aplicaţiile tehnologiei ADN recombinant
- în obţinerea animalelor transgenice;
- în mutageneza in vitro;
- în modificarea genetică a microorganismelor industriale prin transferul de
gene sintetizatoare de antibiotice de la tulpini de streptomicete la tulpini de
E.coli;
- modificarea genetică a unor bacterii care produc proteine biologice active;
- terapia genică.
Obţinerea animalelor transgenice
Prin animal transgenic se înţelege un animal modificat genetic (OMG),
respectiv un organism căruia i s-a transferat o genă străină.
Tehnologia de obţinere a animalelor şi plantelor transgenice poartă numele
de transgeneză. Transgeneza presupune nu numai transferul unor gene, ci şi
înlocuirea sau inactivarea unor gene.
Metode de transfer a genelor în vederea obţinerii organismelor transgenice
1. Transferul de gene în gonade.
2. Transferul de gene în gameţi.
3. Microinjecţia genelor în embrioni (pronucleul spermatizoidului) – a dat
cele mai bune rezultate.
4. Metode de introducere mecanică a ADN-ului în embrioni: metode
biolistice şi electroporarea.
5. Transferul genelor prin intermediul celulelor embrionare.
6. Transferul genelor cu ajutorul vectorilor retrovirali.
Aplicaţiile animalelor transgenice
1. Aplicaţii în cercetarea fundamentală
pentru studiul modului de funcţionare a genelor transferate şi a mecanismului
de reglaj a funcţiilor biologice.
crearea de modele animale utilizate pentru studiul unor boli genetice la om:
hemofilia, ateroscleroza. Astfel, s-au obţinut câini şi ovine transgenice cu
hemofilie, iepuri, porcine cu ateroscleroză.
2. Aplicaţii practice
producerea de proteine recombinate în laptele animalelor transgenice.
Page 49
49
Această direcţie vizează trei obiective: modificarea componemţilor naturali ai
laptelui, adăugarea de noi componente în lapte, producerea de proteine de interes
farmaceutic în lapte. Dintre proteinele de uz farmaceutic şi veterinar produse în
laptele animalelor transgenice putem aminti:
- hormonul de creştere uman, ce se foloseşte în tratamentul piticismului;
- factorii VIII, IX de coagulare a sângelui, care sunt folositi în tratamentul
hemofiliei A şi B;
- interleukina 2, care este folosită în tratamentul unor boli ale sistemului
imunitar;
- α1 antitripsina, care este medicament pentru tratamentul emfizemului
pulmonar;
- factorul 1 de creştere, asemănător insulinei (IGF1), care este un regulator de
creştere.
modificarea organelor de la porcii transgenici, care să fie destinate grefelor la
om.
terapia genică vizează lupta contra bolilor, prin transferul de gene in vivo în
celulele somatice ale unui animal ce suferă de o anumită boală.
ameliorarea genetică a performanţelor de creştere la animale.
12.2. Hibridarea celulară
Hibridarea celulară presupune fuzionarea unor celule somatice provenite
de la specii diferite, în prezenţa unor agenţi care măresc frecvenţa celulelor care
fuzionează: polietilenglicolul, virusul Sendai inactivat.
Prin hibridare celulară s-a reuşit obţinerea unor hibrizi celulari: om –
ţânţar, celule umane – protoplaşti de morcov, tomate – cartofi.
Celulele hibride nu pot regenera organisme hibride, ci ele pot numai să se
înmulţească şi să formeze clone celulare hibride.
Aplicaţii ale hibridării celulare
- în alcătuirea hărţilor genetice cromozomiale;
- formarea unor celule hibride numite hibridoma, care sintetizează diverse tipuri
de anticorpi monoclonali.
Celulele hibridoma sunt celule hibride între celulele limfocitare, producătoare
de anticorpi şi celulele tumorale sau mielomatoase. Aceste celule au capacitatea
de a creşte pe mediile de cultură.
