Bolfă Pompei Florin – Rezumat teză de doctorat 1 UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINǍ VETERINARǍ CLUJ NAPOCA – ŞCOALA DOCTORALĂ – FACULTATEA DE MEDICINǍ VETERINARǍ BOLFĂ POMPEI FLORIN R R E E Z Z U U M M A A T T U U L L T T E E Z Z E E I I D D E E D D O O C C T T O O R R A A T T CORELAREA MODIFICĂRILOR IMUNOLOGICE CU STRESUL OXIDATIV ÎN ANEMIA INFECŢIOASĂ ECVINĂ CONDUCǍTOR ŞTIINŢIFIC PROF. DR. MARINA SPÎNU CLUJ-NAPOCA – 2010 –
CUANTIFICAREA INFLUENŢEI INFECŢIEI CU VIRUSUL ANEMIEI INFECŢIOASE
ECVINE ASUPRA CELULELOR IMUNOCOMPETENTE IN VITRO Virusul anemiei infecţioase ecvine (VAIE) este clasificat în subfamilia Lentivirusurilor, din
cadrul familiei Retrovirusurilor pe baza ultrastructurii sale (Gonda şi col., 1978), a reactivităţii serologice încrucişate (Henderson şi col., 1987), a activităţii revers transcriptazei (Archer şi col., 1977), a compoziţiei peptidice (Ishizaki şi col., 1978) şi a organizării genomice (Stephes şi col., 1986). În timp ce structura şi funcţiile VAIE sunt similare altor lentivirusuri, organizarea genomică şi replicarea materialului genetic este mai puţin complexă.
În prezent, multe studii oferă dovezi ştiintifice privind beneficiile terapeutice ale extractelor de plante în tratamentul sau prevenţia a diferite boli, inclusiv unele forme de cancer (Tajima si Aida, 2000, Vickers, 2002). Capacitatea de a modula diferite funcţii imunologice în patologia diverselor boli, precum şi folosirea ca adjuvanţi în prepararea de vaccinuri a fost menţionată pentru unele plante medicinale (Florins si col., 2007, Kaileh si col., 2007).
Numeroase demersuri experimentale efectuate cu extracte vegetale sau principii active izolate din plante au demonstrat că acestea pot modula răspunsul imunitar pe diverse căi: stimulare nespecifică a sistemului imun, stimulare specifică precum şi indirect prin modulare hormonală.
Planta Aloe vera este cunoscută de multe secole pentru proprietăţile sale benefice asupra sănătăţii şi în industria cosmetică. Efectele plantei asupra sistemului imun, au la bază următoarele mecanisme: alprogenul inhibă influxul de calciu în mastocite, inhibând astfel eliberarea mediată antigen-anticorp de histamină şi leucotriene din aceste celule (Ro şi col., 2000). Mai multe componente cu greutate moleculară mică sunt de asemenea capabile să inhibe eliberarea de specii libere de radicali ai oxigenului din neutrofilele umane activate (Hart şi col., 1990).
Planta Calendula officinalis, inclusă în familia Asteraceae, posedă o vastă aplicabilitate în medicină, având aplicaţii în terapia plăgilor, arsurilor, ulcerului gastric şi duodenal, în ulceraţii cutanate (efect cicatrizant), stări inflamatorii. Testări fitofarmacologice cu diferite extracte de Calendula officinalis au indicat şi proprietăţi antivirale, anti-HIV şi anti-genotoxice. Diferite studii clinice au indicat eficienţa crescută a acestui extract în prevenirea dermatitelor acute la pacienţii cu cancer, trataţi prin radioterapie. In vitro au fost observate efecte imonomodulatoare faţă de limfocite umane (Jimenez-Medina şi col., 2006).
De departe unul dintre cei mai utilizaţi agenţi imunostimulatori de origine vegetală este reprezentat de extractul de Echinacea. De secole, acest extract se recomandă în terapia infecţiilor căilor respiratorii anterioare cu etiologie virală. Cel mai probabil, protecţia antiinfecţioasă antivirală este asigurată de proprietăţile de activare sau augmentare a sistemului imunitar – gazdă, stimularea macrofagelor şi sinteza de citokine constituind două mecanisme importante. Cu toate că extractele de Echinacea se folosesc ca imunostimulatoare, dovezile ştiinţifice care să le justifice valorificarea potenţialului terapeutic sunt încă controversate. Datele referitoare la efectele imunomodulatorii induse in vitro de administrarea acestui extract vegetal, subliniază faptul că acestea interesează preponderent elementele structurale ale imunităţii nespecifice (Bukovsky şi col., 1993; Burger şi col., 1997; Rininger şi col., 2000; Vilma Jurkštienė şi col., 2004; Mishima şi col., 2004). Sunt necesare studii ştiinţifice care să elucideze în totalitate aspecte ca: mecanismele de acţiune, biodisponibilitatea, potenţa-puterea relativă sau efectele sinergice dintre componentele bioactive
Există numeroase studii care dovedesc că cele peste 100 de componente chimice din urzică (Urtica dioica) ajută sistemului imun dar şi sistemul articular al organismelor animale (Randall şi col, 1999
Sistemul imun este capabil să recunoscă substanţele străine (antigenii), care determină o stimulare a acestuia şi duce la formarea de anticorpi (imunitatea mediată umoral) sau la un răspuns imun celular (imunitatea mediată celular). În răspunsul imun umoral celulele efectoare sunt limfocitele B transformate în plasmocite responsabile de sinteza anticorpilor. Răspunsul celular implică limfocitele T prin intermediul cărora sunt eliminate celulele infectate cu microorganisme, se realizează supravegherea imunologică (apărarea anticanceroasă) şi reacţia alogrefelor. Limfocitele NK (natural-killer) reprezintă o a treia categorie de limfocite, care nu au nevoie să fie activate pentru a-şi îndeplini funţiile de apărare.
Bolfă Pompei Florin – Rezumat teză de doctorat
3
Scopul lucrării de faţă a fost cuantificarea in vitro, efectelor infecţiei cu VAIE asupra sistemului imun de la cai infectaţi, de diferite vârste şi la diverse perioade de la diagnosticare, comparativ cu răspunsul imun înregistrat la ecvinele libere de boală. Monitorizarea răspunsului imun la cai, s-a efectuat pe mai multe direcţii. Astfel, a fost cuantificat răspunsul imun celular specific (TTB) şi nespecific (fagocitoza), dar şi răspunsul umoral specific (CIC) şi nespecific (Ig totale, capacitatea bactericidă a serului). A fost monitorizat efectul infecţiei cu virusul AIE asupra subsistemului celular de apărare nespecifică dar şi asupra capacităţii funcţionale a subsistemului limfocitar T. Complexele imune circulante, imunoglobinele totale dar şi capacitatea bactericidă a serului de la cai infectaţi cu VAIE şi de la cai sănătoşi, au fost de asemenea monitorizate. S-a dorit astfel realizarea unui tablou cât mai complet în ceea ce priveşte metabolismul celulelor imunocompetente in vitro.
Influenţa infecţiei cu VAIE asupra nivelurilor de imunoglobuline totale din ser s-a testat in
vitro prin intermediul testului de disproteinemie cu sulfat de zinc. Anticorpii faţă de glicoproteinele virale de suprafaţă ale monocitelor şi macrofagelor infectate cu VAIE, sunt detectabili în termen de o lună de la infecţie, dar nu pot media citoliza celulară anticorpo-dependentă (ADCC), indicând faptul că ADCC nu este implicată în controlul VAIE la caii purtători (Tschetter şi col., 1997). Caii purtători inaparenţi prezintă concentraţii crecute de globulină serică totală si de imunoglobuline, şi scazută de albumine, respectiv raport albumine/globuline (Russell şi col., 1998; Valpotic, 2000). La majoritatea cailor este detectabilă o hiperimunoglobulinemie cu o creştere a IgG, IgG(T) si IgM la 60 de zile de la infecţie (McGuire şi col., 1971a).
Prin testul de disproteinemie cu sulfat de zinc, care reprezintă o metodă relativ simplă de observare a dezechilibrului în raportul albumine/globuline, s-a urmărit aprecierea modificărilor intervenite în titrele totale ale anticorpilor antivirali prin estimarea comparativă a concentraţiei imunoglobulinelor totale, fără a detalia funcţionalitatea acestora.
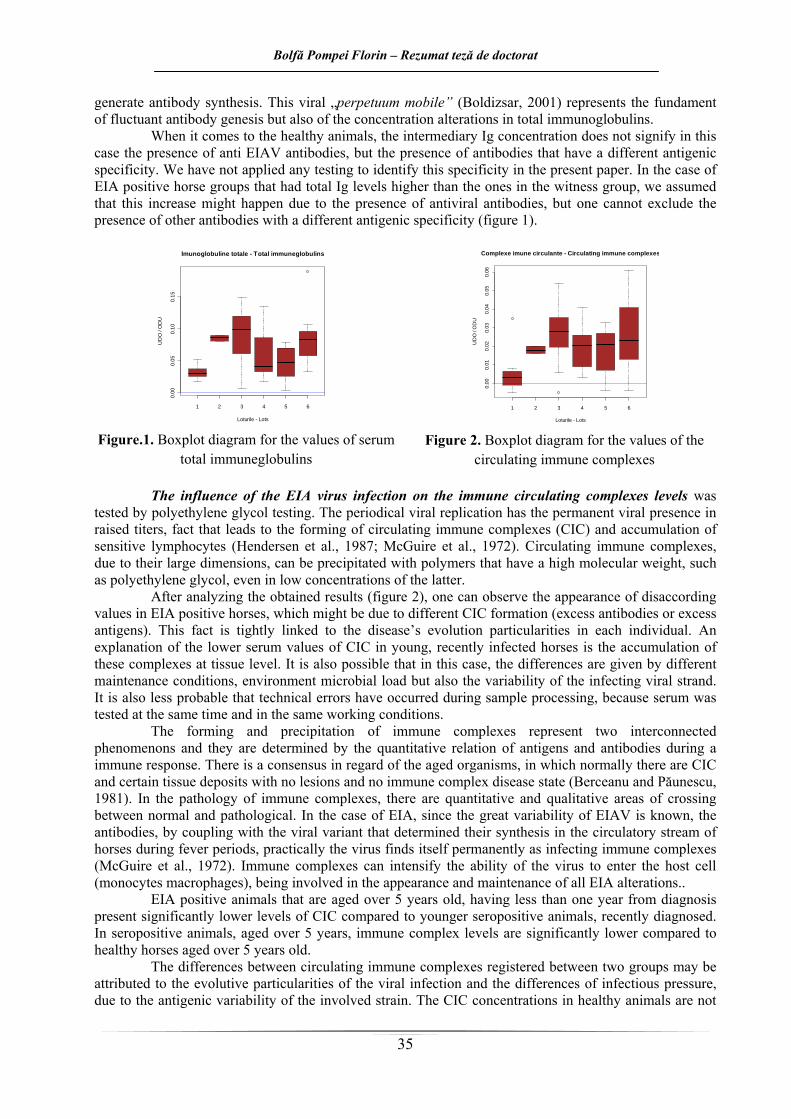
Rezultatele obţinute au arătat că, în cazul cabalinelor cu vârste sub 5 ani, la mai puţin de un de la diagnosticarea infecţiei cu VAIE nivelurile Ig serice snt semnificativ mai scăzute faţă de toţi caii cu vârste de peste 5 ani (sănătoşi si bolnavi). Concentraţia serică a imunoglobulinelor de la această categorie (lot 1) este semnificativ mai redusă faţă de cea a cailor AIE pozitivi de aceleaşi vârste, dar seropozitive de peste 1 an.
Nu au fost găsite diferenţe semnificative din punct de vedere statistic între caii sănătoşi şi cei AIE pozitivi, recent infectaţi, de aceleaşi vârste.
Nivelurile diferite de Ig totale din serurile cailor testaţi pot fi puse pe seama permanentei fluctuaţii a titrulilor de Ig de la caii AIE pozitivi (mai frecvente şi mai evidente în primul an după infecţie), dependente de perioadele de viremie, replicare virală şi selectarea unor noi variante de virus.
În cazul Ig totale, valorile ar trebui să fie concordante cu reactivitatea serologică şi imunoglobulinele să fie prezente în concentraţie maximă la animalele intens pozitive serologic. La momentul recoltării probelor de sânge pentru experimentul de faţă, nu există informaţii despre titrul anticorpilor anti VAIE din sângele cailor cercetaţi. Această informaţie în plus, ar fi permis realizarea unor corelări între rezultatele unui test Coggins şi nivelurile Ig totale al cailor testaţi. Reducerea seropozitivităţii este legată şi de reducerea concentraţiei imunoglobulinelor. Este posibil ca în cazul lotului 1, nivelurile reduse de Ig totale să fie explicate prin scurgerea unui interval redus după infecţie, sau chiar reactivităţii reduse la unele animale. Situaţia se inversează în cazul lotului 3, unde în momentul recoltării, animalele prezntau cele mai crescute titruri serice de Ig totale, aşa cum ar fi de aşteptat în cadrul categoriei de cai AIE pozitivi la mai puţin de un an de la diagnosticare. Valorile crescute în cazul lotului 2 pot fi atribuite numărului scăzut de animale care intră în compoziţia sa (doar doi cai), ceea ce îi reduce valoarea statistică.
Pe perioada de evoluţie a bolii, fluctuaţia titrului imunoglobulinelor ar pute fi explicată şi prin intervenţia unor clase protectoare de Ig, realizarea cuplurilor antigen-anticorp, parte din ele neutralizante iar altele constituind complexele imune circulante infecţioase. Variabilitatea in vivo a virusului favorizează aceste aspecte, ca o consecinţă a formării de variante noi în urma infecţiei primare. VAIE persistă în organism şi în curentul circulator cu perioade de absenţă variabile ca durată, necesare sintezei de noi variante care la rândul lor vor genera sinteză de anticorpi. Acest „perpetuum mobile” (Boldizsar, 2001) viral reprezintă fundamentul anticorpogenezei fluctuante şi totodată a modificării concentraţiei imunoglobulinelor totale.
Bolfă Pompei Florin – Rezumat teză de doctorat
4
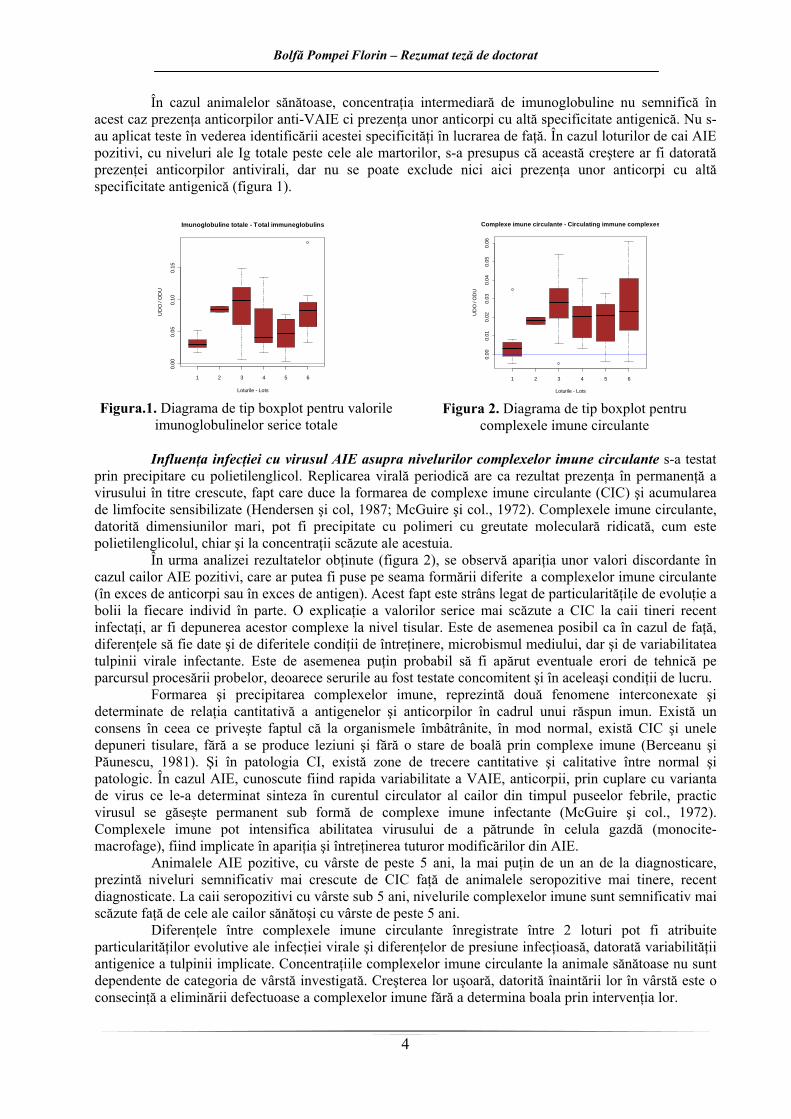
În cazul animalelor sănătoase, concentraţia intermediară de imunoglobuline nu semnifică în acest caz prezenţa anticorpilor anti-VAIE ci prezenţa unor anticorpi cu altă specificitate antigenică. Nu s-au aplicat teste în vederea identificării acestei specificităţi în lucrarea de faţă. În cazul loturilor de cai AIE pozitivi, cu niveluri ale Ig totale peste cele ale martorilor, s-a presupus că această creştere ar fi datorată prezenţei anticorpilor antivirali, dar nu se poate exclude nici aici prezenţa unor anticorpi cu altă specificitate antigenică (figura 1).
complexele imune circulante Influenţa infecţiei cu virusul AIE asupra nivelurilor complexelor imune circulante s-a testat
prin precipitare cu polietilenglicol. Replicarea virală periodică are ca rezultat prezenţa în permanenţă a virusului în titre crescute, fapt care duce la formarea de complexe imune circulante (CIC) şi acumularea de limfocite sensibilizate (Hendersen şi col, 1987; McGuire şi col., 1972). Complexele imune circulante, datorită dimensiunilor mari, pot fi precipitate cu polimeri cu greutate moleculară ridicată, cum este polietilenglicolul, chiar şi la concentraţii scăzute ale acestuia.
În urma analizei rezultatelor obţinute (figura 2), se observă apariţia unor valori discordante în cazul cailor AIE pozitivi, care ar putea fi puse pe seama formării diferite a complexelor imune circulante (în exces de anticorpi sau în exces de antigen). Acest fapt este strâns legat de particularităţile de evoluţie a bolii la fiecare individ în parte. O explicaţie a valorilor serice mai scăzute a CIC la caii tineri recent infectaţi, ar fi depunerea acestor complexe la nivel tisular. Este de asemenea posibil ca în cazul de faţă, diferenţele să fie date şi de diferitele condiţii de întreţinere, microbismul mediului, dar şi de variabilitatea tulpinii virale infectante. Este de asemenea puţin probabil să fi apărut eventuale erori de tehnică pe parcursul procesării probelor, deoarece serurile au fost testate concomitent şi în aceleaşi condiţii de lucru.
Formarea şi precipitarea complexelor imune, reprezintă două fenomene interconexate şi determinate de relaţia cantitativă a antigenelor şi anticorpilor în cadrul unui răspun imun. Există un consens în ceea ce priveşte faptul că la organismele îmbâtrânite, în mod normal, există CIC şi unele depuneri tisulare, fără a se produce leziuni şi fără o stare de boală prin complexe imune (Berceanu şi Păunescu, 1981). Şi în patologia CI, există zone de trecere cantitative şi calitative între normal şi patologic. În cazul AIE, cunoscute fiind rapida variabilitate a VAIE, anticorpii, prin cuplare cu varianta de virus ce le-a determinat sinteza în curentul circulator al cailor din timpul puseelor febrile, practic virusul se găseşte permanent sub formă de complexe imune infectante (McGuire şi col., 1972). Complexele imune pot intensifica abilitatea virusului de a pătrunde în celula gazdă (monocite-macrofage), fiind implicate în apariţia şi întreţinerea tuturor modificărilor din AIE.
Animalele AIE pozitive, cu vârste de peste 5 ani, la mai puţin de un an de la diagnosticare, prezintă niveluri semnificativ mai crescute de CIC faţă de animalele seropozitive mai tinere, recent diagnosticate. La caii seropozitivi cu vârste sub 5 ani, nivelurile complexelor imune sunt semnificativ mai scăzute faţă de cele ale cailor sănătoşi cu vârste de peste 5 ani.
Diferenţele între complexele imune circulante înregistrate între 2 loturi pot fi atribuite particularităţilor evolutive ale infecţiei virale şi diferenţelor de presiune infecţioasă, datorată variabilităţii antigenice a tulpinii implicate. Concentraţiile complexelor imune circulante la animale sănătoase nu sunt dependente de categoria de vârstă investigată. Creşterea lor uşoară, datorită înaintării lor în vârstă este o consecinţă a eliminării defectuoase a complexelor imune fără a determina boala prin intervenţia lor.
Bolfă Pompei Florin – Rezumat teză de doctorat
5
Se poate observa o asemănare destul de mare între rezultatele obţinute în cazul dozării Ig totale şi cele ale dozării CIC (ca şi tendinţe), ceea ce ar confirma faptul că cele două evoluează sincron în sângele animalor bolnave, dar şi sănătoase, şi că depind de vârsta animalelor precum şi de timpul scurs de la diagnosticarea infecţiei.
Umătorul test care a fost aplicat pentru monitorizarea răspunsului imun în cercetarea de faţă, a
fost reprezentat de evaluarea capacităţii bactericide a serului de la cai infectaţi cu VAIE.Acesta reprezintă unul dintre testele care, prin folosirea diluţiilor seriate, determină titrul serului (şi a componentelor antimicrobiene din ser) care prezintă o activitate bactericidă eficientă. Investigarea capacităţii bactericide in vitro permite aprecierea reactivităţii in vivo a organismelor supuse infecţiilor bacteriene.
Experimentele realizate pe alte modele animale de Hansen �i Hirsh au indicat un nivel redus al susceptibilităţii în cazul unor tulpini aviare de Pasteurella multocida. Autorii au corelat rezistenţa totală faţă de seruri cu prezenţa unor factori de patogenitate, adesea fiind puse în evidenţă pentru aceste tulpini structuri de tip capsular (Diallo şi Frost, 2000; Morishita şi col., 1990). Observaţii similare se referă la lipsa de sensibilitate faţă de serurile normale umane în cazul unor tulpini clinice de: Streptococcus pneumoniae, Staphylococcus aureus, Mannheimia haemolytica (Trakulsombon şi col., 1989).
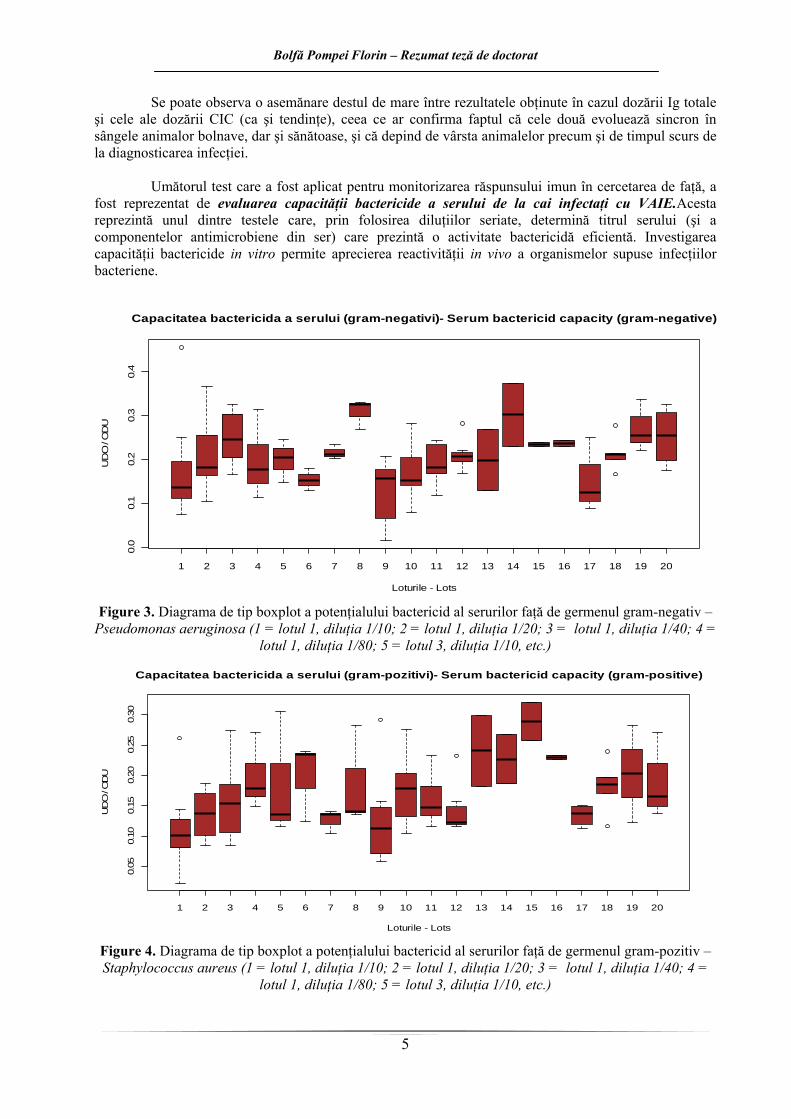
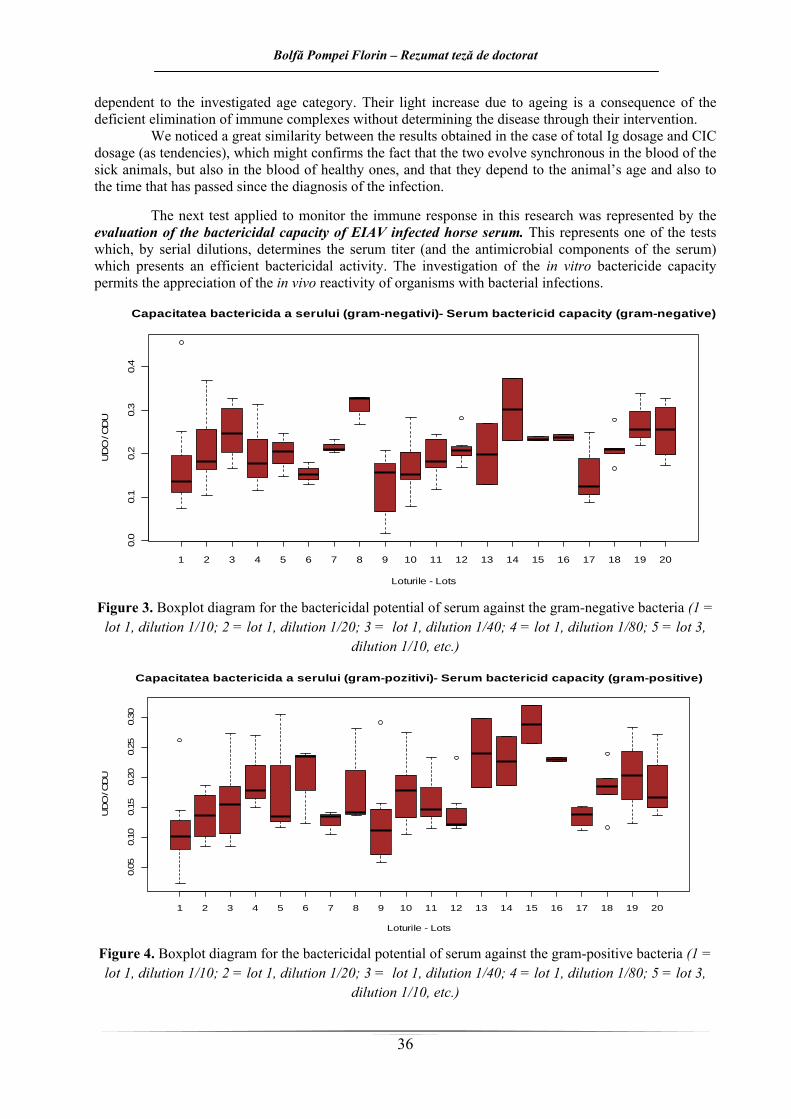
În urma evaluării capacităţii bactericide in vitro a serurilor provenind de la cai sănătoşi şi respectiv cai infectaţi cu VAIE (figura 3 şi 4), se observă o activitate bactericidă atât împotriva unui germen bacterian Gram-pozitiv, cât şi a unuia Gram-negativ. Efectul bactericid maxim a fost obţinut, în majoritatea situaţiilor, la diluţia minimă utilizată.