Page 50
50
13.3 Clonarea animalelor
Este considerată o biotehnologie asociată transferului de embrioni, iar
după unii o tehnică a ingineriei genetice prin care se manipulează genomul la
animale. Clonarea este o metodă foarte frecvent întâlnită la plante şi la
microorganisme ce se înmulţesc.
Clonarea la animale presupune transferul nucleului unei celule somatice
embrionare sau de adult într-o ovocită enucleată. În acest fel un embrion sau un
animal adult poate fi multiplicat într-un număr mare de copii. Toate animalele
rezultate prin clonare sunt identice din punct de vedere genetic. La animale,
clonarea vizează multiplicarea unor animale foarte valoroase într-un număr mare
de exemplare.
Există două metode de clonare:
1. Clonarea embrionară prin transferul unor nuclei diploizi proveniţi din
celule embrionare (blastomere) în ovocite enucleate. Primele rezultate s-au
obţinut la oaie, apoi la taurine, porcine, iepuri.
2. Clonarea animalelor prin transferul de nuclei diploizi proveniţi din celule
de animal adult. Prin această metodă s-a obţinut la sfârşitul anului 1996, oaia
Dolly. A fost obţinută prin transferul unor nuclei diploizi proveniţi din celulele
epiteliale ale glandei mamare în ovocite enucleate. Celulele epiteliului mamar au
fost la început multiplicate pe medii de cultură. Aceste celule au fost oprite în
faza G0 a ciclului celular. Aceasta s-a făcut prin înfometarea celulelor aflate în
cultură, adică s-a redus serul fetal de viţel, determinând celulele să intre în faza
G0. Celulele sunt stimulate prin electrofuziune şi încep să se dividă, rezultând un
embrion care este apoi transferat într-o femelă receptoare.
Page 51
51
TEST 1
1. Care sunt structurile celulare cu rol genetic din nucleul şi citoplasma
celulelor animale?
a) cromozomii, nucleolul, reticulul endoplasmatic;
b) cromozomii, nucleolul, mitocondriile, centrozomul;
c) cromozomii, sucul nuclear, aparatul Golgi, cloroplastele.
2. Cromozomul metafazic conţine:
a) două cromatide, centromer, organizator nucleolar, satelit şi telomere;
b) o cromatidă, centromer, satelit, telomere şi centrozom;
c) o cromatidă, centriol, satelit şi telomere.
3. Caţi autozomi şi câţi cromozomi de sex conţin celulele somatice la om?
a) 44 + X; 44 + Y; 22 + XX; 22 + XY;
b) 44 + XX; 44 + ZZ; 22 + XY;
c) 44 + XX; 44 + XY.
4. Caţi autozomi şi caţi cromozomi de sex (heterozomi) conţin celulele
sexuale (gameţii) de la taur?
a) 58 + XY; 28 + XY; 58 + XX;
b) 29 + X şi 29 + Y;
c) 29 + XY; 29 + XX.
5. Cate celule rezultă în urma mitozei şi caţi cromozomi conţin aceste celule
la om?
a) 4 celule cu 46 cromozomi;
b) 2 celule cu 23 cromozomi;
c) 2 celule cu 46 cromozomi.
6. Cate celule (gameţi) rezultă în urma meiozei II şi caţi cromozomi conţin
acestea la vacă?
a) 4 celule cu cate 30 cromozomi;
b) 2 celule cu cate 60 cromozomi;
c) 2 celule cu cate 30 cromozomi.
Page 52
52
7. Ce structură (monocromatidică sau bicromatidică) au cromozomii rezultaţi
în urma:
- mitozei;
- meiozei I;
- meiozei II;
8. Cum se numesc celulele rezultate în urma meiozei la mascul şi femelă?
a) spermatocit de ordinul I şi ovocit de ordinul I;
b) spermatozoid şi ovulă;
c) spermatogonii şi ovogonii.
9. Care sunt cele două procese care au loc în cursul ciclului celular care
asigură menţinerea constantă a numărului de cromozomi în celulele
somatice?
a) replicarea cromozomilor şi transcripţia;
b) replicarea şi duplicarea cromozomilor;
c) duplicarea cromozomilor şi dansul cromozomilor.
10. Ce gameţi vor produce indivizii cu următoarele genotipuri, ca urmare a
grupării independente a materialului genetic care are loc în meioză?
- AABB; AABb; Aabb; AaBb, AABbCC.
11. Ştiind că gena A determină culoarea roşie a florilor la garoafe iar gena a
determină culoarea albă, precizaţi ce culoare vor avea descendenţii
heterozigoţi (Aa) din generaţia F1 şi F2, dacă acest caracter se transmite
după:
a) dominanţa completă, culoarea va fi ...................................
b) dominanţa incompletă, culoarea va fi.................................
12. Din încrucişarea unei rase de găini cu penajul negru şi creastă bătută
(AABB) cu altă rasă cu penaj alb şi creastă dinţată (aabb), în generaţia F1
rezultă hibrizi negri cu creastă bătută iar din încrucişarea hibrizilor F1 între
ei, în F2 rezultă 16 combinaţii genotipice grupate în 4 clase fenotipice. Ce
caractere prezintă cele 4 clase fenotipice şi în ce proporţii se obţin ele?
A–B– aaB–
A–bb aabb
După ce interacţiune a genelor s-au transmis caracterele?
13. După ce interacţiuni a genelor se transmit grupele sanguine din sistemul
ABO la om şi ce grupe sanguine vor avea copiii rezultaţi din căsătoria
unui bărbat cu grupa sanguină A (LAL
A) cu o femeie cu grupa B (L
Bl).
14. Ce se înţelege prin letalitate determinată de gene letale recesive?
Ce caractere vor avea descendenţii rezultaţi din încrucişarea unor găini şi
cocoşi cu membre scurte (creeper) purtătoare a unei gene letale (Aa+)?
15. În interacţiunea complementară a genelor (la nurcă) cei doi părinţi prezintă
fenotipuri diferite: AAbb (culoare platinată) şi aaBB (culoare aleutină). În
Page 53
53
F2, din interacţiunea genelor nealele (A şi B) şi genelor recesive (a şi b)
rezultă indivizi cu fenotipuri diferite de ale părinţilor: culoarea standard şi
culoarea safir. Ce culori vor avea indivizii din F2?
16. Dacă există o genă recesivă în stare homozigotă (aa) care este epistatică şi
genele cromogene nealele B = negru şi b = roşu, ce fenotipuri (culori) vor
avea indivizii cu următoarele genotipuri:
A – B – 9/16
A – bb 3/16
aaB – 3/16
aabb 1/16
17. Caracterele cantitative sunt determinate în principal de: a) gene epistatice;
b) gene complementare; c) gene aditive (poligene); d) gene letale.
Dacă cele două perechi de gene nealele determină următoarele producţii:
A = 6 kg lapte/zi, a = 4 kg lapte/zi, B = 6 kg lapte/zi şi b = 4 kg lapte/zi, ce
producţii de lapte vor avea vacile cu următoarele genotipuri?
AABB
aaBb
AaBb
aabb
18. În urma unei retroîncrucişări dintre un individ heterozigot (AaBb) şi un
individ homozigot recesiv (aabb) se pot obţine trei rapoarte de segregare
diferite.
Cum sunt aşezate cele două perechi de gene pe cromozomi (pe o pereche
de cromozomi sau pe două perechi diferite) şi cum se transmit la
descendenţi (linkage, crossingover sau segregă independent) dacă:
Rf = 1 : 1 : 1 : 1
Rf = 1 : 1
Rf ≠ 1 : 1 : 1 : 1
19. Caţi gameţi produce femela dublu heterozigotă din F1 dacă nu se produce
crossingoverul (a) şi dacă se produce crossingoverul (b) între genele
nealele plasate pe acelaşi cromozom:
a) A a b) A a
B b B b
20. Definiţi fenomenul de sex-linkage şi precizaţi ce aplicaţii are în avicultură.
Efectuaţi schema de încrucişare pentru obţinerea unui hibrid autosexabil la
găină, ştiind că gena heterozomală L determină îmbrăcarea lentă cu penaj
iar gena l determină îmbrăcarea rapidă cu penaj.