Activitatea bactericidă a serului contra celor două tulpini din experiment este mai evidentă în cazul serurilor provenind de la cai AIE pozitivi. Efectul este mai evident în cazul testării faţă de germenul Gram-pozitiv. În cazul germenului Gram-pozitiv, serul provenind de la animalele sănătoase din ambele loturi dovedeşte un efect inhibitor mai scăzut asupra dezvoltării bacteriei testate, faţă de serurile provenind de la animalele seropozitive.
In vitro, efectul bactericid minim faţă de ambii germeni testaţi, apare în cazul animalor sănătoase, cu vârste de până la 5 ani, iar cel maxim în cazul cailor tineri recent infectaţi (pentru Pseudomonas aeruginosa), sau în cazul cailor cu vârste de peste 5 ani, infectaţi cu VAIE de mai mult de un an (în cazul lui Staphylococcus aureus).
Influenţa infecţiei cu virusul AIE asupra subsistemului celular de apărare nespecifică a fost
evaluată prin intermediul testului de fagocitoză. Activitatea fagocitară poate fi modificată în sens pozitiv (stimulator) sau negativ (inhibitor) prin unele extracte vegetale.
Rezultatele obţinute înregistreză o tendinţă conform căreia, activitatea fagocitară este mai crecută în cea de a doua perioadă monitorizată, dovendind o activare tardivă in vitro a celulelor cu rol fagocitar din sânge. Doar în cazul cailor tineri, infectaţi recent cu VAIE, capacitatea reactivă a fagocitelor este semnificativ mai crescută într-o primă fază, urmată de o inhibare în cea de a doua etapă (p<0,05).
La caii sănătoşi, extractul alcoolic din Urtica dioica se dovedeşte mai degrabă un inhibitor al fagocitozei într-o primă fază, efectul fiind mai intens la categoria de vârstă 1 – 5 ani (p<0,05), urmat de un efect stimulator în cea de a doua fază.
La caii bolnavi tineri, cu vârste de până la 5 ani, la mai puţin de un an de la depistare, extractul alcoolic de urzică demonstrează un efect stimulator al fagocitozei într-o primă fază, pentru ca mai apoi efectul sa fie unul inhibitor, fapt ce s-ar putea explica printr-o posibilă înglobare rapidă urmată de instituirea unei fragilităţi ridicate la nivel membranal care duce la distrucţii celulare (p<0,05).
În schimb, extractul alcoolic din urzică pare sa aibă un efect inhibitor al fagocitozei într-o primă fază la caii bolnavi cu vârste de peste 5 ani indiferent de intervalul scurs de la diagnosticare şi la cei cu vârste cuprinse între 1 şi 5 ani la mai mult de un an de la diagnosticare, urmat intr-o a doua fază de un efect uşor stimulator.
La caii infectaţi cu VAIE, cu vârste de peste 5 ani, şi la mai mult de un an de la diagnosticare, în cazul loturior netratate şi a celor tratate cu alcool, activitatea fagocitară este semnificativ mai crescută într-o a doua fază (intervalul 45-60 de minute) faţă de primul interval (0-15 minute), ceea ce sugerează faptul că la această categorie de animale capacitatea reactivă a fagocitelor este incă pastrată (p<0,05).
Tratamentul cu extract de urzică in vitro în cuantificarea activităţii fagocitare, urmăreşte un tipic asemănător cu cel al fagocitozei spontane. Urmărindu-se efectele extractului de urzică asupra celulelor izolate de la cabaline sănătoase, pe parcursul primei perioade de citire, se poate spune că acesta acţionează mai degrabă inhibitor. Pe parcursul celei de-a doua perioade, efectul extractului de urzică este unul predominant stimulator.
Cum a fost amintit anterior, într-un alt studiu pe cai cu AIE (Boldizsar, 2001) s-a observat că activitatea fagocitară faţă de particulele de tuş a fost mai puţin intensă la animalele bolnave (AIE pozitive), comparativ cu cele sănătoase, subliniindu-se importanţa intervalului scurs de la depistare/infecţie, la evaluarea activităţii fagocitare. Tratarea cu un extract de Calendula officinalis, demonstrează că activitatea cumulată a etilenglicolului şi a extractului vegetal este inhibitoare la caii sănătoşi. În cadrul studiului respectiv, intervalul pentru care a fost monitorizată activitatea fagocitară in vitro, a fost de 30 de minute.
Bolfă Pompei Florin – Rezumat teză de doctorat
7
În lucrarea de faţă (în principal primul interval monitorizat – respectiv primele 45 de minute de incubaţie), se constată câteva aspecte comune, dar şi unele diferenţe în ceea ce priveşte activitatea fagocitară cu studiul de mai sus. Astfel, la caii bolnavi, activitatea fagocitară scade odată cu înaintarea în vârstă şi cu cât trece mai mult timp de la diagnosticarea infecţiei. În schimb, cu cât trece mai mult timp de la incubarea probelor, activitatea fagocitară pare să se redreseze (pe intervalul 45 – 60 de minute).
Aprecierea efectului infecţiei cu virus al AIE asupra capacităţii funcţionale a subsistemului
limfocitar T s-a efectuat prin aplicarea testului de transformare limfoblastică in vitro, varianta cu sânge integral.
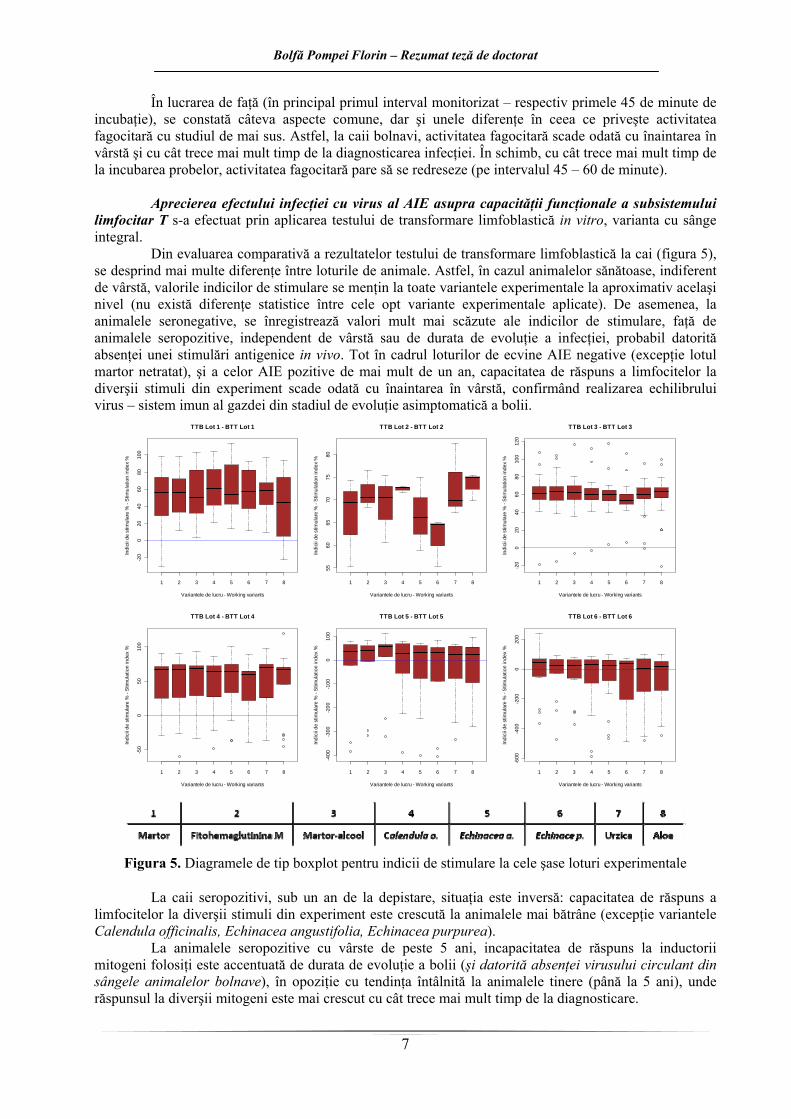
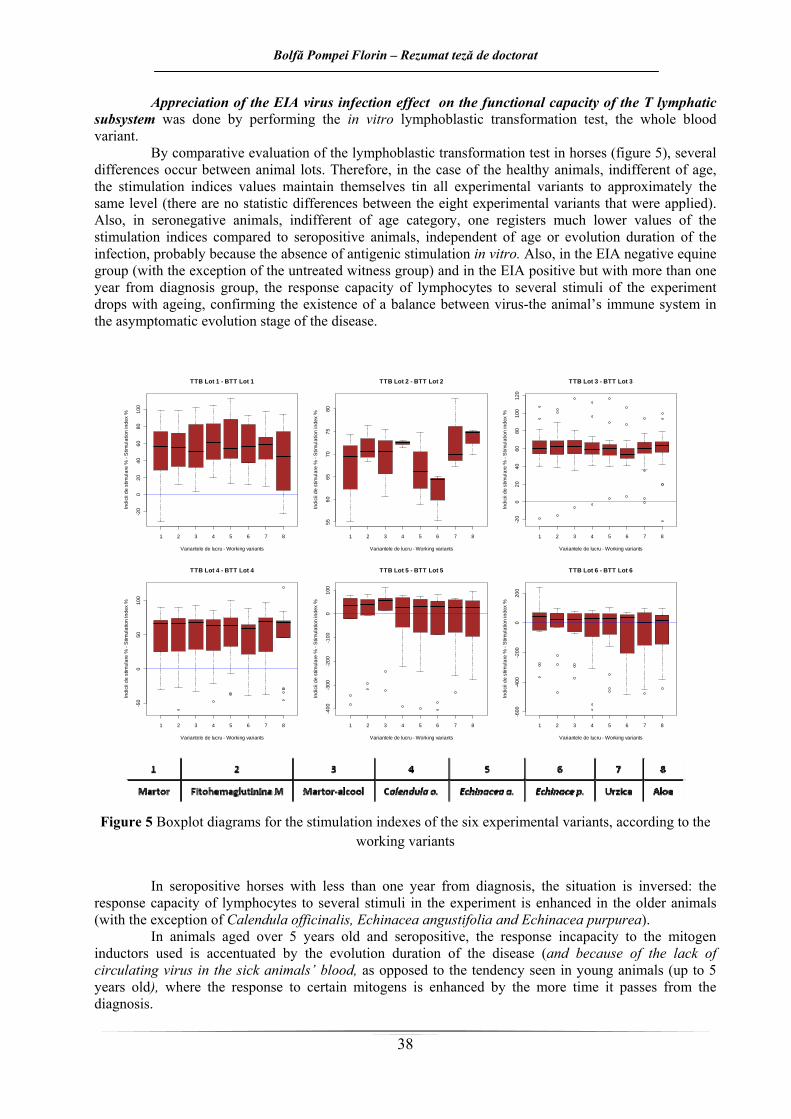
Din evaluarea comparativă a rezultatelor testului de transformare limfoblastică la cai (figura 5), se desprind mai multe diferenţe între loturile de animale. Astfel, în cazul animalelor sănătoase, indiferent de vârstă, valorile indicilor de stimulare se menţin la toate variantele experimentale la aproximativ acelaşi nivel (nu există diferenţe statistice între cele opt variante experimentale aplicate). De asemenea, la animalele seronegative, se înregistrează valori mult mai scăzute ale indicilor de stimulare, faţă de animalele seropozitive, independent de vârstă sau de durata de evoluţie a infecţiei, probabil datorită absenţei unei stimulări antigenice in vivo. Tot în cadrul loturilor de ecvine AIE negative (excepţie lotul martor netratat), şi a celor AIE pozitive de mai mult de un an, capacitatea de răspuns a limfocitelor la diverşii stimuli din experiment scade odată cu înaintarea în vârstă, confirmând realizarea echilibrului virus – sistem imun al gazdei din stadiul de evoluţie asimptomatică a bolii.
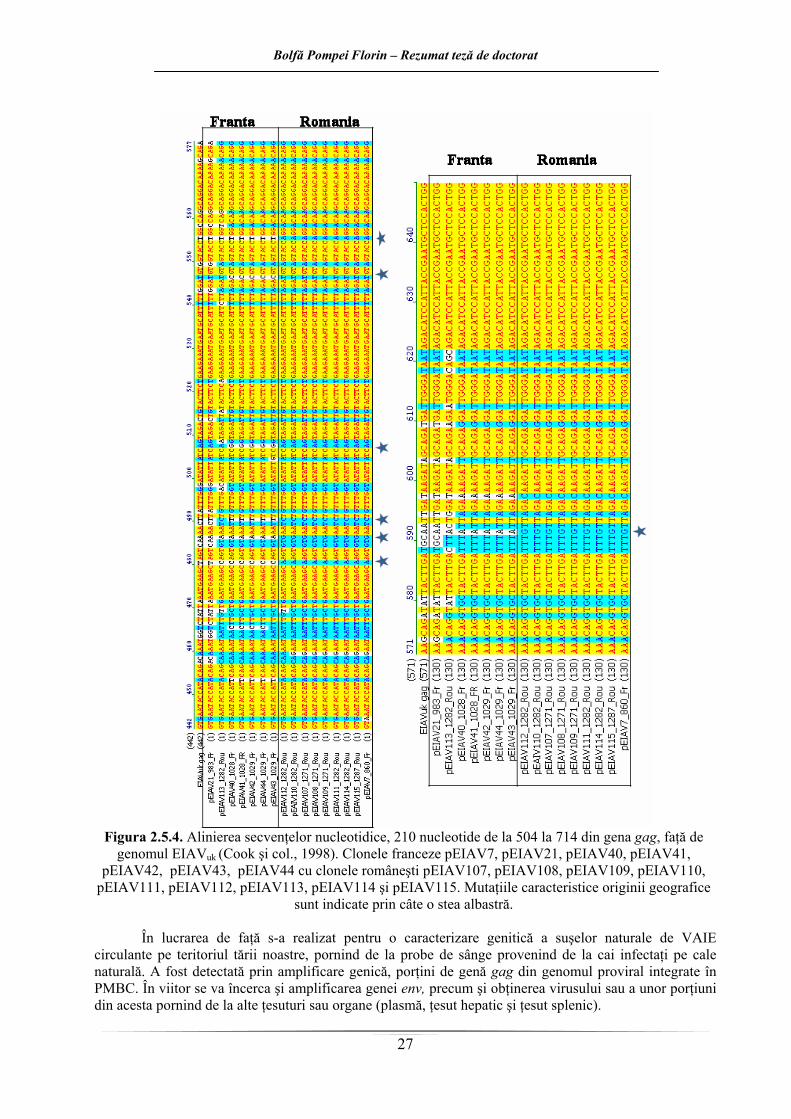
1 2 3 4 5 6 7 8
-20
020
4060
8010
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 1 - BTT Lot 1
1 2 3 4 5 6 7 8
5560
6570
7580
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 2 - BTT Lot 2
1 2 3 4 5 6 7 8
-20
020
4060
8010
012
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 3 - BTT Lot 3
1 2 3 4 5 6 7 8
-50
050
100
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 4 - BTT Lot 4
1 2 3 4 5 6 7 8
-400
-300
-200
-100
010
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 5 - BTT Lot 5
1 2 3 4 5 6 7 8
-600
-400
-200
020
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 6 - BTT Lot 6
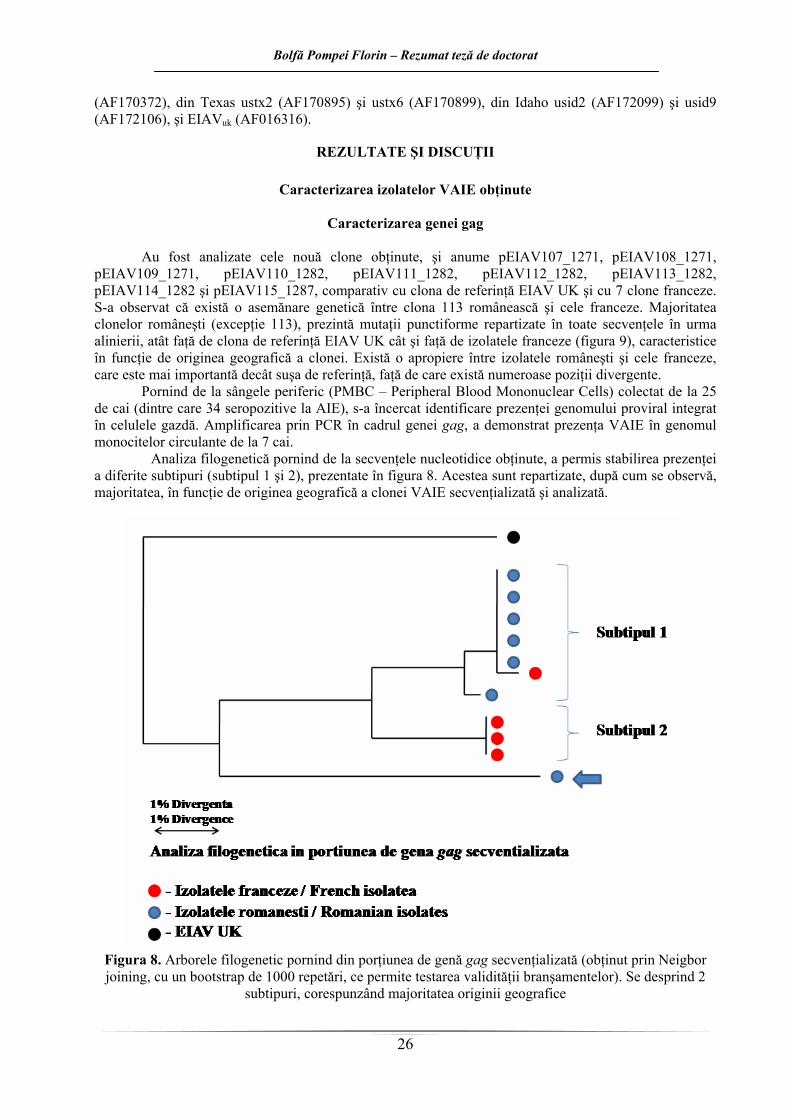
Figura 5. Diagramele de tip boxplot pentru indicii de stimulare la cele şase loturi experimentale
La caii seropozitivi, sub un an de la depistare, situaţia este inversă: capacitatea de răspuns a
limfocitelor la diverşii stimuli din experiment este crescută la animalele mai bătrâne (excepţie variantele Calendula officinalis, Echinacea angustifolia, Echinacea purpurea).
La animalele seropozitive cu vârste de peste 5 ani, incapacitatea de răspuns la inductorii mitogeni folosiţi este accentuată de durata de evoluţie a bolii (şi datorită absenţei virusului circulant din sângele animalelor bolnave), în opoziţie cu tendinţa întâlnită la animalele tinere (până la 5 ani), unde răspunsul la diverşii mitogeni este mai crescut cu cât trece mai mult timp de la diagnosticare.
Bolfă Pompei Florin – Rezumat teză de doctorat
8
Caii cu vârste de până la 5 ani, la mai puţin de un an de la diagnosticarea infecţiei cu VAIE, nu prezintă diferenţe semnificative statistic ale indicilor de stimulare, între cele opt variante de lucru (reactivitatea imună celulară nu este alterată, la fel ca în cazul cailor sănătoşi).
În cadrul cabalinelor cu vârste de până la 5 ani, la mai mult de un an de la diagnosticare, dintre imunomodulatoarele utilizate, Aloe vera (p<0,05) fitohemaglutinina, şi Calendula (0,1>p>0,05) produc o stimuare semnificativ crescută comparativ cu Echinacea purpurea (care are mai degrabă un efect inhibitor).
La lotul de animale cu vârste de peste 5 ani, la mai mult de un an de la diagnosticare, efectul Echinacea purpurea este unul inhibitor, fiind mai scăzut (0,1>p>0,05) faţă de variantele martor, martor alcool şi PHA M.
Efectul inhibitor al extractului de Echinacea purpurea, este vizibil şi în cadrul lotului format din cabaline cu vârste de peste 5 ani, la mai puţin de un an de la diagnosticare (p<0,05) faţă de lotul martor, martor-alcool, PHA M, urzică şi aloe, dar şi faţă de Calendula (0,1>p>0,05).
Efecte imunoredresante ale extractelor folosite sunt înregistrate cu precădere la animalele tinere; astfel de efecte sunt înregistrate la caii cu vârste de până la 5 ani, la mai puţin de un an de la diagnosticare, în urma tratării celulelor cu Calendula officinalis, Echinacea angustifolia şi Echinacea purpurea; la aceeaşi categorie de vârstă, dar la mai mult de un an de la diagnosticare de tratamentele cu Aloe vera, Urtica dioica şi Calendula officinalis; la caii cu vârste mai mari de 5 ani, la mai mult de un an de la diagnosticare, doar extractul de Echinacea angustifolia a înregistrat indici de transformare mai mari faţă de cei doi martori (în toate situaţiile de mai sus, p>0,05).
În cazul celor două specii de Echinacea folosite în experiment, cu toate că este vorba de plante diferite foarte puţin taxonomic, care fac parte din acelaşi gen, efectele acestora in vitro sunt diferite; acest fapt demonstrează prezenţa unor principii activi diferiţi, pentru care organismele testate prezintă într-o măsură mai mică sau mai mare, receptori.
În lucrarea de faţă, rezultatele obţinute de Boldizsar sunt parţial confirmate, dar şi completate, din punct de vedere al reactivităţii imune celulare. Astfel, se confirmă tendinţa de scădere a răspunsului imun celular odată cu înaintarea în vârstă a animalelor. Valorile indicilor de stimulare nu se menţin în aceleaşi limite la animalele bolnave cu cele sănătoase, fiind mult mai scăzuţi la cele din urmă, probabil şi datorită lipsei de stimulare antigenică in vivo. Extractele alcoolice de Echinacea purpurea, Urtica dioica şi Aloe vera dovedesc un foarte slab efect imunostimulator, doar la anumite categorii de cai infectaţi, pe când extractul de Calendula officinalis şi cel de Echinace angustifolia au un efect stimulator ceva mai vizibil. Niciunul dintre extrasele utilizate nu au exprimat un efect imunostimulator/imunomodulator la animalele sănătoase, nedovedindu-se utile în scopul redresării reactivităţii celulare în dozele aplicate, impunându-se eventuale testării ulterioare.
În urma evaluării corelative a rezultatelor de mai sus, se confirmă faptul că, anemia infecţioasă ecvină este o boală sistemică, în care sistemul imun al cailor infectaţi este afectat în diferite grade determinând numeroase implicaţii în patogeneza bolii. Au fost monitorizate componentele sistemului imun specific şi nespecific, celular şi umoral, de la cai sănătoşi şi infectaţi cu VAIE, de vârste diferite şi în diferite faze de evoluţie a bolii. Cumularea rezultatelor obţinute, permite conturarea unui tablou imunopatogenetic, care nu este nici pe departe complet, ce s-ar putea dovedi util medicilor veterinari, dar mai ales cercetătorilor care au ca şi teme de interes protecţia sănătăţii efectivelor de cabaline şi fabricarea unui vaccin eficient antilentiviral.
Bolfă Pompei Florin – Rezumat teză de doctorat
9
CUANTIFICAREA DEZECHILIBRULUI OXIDANŢI/ANTIOXIDANŢI (STRESULUI OXIDATIV) LA CAI INFECTAŢI CU VIRUSUL AIE
De când a început „cercetarea radicalilor liberi”, în 1954, înţelegerea rolului oxidanţilor şi al
antioxidanţilor în stări fiziologice şi patologice a crescut continuu. Oxidanţii sunt în principal generaţi de către enzime metabolice, celule inflamatorii şi pierderi mitocondriale de electroni; ei sunt indispensabili reglării echilibrului redox celular şi pot, în anumite condiţii, să aibă un rol stimulator proinflamator. Antioxidanţii endogeni şi cei exogeni contrabalansează procesele oxidative şi menţin astfel echilibrul oxidanţi/antioxidanţi. Formarea de oxidanţi în exces, sau cantităţi insuficiente de antioxidanţi, poate duce la apariţia stresului oxidativ (Kirskvink şi col., 2008).
La pacienţii cu SIDA a fost observat un deficit în micronutrienţi antioxidanţi. Aceste observaţii privind doar nişte nutrienţi izolaţi demonstrează un defecit de zinc, seleniu şi glutation. O creştere a producţiei de radicali liberi şi a peroxidării lipidelor a fost de asemenea întâlnită la aceşti pacienţi. Acest fapt are o foarte mare importanţă ţinând cont de cercetări recente care prezintă o imunodeficienţă, dar mai important o creştere în replicarea HIV-1 datorită unei superproducţii de radicali liberi. Cea mai semnificativă scădere a fost a carotenoidelor care, în stadiul al II-lea de boală erau la jumătate din valorile normale. S-a pus normal întrebarea dacă aceste scăderi ale antioxidanţilor şi creştere a stresului oxidativ are loc secundar agravării bolii sau, invers, sunt responsabile pentru avansarea ei. În mod paradoxal, peroxidarea lipidelor era la un nivel mai ridicat în stadiul II faţă de stadiul IV de boală. Aceasta poate fi consecutiv unei producţii ridicate de radicali liberi de către mai multe leucocite polimorfonucleate (PMN – neutrofile) în stadiul asimptomatic. Producţia de radicali liberi şi peroxidarea lipidelor par să fie secundare unei inducţii directe de către virus a stimulării neutrofilelor şi a secreţiei de citokine.) (Favier şi col, 1994).
Cercetări efectuate pe viusul HIV al imunodeficienţei umane au demonstrat implicarea speciilor reactive de oxigen. S-a demonstrat că speciile reactive de oxigen şi radicalii liberi sunt implicaţi în activarea virusului HIV latent (Greenspan, 1993).
Până în prezent, stresul oxidativ a fost corelat cu câteva boli ecvine, dar nu cu AIE. În medicina ecvină, numărului studiilor consacrate stresului oxidativ este în creştere şi se crede că stresul oxidativ joacă un rol important în mai multe boli. A fost până în prezent investigată relaţia dintre stresul oxidativ şi boala tractului respirator inferior. S-a demonstrat că ecvinele afectate de această boală suferă de stres oxidativ pulmonar care este corelat cu disfuncţie pulmonară şi inflamaţie a căilor aeriene (Lykkesfeldt şi Svendsen, 2007). Mai mult, există indici de stres oxidativ sistemic detectabil în sângele cailor afectaţi. Există şi studii conform cărora hemoragia pulmonară indusă de exerciţiu şi rabdomioliza indusă de exerciţiu sunt favorizate de antioxidanţi. Se crede că stresul oxidativ este dăunător în leziunea de ischemie-reperfuzie musculară din timpul şi de după anestezie. În prezent este investigat şi rolul stresului oxidativ în boli neuro-motorii (Lykkesfeldt şi Svendsen, 2007; Moffarts şi col., 2005). La caii de exerciţiu, nevoia de oxigen în timpul efortului creşte semnificativ. Majoritatea oxigenului (98%) este redus de lanţul respirator mitocondrial la apă, dar 2% se eliberează ca şi SRO. În timpul exerciţiului, creşte cantitatea de SRO ceea ce implică şi o creştere a cantităţii de antioxidanţi necesari pentru prevenirea stresului oxidativ. Se ajunge astfel la scăderea performanţelor cailor respectivi dar nu se ştie încă exact pe ce căi patogenetice (Moffarts şi col., 2005). Laminita este o altă afecţiune investigată pentru descoperirea unei conexiuni între inflamaţie şi stres oxidativ (Treiber şi col., 2007) dar fără rezultate edificatoare în această direcţie. Sunt necesare studii ulterioare pentru clarificarea acestei situaţii.
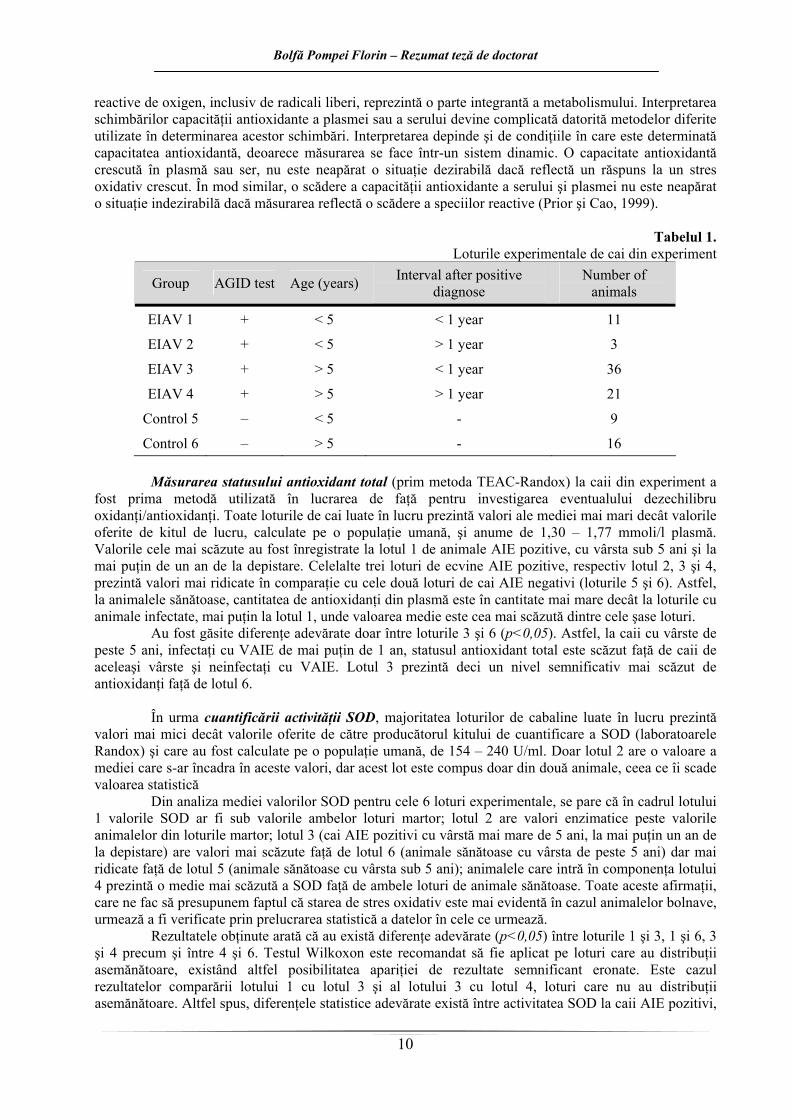
Obiectivul acestui capitol a fost de a cuantifica echilibrul oxidanţi/antioxidanţi (stresul oxidativ)
la cai infectaţi cu VAIE. Astfel ne propunem să realizăm determinarea activităţii a patru sisteme antioxidante enzimatice, a patru sisteme antioxidante neenzimatice precum şi cuantificarea statusului antioxidant total de la cai infectaţi şi neinfectaţi cu VAIE (tabelul 1). Utilizând aceşti indici biochimici dorim să stabilim dacă la cai infecţia cu VAIE duce la apariţia stării de stres oxidativ la nivel celular sau dacă starea de stres oxidativ favorizează infecţia cu virus AIE, sau grăbeşte evoluţia bolii, ducând la o agravare a semnelor clinice şi să evaluăm implicarea stresului oxidativ în modificările care apar în această boală. Abordarea acestui obiectiv se poate dovedi utilă în clarificarea unor aspecte ce ţin de patogeneza bolii, la o mai bună înţelegere a mecanismelor de interacţiune virus – organism şi poate va ajuta în încercările care se fac pentru dezvoltarea unui vaccin eficient anti lentiviral la cai. Producerea de specii
Bolfă Pompei Florin – Rezumat teză de doctorat
10
reactive de oxigen, inclusiv de radicali liberi, reprezintă o parte integrantă a metabolismului. Interpretarea schimbărilor capacităţii antioxidante a plasmei sau a serului devine complicată datorită metodelor diferite utilizate în determinarea acestor schimbări. Interpretarea depinde şi de condiţiile în care este determinată capacitatea antioxidantă, deoarece măsurarea se face într-un sistem dinamic. O capacitate antioxidantă crescută în plasmă sau ser, nu este neapărat o situaţie dezirabilă dacă reflectă un răspuns la un stres oxidativ crescut. În mod similar, o scădere a capacităţii antioxidante a serului şi plasmei nu este neapărat o situaţie indezirabilă dacă măsurarea reflectă o scădere a speciilor reactive (Prior şi Cao, 1999).
Tabelul 1.
Loturile experimentale de cai din experiment
Group AGID test Age (years) Interval after positive diagnose
Number of animals
EIAV 1 + < 5 < 1 year 11
EIAV 2 + < 5 > 1 year 3
EIAV 3 + > 5 < 1 year 36
EIAV 4 + > 5 > 1 year 21
Control 5 – < 5 - 9
Control 6 – > 5 - 16 Măsurarea statusului antioxidant total (prim metoda TEAC-Randox) la caii din experiment a
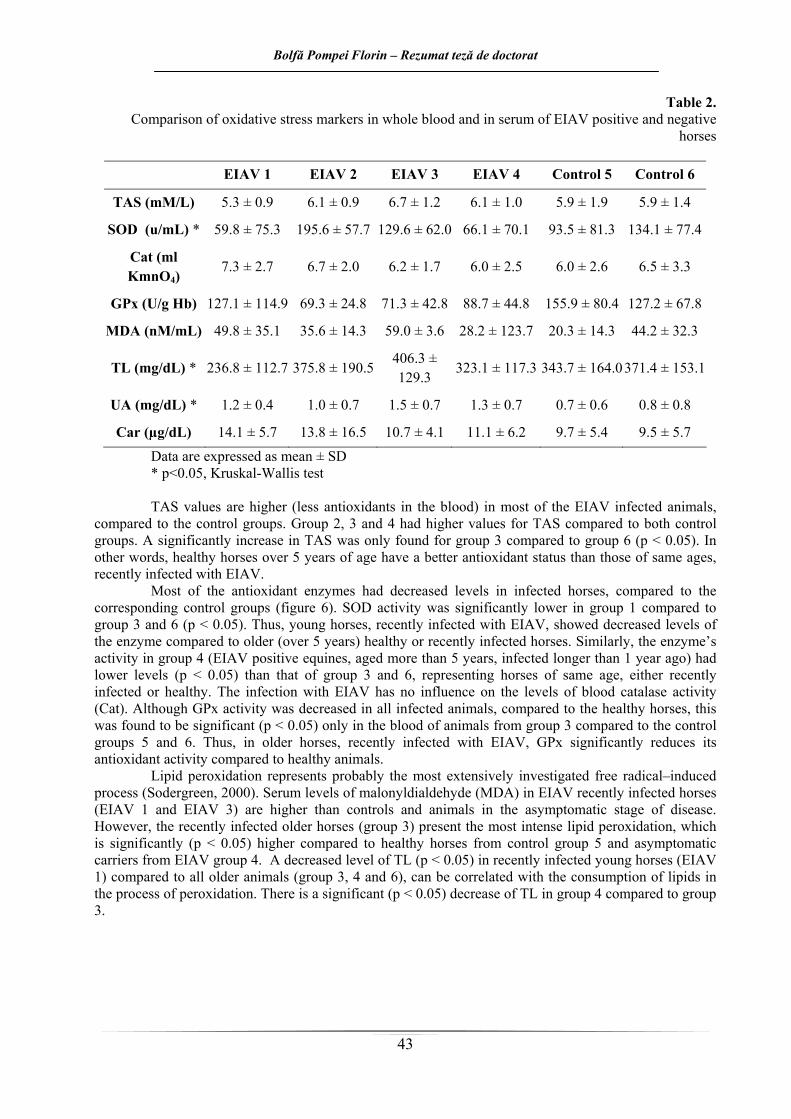
fost prima metodă utilizată în lucrarea de faţă pentru investigarea eventualului dezechilibru oxidanţi/antioxidanţi. Toate loturile de cai luate în lucru prezintă valori ale mediei mai mari decât valorile oferite de kitul de lucru, calculate pe o populaţie umană, şi anume de 1,30 – 1,77 mmoli/l plasmă. Valorile cele mai scăzute au fost înregistrate la lotul 1 de animale AIE pozitive, cu vârsta sub 5 ani şi la mai puţin de un an de la depistare. Celelalte trei loturi de ecvine AIE pozitive, respectiv lotul 2, 3 şi 4, prezintă valori mai ridicate în comparaţie cu cele două loturi de cai AIE negativi (loturile 5 şi 6). Astfel, la animalele sănătoase, cantitatea de antioxidanţi din plasmă este în cantitate mai mare decât la loturile cu animale infectate, mai puţin la lotul 1, unde valoarea medie este cea mai scăzută dintre cele şase loturi.
Au fost găsite diferenţe adevărate doar între loturile 3 şi 6 (p<0,05). Astfel, la caii cu vârste de peste 5 ani, infectaţi cu VAIE de mai puţin de 1 an, statusul antioxidant total este scăzut faţă de caii de aceleaşi vârste şi neinfectaţi cu VAIE. Lotul 3 prezintă deci un nivel semnificativ mai scăzut de antioxidanţi faţă de lotul 6.
În urma cuantificării activităţii SOD, majoritatea loturilor de cabaline luate în lucru prezintă
valori mai mici decât valorile oferite de către producătorul kitului de cuantificare a SOD (laboratoarele Randox) şi care au fost calculate pe o populaţie umană, de 154 – 240 U/ml. Doar lotul 2 are o valoare a mediei care s-ar încadra în aceste valori, dar acest lot este compus doar din două animale, ceea ce îi scade valoarea statistică
Din analiza mediei valorilor SOD pentru cele 6 loturi experimentale, se pare că în cadrul lotului 1 valorile SOD ar fi sub valorile ambelor loturi martor; lotul 2 are valori enzimatice peste valorile animalelor din loturile martor; lotul 3 (cai AIE pozitivi cu vârstă mai mare de 5 ani, la mai puţin un an de la depistare) are valori mai scăzute faţă de lotul 6 (animale sănătoase cu vârsta de peste 5 ani) dar mai ridicate faţă de lotul 5 (animale sănătoase cu vârsta sub 5 ani); animalele care intră în componenţa lotului 4 prezintă o medie mai scăzută a SOD faţă de ambele loturi de animale sănătoase. Toate aceste afirmaţii, care ne fac să presupunem faptul că starea de stres oxidativ este mai evidentă în cazul animalelor bolnave, urmează a fi verificate prin prelucrarea statistică a datelor în cele ce urmează.
Rezultatele obţinute arată că au există diferenţe adevărate (p<0,05) între loturile 1 şi 3, 1 şi 6, 3 şi 4 precum şi între 4 şi 6. Testul Wilkoxon este recomandat să fie aplicat pe loturi care au distribuţii asemănătoare, existând altfel posibilitatea apariţiei de rezultate semnificant eronate. Este cazul rezultatelor comparării lotului 1 cu lotul 3 şi al lotului 3 cu lotul 4, loturi care nu au distribuţii asemănătoare. Altfel spus, diferenţele statistice adevărate există între activitatea SOD la caii AIE pozitivi,
Bolfă Pompei Florin – Rezumat teză de doctorat
11
cu vârsta sub 5 ani, la mai puţin de un an de la depistare şi caii AIE negativi, cu vârste peste 5 ani, activitatea enzimei fiind scăzută la cei bolnavi. De asemenea, caii AIE pozitivi, cu vârste peste 5 ani şi la mai mult de un an de la diagnosticare înregistrează valori mai scăzute faţă de aceeaşi categorie de vârstă de animale sănătoase. Aşadar, la caii sănătoşi cu vârste de peste 5 ani, nivelurile sanguine de superoxid dismutază sunt semnificativ mai crescute faţă de aceeaşi categorie de vârstă de cai infectaţi asimptomatic cu virusul AIE şi de cai sub această vârstă şi la mai puţin de 1 an de la diagnosticare. De asemenea, se pare că odată cu înaintarea în vârstă, activitatea SOD scade la caii infectaţi. Este cazul comparării cailor infectati de peste un an şi cu vârste sub 5 ani cu caii de vârste de peste 5 ani, infectaţi tot de peste un an. Diferenţele în activitatea SOD la aceste categorii, sugerează faptul că odată cu înaintarea în vârstă la caii infectaţi activitatea SOD scade.
Valorile obţinute în cuantificarea activităţii superoxid dismutazei, ne permit să presupunem prezenţa stării de stres oxidativ la caii din loturile 1 şi 4. Se pare astfel că indiferent de intervalul de timp scurs de la instalarea infecţiei cu VAIE, dezechilibrul oxidanţi/antioxidanţi îsi face simţită prezenţa la orice vârstă la cai, prin scăderea activităţii SOD faţă de activitatea enzimatică la animalele sănătoase şi cu vârste de peste 5 ani.
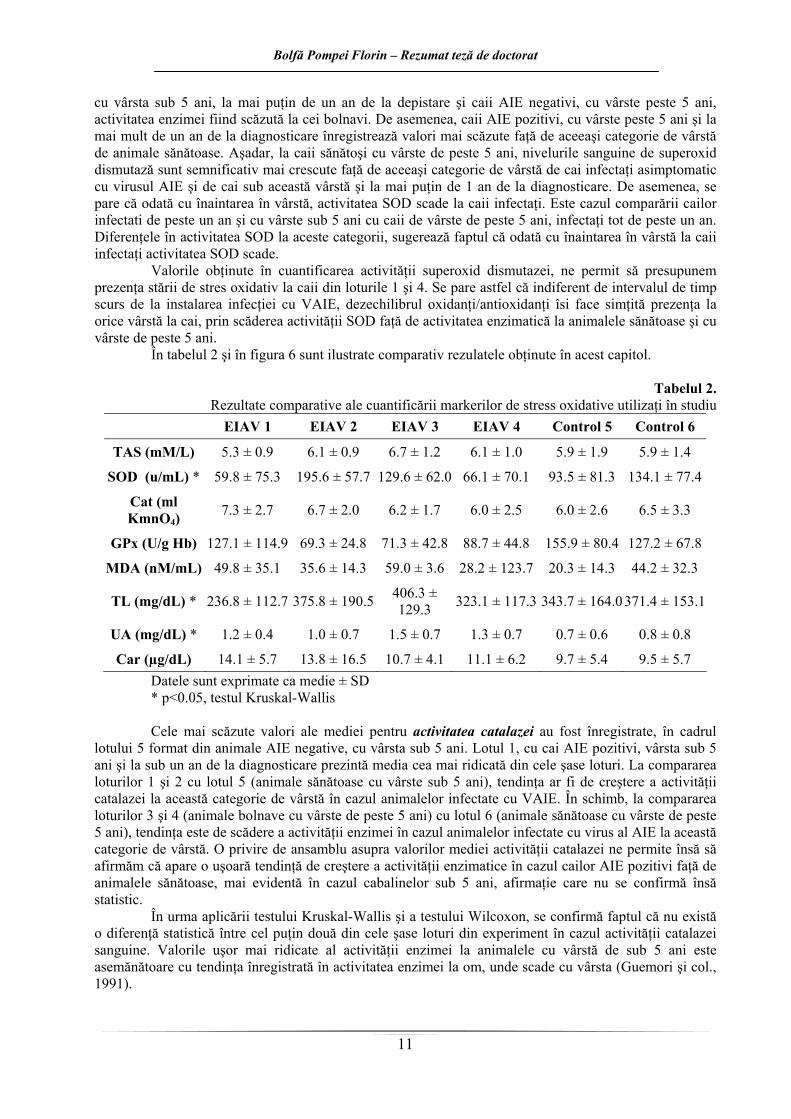
În tabelul 2 şi în figura 6 sunt ilustrate comparativ rezulatele obţinute în acest capitol.
Tabelul 2. Rezultate comparative ale cuantificării markerilor de stress oxidative utilizaţi în studiu
Car (µg/dL) 14.1 ± 5.7 13.8 ± 16.5 10.7 ± 4.1 11.1 ± 6.2 9.7 ± 5.4 9.5 ± 5.7 Datele sunt exprimate ca medie ± SD * p<0.05, testul Kruskal-Wallis Cele mai scăzute valori ale mediei pentru activitatea catalazei au fost înregistrate, în cadrul
lotului 5 format din animale AIE negative, cu vârsta sub 5 ani. Lotul 1, cu cai AIE pozitivi, vârsta sub 5 ani şi la sub un an de la diagnosticare prezintă media cea mai ridicată din cele şase loturi. La compararea loturilor 1 şi 2 cu lotul 5 (animale sănătoase cu vârste sub 5 ani), tendinţa ar fi de creştere a activităţii catalazei la această categorie de vârstă în cazul animalelor infectate cu VAIE. În schimb, la compararea loturilor 3 şi 4 (animale bolnave cu vârste de peste 5 ani) cu lotul 6 (animale sănătoase cu vârste de peste 5 ani), tendinţa este de scădere a activităţii enzimei în cazul animalelor infectate cu virus al AIE la această categorie de vârstă. O privire de ansamblu asupra valorilor mediei activităţii catalazei ne permite însă să afirmăm că apare o uşoară tendinţă de creştere a activităţii enzimatice în cazul cailor AIE pozitivi faţă de animalele sănătoase, mai evidentă în cazul cabalinelor sub 5 ani, afirmaţie care nu se confirmă însă statistic.
În urma aplicării testului Kruskal-Wallis şi a testului Wilcoxon, se confirmă faptul că nu există o diferenţă statistică între cel puţin două din cele şase loturi din experiment în cazul activităţii catalazei sanguine. Valorile uşor mai ridicate al activităţii enzimei la animalele cu vârstă de sub 5 ani este asemănătoare cu tendinţa înregistrată în activitatea enzimei la om, unde scade cu vârsta (Guemori şi col., 1991).
Bolfă Pompei Florin – Rezumat teză de doctorat
12
1 2 3 4 5 6
56
78
910
mM
/LTAS
1 2 3 4 5 6
5015
025
0
u/m
L
SOD
1 2 3 4 5 6
26
1014
ml K
MnO
4
Cat
1 2 3 4 5 6
5010
015
020
0
u/g
Hb
GPx
1 2 3 4 5 6
040
80
nM/m
L
MDA
1 2 3 4 5 6
100
300
500
700
mg/
dL
TL
1 2 3 4 5 6
0.0
1.0
2.0
3.0
mg/
dL
UA
1 2 3 4 5 6
510
1520
ug/d
L
Car
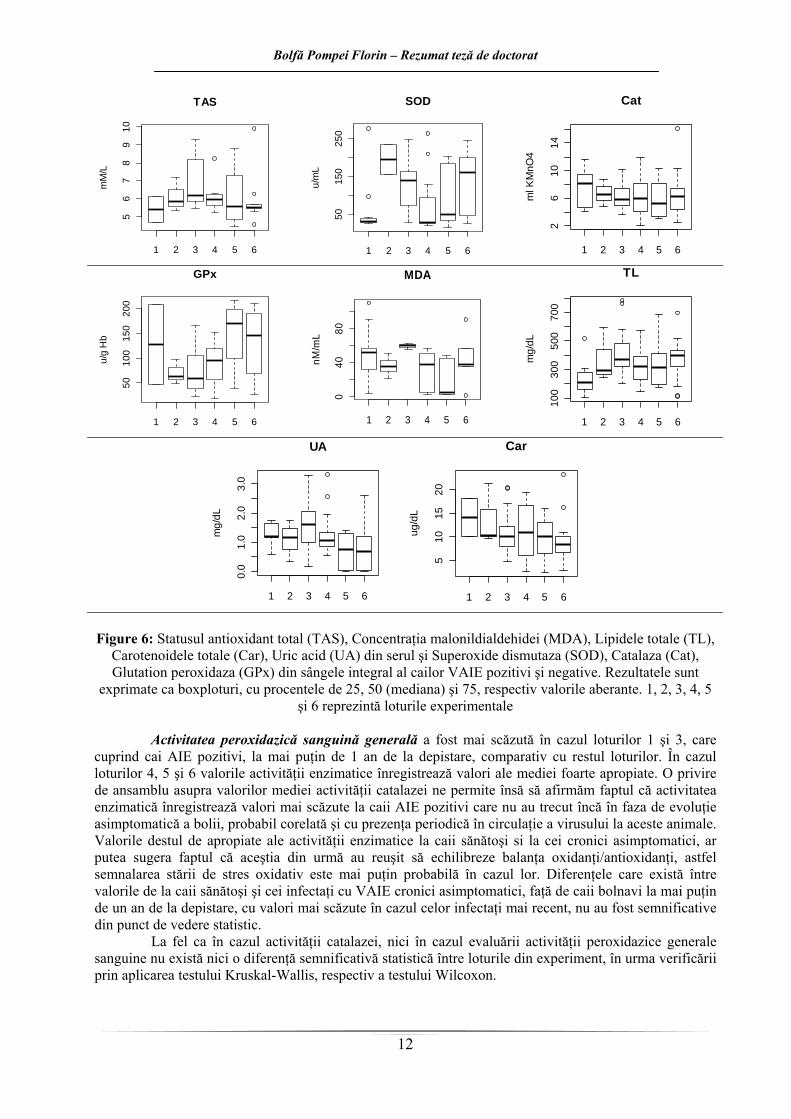
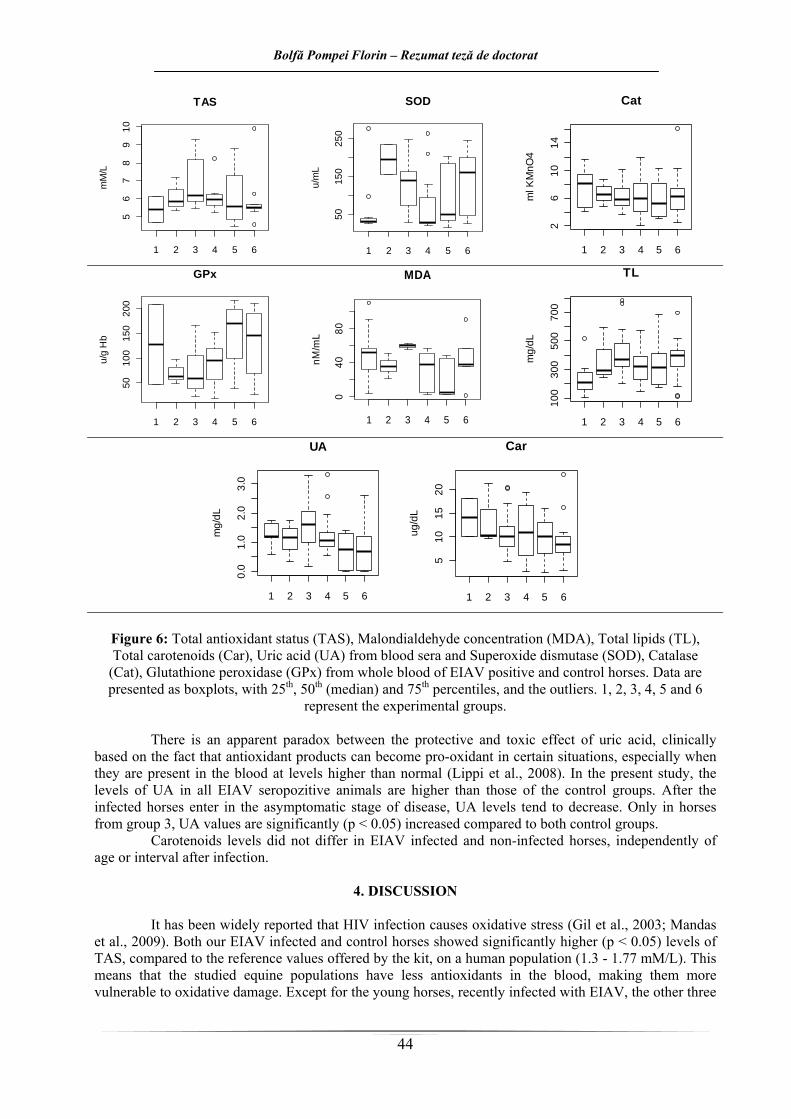
Figure 6: Statusul antioxidant total (TAS), Concentraţia malonildialdehidei (MDA), Lipidele totale (TL),
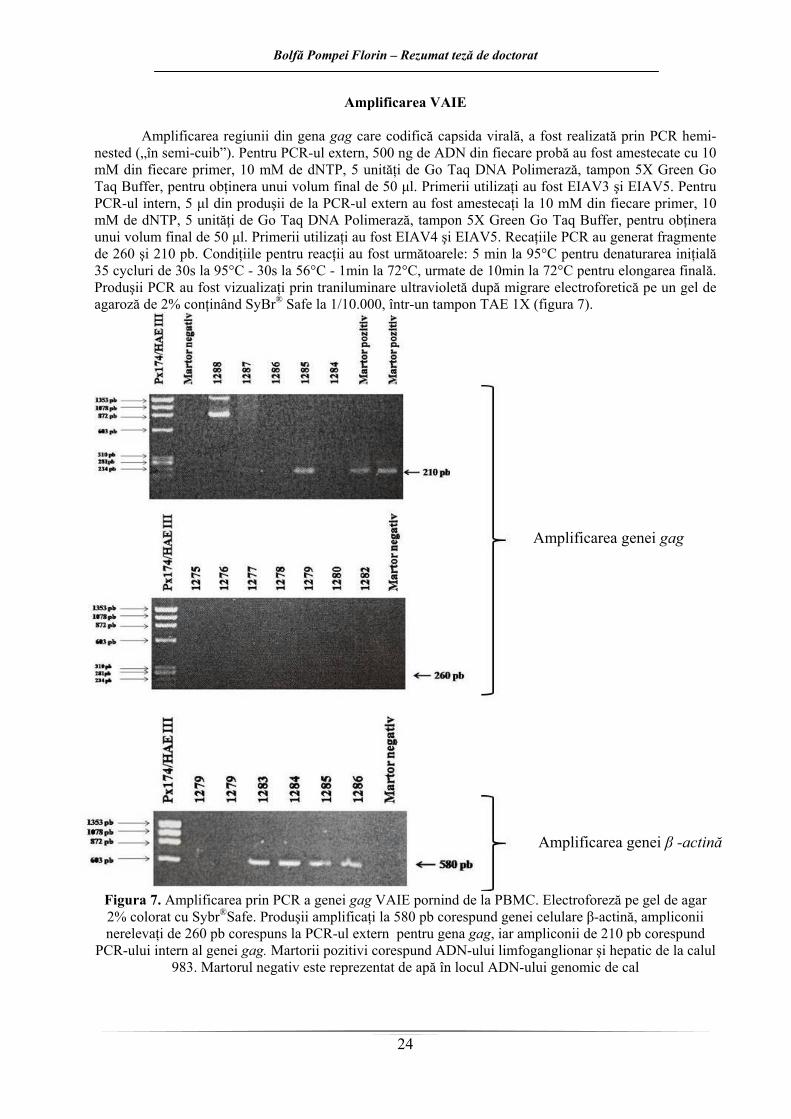
Carotenoidele totale (Car), Uric acid (UA) din serul şi Superoxide dismutaza (SOD), Catalaza (Cat), Glutation peroxidaza (GPx) din sângele integral al cailor VAIE pozitivi şi negative. Rezultatele sunt
exprimate ca boxploturi, cu procentele de 25, 50 (mediana) şi 75, respectiv valorile aberante. 1, 2, 3, 4, 5 şi 6 reprezintă loturile experimentale
Activitatea peroxidazică sanguină generală a fost mai scăzută în cazul loturilor 1 şi 3, care
cuprind cai AIE pozitivi, la mai puţin de 1 an de la depistare, comparativ cu restul loturilor. În cazul loturilor 4, 5 şi 6 valorile activităţii enzimatice înregistrează valori ale mediei foarte apropiate. O privire de ansamblu asupra valorilor mediei activităţii catalazei ne permite însă să afirmăm faptul că activitatea enzimatică înregistrează valori mai scăzute la caii AIE pozitivi care nu au trecut încă în faza de evoluţie asimptomatică a bolii, probabil corelată şi cu prezenţa periodică în circulaţie a virusului la aceste animale. Valorile destul de apropiate ale activităţii enzimatice la caii sănătoşi si la cei cronici asimptomatici, ar putea sugera faptul că aceştia din urmă au reuşit să echilibreze balanţa oxidanţi/antioxidanţi, astfel semnalarea stării de stres oxidativ este mai puţin probabilă în cazul lor. Diferenţele care există între valorile de la caii sănătoşi şi cei infectaţi cu VAIE cronici asimptomatici, faţă de caii bolnavi la mai puţin de un an de la depistare, cu valori mai scăzute în cazul celor infectaţi mai recent, nu au fost semnificative din punct de vedere statistic.