21. Prin replicarea semiconservativă a ADN-ului, care are loc în faza S a
interfazei mitozei, se înţelege că:
Page 54
54
a) dintr-o moleculă de ADN (bicatenară) rezultă două molecule fiice, fiecare
avand o catenă de la molecula mamă, care a funcţionat drept matriţă şi una
nou sintetizată;
b) dintr-o moleculă de ADN rezultă tot o moleculă de ADN, formată din
două catene identice cu ale moleculei mamă.
22. O catenă polinucleotidică de ADN este rezultatul polimerizării mai multor
nucleotide. Care sunt elementele care intră în structura unei nucleotide din
ADN?
23. Transcripţia constă în:
a) decodificarea informaţiei genetice din ARN-ul mesager şi sinteza unei
proteine;
b) copierea informaţiei genetice din ADN în ARN-ul mesager cu ajutorul
enzimei ARN polimeraza.
24. Codul genetic este asemănător unui alfabet cu ajutorul căruia se pot forma
cuvinte de cate trei litere. Ce înţelegeţi prin caracteristica codului genetic
de universalitate:
a) acelaşi aminoacid este codificat de mai mulţi codoni;
b) acelaşi codon codifică acelaşi aminoacid la toate speciile.
25. Cum se numesc secvenţele informaţionale şi cele noninformaţionale din
structura unei gene eucariote?
26. Care este ordinea corectă a intrării în funcţie a enzimelor în procesul
replicării ADN:
a) ADN-polimeraza, ADN-helicazele, primaza, ADN-ligaza;
b) Topoizomeraza, ADN-helicazele, primaza, ADN polimeraza, ADN-ligaza;
c) ARN-polimeraza, ADN-ligaza, primaza, ADN-helicazele.
27. Cum se numeşte la procariote catena de ADN care se sintetizează
discontinuu?
a) catenă conducătoare;
b) catenă progresivă;
c) catenă decalată (întarziată).
28. ADN-polimeraza catalizează:
a) reacţia de transcripţie;
b) reacţia de formare a unor legături fosfodiesterice între gruparea OH – 3’ şi
atomul P - 5’ a doua nucleotide învecinate (polimerizare);
c) reacţia de dezaminare.
29. ADN-ul denaturat se obţine prin:
a) denaturarea termică, urmată de răcirea bruscă a soluţiei în care se află
ADN-ul şi este monocatenar;
b) prin răcirea bruscă a soluţiei în care se află ADN-ul şi este bicatenar;
c) denaturarea termică, urmată de răcirea bruscă şi este bicatenar.
30. Minisateliţii şi microsateliţii sunt:
Page 55
55
a) secvenţe de ADN unice şi servesc la genotipizare;
b) secvenţe înalt repetate şi servesc la întocmirea amprentei genetice;
c) secvenţe mediu repetate şi servesc la studiul mutaţiilor.
TEST 2
1. Legea grupării independente a materialului genetic acţionează în:
a) gametogeneză;
b) fecundare;
c) gametogeneză şi fecundare;
d) nici un răspuns corect.
2. Care sunt cele două procese care au loc în cursul ciclului celular care
asigură menţinerea constantă a numărului de cromozomi din celule de la o
generaţie la alta:
a) duplicaţia cromozomilor în interfaza ciclului celular şi replicarea
cromozomilor în metafaza mitozei;
b) replicarea cromozomilor în interfază şi duplicaţia cromozomilor în
metafază;
c) duplicaţia cromozomilor între două diviziuni succesive şi replicarea
cromozomilor la sfârşitul metafazei.
3. Chiasmele sunt fenomene genetice care sunt implicate în:
a) linkage;
b) crossing-over;
c) duplicaţia cromozomilor;
d) toate răspunsurile sunt corecte.