La fel ca în cazul activităţii catalazei, nici în cazul evaluării activităţii peroxidazice generale sanguine nu există nici o diferenţă semnificativă statistică între loturile din experiment, în urma verificării prin aplicarea testului Kruskal-Wallis, respectiv a testului Wilcoxon.
Bolfă Pompei Florin – Rezumat teză de doctorat
13
Valorile scăzute ale activităţii enzimelor antioxidante au drept rezultat acumularea anionilor radical superoxid şi peroxidului de hidrogen în cazul animalelor proaspăt infectate cu VAIE, rezultând astfel o stare prooxidantă persistentă, la nivel celular, care poate favoriza instalarea stresului oxidativ. Această afirmaţie este valabilă şi în cazul activităţii sanguine peroxidazice generale mai ales în cazul cailor infectaţi cu VAIE, la mai puţin de un an de la diagnosticare, indiferent de vârstă, deşi diferenţele dintre lot
i uşoară a unei stări prooxidante la acestea. Acest dezechilibru antioxidanţi/prooxidanţi în favoarea
înregistra
usul seleniului în organism, deoarece GPx este o selenoenzimă ce conţine Se organic sub formă de selenocisteină în centrul activ. Valori scăzute ale GPx pot
ri ar putea fi puse pe seama prezenţei ciclice în circulaţie a virusului la această c
funcţii specializate în organism. Sistemele neenzimatice conţin o varietate mare de biomolecule şi acţioneaz
uri nu sunt semnificative din punct de vedere statistic. În cazul activităţii GPx, la o primă analiză a valorilor obţinute, caii infectaţi cu VAIE par să
înregistreze valori mai scăzute ale enzimei comparativ cu cei sănătoşi. Cea mai scăzută medie pentru activitatea glutation peroxidazei sanguine au fost înregistrate în cadrul lotului 2 compus din animale AIE pozitive, cu vârsta sub 5 ani, la mai mult de un an de la diagnosticare. În schimb, media cea mai crescută pentru activitatea GPx la caii luaţi în studiu a fost înregistrată la lotul 5, de cai AIE negativi, cu vârste sub 5 ani. Din aceste două afirmaţii de mai sus reiese că între caii AIE pozitivi cronici asimptomatici şi caii sănătoşi de aceleaşi vârste, se înregistrează diferenţele cele mai mari, cu o activitate enzimatică mai crescută la animalele sănătoase. Privind mediile activităţii GPx la caii din loturile 1, 2, 3 şi 4 (toţi infectaţi cu VAIE), valorile mediei sunt, în general, aproximativ la jumătatea valorilor înregistrate pentru caii din loturile 5 şi 6 (cai liberi de AIE). Valorile ceva mai crescute ale activităţii GPx la lotul 1 s-ar putea explica în primul rând prin prezenţa a doar 2 animale în acest lot, ceea ce nu este foarte semnificativ din punct de vedere statistic. Această activitate mai scăzută a GPx în cazul animalelor bolnave predispune la instalarea ma
prooxidanţilor ar putea fi probabil corelată şi cu prezenţa ciclică în circulaţie a virusului la aceste animale.
La fel ca şi pentru activitatea SOD şi a catalazei, în cazul GPx, se înregistrează o uşoară scădere a activităţii enzimatice în cadrul cailor bolnavi odată cu înaintarea în vârstă. Această tendinţă este
tă şi în cazul cailor din loturile martor şi este asemănătoare cu cea de la om, unde odată cu înaintarea în vârstă activitatea GPx, dar şi a SOD şi catalazei au tendinţa să scadă (Guemori şi col, 1991).
În cazul activităţii glutation peroxidazei sanguine la cai, se poate afirma faptul că există diferenţe semnificative între caii infectaţi cu virus al AIE şi caii sănătoşi. Astfel, în cazul animalelor sănătoase, valorile activităţii enzimatice sunt semnificativ mai mari faţă de cele înregistrate la caii bolnavi. Activitatea enzimatică a GPx este mai scăzută la animalele cu vârsta de peste 5 ani comparativ cu cele cu vârsta sub 5 ani, atât în cazul cailor sănătoşi cât şi în cazul animalelor infectate cu VAIE, indiferent de perioada scursă de la diagnosticare. Într-un studiu pe cai, Delguste sugerează că o creştere a activităţii GPx ca răspuns la o stare de stres oxidativ ar putea exista dar ar fi de durată limitată în timp (Delguste şi col., 2007). Activitatea GPx este corelată şi cu stat
indica şi un status deficitar al seleniului în organism. Analizând în continuare valorile mediei obţinute în cuantificarea peroxidării lipidelor (MDA),
se pare că în cazul animalelor bolnave la care nu a trecut mai mult de 1 an de la diagnosticare (loturile 1 şi 3) lipoperoxidarea este mai evidentă indiferent de vârstă faţă de cabalinele sănătoase (loturile 5 şi 6). Activităţile scăzute ale enzimelor antioxidante se corelează cu nivelul ridicat al peroxidării lipidelor, manifestări ce denotă prezenţa stării de stres oxidativ la animalele AIE pozitive, la mai puţin de un an de la infectare. Aceste modifică
ategorie de animale. Toate aceste supoziţii urmează a fi verificate prin aplicarea testelor statistice asupra datelor în continuare.
În organismele animale, un rol important în contracararea acţiunii nocive a SRO îl ocupă şi antioxidanţii neenzimatici. Aceştia sunt biomolecule ce au proprietatea de a se oxida uşor şi rapid în prezenţa speciilor reactive de oxigen, protejând astfel împotriva oxidării biomoleculelor cu rol structural sau cu
ă mai ales în spaţiul extracelular, acolo unde enzimele sunt absente sau apar în cantităţi foarte mici.
Din analiza comparativă a nivelului lipidelor totale şi a gradului de peroxidare al lipidelor serice, am constatat ceva interesant: făcându-se o medie a nivelului lipidelor totale de la caii sănătoşi pe care am raportat-o la gradul de peroxidare lipidic de la aceeaşi categorie de animale, am luat în calcul
Bolfă Pompei Florin – Rezumat teză de doctorat
14
valoare lipidelor totale de la fiecare lot în parte şi am calculat valoarea predictivă a peroxidării lipidelor, la care am raportat apoi valoarea reală. Astfel, în cazul lotului 1 (animale AIE pozitive, cu vârsta sub 5 ani şi la mai puţin de 1 an de la diagnosticare) valoarea obţinută în cazul peroxidării lipidelor a fost de aproximativ două ori (2x) mai mare, în cazul lotului 3 (animale AIE pozitive, cu vârsta de peste 5 ani şi la mai puţin de 1 an de la diagnosticare) de aproximativ 1,4x mai mare iar în cazul lotului 4 (animale AIE pozitive, cu vârsta de peste 5 ani şi la mai mult de 1 an de la diagnosticare) de aproximativ 1,19x mai scăzută. De aici am putea deduce faptul că lipoperoxidarea este mai evidentă în cazul cailor recent infectaţi,
ut al lipidelor totale semnificativ statistic se înregistrează între animalele din lotul 1 faţă de caii cu
d ipoteza că odată cu trecerea în faza de purtători
de apărare antioxidantă scăzut, rezultate similare
rea în vârstă creşte si gradul de peroxidare lipidic. Activităţile scăzute a
probabil şi datorită prezenţei antigenului în sânge, şi dintre aceştia la cei cu vârste de sub 5 ani modificările sunt mai evidente.
Diferenţe semnificative statistic (p<0,05) există între loturile 1 cu 3, 1 cu 6 precum şi între loturile 3 şi 4. Această scădere ar putea fi cauzată de un consum mai crescut al lipidelor datorită procesului de lipoperoxidare la caii bolnavi cu vârste sub 5 ani, la mai puţin de un an de la diagnosticare. Un nivel mai scăz
vârste de peste 5 ani, sănătoşi sau bolnavi, indiferent de stadiul de evoluţie al bolii, respectiv loturile 3, 4 şi 6.
În ceea ce priveşte gradul de peroxidare al lipidelor, rezultatele aplicării testului t şi ale testului Wilcoxon sunt similare, respectiv relevă diferenţe statistice reale (p<0,05) între loturile 3 şi 4, respectiv 3 şi 5. Astfel, se observă faptul că gradul de peroxidare al lipidelor din ser este mai crescut în cazul cabalinelor AIE pozitive până la un an de la diagnosticare (lotul 1 dar mai ales lotul 3). Această activitate prooxidantă crescută ar putea fi pusă şi pe seama prezenţei în circulaţie a virusului, animalele nefiind intrate încă în faza asimptomatică a bolii, când se pare că şi balanţa oxidanţi/antioxidanţi se echilibrează. Totuşi, o lipoperoxidare mai accentuată pare să fie prezentă în cazul lotului 3, care este format din animale cu vârste de peste 5 ani, ceea ce sugerează şi faptul că odată cu înaintarea în vârstă, capacitatea de apărare a organismului împotriva speciilor reactive de oxigen scade. De asemenea, în cadrul loturilor de cai infectaţi cu vârste de peste 5 ani, gradul de peroxidare lipidic este semnificativ mai crescut la animalele sub 1 an de la diagnosticare, aceasta confirmân
asimptomatici (când se reduc şi semnele clinice de boală), animalele reuşesc să creeze un echilibru şi în ceea ce priveşte balanţa oxidanţi/antioxidanţi.
Peroxidarea lipidelor reprezintă probabil cel mai investigat proces indus de radicalii liberi de oxigen. Prezenţa din abundenţă a fosfolipidelor de membrană la locurile unde se formează radicali şi în special SRO, le transformă în ţinte endogene, rapid afectate de către radicalii liberi. Grupul acizilor graşi polinesaturaţi (PUFA) este cel mai susceptibil la reacţii cu radicalii liberi. Peroxidarea lipidelor la acizi graşi poate conduce la o reacţie în lanţ care se acompaniază de obicei cu formarea a numeroşi produşi inclusiv alcani şi compuşi carbonilici. Deoarece mulţi dintre aceşti produşi, îndeosebi hidroxi-alchenele, sunt toxici prin ei înşişi, ei pot servi ca mesageri secundari pentru daune produse de radicali (Loeckie şi col., 1999). Creşterea gradului de peroxidare al lipidelor indusă de speciile reactive de oxigen, poate juca un rol în stimularea replicării virusului HIV. Se arată de asemenea în literatura de specialitate că pacienţii HIV pozitivi prezintă un stres oxidativ crescut şi un sistem
cu cele obţinute în lucrarea de faţă. Nu se cunoaşte încă dacă suplimentarea cu antioxidanţi la aceşti pacienţi ar reduce stresul oxidativ (Allard şi col., 1998).
Rezultatele din lucrarea de faţă, în ceea ce priveşte nivelul lipidelor totale şi gradul de peroxidare al lipidelor serice, sugerează faptul că modificările care indică instalarea stării de stres oxidativ din cadrul loturilor de animale AIE pozitive ar fi de durată limitată în timp. Astfel, într-o primă fază, în primul an de la diagnosticare, asemănător populaţiei umane infectate cu virusul HIV, echilibrul oxidanţi/antioxidanţi este în favoarea celor dintâi, urmând ca într-o etapă următoare, respectiv odată cu trecerea în faza asimptomatică de evoluţie a bolii, semnele de stres oxidativ să se estompeze. De asemenea, lipoperoxidarea este mai evidentă în cazul cailor recent infectaţi, indiferent de vârstă, probabil şi datorită prezenţei antigenului din sânge, comparativ cu animalele sănătoase, cu vârste sub 5 ani (lotul 3 cu lotul 5, p<0,05, respectiv lotul 1 cu lotul 5, 0,1<p<0,05), care sunt se pare cel mai bine protejate faţă de stresul oxidativ. Dintre animalele infectate, la cele cu vârste de peste 5 ani, gradul de peroxidare al lipidelor este mai ridicat (lotul 3 cu lotul 4, p<0,05), tendinţă asemănătoare cu animalele sănătoase şi cu populaţia umană, unde odată cu înainta
le enzimelor antioxidante se corelează cu nivelul ridicat al peroxidării lipidelor, manifestări ce denotă prezenţa stării de stres oxidativ.
Bolfă Pompei Florin – Rezumat teză de doctorat
15
Creşterea gradului de peroxidare al lipidelor indusă de speciile reactive de oxigen, poate juca un rol în stimularea replicării virusului HIV (un alt retrovirus, cu mecanisme patogenetice asemănătoare VAIE). A emănător cu pacienţii HIV pozitivi, în cazul cailor AIE pozitivi, există un stres oxidativ crescut şi un siste
către valorile întâlnite la caii sănătoşi. Cu alte cuvi
raţii crescute, acidul uric conferă o protecţie împotriva activităţii radicalilor liberi in vivo, şi indică faptul că
peste 5 ani, la mai puţin de un an de la diagnosticare faţă de caii AIE negativi, indiferent de vârstă. Nici în cazul aplicării acestui test nu au fost înregistra
sm de apărare antioxidantă scăzut. Nivelurile acidului uric la caii infectaţi cu VAIE sunt ceva mai ridicate faţă de cei sănătoşi.
Toate valorile mediei însă, se încadrează în valorile de referinţă 0 – 1,8 mg/dl (Pintea şi col., 2008). Media cea mai scăzută a nivelului acidului uric a fost înregistrată în cadrul lotului 5 compus din
animale AIE negative, cu vârsta de sub 5 ani. La polul opus, media cea mai ridicată a fost înregistrată în dreptul lotului 3, format din animale AIE pozitive, cu vârsta de peste 5 ani, la mai puţin de un an de la diagnosticare. Toate loturile de animale infectate prezintă valori serice mai crescute ale mediei acidului uric faţă de animalele sănătoase. În cadrul loturilor de animale sănătoase, cele cu vârste de peste 5 ani prezintă valori uşor mai ridicate ale acidului uric, tendinţă care se păstrează şi în cazul loturilor de cai infectaţi cu VAIE. În cadrul acestor loturi de animale infectate, nivelul mediei acidului uric este mai crescut la ecvinele la mai puţin de un an de la diagnosticare. După ce animalele trec în faza asimptomatică de evoluţie a bolii, nivelurile acidului uric au tendinţa să scadă,
nte, cu cât se înaintează în vârstă, respectiv cu cât a trecut mai puţin timp de la instalarea infecţiei, nivelele serice ale acidului uric sunt tot mai crescute.
Există un paradox aparent între efectul protector şi cel toxic al acidului uric, care se bazează clinic pe faptul că, compuşi antioxidanţi pot deveni prooxidanţi în anumite situaţii, mai ales atunci când sunt prezenţi în sânge la niveluri supranormale (Lippi şi col., 2008). Astfel, valorile mai crescute ale acidului uric, care apar la animalele recent infectate (sub un an de la diagnosticare), ar putea denota prezenţa unei stări de stres oxidativ. Acidul uric reacţionează cu radicalii liberi derivaţi din oxigen şi se oxidează în muşchii scheletici în timpul exerciţiului intens. Concentraţiile de acid uric intracelular sunt rapid înlocuite de cel din plasmă după exerciţiu. Există o relaţie invers proporţională între concentraţia acidului uric seric şi stres oxidativ în timpul unui exerciţiu aerob acut. Aceste observaţii sugerează că, la concent
acidul uric poate avea o importanţă biologică în instalarea stresului oxidativ acut (Waring şi col., 2003).
Există diferenţe semnificative statistic între mai multe loturi experimentale (p<0,05). Aceste două afirmaţii au fost verificate prin aplicarea testului Wilcoxon, în urma căruia rezultatele obţinute au fost similare. Astfel, s-au înregistrat diferenţe statistice adevărate între loturile 3 cu 5 şi 3 cu 6, la ambele p<0,05. Valori destul de scăzute ale lui p au fost înregistrate şi la analiza comparativă a loturilor 1 cu 5, 1 cu 6, 4 cu 5 şi 4 cu 6. În toate cazurile anterior menţionate, diferenţele au fost între un lot de animale bolnave şi unul de animale sănătoase, cu valori superioare ale acidului uric de partea loturilor de cai bolnavi, în fiecare din situaţii. Se înregistrează astfel un nivel semnificativ statistic (p<0,05), mai ridicat, al acidului uric în cazul cailor AIE pozitivi, cu vârste de
te diferenţe reale între loturile de animale sănătoase. Un ultim marker al stresului oxidativ cuantificat în lucrarea de faţă au fost carotenoidele totale.
În serul cailor infectaţi cu VAIE carotenoidele sunt prezente la un nivel ceva mai ridicat faţă de caii sănătoşi. Carotenoidele totale din serul cailor testaţi au înregistrat valorile cele mai scăzute ale mediei în cadrul lotului 6, de animale sănătoase, compus din cabaline cu vârste mai mari de 5 ani. În schimb, valorile cele mai ridicate ale mediei au fost înregistrate la lotul 1 de animale bolnave, format din cai cu vârsta sub 5 ani, la mai puţin de un an de la diagnosticare. La fel ca şi în cazul acidului uric, şi în cazul carotenoidelor serice totale, toate loturile de animale infectate prezintă valori ale mediei mai crescute faţă de animalele sănătoase. Atât în cadrul loturilor de animale sănătoase cât şi în cadrul celor AIE pozitive, carotenoidele sunt prezente în cantităţi mai reduse odată cu înaintarea în vârstă. În ceea ce priveşte intervalul trecut de la diagnosticare, în cazul cailor AIE pozitivi, mai tineri de 5 ani, asemănător cu tendinţa înregistrată în cazul acidului uric, cu cât trece mai mult timp de la diagnosticarea infecţiei, valorile carotenoidelor au tendinţa să scadă către nivelul celor din lotul martor de aceeaşi vârstă. Un nivel seric mai crescut al carotenoidelor ar putea avea două explicaţii, una fiind un aport alimentar crescut, iar cealaltă explicaţie ar fi prezenţa în cantitate crescută a radicalilor liberi la caii din loturile cu AIE, iar
Bolfă Pompei Florin – Rezumat teză de doctorat
16
antioxidanţii serici neenzimatici (printre care şi carotenoidele) apar în cantitate crescută pentru a-i neutraliza. Acest nivel ridicat al SRO ar putea fi datorat prezenţei virusului anemiei infecţioase ecvine în circulaţie. Pe de altă parte, s-ar sugera faptul că la animalele mai tinere sistemul de protecţie împotriva oxidării e
. u există diferenţe semnificative statistic întreloturi în ceea ce priveşte nivelul carotenoidelor
totale.
stic care să su
există şi
lângă scăderea activităţii antioxida
e diagnostic aplicabile in condiţii de teren trebuie dezvoltate pentru a putea evalua statusul antioxidant al cailor, mai ales a celor de competiţie, în vederea definirii nevoilor specifice în antioxidanţi ale acestora.
ste mai bine pus la punct, aceste animale adaptându-se mai uşor unei eventuale stări de stres oxidativ
N
Rezumând toate informaţiile de mai sus, constatăm că există diferenţe semnificative statigereze prezenţa unui dezechilibru oxidanţi/antioxidanţi, în primul rând în cazul lotului 3 (şase
markeri), apoi în cazul lotului 1 (doi markeri) dar şi în cazul lotului 2 şi respectiv 4 (câte un marker). Se pare aşadar că infecţia cu virus AIE determină la caii cu vârste de peste 5 ani, în primul an de
la diagnosticare un dezechilibru major oxidanţi/antioxidanţi care determină instalarea la aceştia a unei stări de stres oxidativ. Caii mai tineri, cu vârste sub 5 ani, au un status antioxidant mai bine pus la punct, şi în cazul unei infecţii virale (VAIE), se adaptează ceva mai bine în primul an de la diagnosticare faţă de caii mai vârstnici. În schimb, odată cu trecerea a mai mult timp de la diagnosticare (peste un an), organismul reuşeşte la aceşti cai în faza asimptomatică de evoluţie a bolii, să echilibreze balanţa prooxidanţi/antioxidanţi, probabil şi datorită prezenţei sporadice a virusului în circulaţie. Cu toate acestea,
în cadrul acestor loturi semne, într-adevăr mai discrete, de stres oxidativ. În schimb, nu există diferenţe semnificative statistic între cele două categorii de vârstă (sub 5 ani şi peste 5 ani) de cai sănătoşi.
Din punct de vedere experimental, în starea de stres oxidativ, pe nte enzimatice şi neenzimatice se remarcă o creştere a concentraţiei SRO respectiv a produşilor
rezultaţi prin degradarea oxidativă a lipidelor, proteinelor şi a acizilor nucleici. Dezechilibrul oxidanţi/antioxidanţi (indus de exerciţiu sau corelat cu o boală) este cercetat în
prezent de mai multe grupuri de cercetători. Prevenirea şi modularea stresului oxidativ se conturează ca o nouă ţintă in medicina ecvină. Pornind de la aceste date se poate ajunge şi la modularea nutriţiei ecvinelor, în funcţie de efortul depus şi de cerinţele în antioxidanţi (Moffarts şi col., 2005, Williams, 2003). De aceea, metodele d
Bolfă Pompei Florin – Rezumat teză de doctorat
17
EXAMENUL PARACLINIC LA CAI INFECTAŢI CU VIRUSUL AIE
MATERIAL BIOLOGIC ŞI METODE DE LUCRU
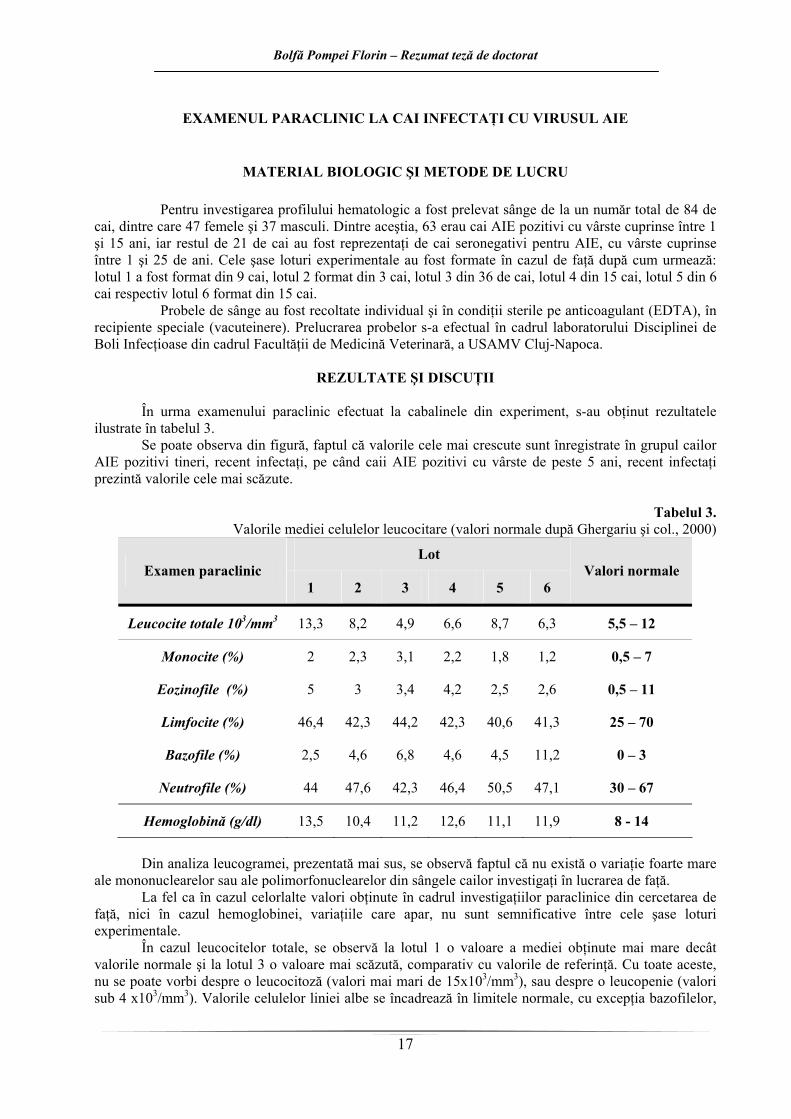
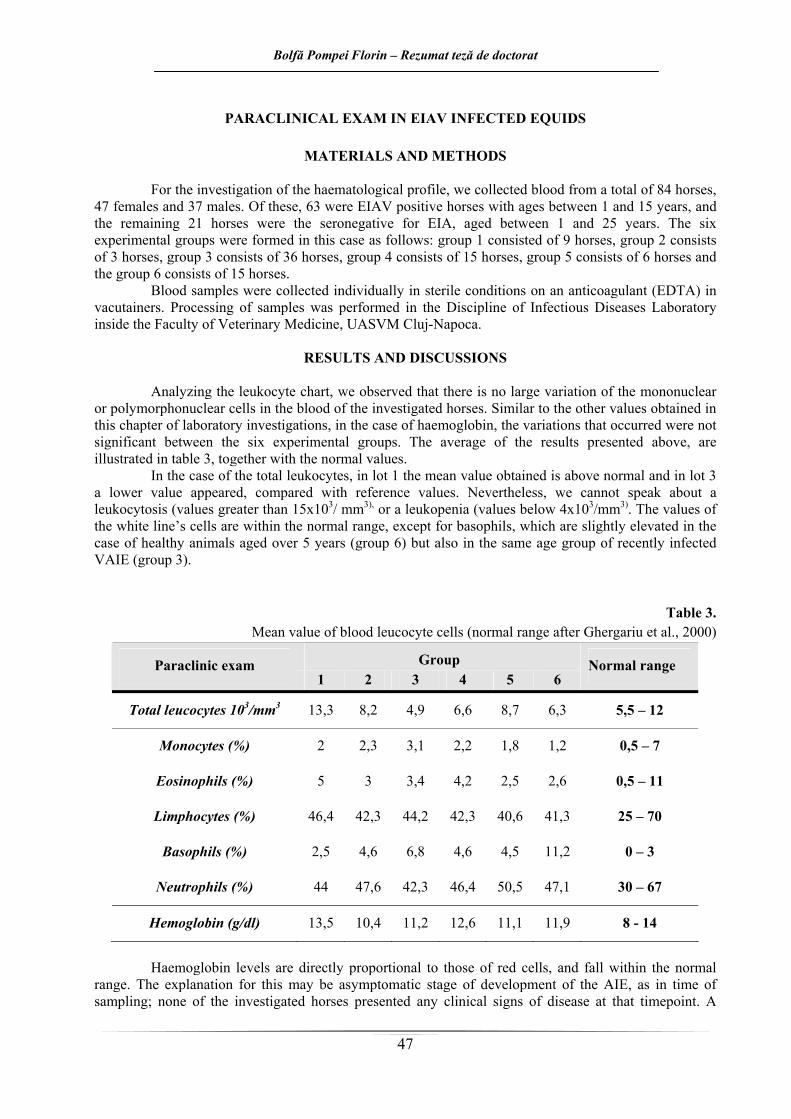
Pentru investigarea profilului hematologic a fost prelevat sânge de la un număr total de 84 de cai, dintre care 47 femele şi 37 masculi. Dintre aceştia, 63 erau cai AIE pozitivi cu vârste cuprinse între 1 şi 15 ani, iar restul de 21 de cai au fost reprezentaţi de cai seronegativi pentru AIE, cu vârste cuprinse între 1 şi 25 de ani. Cele şase loturi experimentale au fost formate în cazul de faţă după cum urmează: lotul 1 a fost format din 9 cai, lotul 2 format din 3 cai, lotul 3 din 36 de cai, lotul 4 din 15 cai, lotul 5 din 6 cai respectiv lotul 6 format din 15 cai.