4. Care este numărul de cromozomi în celulele sexuale şi în cele somatice la
taurine:
a) 30 autosomi şi 1 heterosom şi 58 autosomi şi 2 heterosomi;
b) 29 autosomi şi 1 heterosom şi 58 autosomi şi 2 heterosomi;
c) 28 autosomi şi 2 heterosomi şi 58 autosomi şi 2 heterosomi.
5. La tipul de determinare a sexelor Drosophila, femela este sexul:
a) homogametic;
b) heterogametic;
c) homozigotic;
d) heterozigotic.
6. Câte celule rezultă în urma mitozei şi a meiozei secundare şi care este
numărul de cromozomi din aceste celule:
a) două celule cu 2n şi respectiv două celule cu n cromozomi:
Page 56
56
b) o celulă cu 2n şi respectiv două celule cu n cromozomi;
c) două celule cu 2n şi respectiv patru celule cu n cromozomi;
d) două celule cu 2n şi respectiv două celule cu 2n cromozomi.
7. Arătaţi în ce moment al diviziunii meiotice se realizează crossing-over-ul:
a) diplonem;
b) metafaza I;
c) zigonem;
d) metafaza II.
8. Numărul de cromozomi este caracteristic:
a) fiecărui individ;
b) fiecărei specii;
c) categoriilor înrudite de specii;
d) toate răspunsurile sunt corecte.
9. Genele epistatice acţionează:
a) intensificând manifestarea unei gene;
b) inhibând manifestarea unei gene alele;
c) inhibând manifestarea unei gene nealele;
d) toate răspunsurile sunt corecte.
10. La taurine prezenţa coarnelor este determinată de o genă recesivă, iar
absenţa de cel puţin o genă dominantă (P-). De câte generaţii aveţi nevoie
pentru a ecorna genetic produşii la o turmă de 100 de vaci cu coarne,
folosind un taur fără coarne, dar homozigot?
a) trei generaţii;
b) două generaţii,
c) o generaţie;
d) nu se poate ecorna genetic.
11. Transcripţia reprezintă:
a) sinteza hidraţilor de carbon,
b) sinteza ATP;
c) sinteza ARN
d) sinteza proteinelor.
12. Albinismul:
a) este rezultatul unei mutaţii cromozomiale;
b) este rezultatul unei mutaţii genice;
c) se exprimă în stare heterozigotată;
d) se exprimă numai în stare de homozigoţie dominantă.
13. De recunoaşterea iniţierii transcripţiei la procariote răspunde:
a) factorul β;
b) factorul Ψ;
c) factorul σ;
d) factorul ρ.
14. ARN mesager reprezintă din totalul cantităţii de ARN din celulă:
a) 80 – 85%;
Page 57
57
b) 50 – 55%;
c) 10 – 15%;
d) 2 – 5%.
15. Nucleolul este ataşat de:
a) cromozom la nivelul centromerului;
b) cromozom la nivelul constricţiei secundare;
c) centrosom;
d) ribozomi.
16. Transpozomii reprezintă:
a) elemente genetice mobile;
b) materialul genetic al virusurilor;
c) materialul genetic al unor bacterii;
d) nici un răspuns nu este corect.
17. În procesul de transcriere a informaţiei genetice intervine enzima:
a) ADN-polimeraza;
b) ARN-polimeraza;
c) ADN-transcriptaza;
d) ARN-replicaza.
18. Rezultatul procesului de translaţie a mesajului genetic este:
a) sinteza unei molecule de ARN mesager;
b) sinteza unui lanţ polipeptidic;
c) sinteza unui lanţ de ADN;
d) sinteza unui precursor ribozomal.
19. Codul genetic este degenerat pentru că:
a) un anumit aminoacid este codificat de mai mulţi codoni;
b) nu la toate organismele acelaşi codon codifică acelaşi aminoacid;
c) între codoni nu există suprapunere;
d) totdeauna un codon codifică un singur aminoacid.
20. Reprezentarea grafică a poziţiei genelor în structura unui cromozom
prezintă:
a) idiogramă;
b) hartă cromozomală;
c) cariogramă;
d) cariotip.
Testul 3
1. Catârul şi bardoul rezultă din încrucişarea reciprocă a cabalinelor şi asinilor.
Câţi cromozomi are un astfel de hibrid:
a) 60;
b) 61;
c) 62;
d) 63.
2. Ce este genomul:
Page 58
58
a) totalitatea genelor dintr-un organism;
b) totalitatea genelor dintr-o celulă;
c) totalitatea genelor dintr-un gamet;
d) toate răspunsurile sunt corecte.
3. Câţi gameţi rezultă dintr-un spermatocit de ordinul I:
a) 4;
b) 8;
c) 12;
d) 16.
4. În cazul înlănţuirii incomplete a genelor în poziţia cis, dintr-o încrucişare a
găinilor cu pene frizate albe (dublu heterozigoţi), cu cocoşi cu pene normale şi
colorate, dublu homozigoţi, ce combinaţii pot ieşi la descendenţi:
a) indivizi cu pene frizate colorate;
b) indivizi cu pene albe normale;
c) indivizi cu pene frizate colorate şi albe normale;
d) indivizi cu pene frizate albe; normal colorate; frizate colorate; normal albe.
5. La unele rase de câini lipsa cozii este determinată de o genă cu dominanţă
incompletă. O căţea cu coada normală a fătat opt pui din care trei aveau coada
mai scurtă. Ce fel de genotip a avut câinele:
a) homozigot dominant;
b) heterozigot;
c) homozigot recesiv;
d) hemizigot.
6. Care din elementele enumerate mai jos măreşte variabilitatea genetică într-o
populaţie:
a) disjuncţia cromozomilor;
b) selecţia;
c) consangvinizarea;
d) toate răspunsurile sunt corecte.
7. În cazul interacţiunii complementare a genelor, în descendenţă va rezulta:
a) un fenotip nou diferit de al formelor parentale;
b) se va intensifica manifestarea caracterelor parentale;
c) va fi inhibată manifestarea caracterelor parentale;
d) toate răspunsurile sunt corecte.
8. Codul genetic reprezintă:
a) „limbajul” necesar traducerii informaţiei genetice dintr-o genă într-o
proteină;
b) modul de înregistrare a informaţiei genetice;
c) succesiunea nucleotidelor în genom;
d) toate răspunsurile sunt corecte.
9. Teoria cromozomială a eredităţii explică:
a) modul de organizare al genomului;
b) linkage-ul şi crossing-overul;
Page 59
59
c) structura cromozomilor.
10. Duplicarea cromozomului (centromerului) în diviziunea meiotică are loc în:
a) zigonem;
b) metafaza I;
c) telofaza I;
d) metafaza II.
11. La taurine absenţa coarnelor este dată de o genă dominantă P, iar prezenţa
coarnelor de alela sa recesivă p. Culoarea neagră este dată de gena dominantă
B, iar culoarea roşie de alela sa recesivă b. Ce fel de descendenţi şi cu ce
probabilităţi se vor obţine din împerecherea unui taur negru, fără coarne
(dublu heterozigot) cu vaci roşii cu coarne:
a) toţi descendenţii negri şi fără coarne;
b) 25% descendenţi negri şi fără coarne, 25% descendenţi roşii şi fără coarne,
25% descendenţi negri şi cu coarne, 25% descendenţi roşii şi cu coarne;
c) 50% descendenţi negri şi fără coarne, 50% descendenţi roşii şi cu coarne;
d) 50% descendenţi negri şi cu coarne, 50% descendenţi roşii şi fără coarne.
12. Codul genetic este universal pentru că:
a) un anumit aminoacid este codificat de mai mulţi codoni;
b) la toate organismele acelaşi codon codifică acelaşi aminoacid;
c) între codoni nu există suprapunere;
d) are un grad redus de ambiguitate.