Probele de sânge au fost recoltate individual şi în condiţii sterile pe anticoagulant (EDTA), în recipiente speciale (vacuteinere). Prelucrarea probelor s-a efectual în cadrul laboratorului Disciplinei de Boli Infecţioase din cadrul Facultăţii de Medicină Veterinară, a USAMV Cluj-Napoca.
REZULTATE ŞI DISCUŢII
În urma examenului paraclinic efectuat la cabalinele din experiment, s-au obţinut rezultatele
ilustrate în tabelul 3. Se poate observa din figură, faptul că valorile cele mai crescute sunt înregistrate în grupul cailor
AIE pozitivi tineri, recent infectaţi, pe când caii AIE pozitivi cu vârste de peste 5 ani, recent infectaţi prezintă valorile cele mai scăzute.
Tabelul 3.
Valorile mediei celulelor leucocitare (valori normale după Ghergariu şi col., 2000)
Din analiza leucogramei, prezentată mai sus, se observă faptul că nu există o variaţie foarte mare
ale mononuclearelor sau ale polimorfonuclearelor din sângele cailor investigaţi în lucrarea de faţă. La fel ca în cazul celorlalte valori obţinute în cadrul investigaţiilor paraclinice din cercetarea de
faţă, nici în cazul hemoglobinei, variaţiile care apar, nu sunt semnificative între cele şase loturi experimentale.
În cazul leucocitelor totale, se observă la lotul 1 o valoare a mediei obţinute mai mare decât valorile normale şi la lotul 3 o valoare mai scăzută, comparativ cu valorile de referinţă. Cu toate aceste, nu se poate vorbi despre o leucocitoză (valori mai mari de 15x103/mm3), sau despre o leucopenie (valori sub 4 x103/mm3). Valorile celulelor liniei albe se încadrează în limitele normale, cu excepţia bazofilelor,
Bolfă Pompei Florin – Rezumat teză de doctorat
18
care sunt uşor crescute mai ales în cazul animalelor cu vârste de peste 5 ani sănătoase (lotul 6) dar şi recent infectate cu VAIE (lotul 3).
Valorile hemoglobinei sunt direct proporţionale cu cele ale hematiilor, şi se încadrează în limitele normale în cazul de faţă. Explicaţia acestui fapt poate fi stadiul asimptomatic de evoluţie al AIE, deoarece în momentul recoltării, niciunul dintre caii investigaţi nu prezenta semne clinice de boală. Un singur examen hematologic însă, este insuficient pentru diagnostic, fiind necesară efectuarea unor exemene repetate, concordante cu ascensiunile termice, deoarece între episoadele febrile, parametri hematologici pot reveni la normal (Moga-Mânzat, 2005).
Modificările paraclinice asociate cu AIE sunt: trombocitopenia moderată până la gravă, anemia severă normocitară, normocromă, oligocitemică, cu hematocrit până la 14-20%, leucopenie (până la 2000 celule/mm3) cu neutropenie şi limfopenie, hipergammaglobulinemie şi activitatea crescută a enzimelor biliare, prezenţa siderocitelor, considerate specifice AIE (Barna şi col., 1995). Dintre aceste modificări, au fost identificate în lucrarea de faţă doar o hipergamaglobulinemie la caii bolnavi, parametru care se pare că este cel mai fiabil dintre cei investigaţi în a reda adevăratul status al calului investigat (infectat sau nu cu VAIE).
Bolfă Pompei Florin – Rezumat teză de doctorat
19
COMPARAREA UNUI TEST COMERCIAL ELISA COMPETITIV CU IMUNODIFUZIA ÎN GEL DE AGAR PENTRU SERODIAGNOSTICUL ANEMIEI INFECŢIOASE ECVINE
Testul serologic utilizat în prezent în multe ţări în cadrul programelor de diagnostic şi control al
AIE este testul de imunodifuzie în gel de agar (Bolfă şi col., 2008b; Toledo şi col., 2007), ce detectează anticorpii precipitanţi împotriva proteinei interne specifice de grup p 26, care este un antigen stabil (Toledo şi col., 2007).
Deoarece prezintă o sensibilitate mai crescută, au fost propuse diferite metode imunoenzimatice (ELISA) pentru diagnosticul serologic al AIE, care folosesc particule virale purificate ca antigen (Pare şi Simard, 2004) sau proteine virale recombinate (Birket şi col., 1997; Toledo şi col., 2007). În câteva situaţii rare, putem întâlni rezultate înşelătoare, atunci când nivelul de virus circulant din timpul unui episod acut al bolii este suficient încât să lege toţi anticorpii disponibili, şi dacă răspunsul imun umoral iniţial nu atinge praguri detectabile (Toma, 1980). Deşi ELISA detectează anticorpii mai devreme şi la concentraţii mai scăzute decât testul AGID, rezultatele pozitive ELISA se confirmă prin testul AGID. Se recurge la aceasta deoarece, în cazul ELISA au fost observate rezultate fals pozitive. Testul AGID mai prezintă şi avantajul de a distinge între răspunsuri antigen – anticorp AIE şi non-AIE prin linii de identitate (OIE, 2010). Pentru identificarea agentului etiologic în AIE se poate recurge la izolarea şi identificarea virusului sau la reacţia în lanţ a polimerazei (PCR).
În contextul în care AIE reprezintă boala infecţioasă care produce cele mai importante daune economice în cadrul efectivelor de cabaline din România, s-ar impune în viitor introducerea în practica veterinară a unui test rapid şi eficace, care permite testarea eficace, cu acurateţe a unui număr mare de probe serologice într-o perioadă scurtă de timp şi a cărui citire este una obiectivă prin utilizarea unui spectrofotometru
MATERIAL ŞI METODE Experimentul de faţă a fost împărţit pe două direcţii: într-o primă etapă s-au testat probe de ser
recoltate de la cai diagnosticai ca AIE pozitivi în urmă cu cel puţin 1 an şi maxim 7 ani şi 6 luni (28 cabaline) pentru a observa eventuale diferenţe de reacţie faţă de AGID odată cu trecerea diverselor perioade de la diagnosticare. O a doua etapă a vizat retestarea serurilor pozitive de la un număr de 64 de cai, pentru a verifica procentul de corelaţie/concordanţă între cele două teste (AGID şi cELISA). Toate probele de sânge de la cei 92 de cai testaţi, au provenit de la animale de pe raza judeţului Bistriţa-Năsăud.
Pentru prima parte a studiului de faţă au fost utilizate probe de ser provenind de la 28 de cai, dintre care 20 de femele şi 8 masculi, cu vârste cuprinse între 1 şi 18 ani. AIE a fost diagnosticată prin testul oficial (AGID) în cadrul Laboratorului Sanitar Veterinar şi pentru Sănătate Animală (LSVSA) judeţean din Bistriţa, în cadrul planului bianual de testare prevăzut de ANSVSA. Animalele au fost diagnosticate ca pozitive la AIE între anii 1998 şi 2005, iar serul de la aceste animale, recoltat în 2005 şi 2006 a fost retestat prin metoda imunoenzimatică (kitul IDEXX EIA, cELISA), conform recomandărilor producătorului. Retestarea s-a efectuat la perioade între 1 şi 7 ani 6 luni după diagnosticarea iniţială. Testarea a avut loc în cadrul Laboratorului Disciplinei de Boli Infecţioase al USAMV Cluj-Napoca, la finalul anului 2006.
Pentru cea de a doua parte a experimentului, cele 64 probe de ser au provenit prin amabilitatea LSVSA Bistriţa, în primăvara anului 2008, după testarea lor ca pozitive prin metoda AGID, şi au fost retestate prin cELISA în toamna anului 2008 în cadrul Laboratorului Disciplinei de Anatomie Patologică, Diagnostic Necropsic şi Medicină Legală şi cea de Parazitologie din cadrul USAMV Cluj-Napoca, la finalul anului 2006.
Analiza statistică a rezultatelor a fost efectuată cu ajutorul programului R, utilizând testul Fisher exact (pentru analiza tabelelor de contingenţă unde numărul de probe este scăzut).
Bolfă Pompei Florin – Rezumat teză de doctorat
20
REZULTATE ŞI DISCUŢII
În Statele Unite ale Americii există în prezent patru metode ELISA care sunt aprobate pentru diagnosticul AIE de către Departamentul de Agricultură, şi pot fi aplicate la nivel internaţional: o metodă ELISA competitivă şi trei metode ELISA non-competitive. Metoda ELISA competitivă şi o metodă non-competitivă detectează anticorpii produşi împotriva antigenului proteinei interne p26. O altă metodă non-competitivă încorporează atât antigen al proteinei interne p26 şi gp45 (proteina virală transmembranară), pe când ultima metodă non-competitivă detectează anticorpii anti proteină gp45. În toate testele se utilizează protocoale ELISA tipice (OIE, 2010).
Un rezultat pozitiv ELISA trebuie retestat prin intermediul testului AGID pentru a confirma diagnosticul, datorită posibilităţii apariţiei unor rezultate fals pozitive. Rezultatele pot fi de asemenea confirmate prin tehnica imunoblot (OIE, 2010).
Kitul Idexx ELISA competitiv (EIA cELISA) reprezintă un test rapid, convenabil şi specific pentru detecţia anticorpilor anti VAIE în ser de cabaline. Antigenul p26 purificat şi anticorpii monoclonali au rolul de a reduce reacţiile nespecifice întâlnite frecvent în testele ELISA. Corelaţia între Idexx EIA cELISA şi testul de imunodifuzie în gel de agar este de peste 99%, conform producătorului.
Rezultatele obţinute în cercetarea de faţă au fost interpretate atât vizual urmărind schimbările de culoare) cât şi spectrofotometric (citirea densităţilor optice la spectrofotometrul PR1100 Bio- Rad, λ=620 nm).
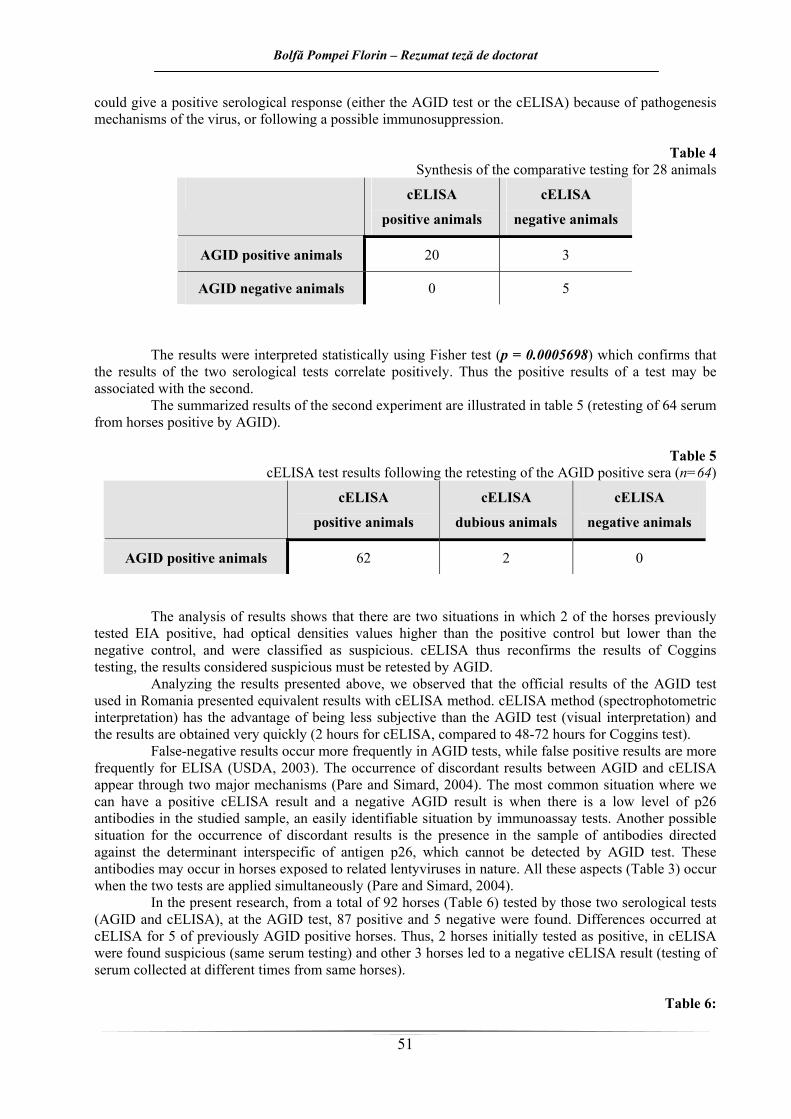
Rezultatele de la primul experiment sunt ilustrate în tabelul 4. Din cele 28 de animale testate, 23 erau pozitive la testul Coggins (diagnosticare efectuată în urmă cu până la 7 ani şi 6 luni) şi 5 au fost animale negative la AIE. Numărul de reacţii negative utilizând metoda cELISA a crescut de la 5 la 8 (apariţia unei culori albastru închis, respectiv o lungime de undă mai mare decât cea a martorului negativ), aşa cum se observă şi în continuare.
Tabelul 4. Sinteza rezultatelor testării comparative a 28 de cabaline
Test Animale cELISA pozitive
Animale cELISA negative
Animale AGID pozitive 20 3
Animale AGID negative 0 5
Din cei trei cai anterior testaţi ca şi pozitivi la AIE prin intermediul AGID şi care au fost testaţi ca
negativi la cELISA, unul a fost diagnosticat în urmă cu 7 ani şi 6 luni, altul în urmă cu 6 ani şi 6 luni şi unul cu 1 an mai devreme faţă de perioada recoltării pentru testare. Această negativizare poate fi explicată prin trecerea cailor respectivi în stadiul asimptomatic de evoluţie a bolii (Bolfă şi col., 2008a). În afară de mânjii proveniţi din mame AIE pozitive şi care consumă sau au consumat colostru, acest tip de rezultate (fals negative) pot fi întâlnite de asemenea la caii care au fost infectaţi şi nu dezvoltă un răspuns imun. De asemenea, atunci când un cal se află într-un episod acut de AIE, nivelul de viremie prezent este suficient pentru a lega toţi anticorpii serici disponibili, făcând imposibilă detectarea unui astfel de animal (USDA, 2003). Acesta este şi unul din motivele pentru care un animal ce prezintă un rezultat serologic pozitiv la testul Coggins este considerat pozitiv pe viaţă şi nu mai este retestat. Oricum, în cazul unei eventuale retestări, la un moment ulterior, s-ar putea ca cele trei animale să prezinte din nou un răspuns serologic pozitiv (fie la testul AGID, fie la cELISA) datorită mecanismelor patogenetice ale virusului, sau în urma unei eventuale imunosupresii.
Rezultatele obţinute au fost interpretate statistic prin testul Fisher (p=0,0005698 – foarte semnificativ statistic) care confirmă faptul că rezultatele la cele două teste serologice se corelează pozitiv. Astfel rezultatele pozitive ale unui test se pot asocia cu cel de al doilea.
În tabelul 5 sunt ilustrate rezultatele sintetizate ale celui de al doilea experiment (retestarea celor 64 de seruri de la cai AGID pozitivi).
Bolfă Pompei Florin – Rezumat teză de doctorat
21
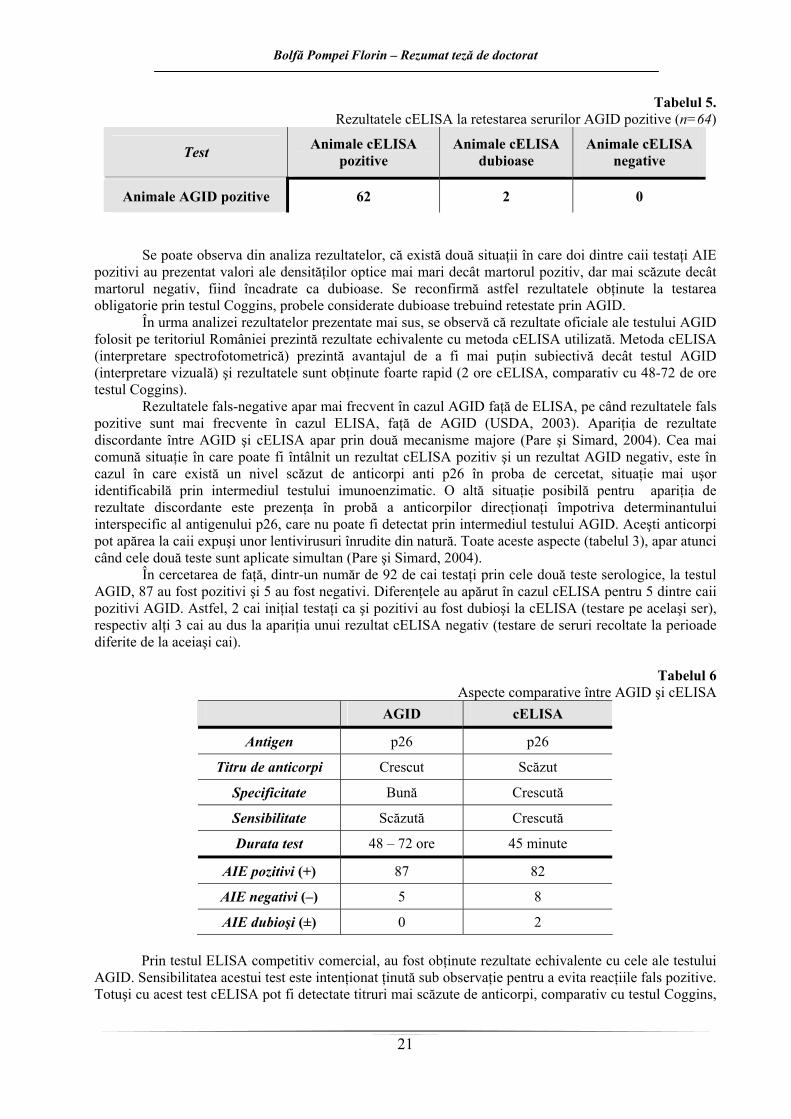
Tabelul 5. Rezultatele cELISA la retestarea serurilor AGID pozitive (n=64)
Test Animale cELISA pozitive
Animale cELISA dubioase
Animale cELISA negative
Animale AGID pozitive 62 2 0
Se poate observa din analiza rezultatelor, că există două situaţii în care doi dintre caii testaţi AIE
pozitivi au prezentat valori ale densităţilor optice mai mari decât martorul pozitiv, dar mai scăzute decât martorul negativ, fiind încadrate ca dubioase. Se reconfirmă astfel rezultatele obţinute la testarea obligatorie prin testul Coggins, probele considerate dubioase trebuind retestate prin AGID.
În urma analizei rezultatelor prezentate mai sus, se observă că rezultate oficiale ale testului AGID folosit pe teritoriul României prezintă rezultate echivalente cu metoda cELISA utilizată. Metoda cELISA (interpretare spectrofotometrică) prezintă avantajul de a fi mai puţin subiectivă decât testul AGID (interpretare vizuală) şi rezultatele sunt obţinute foarte rapid (2 ore cELISA, comparativ cu 48-72 de ore testul Coggins).
Rezultatele fals-negative apar mai frecvent în cazul AGID faţă de ELISA, pe când rezultatele fals pozitive sunt mai frecvente în cazul ELISA, faţă de AGID (USDA, 2003). Apariţia de rezultate discordante între AGID şi cELISA apar prin două mecanisme majore (Pare şi Simard, 2004). Cea mai comună situaţie în care poate fi întâlnit un rezultat cELISA pozitiv şi un rezultat AGID negativ, este în cazul în care există un nivel scăzut de anticorpi anti p26 în proba de cercetat, situaţie mai uşor identificabilă prin intermediul testului imunoenzimatic. O altă situaţie posibilă pentru apariţia de rezultate discordante este prezenţa în probă a anticorpilor direcţionaţi împotriva determinantului interspecific al antigenului p26, care nu poate fi detectat prin intermediul testului AGID. Aceşti anticorpi pot apărea la caii expuşi unor lentivirusuri înrudite din natură. Toate aceste aspecte (tabelul 3), apar atunci când cele două teste sunt aplicate simultan (Pare şi Simard, 2004).
În cercetarea de faţă, dintr-un număr de 92 de cai testaţi prin cele două teste serologice, la testul AGID, 87 au fost pozitivi şi 5 au fost negativi. Diferenţele au apărut în cazul cELISA pentru 5 dintre caii pozitivi AGID. Astfel, 2 cai iniţial testaţi ca şi pozitivi au fost dubioşi la cELISA (testare pe acelaşi ser), respectiv alţi 3 cai au dus la apariţia unui rezultat cELISA negativ (testare de seruri recoltate la perioade diferite de la aceiaşi cai).
Tabelul 6
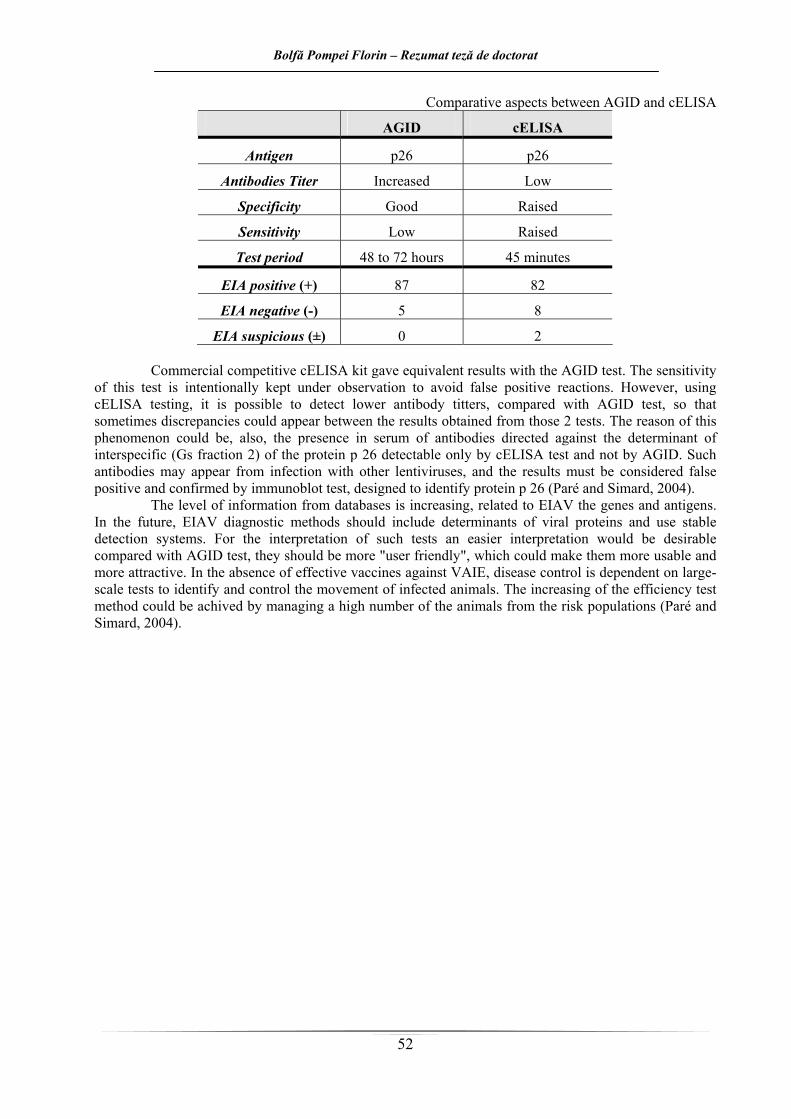
Aspecte comparative între AGID şi cELISA AGID cELISA
Antigen p26 p26
Titru de anticorpi Crescut Scăzut
Specificitate Bună Crescută
Sensibilitate Scăzută Crescută
Durata test 48 – 72 ore 45 minute
AIE pozitivi (+) 87 82
AIE negativi (–) 5 8
AIE dubioşi (±) 0 2 Prin testul ELISA competitiv comercial, au fost obţinute rezultate echivalente cu cele ale testului
AGID. Sensibilitatea acestui test este intenţionat ţinută sub observaţie pentru a evita reacţiile fals pozitive. Totuşi cu acest test cELISA pot fi detectate titruri mai scăzute de anticorpi, comparativ cu testul Coggins,
Bolfă Pompei Florin – Rezumat teză de doctorat
22
astfel că, uneori apar discordanţe între rezultatele obţinute la cele 2 teste. Cauza acestui fenomen poate fi pusă şi pe seama prezenţei în ser a anticorpilor direcţionaţi împotriva determinantului interspecific (fracţiunea Gs 2) al proteinei p 26 decelabilă numai prin testul C-ELISA şi nu prin AGID. Asemenea anticorpi pot apărea ca urmare a infecţiei cu alte lentivirusuri, iar rezultatele trebuie considerate fals pozitive şi confirmate prin teste immunoblot, destinate să identifice proteina p 26 (Paré şi Simard, 2004).