13. Câte categorii de genotipuri există într-o populaţie la un locus heterosomal, cu
alelism simplu:
a) 2;
b) 3;
c) 4;
d) 5.
14. Heterosomii reprezintă:
a) cromozomi care se găsesc doar în celulele somatice;
b) o pereche de cromozomi care diferă la cele două sexe;
c) cromozomi care se găsesc doar în celulele gametice;
d) o pereche de cromozomi care se găseşte doar la masculi.
15. Structura primară a unei proteine este dată de:
a) tipurile de aminoacizi implicaţi;
b) numărul lanţurilor polipeptidice;
c) legăturile de sulf şi hidrogen prezente;
d) numărul şi secvenţa aminoacizilor implicaţi.
16. Codonii reprezintă o secvenţă de trei nucleotide din structura:
a) ADN-ului;
b) ARN-ului de transfer;
c) ARN mesager;
d) ARN viral;
e) toate răspunsurile sunt corecte.
Page 60
60
BIBLIOGRAFIE
Obligatorie
1. VLAIC, A., T. OROIAN (2005) – Elemente de genetică pentru zootehnişti. Ed.
AcademicPres Cluj-Napoca;
2. VLAIC, A. (2007) – Genetica peştilor, Ed. Risoprint Cluj-Napoca
3. COŞIER VIORICA, A. VLAIC (2007) – Abordarea practică a problemelor de
genetică animală. Ed. Todesco Cluj-Napoca;
4. VLAIC, A. (1997) – Inginerie genetică. Realizări, speranţe şi nelinişti. Ed.
Promedia Plus Cluj-Napoca;
5. PETRE, A, A. VLAIC (1991) – Genetică animală. Tipo Agronomia Cluj-
Napoca;
6. PETRE, A., E. NEGRUŢIU (1975) – Genetica animală. Ed. Didactică şi
Pedagogică Bucureşti;
7. PETRE, A., A. VLAIC, MARIOARA POP (1989) – Lucrări practice de
genetică animală. Tipo Agronomia Cluj-Napoca;
Facultativă
1. BENCSIK, I. (2005) – Genetica generală. Ed. Mirton, Timişoara;
2. BROWN, T.A. (2002) – Genomes 2 nd
. Ed. Bios Oxford UK;
3. CARLAN, M. (1996) – Elemente de genetică animală normală. Ed. Polirom
Iaşi;
4. COŞIER VIORICA (2007) – Inginerie genetică. Ed. Risoprint Cluj-Napoca;
5. CREANGĂ ŞTF., CÎRLAN, M. (2005) – Ereditate şi variabilitate. Ed. Alfa
Iaşi;
6. DRĂGOTOIU TOMIŢA (2001) – Eredopatologie animală. Ed. Xant
Bucureşti;
7. POPESCU-VIFOR, ŞT., N. PIPERNEA, A. PETRE, I. VINTILĂ (1979) –
Genetica animală. Ed. Didactică şi Pedagogică Bucureşti;
8. POPESCU-VIFOR, ŞT., (1990) – Genetica populaţiilor de animale domestice.
Ed. Ceres Bucureşti.
Site-uri de internet
http://www.contexo.info/DNA_Basics/Mitosis.htm
http://www.bio.miami.edu/dana/mov/mitosis.mov
http://www.bio.miami.edu/dana/mov/meiosis.mov
http://www.cellsalive.com/cell_cycle.htm
http://www.lcusd.net/lchs/mewoldsen/ebiolink.htm
http://www.fli-leibniz.de/~sweta/genetic_code_and_evolution/
http://old.ournet.md/~biochim/ghid/genele/genele.html
http://www.phschool.com/science/biology_place/biocoach/dnarep/helix.html
http://seqcore.brcf.med.umich.edu/doc/educ/dnapr/pg2.html
http://ghr.nlm.nih.gov/BrowseGenes
http://www.biology.arizona.edu/Mendelian_genetics/mendelian_genetics.html