Pe măsură ce cresc informaţiile din bazele de date în ceea ce privesc genele şi antigenele VAIE, vor fi cu siguranţă încercate abordări noi în ceea ce priveşte diagnosticul acurat al AIE. Dacă s-ar putea efectua deja adăugiri la protocoalele testelor deja în uz, acestea ar trebui să includă determinanţi de la câteva proteine virale ale VAIE, precum şi să folosească un format cu reactivi şi sisteme de detectare stabile. Pentru interpretarea unor astfel de teste ar fi nevoie de o interpretare mai facilă comparativ cu testul AGID, ar trebui să fie „prietenoase” cu utilizatorul, ceea ce le-ar face mult mai uşor de utilizat şi mai atractive. În absenţa unor vaccinuri eficiente împotriva VAIE, controlul bolii este dependent de testările la scară largă pentru identificarea şi controlul mişcării animalelor infectate. Creşterea eficienţei metodei de testare se realizează prin controlul unui număr cât mai crescut de animale din populaţiile cu risc (Paré şi Simard, 2004).
Bolfă Pompei Florin – Rezumat teză de doctorat
23
PRIMA CARACTERIZARE GENETICĂ A IZOLATELOR ROMÂNEŞTI DE TEREN ALE VAIE
MATERIAL ŞI METODE Pentru testele de biologie moleculară prezentate în continuare, s-au folosit ca materie primă
celulele albe („buffy coat”) ale sângelui recoltat pe anticoagulant (EDTA) de la 25 de cai din judeţul Bistriţa-Năsăud, Ecvinele au fost testate ca şi pozitive pentru AIE înainte de recoltare cu 3 săptămâni până la 7 ani, în cadrul LSVSA Bistriţa, tstările fiind cuprinse în planul naţional de testare bianulă a tuturor ecvinelor cu vârste de peste 6 luni din regiunea respectivă, considerată cu risc epidemiologic înalt. După recoltarea pe anticoagulant, sângele a fost transportat în cadrul USAMV Cluj Napoca, Disciplina de Boli Infecţioase. Aici s-a recurs la centrifugarea (6000 rpm timp de 10 minute) şi îndepărtarea plasmei, respectiv decolarea cu o pipetă cu vârf a fracţiunii albe obţinute între plasmă şi hematii. Aceasta a fost în continuare menţinută la temperatură scăzută (-20°C), apoi transportată în cadrul laboratorului de Retrovirusologie, Evoluţie şi Cancer, UMR 754 din cadrul Universităţii Claude Bernard, Lyon, Franţa unde au fost procesate în continuare.
Analiza moleculară a probelor
Extracţia ADN-ului genomic
ADN-ul genomic a fost extras din probele de „buffy coat” cu ajutorul kitului „Qiamp®” (Qiagen),
respectând recomandările de lucru ale furnizorului, cu câteva excepţii: a fost dublată cantitatea de tampon AL, proteinază K şi etanol absolut, datorită volumului crescut (≈ 200μl) de „buffy coat”. Întegritatea ADN-ului extras a fost verificată prin migrare electroforetică (3 μl probă,3 μl ladder 1 kb şi 2 μl mass loading dye 6X) pe un gel de agaroză cu concentraţia de 0,8%, conţinând „SYBR® Safe”, (Molecular Probes, Franţa), agent colorant intercalar de acizi nucleici diluat 1/10.000, în tampon TAE 1X (40mM de Tris, 20 mM acetat de sodiu şi 1mM de EDTA, pH 8,3). ADN-urile au fost vizualizate prin transiluminare cu ultraviolete. Concentraţia de ADN a fost măsurată prin citire a absorbanţei la 260 nm cu ajutorul spectrofotometrului „SmartSpec® 3000” (BIORAD). Puritatea materialului genetic a fost verificată prin raportul 260nm/280nm.
Amplificarea prin reacţia în lanţ a polimerazei (PCR)
Controlul ADN-ului Controlul calitativ al ADN-ului pentru amplificare a fost efectuat în urma amplificării prin reacţia
în lanţ a polimerazei (PCR) a unei părţi din gena celulară β-actină (Nagarajan şi Simard, 2001). Amestecul PCR a conţinut 500 ng de ADN/10μl , 1 μl de 10 μM din fiecare primer specific pentru gena β-actinei (GnC5 şi GnC6), 1 μl de 10 mM de dNTP (dezoxinucleotid-trifosfat), 0,25 μl de Go Taq DNA Polimerază 5 U/μl, 10 μl de tampon 5X Green Go Taq Buffer, pentru un volum final de 50 μl.
Verificarea calităţii a fost efectuată cu amorsele (primerii) β-actină 5’ (5’-ATG TGC AAG GCC GGC TTC G-3’) şi β-actină 3’ (5’-TTA ATG TCA CGC ACG ATT TCC-3’) (Nagarajan şi Simard, 2001), concepuţi după secvenţă umană de β-actină, care generează un fragment de 580 perechi de baze (pb). Condiţiile desfăşurării PCR-ului au fost următoarele: 5 minute (min) la 95°C pentru denaturarea iniţială, 35 cicluri de 30 secunde (s) la 95°C apoi 30 s la 56°C apoi 30 s la 72°C, şi 10 min la72°C pentru elongarea finală. Produşii PCR au fost vizualizaţi prin transiluminare cu ultraviolete după migrare electroforetică pe un gel de agaroză de 1,5 % conţinând „SYBR® Safe”, 1/10.000, în tampon TAE 1X.
În afară de proba 1279, ADN-ul provenind de la probele din experiment a fost amplificabil.
Bolfă Pompei Florin – Rezumat teză de doctorat
24
Amplificarea VAIE Amplificarea regiunii din gena gag care codifică capsida virală, a fost realizată prin PCR hemi-
nested („în semi-cuib”). Pentru PCR-ul extern, 500 ng de ADN din fiecare probă au fost amestecate cu 10 mM din fiecare primer, 10 mM de dNTP, 5 unităţi de Go Taq DNA Polimerază, tampon 5X Green Go Taq Buffer, pentru obţinera unui volum final de 50 μl. Primerii utilizaţi au fost EIAV3 şi EIAV5. Pentru PCR-ul intern, 5 μl din produşii de la PCR-ul extern au fost amestecaţi la 10 mM din fiecare primer, 10 mM de dNTP, 5 unităţi de Go Taq DNA Polimerază, tampon 5X Green Go Taq Buffer, pentru obţinera unui volum final de 50 μl. Primerii utilizaţi au fost EIAV4 şi EIAV5. Recaţiile PCR au generat fragmente de 260 şi 210 pb. Condiţiile pentru reacţii au fost următoarele: 5 min la 95°C pentru denaturarea iniţială 35 cycluri de 30s la 95°C - 30s la 56°C - 1min la 72°C, urmate de 10min la 72°C pentru elongarea finală. Produşii PCR au fost vizualizaţi prin traniluminare ultravioletă după migrare electroforetică pe un gel de agaroză de 2% conţinând SyBr® Safe la 1/10.000, într-un tampon TAE 1X (figura 7).
Figura 7. Amplificarea prin PCR a genei gag VAIE pornind de la PBMC. Electroforeză pe gel de agar 2% colorat cu Sybr®Safe. Produşii amplificaţi la 580 pb corespund genei celulare β-actină, ampliconii nerelevaţi de 260 pb corespuns la PCR-ul extern pentru gena gag, iar ampliconii de 210 pb corespund
PCR-ului intern al genei gag. Martorii pozitivi corespund ADN-ului limfoganglionar şi hepatic de la calul 983. Martorul negativ este reprezentat de apă în locul ADN-ului genomic de cal
Amplificarea genei β -actină
Amplificarea genei gag
Bolfă Pompei Florin – Rezumat teză de doctorat
25
Clonajul produşilor gag Ampliconii genei gag au fost obţinuţi prin clonaj TA. Această tehnică se bazează pe o proprietate
a ADN polimerazei Taq care adaugă o deoxiadenozină (A) singură la capătul 3’ al produsului PCR. Inserarea produşilor obţinuţi în urma amplificării se face la nivelul extremităţii T (deoxitimidină) 5’ al plamedei vector. Legarea ampliconilor gag cu plasmida vector pCR®2.1 (Promega, Franţa) (figura 2.5.2), a fost realizată respectând recomandările de lucru ale furnizorului. Produşii de ligatură au fost utilizaţi pentru transformarea în bacterii competente Escherichia coli JM 109. Plasmida pCR®2.1 conţine o genă de rezistenţă la ampicilină, care codifică β-lactamaza, ce permite selecţionarea bacteriilor transformate conţinând vectorul, precum şi gena LacZ ce codifică β-galactozidaza asupra controlului promotorului operonului lactozei, care permite identificarea bacteriilor transformate care au integrat şi insertul plasmidic. Introducerea unui insert în gena LacZ, întrerupe cadrul său de lectură.
Bacteriile au fost în cultivate pe un mediu selectiv agar LB (10g/l de bacto-tryptonă, 5g/l de extract levuric, 10g/l de clorură de sodiu, 15g/l de agar) care conţine ampicilină la 50 μg/ml, ce permite selecţionarea bacteriilor transformate. Mediul a fost suplimentat cu 10μmol de IPTG (IsoPropyl-β-D-ThioGalactopyranozidă), inductorul operonului lactozei, şi cu 1μg de X-gal (5-bromo-4-chloro-3-indolyl-β-D-Galactozidă), substrat cromogenic al β galactozidazei. În prezenţa lactozei, bacteriile în care plasmida este integrată nu au fost capabile să exprime β galactozidaza şi au determinat apariţia de colonii albe. Coloniile albastre reprezintă bacteriile capabile să hihrolizeze substratul X-gal datorită prezenţei plasmidei integrate, funcţionalizând gena Lac-Z.
Plasmidele au fost în continuare extrase utilizând kitul „Wizard® Plus Minipreps DNA Purification System” (Promega, Franţa), apoi digerate cu enzima de restricţie EcoRI ale cărei situsuri sunt localizate de o parte şi de alta a situsurilor de clonaj. Produşii de digestie au fost vizualizaţi prin transiluminare ultravioletă după electroforeză pe un gel de agaroză 1,2% conţinând SyBr® Safe la 1/10.000, într-un tampon TAE 1X. Coloniile (câte 10) au fost reînsămânţate în într-un bulion LB (Luria-Bertani: 10g/l de bacto-triptonă, 5g/l de extract de levură, 10g/l de clorură de sodiu) conţinând ampicilină la 50 μg/ml, şi incubate timp de o noapte la 37°C într-un agitator mecanic.
Extracţia plasmidică
Pornind de la culturile individuale, ADN-ul plasmidic a fost extras utilizând kitul „PLASMID
MIDI KIT® DNA purification system” (Promega, France) (Sambrook şi Rossel, 2001). Produşii de extracţie digeraţi cu ajutorul enzimei de restricţie EcoRI (Promega, Franţa) au fost vizualizaţi după migrare electroforetică pe un gel de agaroză 0,8% conţinând SyBr® Safe la 1/10.000, într-un tampon TAE 1X, prin transiluminare cu ultraviolete. Concentraţia ADN-ului plasmidic a fost măsurată prin citirea absorbanţei la 260 nm la spectrofotometrul „SmartSpec® 3000”. Puritatea ADN-ului a fost verificată prin citirea raportului 260nm/280nm. Clonele obţinute au fost identificate după următoarea nomenclatură: pEIAVX, unde „p” reprezintă originea plasmidică a ADN-ului, „EIAV” – equine infectious anemia virus (virusula anemiei infecţioase ecvine) şi „X” corespunde numărului de identificare.
Secvenţializarea regiunii gag
ADN-ul plasmidic al clonelor conţinând un insert gag au fost secvenţializate (Genomexpress,
Grenoble, Franţa), utilizând amorsele universale T7 (5’-TAA TAC GAC TCA CTA TAG GGC GA -3’) şi SP6 (5’-ATT TAG GTG ACA CTA TAG AAT ACT-3’).
Analiza secvenţelor
Alinierea secvenţelor, contrucţia arborilor filogenetici şi calculul matricilor de divergenţă au fost
realizate utilizând programul Vector NTI Advance (TM) 11.0 (Invitrogen). Secvenţele utilizate pentru comparare au fost secvenţele franceze pEIAV21_983_Fr, pEIAV40_1028_Fr, pEIAV41_1028_Fr, pEIAV42_1029_Fr, pEIAV44_1029_Fr, pEIAV43_1029_Fr, pEIAV7_860_Fr, secvenţele japoneze japV70 (număr acces Genbank: AB008196), americane Wyoming uswy1 (AF170360) şi uswy12
Bolfă Pompei Florin – Rezumat teză de doctorat
26
(AF170372), din Texas ustx2 (AF170895) şi ustx6 (AF170899), din Idaho usid2 (AF172099) şi usid9 (AF172106), şi EIAVuk (AF016316).
REZULTATE ŞI DISCUŢII
Caracterizarea izolatelor VAIE obţinute
Caracterizarea genei gag Au fost analizate cele nouă clone obţinute, şi anume pEIAV107_1271, pEIAV108_1271,
pEIAV109_1271, pEIAV110_1282, pEIAV111_1282, pEIAV112_1282, pEIAV113_1282, pEIAV114_1282 şi pEIAV115_1287, comparativ cu clona de referinţă EIAV UK şi cu 7 clone franceze. S-a observat că există o asemănare genetică între clona 113 românească şi cele franceze. Majoritatea clonelor româneşti (excepţie 113), prezintă mutaţii punctiforme repartizate în toate secvenţele în urma alinierii, atât faţă de clona de referinţă EIAV UK cât şi faţă de izolatele franceze (figura 9), caracteristice în funcţie de originea geografică a clonei. Există o apropiere între izolatele româneşti şi cele franceze, care este mai importantă decât suşa de referinţă, faţă de care există numeroase poziţii divergente.
Pornind de la sângele periferic (PMBC – Peripheral Blood Mononuclear Cells) colectat de la 25 de cai (dintre care 34 seropozitive la AIE), s-a încercat identificare prezenţei genomului proviral integrat în celulele gazdă. Amplificarea prin PCR în cadrul genei gag, a demonstrat prezenţa VAIE în genomul monocitelor circulante de la 7 cai.
Analiza filogenetică pornind de la secvenţele nucleotidice obţinute, a permis stabilirea prezenţei a diferite subtipuri (subtipul 1 şi 2), prezentate în figura 8. Acestea sunt repartizate, după cum se observă, majoritatea, în funcţie de originea geografică a clonei VAIE secvenţializată şi analizată.
Figura 8. Arborele filogenetic pornind din porţiunea de genă gag secvenţializată (obţinut prin Neigbor joining, cu un bootstrap de 1000 repetări, ce permite testarea validităţii branşamentelor). Se desprind 2
subtipuri, corespunzând majoritatea originii geografice
Bolfă Pompei Florin – Rezumat teză de doctorat
27
Figura 2.5.4. Alinierea secvenţelor nucleotidice, 210 nucleotide de la 504 la 714 din gena gag, faţă de
pEIAV111, pEIAV112, pEIAV113, pEIAV114 şi pEIAV115. Mutaţiile caracteristice originii geografice sunt indicate prin câte o stea albastră.
În lucrarea de faţă s-a realizat pentru o caracterizare genitică a suşelor naturale de VAIE
circulante pe teritoriul tării noastre, pornind de la probe de sânge provenind de la cai infectaţi pe cale naturală. A fost detectată prin amplificare genică, porţini de genă gag din genomul proviral integrate în PMBC. În viitor se va încerca şi amplificarea genei env, precum şi obţinerea virusului sau a unor porţiuni din acesta pornind de la alte ţesuturi sau organe (plasmă, ţesut hepatic şi ţesut splenic).
Bolfă Pompei Florin – Rezumat teză de doctorat
28
Niciunul dintre caii infectaţi utilizaţi în experiment nu a prezentat semne clinice de AIE în momentul recoltării probelor biologice, ei aflându-se într-un stadiu asimptomatic de evoluţie a infecţiei.
Studiul de faţă, reprezintă prima caracterizare genetică a VAIE în România. S-a reuşit demonstrarea faptului că, clonele româneşti de virus, formează o populaţie clar distinctă faţă de alte virusuri deja cunoscute în Franţa sau în Statele-Unite. Dintre cele nouă clone studiate, porţiunea de genă gag analizată este foarte apropiată la opt dintre ele.
Bolfă Pompei Florin – Rezumat teză de doctorat
29
II.5.3. Rezultate comparative ale diferitelor tehnici de diagnostic în AIE
Testul practicat astăzi larg în activitatea de rutină a laboratoarelor de profil pentru confirmarea AIE, este testul Coggins. Acesta ajută la diagnosticarea formelor clinice, latente şi certificarea indemnităţii cabalinelor care se deplasează în interiorul statului sau în afara frontierelor lui.
Odată cu introducerea noilor metode de diagnostic serologic, aria de extindere a AIE a putut fi controlată mai eficient, cuprinzând totalul efectivelor de cabaline infectate (diagnosticate prin testul Coggins), ca şi altele cu un grad mai redus de anticorpi, evidenţiate prin teste de mare fineţe (ELISA, Western Blot sau PCR).
Dintre toate testele, cel mai fidel şi cu valoare diagnostică ridicată rămâne testul Coggins, care are însă dezavantajul că, la acelaşi animal nu apare constant pozitiv, ceea ce impune repetarea lui. Surprinderea animalului cu AIE într-o fază de eclipsă sub raportul prezenţei anticorpilor în cazul efectuării unui singur control, ar putea duce la interpretarea că, animalul este liber de AIE.
După testul Coggins, imediat ca fidelitate se situează examenul histopatologic. Acesta are avantajul în plus, că în multe cazuri confirmă diagnosticul de boală, chiar la caii reacţionaţi negativ serologic.
Directiva OIE, prevăzută în Manualul de standarde pentru testele de diagnostic şi vaccinuri, ediţia 2008, stipulează că testele autorizate pentru diagnosticul curent al AIE sunt testul ID (Coggins) şi testul ELISA, cu menţiunea că, serurile pozitive la testul ELISA trebuie să fie confirmate prin testul Coggins. Pentru confirmarea acestor teste este indicat testul immunoblot (OIE, 2009). Există câteva situaţii în care testul PCR poate folosit pentru detectarea infecţiei cu VAIE la cai, şi anume: în cazul unor rezultate contradictorii privind testele serologice; infecţie suspectă, dar rezultate serologice negative sau contestabile; teste complementare celor serologice pentru confirmarea rezultatelor pozitive; confirmarea infecţiei devreme, înainte ca anticorpii serici contra VAIE să se dezvolte; asigurarea că în cazul cailor care urmează să fie utilizaţi pentru antiser sau producţie de vaccin sau ca donatori de sânge sunt liberi de VAIE; confirmarea statusului unui mânz de la o iapă infectată (OIE, 2009).
Bolfă Pompei Florin – Rezumat teză de doctorat
30
CONCLUZII 1. Ig totale şi CIC evoluează interdependent în patogeneza AIE. Creşterea nivelurilor acestor indicatori
imunologici este asociată cu infecţiile virale recente, în care organismul nu a ajuns la un echilibru cu virusul infectant, respectiv cu înaintarea în vârstă, când capacitatea de clearance imunologic scade.
2. Serurile cailor AIE pozitivi prezintă un efect bactericid mai evident, la o concentraţie mai mare, posibil datorită intervenţiei unor componente activate de virus, respectiv a complementului.
3. Efectul stimulator iniţial al extractului de urzică asupra fagocitozei la caii AIE pozitivi tineri, recent infectaţi, urmat de un efect inhibitor, este explicabil printr-o posibilă înglobare rapidă a carbonului, urmată de instituirea unei fragilităţi membranale celulare.
4. Datorită absenţei unei stimulări antigenice in vivo, valorile indicilor de stimulare sunt mult mai scăzute la animalele sănătoase faţă de cele VAIE pozitive, independent de vârsta sau de durata de evoluţie a bolii.
5. Incapacitatea de răspuns la inductorii mitogeni folosiţi (la caii cu vârste de peste 5 ani, VAIE pozitivi), este accentuată de durata de evoluţie a bolii, în opoziţie cu tendinţa întâlnită la animalele tinere, probabil datorită diminuării reactivităţii imune cu înaintarea în vârstă.
6. Animalele tinere, recent infectate, dovedesc o reactivitate celulară crescută faţă de animalele vârstnice, datorită infecţiei, pe de-o parte şi imunoreactivităţii generale crescute la tineret, efectele imunoredresante ale extractelor folosite fiind mai evidente.
7. Modificările ce indică un dezechilibru oxidanţi/antioxidanţi, sunt mai evidente în cazul cailor infectaţi cu VAIE comparativ cu cei sănătoşi, atât la nivel enzimatic, non-enzimatic cât şi din punct de vedere al statusului antioxidant total.
8. Datorită prezenţei mai frecvente a virusului în circulaţie la caii recent infectaţi faţă de cei intraţi în stadiul asimptomatic, daunele oxidative sunt mai evidente la prima categorie, agravându-se odată cu înaintarea în vârstă, pe fondul reducerii capacităţii de apărare antioxidantă.
9. Modificările valorilor parametrilor hematologici în AIE se asociază cu episoadele febrile, iar lipsa virusului circulant sau a febrei în momentul recoltării confirmă perioada de „linişte” antigenică în momentul recoltării.
10. Studiul de faţă este primul care demonstrează existenţa unei corelaţii între o boală virală a cabalinelor şi stresul oxidativ.
11. Eventuala introducere în practica diagnosticului AIE a metodei cELISA, ar permite un diagnostic mai precoce şi mai sensibil, dar nu ar înlătura definitiv testul Coggins, recomandat în continuare de OIE pentru confirmarea rezultatelor pozitive.
12. Studiul genetic al clonelor VAIE româneşti, pornind de la o porţiune de genă gag, demonstrează faptul că se disting de alte virusuri deja cunoscute în SUA sau în Franţa.
13. Analiza secvenţelor nucleotidice obţinute, ne permite să afirmăm că suşele româneşti şi cele franceze sunt mult mai apropiate între ele faţă de suşa nord-americană de referinţă.
14. Repartizarea majorităţii izolatelor analizate în funcţie de originea lor geografică, sugerează circulaţia în Europa a unor suşe virale diferite de VAIE.
Bolfă Pompei Florin – Rezumat teză de doctorat
31
BIBLIOGRAFIE SELECTIVĂ
1. Andrei S., Pintea A., Catoi C., Sevastre B., Varga A., Baba A.I., Antioxidant enzymes activities, lipids and proteins oxidation – markers of oxidative stress in rats i.p. treated with asbestos, Bull. USAMV CN (2002) 59:598-603.
2. Aquaro S., Scopelliti F., Pollicita M., Perno C.F., Oxidative Stress and HIV Infection: Target Pathways for Novel Therapies? Future HIV Therapy (2008) 2:327-338.
3. Calamari L., Ferrari A., Bertin G., Effect of selenium source and dose on selenium status of mature horses, J. Anim. Sci. (2009) 87:167-178.
4. Castejón F., Trigo P., Muñoz A., Riber C., Uric acid responses to endurance racing and relationships with performance, plasma biochemistry and metabolic alterations, Equine Vet. J. Suppl. (2006) 36:70-73.
5. Craigo J.K., Montelaro R.C., EIAV Envelope Diversity: Shaping Viral Persistence and Encumbering Vaccine Efficacy, Curr. HIV Res. 2010 Feb 18, [Epub ahead of print].
6. Delguste C., de Moffarts B., Kirschvink N., Art T., Pincemail J., Defraigne J.O., Amory H., Lekeux P., Change in blood antioxidant status of horses moved from a stable following diagnosis of equine motor neuron disease, Can. Vet. J. (2007) 48:1165–1167.
7. Gil del Valle L., Gonzales I., Tarinas A., Alvarez A., Molina R., Tapanes R., Perez J., Evaluation of oxidative stress in AIDS pediatric patients, Acta Farm. Bonaerense (2004) 23:466-471.
8. Gil L., Martínez G., González I., Tarinas A., Alvarez A., Giuliani A., Molina R., Tápanes R., Pérez J., León O.S., Contribution to characterization of oxidative stress in HIV/AIDS patients, Pharm. Res. (2003) 47:217–224.
9. Jaime M, Ramirez-Boscá A., Soler A., Díez A., Carrión-Gutiérrez M.A., Díaz-Alperi J., Quintanilla-Ripoll E., Bernd A., Quintanilla-Almagr E., Increase with age of serum lipid peroxides: Implications for the prevention of atherosclerosis, Mechanisms of Ageing and Development (1998) 100:17-24.
10. Kirschvink N., de Moffarts B., Lekeux P., The oxidant/antioxidant equilibrium in horses. The Veterinary Journal (2008) 177: 178–191.
11. Kirschvink N., Fievez L., Bougnet V., Art T., Degand G., Smith N., Marlin D., Roberts C., Harris P., Lekeux P., Effect of nutritional antioxidant supplementation on systemic and pulmonary antioxidant status, airway inflammation and lung function in heaves-affected horses. Equine Vet. J. (2002) 34:705–712.
12. Leroux C., Cadore J.L., Montelaro R.C., Equine infectious anemia virus (EIAV): what has HIV’s country cousin got to tell us?, Vet. Res. (2004) 35:485-512.
13. Lippi G., Montagnana M., Franchini M., Favaloro E.J., Targher G., The paradoxical relationship between serum uric acid and cardiovascular disease, Clinica Chimica Acta (2008) 392:1-7.
14. Mandas A., Iorio E.L., Congiu M.G., Balestrieri C., Mereu A., Cau D., Dessì S., Curreli N., Oxidative Imbalance in HIV-1 Infected Patients Treated with Antiretroviral Therapy, J. Biomed. Biotechnol. (2009) doi: 10.1155/2009/749575.
15. Marañón G., Muñoz-Escassi B., Manley W., García C., Cayado P., de la Muela M.S., Olábarri B., León R., Vara E., The effect of methyl sulphonyl methane supplementation on biomarkers of oxidative stress in sport horses following jumping exercise, Acta Vet Scand. (2008) 50(1): 45.
16. Moffarts B., Kirschvink N., van Erck E., Art T., Pincemail J., Lekeux P., Oxidative stress in exercising horses: fact or fiction?, Equine and Comparative Exercise Physiology, (2005) 2: 253-261.
17. Pintea A., Andrei S., Bele C., Biochimie medical veterinară:lucrări practice, Ed. AcademicPres, Cluj-Napoca, 2008, pp. 72-74, 91-92, 108-109, 135-136.
18. Van Regenmortel M.H., Mayo M.A., Fauquet C.M., Maniloff J., Virus nomenclature: consensus versus chaos, Arch. Virol. (2000) 145:2227–2232.
19. Waring W.S., Convery A. , Mishra V., Shenkin A., Webb D.J., Maxwell S.R.J., Uric acid reduces exercise-induced oxidative stress in healthy adults, Clinical Science (2003) 105:425–430.
20. Williams C.A., Kronfeld D.S., Hess T.M., Waldron J.N., Saker K.E., Hoffman R.M., Harris P.A., Oxidative stress in horses in three 80-km races. Equine Nutr. Phys. Soc. Proc. (2003) 18:47- 2.
INVESTIGATION OF THE INFLUENCE OF EQUINE INFECTIOUS ANEMIA VIRUS ON IMMUNE COMPETENT CELL METABOLISM IN VITRO
Equine Infectious Anemia Virus (EIAV) is a lentivirus (Van Regenmortel et al., 2000), of the
Retrovirus family, with an almost worldwide distribution, infecting equids. It causes a persistent infection characterized by recurring febrile episodes associating viremia, fever, thrombocytopenia, and wasting symptoms (Leroux et al., 2004). EIAV and its associated disease have presented a considerable challenge to veterinary medicine worldwide ever and has been utilized as an animal model of HIV-1/AIDS research since the latters identification in the late 20th century (Craigo et al., 2010).
To this day, there are many studies that offer scientific proofs on the therapeutic benefits of plant extracts in the treatment or prophylaxis of different diseases, including some rare forms of cancer (Tajima and Aida, 2000, Vickers, 2002). The capacity to modulate different immunologic functions in the pathology of different diseases, but also their usage as adjutants in the preparation of vaccines has already been mentioned for some herbal plants (Florins et al., 2007, et al., 2007).
Numerous experiments with plant extracts or plant isolated active principles have proven that these can modulate the immune response by different means:
• Non-specifical stimulation of the immune system by activating serum complement and PMN neutrophils respectively macrophages, they stimulate phagocytosis (ex. Echinacea spp., Calendula officinalis, Thuja occidentalis etc.);
• Specific stimulation of the immune system, both towards cellular immune reactions and humoral ones, the result of these two directions being antibody synthesis (ex. Astragalus membracoeus, Ganoderma lucidum etc.);
• They directly influence the immune response by hormonal modulation (via adrenal glands).
The Aloe vera plant is known by many centuries for its benefic properties on health and the cosmetic industry. 2000 years ago, the Greeks regarded this plant as a universal cure. Certain components with a low molecular weight are capable of inhibiting the free species of radical oxygen secretion by the human activated neutrophils (Hart et al., 1990).
Calendula officinalis, has a vast applicability in medicine, with application in wound therapy, gastric and duodenal ulcer, cutaneous ulcerations (has a healing effect), inflammatory states. Phytopharmacological testing with different extracts of Calendula officinalis have also indicated that the plant has antiviral properties, anti-HIV and anti/geno-toxic. Different clinical studies have indicated the high efficiency of this extract in preventing acute dermatitis in patients with cancer that were treated by radiotherapy. In vitro, immune-modulating effects were noticed towards human lymphocytes (Jimenez-Medina şi col., 2006).
By far one of the most widely used immune stimulators of herbal origin, is the Echinacea extract. Most probably, the anti-infectious protection is ensured by the activating or enhancing properties of the host immune system, macrophage stimulation and cytokine synthesis being two important mechanisms. Even though Echinacea extracts have been used as immune stimulators, the scientific proof to justify the value of therapy with it is still controversial. Data concerning immune modulating effects induced in vitro by administration of this vegetal extract show that these mainly interest the structural elements of non-specific immunity (Bukovsky et al., 1993; Burger et al., 1997; Rininger et al., 2000; Vilma Jurkštienė et al., 2004; Mishima et al., 2004). Scientific studies are needed to explain certain aspects such as action mechanism, bioavailability, relative potency and power or the synergic effects between the bioactive components.
There are numerous studies that prove that the over 100 chemical components of nettles (Urtica dioica) are of help for the immune system, but also for the joint system of animals (Randall et al, 1999).
The immune system is capable of recognizing foreign substances (antigens), which determines a stimulation of this system and leads to antibody synthesis (humoral mediated immunity) or to a cellular immune response (cell mediated immunity). During the humoral immune response, the effector cells are the B-lymphocytes transformed into plasmocytes responsible with antibody synthesis. The cellular response involves T lymphocytes by whose means microorganism infected cells are eliminated, immune surveillance is performed (antitumoral defence) and allograft reaction takes place. NK (natural killer)
Bolfă Pompei Florin – Rezumat teză de doctorat
34
lymphocytes represent the third lymphocyte category; they do not need activation to perform their responsibilities.
The objective of this research was an in vitro quantification of EIAV infection effects on the
immune system of infected horses, of different ages and time periods from diagnosis, compared to the immune system of healthy horses. The monitoring of the immune system in horses took place on several directions. Circulating immune complexes (CIC), total immunoglobulines (Ig tot) but also the bactericidal capacity of serum in EIAV infected horses and in healthy horses were also monitored. The effect of EIAV infection on the cellular non-specific defense subsystem and on the functional capacity of the T lymphocyte subsystem was monitored. We wished to compel a picture as complete as possible when it comes to immune competent cell metabolism in vitro. We have also evaluated the influence of active products of different (Calendula officinalis, Echinacea angustifolia, Echinacea purpurea, Urtica dioica and Aloe vera) on the in vitro dynamics of immune effectors involved in the pathogenesis of EIAV infection.
The influence of EIAV infection on the total serum immunoglobulines levels was tested in
vitro through the dysproteinemia by zinc sulfate test. Antibodies towards viral surface glycoproteins of EIAV infected monocytes and macrophages are detectable in a time span of one month from infection, but they cannot mediate antibody dependent cellular cytolysis (ADCC), therefore indicating that ADCC is not involved in EIAV control in carrier (Tschetter et al., 1997). Unapparent carrier horses have increased total serum globulin and immunoglobulin and low albumin, respectively a low albumin/globulin ratio (Russell et al., 1998; Valpotic, 2000). In most horses, hyperimmuneglobulinemia with an increased IgG, IgG (T) and IgM in 60 days from the infection is noticeable (McGuire et al., 1971a).
Through the zinc sulfate dysproteinemia test, which is a relatively simple observation method of the albumin/globulin ratio misbalance, we appreciated modifications that appeared in total antiviral antibody titter by comparatively estimating the total immunoglobulin concentration, without detailing their functionality.
The obtained results have shown that in horses aged less than 5 years old and less than one year from EIAV infection diagnosis, serum Ig levels are significantly lower compared to all horsed aged over 5 years old (healthy and sick). The serum concentration of immunoglobulin in this category (group 1) is significantly lowered compared to EIA positive same aged horses, but seropositive for over 1 year.
There were no statistically significant differences between healthy and EIA positive, recently infected horses of the same age.
Different total Ig levels in the serum of tested horses can appear because of permanent fluctuation of Ig titers in EIA positive horses (more frequent and obvious in the first year of infection), depending on the viremia periods, viral replication and new virus variant selection.
In the case of total Ig, values are expected to be in coordination to serologic reactivity and immunoglobulins should be present in maximum concentrations in serologically intense positive animals. At the time-point of blood sampling for this experiment, there is no information on the anti EIAV antibody titer in the blood of researched horses (not-tested). This extra information would have allowed the realization of a correlation between the results of a Coggins test and total Ig levels in the tested horses. Reducing of seropositivity is also linked to the immunoglobulin concentration decrease. In the case of group 1, low total Ig levels may be explained by the short interval that has passed since the infection or even reduced reactivity in certain animals. The situation becomes inverted in the case of group 3, where at the moment of sampling, the animals had the highest serum titers of total Ig, as if one would expect in EIA positive horses with less than one year from diagnosis. Increased values as in group 2 may be attributed to the low number of animals that it contains (only two horses), which reduces its statistical value.
During the disease’s evolution, the fluctuation of the Ig titer can be explained by the intervention of certain protective Ig’s, realization of antigen-antibody coupling, part of them being neutralizing and part of them being immune infectious circulating complexes. The in vivo variability of the virus does a favor to these aspects, as a consequence of the forming of new variants after the primary infection. EIAV persists in the organism and the circulatory stream, also having periods of absence that are variable as duration, which are necessary to the synthesis of new variants, which will, in their turn,
Bolfă Pompei Florin – Rezumat teză de doctorat
35
generate antibody synthesis. This viral „perpetuum mobile” (Boldizsar, 2001) represents the fundament of fluctuant antibody genesis but also of the concentration alterations in total immunoglobulins.
When it comes to the healthy animals, the intermediary Ig concentration does not signify in this case the presence of anti EIAV antibodies, but the presence of antibodies that have a different antigenic specificity. We have not applied any testing to identify this specificity in the present paper. In the case of EIA positive horse groups that had total Ig levels higher than the ones in the witness group, we assumed that this increase might happen due to the presence of antiviral antibodies, but one cannot exclude the presence of other antibodies with a different antigenic specificity (figure 1).
circulating immune complexes The influence of the EIA virus infection on the immune circulating complexes levels was
tested by polyethylene glycol testing. The periodical viral replication has the permanent viral presence in raised titers, fact that leads to the forming of circulating immune complexes (CIC) and accumulation of sensitive lymphocytes (Hendersen et al., 1987; McGuire et al., 1972). Circulating immune complexes, due to their large dimensions, can be precipitated with polymers that have a high molecular weight, such as polyethylene glycol, even in low concentrations of the latter.
After analyzing the obtained results (figure 2), one can observe the appearance of disaccording values in EIA positive horses, which might be due to different CIC formation (excess antibodies or excess antigens). This fact is tightly linked to the disease’s evolution particularities in each individual. An explanation of the lower serum values of CIC in young, recently infected horses is the accumulation of these complexes at tissue level. It is also possible that in this case, the differences are given by different maintenance conditions, environment microbial load but also the variability of the infecting viral strand. It is also less probable that technical errors have occurred during sample processing, because serum was tested at the same time and in the same working conditions.
The forming and precipitation of immune complexes represent two interconnected phenomenons and they are determined by the quantitative relation of antigens and antibodies during a immune response. There is a consensus in regard of the aged organisms, in which normally there are CIC and certain tissue deposits with no lesions and no immune complex disease state (Berceanu and Păunescu, 1981). In the pathology of immune complexes, there are quantitative and qualitative areas of crossing between normal and pathological. In the case of EIA, since the great variability of EIAV is known, the antibodies, by coupling with the viral variant that determined their synthesis in the circulatory stream of horses during fever periods, practically the virus finds itself permanently as infecting immune complexes (McGuire et al., 1972). Immune complexes can intensify the ability of the virus to enter the host cell (monocytes macrophages), being involved in the appearance and maintenance of all EIA alterations..
EIA positive animals that are aged over 5 years old, having less than one year from diagnosis present significantly lower levels of CIC compared to younger seropositive animals, recently diagnosed. In seropositive animals, aged over 5 years, immune complex levels are significantly lower compared to healthy horses aged over 5 years old.
The differences between circulating immune complexes registered between two groups may be attributed to the evolutive particularities of the viral infection and the differences of infectious pressure, due to the antigenic variability of the involved strain. The CIC concentrations in healthy animals are not
Bolfă Pompei Florin – Rezumat teză de doctorat
36
dependent to the investigated age category. Their light increase due to ageing is a consequence of the deficient elimination of immune complexes without determining the disease through their intervention.
We noticed a great similarity between the results obtained in the case of total Ig dosage and CIC dosage (as tendencies), which might confirms the fact that the two evolve synchronous in the blood of the sick animals, but also in the blood of healthy ones, and that they depend to the animal’s age and also to the time that has passed since the diagnosis of the infection.
The next test applied to monitor the immune response in this research was represented by the evaluation of the bactericidal capacity of EIAV infected horse serum. This represents one of the tests which, by serial dilutions, determines the serum titer (and the antimicrobial components of the serum) which presents an efficient bactericidal activity. The investigation of the in vitro bactericide capacity permits the appreciation of the in vivo reactivity of organisms with bacterial infections.
Capacitatea bactericida a serului (gram-pozitivi)- Serum bactericid capacity (gram-positive)
Figure 4. Boxplot diagram for the bactericidal potential of serum against the gram-positive bacteria (1 = lot 1, dilution 1/10; 2 = lot 1, dilution 1/20; 3 = lot 1, dilution 1/40; 4 = lot 1, dilution 1/80; 5 = lot 3,
dilution 1/10, etc.)
Bolfă Pompei Florin – Rezumat teză de doctorat
37
The experiments realized on other animal models by Hansen and Hirsh have indicated a low susceptibility level in the case of avian strains of Pasteurella multocida. The authors have correlated total resistance to serum to the presence of pathogenicity factors, since there have been evidentiated capsular structures for these strains (Diallo and Frost, 2000; Morishita et al., 1990). Similar observations refer to the lack o sensitivity compared to normal human serums in the case of clinical strains of: Streptococcus pneumoniae, Staphylococcus aureus, Mannheimia haemolytica (Trakulsombon et al., 1989).
The results we obtained (figure 3 and 4) after the evaluation of the bactericidal capacity in vitro of serum obtained from healthy respectively EIAV infected horses, indicated a bactericidal activity both against gram positive and gram negative germs. The optimal dilution, in which the maximum bactericidal effect is obtained, was, in most cases, the minimal dilution.
The bactericidal activity of serum against the two strains of the experiment is more obvious in serum coming from EIA positive horses. The effect is more obvious in the case of testing against gram positives. In gram positives, the serum that comes from healthy animals of both groups proves a decreased inhibitor effect on the development of the tested bacteria compared to serum from seropositive animals.
In vitro, the minimal bactericidal effect towards both tested germs appears in the case of healthy animals aged maximum 5 years old, and the maximal one in the case of young, recently infected horses (for Pseudomonas aeruginosa), or in the case of horses aged over 5 years old infected with EIAV since more than one year (in the case of Staphylococcus aureus).
The influence of the EIA virus infection on the non-specific cell defense subsystem was
evaluated by the phagocytosis test. The phagocytic activity can be modified in a positive (stimulative) or negative (inhibitive) way by certain vegetal extracts.
The results we obtained register a tendency according to which the phagocytic activity is enhanced in the second monitored period, proving a delayed in vitro activation of phagocytic blood cells. In the case of young, recently infected horses with EIAV, the reactive capacity of phagocytes is significantly raised in the first stage, followed by an inhabitation in the second stage (p<0,05).
In healthy horses, the alcoholic extract of Urtica dioica proves itself to be more of a inhibitor of phagocytosis in the first stage, the effect being more intense in the age category of 1-5 years (p<0,05), followed by a stimulative effect in the second stage.
In young, infecte horses, aged up to 5 years, having less then one year from the discovery of the disease, the alcoholic nettle extract demonstrates a stimulatory effect of phagocytosis in the first stage, but after this, the effect is inhibitory, which could be explained by a possible fast recruitment followed by high membrane frailty that leads to cell destruction (p<0,05).
The alcoholic nettle extract seems to have an inhibitory effect on first stage phagocytosis in sick horses aged over 5 years old, indifferent of the interval that has passed from the diagnosis and in the ones with ages between 1 and 5 years where more than 1 year has passed since the diagnosis, followed in the second stage by a slight stimulative effect.
In EIAV infected horses aged over 5 years old, and in more than one year from diagnosis, in the case of the untreated and treated with alcohol groups, phagocytic activity is significantly enhanced in the second stage (the 45-60 minutes interval) compared to the first interval (0-15 minutes), which suggests the fact that in this category of animals, the reactive capacity of the phagocytes is still kept (p<0,05).
The in vitro nettle extract treatment with the purpose of quantifying phagocytic activity follows a similar model to spontaneous phagocytosis. By looking at the nettle extract effects on cells isolated from healthy horses, during the first reading stage, one might state that it acts rather as an inhibitor. During the second stage, the effect of nettle extract is predominantly stimulative.
Comparing our results with those of Boldizsar (mainly the first monitored interval- respectively the first 45 minutes of incubation), one notices some common aspects, but also certain differences regarding phagocytic activity. Therefore, similar to Boldizsar’s results (2001), it was noticed that in EIAV positive horses, phagocytic activity drops with ageing and by the amount of time that passes from diagnosis. On the other hand, the more time passes from sample incubation, phagocytic activity seems to get back to normal (during the 45-60 minute interval).
Bolfă Pompei Florin – Rezumat teză de doctorat
38
Appreciation of the EIA virus infection effect on the functional capacity of the T lymphatic subsystem was done by performing the in vitro lymphoblastic transformation test, the whole blood variant.
By comparative evaluation of the lymphoblastic transformation test in horses (figure 5), several differences occur between animal lots. Therefore, in the case of the healthy animals, indifferent of age, the stimulation indices values maintain themselves tin all experimental variants to approximately the same level (there are no statistic differences between the eight experimental variants that were applied). Also, in seronegative animals, indifferent of age category, one registers much lower values of the stimulation indices compared to seropositive animals, independent of age or evolution duration of the infection, probably because the absence of antigenic stimulation in vitro. Also, in the EIA negative equine group (with the exception of the untreated witness group) and in the EIA positive but with more than one year from diagnosis group, the response capacity of lymphocytes to several stimuli of the experiment drops with ageing, confirming the existence of a balance between virus-the animal’s immune system in the asymptomatic evolution stage of the disease.
1 2 3 4 5 6 7 8
-20
020
4060
8010
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 1 - BTT Lot 1
1 2 3 4 5 6 7 8
5560
6570
7580
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 2 - BTT Lot 2
1 2 3 4 5 6 7 8
-20
020
4060
8010
012
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 3 - BTT Lot 3
1 2 3 4 5 6 7 8
-50
050
100
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 4 - BTT Lot 4
1 2 3 4 5 6 7 8
-400
-300
-200
-100
010
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 5 - BTT Lot 5
1 2 3 4 5 6 7 8
-600
-400
-200
020
0
Variantele de lucru - Working variants
Indi
cii d
e st
imul
are
% -
Stim
ulat
ion
inde
x %
TTB Lot 6 - BTT Lot 6
Figure 5 Boxplot diagrams for the stimulation indexes of the six experimental variants, according to the
working variants
In seropositive horses with less than one year from diagnosis, the situation is inversed: the
response capacity of lymphocytes to several stimuli in the experiment is enhanced in the older animals (with the exception of Calendula officinalis, Echinacea angustifolia and Echinacea purpurea).
In animals aged over 5 years old and seropositive, the response incapacity to the mitogen inductors used is accentuated by the evolution duration of the disease (and because of the lack of circulating virus in the sick animals’ blood, as opposed to the tendency seen in young animals (up to 5 years old), where the response to certain mitogens is enhanced by the more time it passes from the diagnosis.
Bolfă Pompei Florin – Rezumat teză de doctorat
39
In horses aged up to 5 years old and that have less than one year since EIAV infection diagnosis, there are no statistically significant differences in the stimulation indices between the 8 working variants (the cell immune reactivity is not altered, just as in healthy horses).
When it comes to horses aged up to 5 years, with more than one year from diagnosis, among the utilized immune modulators, Aloe vera (p<0,05) phytohemaglutinine, and Calendula (0,1>p>0,05) produce a significantly increased stimulation compared to Echinacea purpurea (with a rather inhibitory effect).
In the animal group of over 5 year olds, having more than one year from diagnosis, the effect of Echinacea purpurea is an inhibitory one, being lower (0,1>p>0,05) compared to the following variants: witness, alcohol witness and PHA M.
The inhibitory effect of Echinacea purpurea extract is visible in the group of horses aged over 5 years old, with less than one year from diagnosis (p<0,05) compared to the witness group, the alcohol-group, PHA M, nettle and aloe, but also compared to Calendula (0,1>p>0,05).
The immune recovery effects of the used extracts are registered preferentially in young animals; these effects are registered in horses aged up to 5 years, with less than one year from the diagnosis, after cell treatment with Calendula officinalis, Echinacea angustifolia and Echinacea purpurea; in the same age category but having more than one year from diagnosis of treatment with Aloe vera, Urtica dioica and Calendula officinalis; in horses aged more than 5 years, having more than one year from diagnosis, only the Echinacea angustifolia extract has registered transformation indices that were larger than the two witnesses (in all the aforementioned situations, p>0,05).
In the case of the two Echinacea species we used during this experiment, even though er are talking of plants that are very similar taxonomically, being a part of the same genre, their in vitro effects are different; this fact demonstrating that there are different active principles, for which all organisms we tested do have more or less receptors.
In a synthesis study focused on different experimental studies, Melchart et al. (1994) have evidentiated au the modulating effects of the immune parameters of Echinacea angustifolia given that further testing should also take place. Focusing on the same directions, Bukovski et al. (1993), Burger et al. (1997), Mishima et al. (2004), Rininger et al. (2000), have evidentiated the fact that plant extracts of Echinacea species are capable of inducing an increased blastic stimulation level. This tendency confirms itself in this thesis. In addition, it is proven that Echinacea purpurea does not have the same active principles and does not have the same benefic effects in vitro.
This thesis we partially confirm Boldizsar’s results but we also complete them from a immune cellular reactivity point of view. Therefore, the tendency of a decrease of the cellular immune response by ageing is confirmed. The values of the stimulation indices do not maintain themselves in the same limits in the sick animals compared to the healthy ones, since they are dropping increasingly in the latter, probably due to the lack of in vivo antigenic stimulation. The alcoholic Echinacea purpurea, Urtica dioica and Aloe vera extra weak immune stimulating effect and only in certain categories of infected horses, while the Calendula officinalis and the Echinacea angustifolia extract seem to have a more noticeable stimulative extract. None of the utilized extracts have expressed a immune stimulative or immune modulation effects in healthy animals, not proving themselves useful in the resetting of cellular reactivity in the applied doses; therefore further testing is required.
After the correlative evaluation of the immune results presented, it is confirmed that EIA is a systemic disease, in which the immune system of the infected horses is affected in different degrees determining numerous implications in the disease’s pathogenesis. The components of the specific and nonspecific immune system, cellular and humoral, from healthy and EIAV infected horses, of different ages and in different evolution staged of the disease. The cumulation of the obtained results allows the contouring of a immune pathogenesis picture, which is not by far complete.
Bolfă Pompei Florin – Rezumat teză de doctorat
40
EVALUATION OF THE OXIDANT-ANTIOXIDANT EQUILIBRIUM IN HORSES INFECTED WITH EQUINE INFECTIOUS ANEMIA VIRUS
In an excellent rewiev, Kirschvink et al. (2008) showed that oxidant/antioxidant equilibrium in
horses has been connected over the past years with many physiological and pathological conditions in horse, with some studies focusing on antioxidant supplementation trials. Thus, some physiological situations where the oxidant/antioxidant equilibrium is imbalanced, regard equine reproduction as well as exercise and training Some situations from equine pathology where the oxidant/antioxidant equilibrium is altered, include lower airway diseases, such as recurrent airway obstruction, obstructive pulmonary disease, and possibly exercise induced pulmonary hemorrhage; neurological disorders such as equine dysautonomia and motor neuron disease; muscle disorders like white muscle disease, steatitis, also known as yellow fat disease, seasonal pasture myopaty reported in USA, atypical myopaty reported in Europe, possibly also exertional rhabdomyolysis; perfusion related disorders like equine post-anaesthetic myositis, ischemia and reperfusion process during colic; laminitis; joint diseases as degenerative joint disease as well as osteoarthritis. The majority of these studies are focused on race horses, unlike our investigation which is focused on horses used for farming and transportation.
Oxidative stress is thought to play an important role in the progression of HIV infection, another lentiviral disease (Gil del Valle et al., 2004; Mäenpää et al., 1988). It has been observed that perturbations in antioxidant defense systems, and consequently redox imbalance, are present in many tissues of HIV-infected patients. Moreover, there is clear evidence that oxidative stress may contribute to several aspects of HIV disease, including viral replication, inflammatory response and decreased immune cell proliferation (Aquaro et al., 2008).
Once infected with the EIAV, horses remain viraemic carriers for the rest of their life. Most of the animals that survive the acute or chronic stage enter in the asymptomatic stage of disease (Bolfă et al., 2008; Leroux et al., 2004). So far, in equine medicine, no study has correlated the oxidant/antioxidant equilibrium with an infectious disease, respectively a viral disease.
In this context, the aim of the present study was to investigate the oxidant/antioxidant equilibrium in EIAV infected horses, compared to healthy ones, on different age groups and at different time points following the initial diagnose. The research aimed to monitor both enzymatic and non-enzymatic blood antioxidant system, as well as serum total antioxidant capacity.
2. MATERIALS AND METHODS
2.1. Subjects and blood collection Blood and serum samples from 96 horses were collected over a period of 2 years (January 2005
– February 2007) in the area of Bistrita-Nasaud and Cluj County, Romania. The animals (52 females and 44 males), with ages between 1 and 25 years, were divided into six experimental groups, according to disease status, age and period after diagnose, as shown in table 1.
Table 1.
Experimental groups of horses in the experiment
Group AGID test Age (years) Interval after positive diagnose
Number of animals
EIAV 1 + < 5 < 1 year 11
EIAV 2 + < 5 > 1 year 3
EIAV 3 + > 5 < 1 year 36
EIAV 4 + > 5 > 1 year 21
Control 5 – < 5 - 9
Control 6 – > 5 - 16
Bolfă Pompei Florin – Rezumat teză de doctorat
41
All animals were clinically healthy at the moment of sampling. The horses are used for agricultural works and blood samples were collected always mornings, before the workday began, from the jugular vein. Any hemolysed samples were discarded. Blood samples were analyzed the same day and serum samples were stored at -20 °C until analysis. All horses were previously tested for EIAV seropositivity or seronegativity with the reference agar gel immunodiffusion test (AGID) at the Sanitary Veterinarian Laboratory and for Animal Health Bistrita, respectively Cluj-Napoca (EIA Kit No. 119, Pasteur Institute, Bucharest, Romania).
2.2. Biochemical measurements
2.2.1. Total antioxidant status (TAS)
For the quantification of TAS from serum samples we used Randox Ltd. (Diamond Road, Crumlin, U.K.) Kit Cat. No. NX2331. Briefly, ABTS (2,2’-azino-di-(3-ethylbenzthiazoline sulfonate)) is incubated with metmyoglobin and hydrogen peroxide (H2O2) to produce the radical cation ABTS®+. This has a relatively stable blue-green colour that can be measured at 600 nm. Antioxidants in the added sample cause a suppression of this colour production, which is proportional to their concentration (Randox, 1996). Values were expressed as mM/L.
2.2.2. Superoxide dismutase (SOD)
Evaluation of SOD activity from whole blood was achieved using Randox Ltd. (Diamond Road, Crumlin, U.K.) Kit Cat. No. SD125. Briefly, the method employs xanthine and xanthine oxidase to generate superoxide radicals, which reacts with 2-(4-iodophenyl)-3-(4-nitrophenol)-5-phenyltretazolium chloride (INT) to form a red formazan dye. The superoxide dismutase activity is then measured by the inhibition degree of this reaction (Randox, 1996). Values were expressed as U/ml.
2.2.3. Catalase (Cat) Catalase activity from whole blood was evaluated using the Bach-Zubkova permanganometric
method. A quantity of H2O2 is actually dosed before and after the enzyme action. The activity of catalase is determined by calculating the difference between the ammount of potassium permanganate used at titration. This is represented by the catalasic number expressed in ml KMnO4, which represents the quantity of hydrogen peroxide in miligrams decomposed by 0.1 ml of blood (Pintea et al., 2008).
2.2.4. Glutathione peroxidase (GPx)
Evaluation of GPx activity from whole blood was determined using Randox Ltd. (Diamond Road, Crumlin, U.K.) Kit Cat. No. RS504. Briefly, GPx catalyses the oxidation of GSH by cumene hydroperoxide. In the presence of glutathione reductase and NADPH the oxidized glutathione is immediately converted to the reduced form with a concomitant oxidation of NADPH to NADP+. The decrease in absorbance at 340 nm is then measured (Randox, 1996). Values were expressed as U/g Hb.
2.2.5. Malondialdehyde concentration (MDA)
Determination of lipid peroxidation from blood sera was assessed by quantification of MDA concentration. Briefly, MDA has the property to react with thiobarbituric acid forming a colored reaction product, which can be dosed spectrophotometrically at 535 nm. Concentrations of MDA were calculated
Bolfă Pompei Florin – Rezumat teză de doctorat
42
using the corresponding standard curve, using acid hydrolysis of tetrametoxipropanol (TMP) and values were expressed in nM/mL (Andrei et al., 2002).
2.2.6. Total lipids (TL)
Total lipids from blood sera were measured using a manual method, also known as sulfo-phospho-vanillin method. Fatty acids from serum lipids are liberated and sulfonated through treatment with concentrated sulphuric acid, under warm conditions. Afterwards, through vanillin and phosphoric acid reaction, a violet colour compound is formed (sulfo-phospho-vanillin complex). For the calculation, standard is considered 1000 mg lipids/100 mL. Absorbance of standard and samples are then read compared to a blank, at 525 nm. Values were expressed as mg/dL (Pintea et al., 2008).
2.2.7. Uric acid (UA)
For the quantification of uric acid from blood sera, we used a Hospitex method (Hospitex Diagnostics, Firenze, Italy). Uric acid in sample is oxidized to allantoin in presence of the enzyme uricase and H2O2 is generated. The H2O2 reacts with TOOS (N-etil-N-hidroxisulfopropil-3-metilalanin) and 4-aminoantipyrine in the presence of peroxidase to form a violet dye. The intensity of colour formed is proportional to the uric acid concentration and can be measured spectrophotometrically at 546 nm. Values were expressed as mg/dL (Pintea et al., 2008).
2.2.8. Total carotenoids (Car)
Quantification of total carotenoids, especially β-carotene, from blood sera in petroleum ether was achieved by a spectrophotometrical method, reading the extinction at 448 nm. The amount of carotenoid pigments from the samples is expressed in µg/dL (Pintea et al., 2008).
Chemicals were purchased from Merck Chemicals (MERCK Romania SRL, Bucharest,
Romania).
2.3. Statistical analysis
Data were expressed as mean ± SD. The results were analyzed for normality of distribution using the Shapiro–Wilk normality test. The majority of data sets were found not to be normally distributed. Mean comparison was done by the Kruskal-Wallis test followed by a Mann Whitney test; a confidence level of 95% (p < 0.05) was considered significant. To assess the influence of EIAV on the oxidant-antioxidant equilibrium, the levels of each parameter from seropositive horses (infected <1 year or >1 year before the experiment) was compared to those of the healthy animals with same age. Different age groups of EIAV positive and negative horses were as well compared, to determine the influence of age on the different parameters investigated. The R free software package was used for all statistical analyses (www.r-project.org).
3. RESULTS
As shown in table 2, only in the case of SOD, TL and UA there is a significant difference
between the six groups (p < 0.05, Kruskal-Walis test).
Bolfă Pompei Florin – Rezumat teză de doctorat
43
Table 2. Comparison of oxidative stress markers in whole blood and in serum of EIAV positive and negative
horses EIAV 1 EIAV 2 EIAV 3 EIAV 4 Control 5 Control 6
Data are expressed as mean ± SD * p<0.05, Kruskal-Wallis test TAS values are higher (less antioxidants in the blood) in most of the EIAV infected animals,
compared to the control groups. Group 2, 3 and 4 had higher values for TAS compared to both control groups. A significantly increase in TAS was only found for group 3 compared to group 6 (p < 0.05). In other words, healthy horses over 5 years of age have a better antioxidant status than those of same ages, recently infected with EIAV.
Most of the antioxidant enzymes had decreased levels in infected horses, compared to the corresponding control groups (figure 6). SOD activity was significantly lower in group 1 compared to group 3 and 6 (p < 0.05). Thus, young horses, recently infected with EIAV, showed decreased levels of the enzyme compared to older (over 5 years) healthy or recently infected horses. Similarly, the enzyme’s activity in group 4 (EIAV positive equines, aged more than 5 years, infected longer than 1 year ago) had lower levels (p < 0.05) than that of group 3 and 6, representing horses of same age, either recently infected or healthy. The infection with EIAV has no influence on the levels of blood catalase activity (Cat). Although GPx activity was decreased in all infected animals, compared to the healthy horses, this was found to be significant (p < 0.05) only in the blood of animals from group 3 compared to the control groups 5 and 6. Thus, in older horses, recently infected with EIAV, GPx significantly reduces its antioxidant activity compared to healthy animals.
Lipid peroxidation represents probably the most extensively investigated free radical–induced process (Sodergreen, 2000). Serum levels of malonyldialdehyde (MDA) in EIAV recently infected horses (EIAV 1 and EIAV 3) are higher than controls and animals in the asymptomatic stage of disease. However, the recently infected older horses (group 3) present the most intense lipid peroxidation, which is significantly (p < 0.05) higher compared to healthy horses from control group 5 and asymptomatic carriers from EIAV group 4. A decreased level of TL (p < 0.05) in recently infected young horses (EIAV 1) compared to all older animals (group 3, 4 and 6), can be correlated with the consumption of lipids in the process of peroxidation. There is a significant (p < 0.05) decrease of TL in group 4 compared to group 3.
Bolfă Pompei Florin – Rezumat teză de doctorat
44
1 2 3 4 5 6
56
78
910
mM
/LTAS
1 2 3 4 5 6
5015
025
0
u/m
L
SOD
1 2 3 4 5 6
26
1014
ml K
MnO
4
Cat
1 2 3 4 5 6
5010
015
020
0
u/g
Hb
GPx
1 2 3 4 5 6
040
80
nM/m
L
MDA
1 2 3 4 5 6
100
300
500
700
mg/
dL
TL
1 2 3 4 5 6
0.0
1.0
2.0
3.0
mg/
dL
UA
1 2 3 4 5 6
510
1520
ug/d
L
Car
Figure 6: Total antioxidant status (TAS), Malondialdehyde concentration (MDA), Total lipids (TL), Total carotenoids (Car), Uric acid (UA) from blood sera and Superoxide dismutase (SOD), Catalase
(Cat), Glutathione peroxidase (GPx) from whole blood of EIAV positive and control horses. Data are presented as boxplots, with 25th, 50th (median) and 75th percentiles, and the outliers. 1, 2, 3, 4, 5 and 6
represent the experimental groups. There is an apparent paradox between the protective and toxic effect of uric acid, clinically
based on the fact that antioxidant products can become pro-oxidant in certain situations, especially when they are present in the blood at levels higher than normal (Lippi et al., 2008). In the present study, the levels of UA in all EIAV seropozitive animals are higher than those of the control groups. After the infected horses enter in the asymptomatic stage of disease, UA levels tend to decrease. Only in horses from group 3, UA values are significantly (p < 0.05) increased compared to both control groups.
Carotenoids levels did not differ in EIAV infected and non-infected horses, independently of age or interval after infection.
4. DISCUSSION
It has been widely reported that HIV infection causes oxidative stress (Gil et al., 2003; Mandas
et al., 2009). Both our EIAV infected and control horses showed significantly higher (p < 0.05) levels of TAS, compared to the reference values offered by the kit, on a human population (1.3 - 1.77 mM/L). This means that the studied equine populations have less antioxidants in the blood, making them more vulnerable to oxidative damage. Except for the young horses, recently infected with EIAV, the other three
Bolfă Pompei Florin – Rezumat teză de doctorat
45
groups of infected animals showed increased values compared to all healthy animals. The cause for this could be the viral infection; the effect is more obvious (p < 0.05) in recently infected older horses compared to uninfected animals of the same age.
Similar to TAS, most of the values for SOD activity were lower than the reference values in humans offered by the kit producer (154 - 240 u/mL). The decrease in SOD activity in EIAV recently infected horses compared to the healthy animals of same age, is not statistically significant. The most severe decrease of the enzyme’s activity was in case of young, recently infected horses (EIAV 1) and older, longer infected animals (EIAV 4). The virus may be responsible for the weak activity of the antioxidant enzyme in the first case, whereas, similar to humans (Guemori et al., 1991), the decrease in SOD activity for the EIAV 4 group may be due to both aging and the viral infection.
Catalase activity in the blood of EIAV infected horses, has the tendency to decrease with aging, as it happens in case of the human population (Guemori et al., 1991). In a study on an EIAV experimentally infected horse, Fujino and Negishi (1957), proved that during febrile episodes the activities of catalase along with alkaline phosphatase have the tendency to decrease, while lipase and amylase have the tendency to increase. However, between our experimental groups, the variation of the enzyme’s activity, showed no statistical differences. In horses recently infected with EIAV, the activity of two antioxidant enzymes investigated (catalase and GPx), was decreased compared to the controls of same age suggesting that infection with EIAV imbalances the oxidant/antioxidant equilibrium in recently infected horses, aged over 5 years.
In a study on horses, Delguste et al. (2007) suggests that an increase of GPx activity as a response to an oxidative stress state may exist, but is limited in time. Seasonal changes, affecting nutrition composition, could have some effect on GPx activity (Erdélyi et al., 1999). GPx activity is known to correlates with selenium status in the organism, as the enzyme contains organic Selenium (Se) as selenocisteine in its active core. Low GPx values might be an indicator of a Se deficit in the organism. Nevertheless, in horses, the quantity and source of Se in food, doesn’t influence the activity of GPx 1 (Calamari et al., 2009). In our experiment, GPx activity in the virally infected groups was decreased compared to both control groups. Only in recently infected older horses (EIAV 3) this decrease was significant (p < 0.05) compared to healthy controls of all ages. The values for blood GPx activity in horses from the current study were higher than the kit’s reference values on a human population (27.5 – 73.6 U/g Hb).
In sport horses following jumping exercise, there is an increase of MDA, nitric oxide (NO) and carbon monoxide (CO), accompanied by a decrease in reduced glutathione and antioxidant enzymes, inducing harmful effects on horses, probably due to an increase in oxidative damage and proinflammatory molecules (Marañón et al., 2008). The decrease in activity (Delguste et al., 2007) of the main antioxidant enzymes (SOD, Cat and GPx), and an increase of lipid peroxidation level (Jaime et al., 1998), with aging, in humans, are indicators of an oxidative stress state. MDA concentration in our study, has similar tendency as in humans, respectively increases at older animals, both healthy and EIAV positive. Similar to HIV infection in humans, (Favier et al., 1994), horses infected with EIAV, have higher lipid peroxidation levels in the first year after diagnose, than later when they enter in the asymptomatic phase of disease. This could be consecutive to a more intense overproduction of oxygen free radicals by more viable polymorphonuclear (PMN) at the asymptomatic stage. The free radicals production and lipid peroxidation seem secondary to a direct induction by the virus of PMN stimulation and cytokines secretion (Favier et al., 1994). In humans, the increase in MDA concentration is proved to play a role in stimulating HIV replication (Allard et al., 1998).
The mean levels of UA in our study are in the normal range for horses (0 - 1.8 mg/dL,( Pintea et al., 2008)). All EIAV infected animals showed higher serum values than those from healthy animals. Animals aged over 5 years, both infected and uninfected had slightly elevated values, suggesting a slightly increase with age of UA. Recently infected horses have increased UA values than those in the asymptomatic phase of disease, with no statistical significance. Only recently infected horses aged over 5 years have significantly (p < 0.05) increased UA levels compared to both control groups. Waring et al. (Waring et al., 2003) suggests that increased concentrations of UA could offer a protective effect in vivo against free radicals, and that UA could have a biologic importance in acute oxidative stress. In horses following prolonged exercise, UA level was increased, especially in fast horses with raised metabolism (Calamari et al., 2009). On the other hand, exercise in horses preceded by antioxidant suplimentation
Bolfă Pompei Florin – Rezumat teză de doctorat
46
(vitamin E and C) showed reduced concentrations of UA (Kirschvink et al., 2002). It is difficult to estimate if the increase in UA levels is based on its antioxidant function or if these levels are responsible for installation of oxidative stress. Lippi et al. (2008) proposed a prooxidant action for UA in serum for levels above the normal range. Further extensive studies are needed for a better clarification of this aspect.
Similar to UA, in EIAV infected horses, serum carotenoids are present at a slightly more elevated level than in healthy horse serum, but within the reference values literature offers (1 - 30 µg/dL, depending on pasture (Pintea et al., 2008)). Both in infected and healthy animals, serum carotenoids have the tendency to decrease with ageing. A higher serum carotenoids level in EIAV infected horses, might have two explanations: one being a higher nutritional intake, the second would consist in an increase in free radicals in the EIAV infected groups and the serum non-enzymatic antioxidants (amongst which are also carotenoids) appear in a larger quantity to neutralize them. This increased ROS level might be a consequence of the presence of the virus in the blood stream. On the other hand, it is suggested that the younger animals have a better oxidation protection system; these animals will adapt better to oxidative stress states. Blood carotenoid concentration depends on the diet, the season and the animal’s general health state (Greiwe-Crandellet al., 1995; Greiwe-Crandell et al., 1997; Mäenpää et al., 1988; McDowell et al., 1989). β-Carotene is at the same time the main pro-vitamin A, and decrease of its plasma concentration leads to a decrease in the vitamin A. Decrease in retinol concentration has adverse effects on growth and reproductive capacity in horses, it also determines an increase in infectious disease risk (Donogue et al., 1981; McDowell et al., 1995).
Prevention of oxidative stress and modulation of oxidant/antioxidant equilibrium looms as a new target in equine medicine. The instrument for that is nutrition modulation of equines, depending on the effort and antioxidant requirements (Kirschvink et al., 2008; Moffarts et al., 2005; Williams et al., 2003).
In this study we proved for the first time the connection between a viral infection in equines and oxidative stress. Changes indicating oxidant/antioxidant disequilibrium, in both enzymatic and non-enzymatic system, as well as in total antioxidant status are more obvious in all EIAV infected horses. Recently infected animals, over 5 years, represent the most vulnerable category to oxidative damage, followed by recently infected younger horses. Oxidative stress in EIAV asymptomatic horses is present, but milder compared to recently infected equines. This suggests that the circulating virus plays an important role in the immediate or cumulative harmful effects of oxidative stress.
Bolfă Pompei Florin – Rezumat teză de doctorat
47
PARACLINICAL EXAM IN EIAV INFECTED EQUIDS
MATERIALS AND METHODS For the investigation of the haematological profile, we collected blood from a total of 84 horses,
47 females and 37 males. Of these, 63 were EIAV positive horses with ages between 1 and 15 years, and the remaining 21 horses were the seronegative for EIA, aged between 1 and 25 years. The six experimental groups were formed in this case as follows: group 1 consisted of 9 horses, group 2 consists of 3 horses, group 3 consists of 36 horses, group 4 consists of 15 horses, group 5 consists of 6 horses and the group 6 consists of 15 horses.
Blood samples were collected individually in sterile conditions on an anticoagulant (EDTA) in vacutainers. Processing of samples was performed in the Discipline of Infectious Diseases Laboratory inside the Faculty of Veterinary Medicine, UASVM Cluj-Napoca.
RESULTS AND DISCUSSIONS
Analyzing the leukocyte chart, we observed that there is no large variation of the mononuclear
or polymorphonuclear cells in the blood of the investigated horses. Similar to the other values obtained in this chapter of laboratory investigations, in the case of haemoglobin, the variations that occurred were not significant between the six experimental groups. The average of the results presented above, are illustrated in table 3, together with the normal values.
In the case of the total leukocytes, in lot 1 the mean value obtained is above normal and in lot 3 a lower value appeared, compared with reference values. Nevertheless, we cannot speak about a leukocytosis (values greater than 15x103/ mm3), or a leukopenia (values below 4x103/mm3). The values of the white line’s cells are within the normal range, except for basophils, which are slightly elevated in the case of healthy animals aged over 5 years (group 6) but also in the same age group of recently infected VAIE (group 3).
Table 3. Mean value of blood leucocyte cells (normal range after Ghergariu et al., 2000)
Haemoglobin levels are directly proportional to those of red cells, and fall within the normal
range. The explanation for this may be asymptomatic stage of development of the AIE, as in time of sampling; none of the investigated horses presented any clinical signs of disease at that timepoint. A
Bolfă Pompei Florin – Rezumat teză de doctorat
48
single haematological examination, however, is insufficient for a paraclinical diagnosis; repeated exams are needed, consistently with the increase in temperature of animals, as between the febrile episodes, haematological parameters may return to normal (Moga-Manzat, 2005).
Laboratory changes associated with the AIE are: moderate to severe thrombocytopenia, severe anaemia normocytic, normochrome, olygocitemic, the hematocrit to 14-20%, leukopenia (up to 2000 cells/mm3) with neutropenia and lymphopenia, hypergammaglobulinemia and increased activity of bile enzymes, the presence of siderocytes, considered specific for EIA (Barna et al., 1995). Among these changes we identified in this paper only hypergammaglobulinemia in EIAV infected equines, parameter that seems to be the most reliable of those investigated to show the real status of the horse investigated (infected or not with VAIE).
Bolfă Pompei Florin – Rezumat teză de doctorat
49
COMPARISON OF A COMMERCIAL ENZYME-LINKED IMMUNOSORBENT ASSAY AND AN AGAR GEL IMMUNODIFFUSION TEST FOR THE SERODIAGNOSIS OF EQUINE
INFECTIOUS ANEMIA
Agar gel immunodiffusion (AGID) tests and enzyme-linked immunosorbent assays (ELISAs) (Suzuki et al., 1982) represent accurate, reliable tests for the detection of EIA in horses, except for animals in the early stages of infection and foals of infected dams. In other rare circumstances, misleading results may occur when the level of circulating virus in the blood during an acute episode of the disease is sufficient to bind available antibody, and if initial antibody levels never raise high enough to be detectable (Toma, 1980). Although the ELISA will detect antibodies somewhat earlier and at lower concentrations than the AGID test, positive ELISAs are confirmed using the AGID test. This is because false-positive results have been noted with ELISAs. The AGID test also has the advantage of distinguishing between EIA and non-EIA antigen–antibody reactions by lines of identity (OIE, 2010).
Virus isolation is usually not necessary to make a diagnosis. Isolation of the virus from suspect horses may be made by inoculating their blood on to leukocyte cultures prepared from horses free of infection. Virus production in cultures can be confirmed by detection of specific EIA antigen by ELISA (Shane et al., 1984), by imunofluorescence assay (Weiland et al., 1982), by molecular tests or by subinoculation into susceptible horses. Virus isolation is rarely attempted because of the difficulty of growing horse leukocyte cultures.
A nested polymerase chain reaction (PCR) assay to detect EIA proviral DNA from the peripheral blood of horses has been described (Nagarajan and Simard, 2001). The nested PCR method is based on primer sequences from the gag region of the proviral genome. It has proven to be a sensitive technique to detect field strains of EAV in white blood cells of EIA infected horses; the lower limit of detection is typically around 10 genomic copies of the target DNA (Nagarajan and Simard, 2001; Nagarajan and Simard, 2007). A real-time reverse-transcriptase PCR assay has also been described (Cook et al., 2002). To confirm the results of these very sensitive assays, it is recommended that duplicate samples of each diagnostic specimen be processed. Because of the risk of cross contamination, it is also important that proper procedures are followed.
Due to the persistence of EIAV in infected equids, detection of serum antibody to EIAV confirms the diagnosis of EIAV infection. Precipitating antibodies are rapidly produced as a result of EIA infection, and can be detected by the AGID test. Specific reactions are indicated by precipitin lines between the EIA antigen and the test serum and confirmed by their identity with the reaction between the antigen and the positive standard serum. Horses in the first 2–3 weeks after infection will usually give negative serological reactions. In rare cases the post-infection time prior to the appearance of detectable antibody may extend up to 60 days (OIE, 2010).
A positive test result by ELISA should be retested using the AGID test to confirm the diagnosis because some false-positive results have been noted with the ELISA. The results can also be confirmed by the immunoblot technique (OIE, 2010).
The purpose of the present study was to estimate the performance characteristics of a commercially available competitive enzyme-linked immunosorbent assay (cELISA) in comparison with a reference AGID kit for the detection of equine infectious anemia (EIA) antibodies in horses from Romania. A total of 87 positive and 5 negative samples by the reference AGID were tested. The cELISA directed against antibodies to the p26 core protein performed relatively well in comparison with the reference AGID, with similar accuracy and precision. Doubtful and negative results obtained at the retesting with the immunosorbent assay, can be explained by the particularities of in vivo evolution and pathogenesis of the viral infection. The cELISA kit can be used somewhere earlier in the evolution of EIA comparatively to AGID, having a higher sensibility and specificity, the possible errors due to subjectivism in colour evaluation being eliminated by the use of a spectrophotometer.
Taking into consideration that the EIA is the infectious disease which causes the most important damages, economically speaking, to all horses in Romania, EILSA testing in the future, could be introduced in veterinary practice as a quickly and effectively test, allowing the efficient and accuracy testing, of a high number of serological tests in a short time and an objective evaluation of the results by
using a spectrophotometer. The possible implementation of the method in Romania in the future, similar to other countries, would still impose the retesting of positive samples through AGID.
MATERIALS AND METHODS
The present study was divided in two experimental directions: in the first stage we tested serum
samples collected from horses diagnosed EIA positive (n=28) at different time points prior to testing. The aim was to observe the differences in serological response compare to the prior AGID testing. A second phase consists of retesting the positive serum from a total of 64 horses. This was performed in order to verify the correlation/accordance of the two tests (AGID and cELISA). All blood samples tested originated from the 92 horses located in Bistriţa-Năsăud County.
For the first part of the present study we used serum samples from 28 horses, 20 females and 8 males, aged between 1 and 18 years. EIA was formally diagnosed by the official test (AGID) in the County Laboratory for Animal Health and Veterinary Health (LSVSA) of Bistrita, inside of bi-annual testing plan requested by the ANSVSA (National Sanitary Veterinarian Agency and for Animal Health). The animals were diagnosed as EIA positive between 1998 and 2005, and serum from these animals, collected in 2005 and 2006 was retested using enzyme immunoassay tests (EIA IDEXX kit, cELISA), in accordance with the producer’s recommendations. Retesting was conducted from 1 year, up to 7 years 6 months after initial diagnosis. Testing took place in the Laboratory of Infectious Diseases Discipline USAMV Cluj-Napoca, at the end of 2006.
For the second part of the experiment, the 64 serum samples were otained by courtesy of LSVSA (Sanitary Veterinarian Laboratoy and for Animal Health) Bistrita, in the spring of 2008, after testing them as positive by AGID method, and were retested by cELISA in the autumn of 2008 in the Laboratory of Pathological Anatomy, Necropsy Diagnosis and Forensic Medicine and the Parasitology discipline of the UASVM Cluj-Napoca, at the end of 2006.
RESULTS AND DISCUSSION
In the U.S.A, there are currently four ELISA methods for diagnosis EIA approved by the
Department of Agriculture, and can be applied worldwide: a competitive ELISA method, and three non-competitive ELISA methods. Competitive ELISA method and a non-competitive method detect antibodies produced against the antigen internal protein p26. Another non-competitive method incorporates both internal protein p26 antigen and gp45 (viral trans membrane protein), while the last non-competitive method detects antibodies against gp45 protein. In all tests use typical ELISA protocol (OIE, 2010). A positive ELISA should be retested by the AGID test to confirm the diagnosis because of the potential occurrence of false positive results. Results can also be confirmed by the immunoblot technique (OIE, 2010).
The competitive kit IDEXX ELISA (cELISA EIA) is a rapid, convenient and specific test for the detection of antibodies anti EIAV in the serum of horses. The purified antigen p26 and monoclonal antibodies are designed to reduce the frequent non-specific reactions in ELISA tests. Correlation between IDEXX EIA cELISA and the immunodiffusion test in agar gel is over 99%, according to the producer.
Results from the first experiment are illustrated in Table 4. Of the 28 animals tested, 23 were positive at the Coggins test (diagnosed 7 years and 6 months ago) and 5 were EIA negative.
The number of negative reactions using cELISA increased from 5 to 8 (appearance of dark blue color, i.e. a wavelength greater than the negative control).
Of the three previously positively tested horses towards EIAV by the AGID technique and
tested as negative by cELISA, one was diagnosed 7 years and 6 months ago, another 6 years and 6 months ago and the last 1 year before the harvest time for testing. The negative progression of these horses could be explained by entering into the asymptomatic stage of disease. This type of results (false negative) can be seen in horses when the level of circulating virus in the blood during an acute episode of the disease is sufficient to bind available antibody, making it impossible to detect such an animal (USDA, 2003). It is also one of the reasons why an animal that has a positive Coggins serological test result is considered positive for life and is not retested. However, in our case, an eventual retesting at a later time,
Bolfă Pompei Florin – Rezumat teză de doctorat
51
could give a positive serological response (either the AGID test or the cELISA) because of pathogenesis mechanisms of the virus, or following a possible immunosuppression.
Table 4
Synthesis of the comparative testing for 28 animals
cELISA
positive animals
cELISA
negative animals
AGID positive animals 20 3
AGID negative animals 0 5
The results were interpreted statistically using Fisher test (p = 0.0005698) which confirms that
the results of the two serological tests correlate positively. Thus the positive results of a test may be associated with the second.
The summarized results of the second experiment are illustrated in table 5 (retesting of 64 serum from horses positive by AGID).
Table 5
cELISA test results following the retesting of the AGID positive sera (n=64)
cELISA
positive animals
cELISA
dubious animals
cELISA
negative animals
AGID positive animals 62 2 0
The analysis of results shows that there are two situations in which 2 of the horses previously
tested EIA positive, had optical densities values higher than the positive control but lower than the negative control, and were classified as suspicious. cELISA thus reconfirms the results of Coggins testing, the results considered suspicious must be retested by AGID.
Analyzing the results presented above, we observed that the official results of the AGID test used in Romania presented equivalent results with cELISA method. cELISA method (spectrophotometric interpretation) has the advantage of being less subjective than the AGID test (visual interpretation) and the results are obtained very quickly (2 hours for cELISA, compared to 48-72 hours for Coggins test).
False-negative results occur more frequently in AGID tests, while false positive results are more frequently for ELISA (USDA, 2003). The occurrence of discordant results between AGID and cELISA appear through two major mechanisms (Pare and Simard, 2004). The most common situation where we can have a positive cELISA result and a negative AGID result is when there is a low level of p26 antibodies in the studied sample, an easily identifiable situation by immunoassay tests. Another possible situation for the occurrence of discordant results is the presence in the sample of antibodies directed against the determinant interspecific of antigen p26, which cannot be detected by AGID test. These antibodies may occur in horses exposed to related lentyviruses in nature. All these aspects (Table 3) occur when the two tests are applied simultaneously (Pare and Simard, 2004).
In the present research, from a total of 92 horses (Table 6) tested by those two serological tests (AGID and cELISA), at the AGID test, 87 positive and 5 negative were found. Differences occurred at cELISA for 5 of previously AGID positive horses. Thus, 2 horses initially tested as positive, in cELISA were found suspicious (same serum testing) and other 3 horses led to a negative cELISA result (testing of serum collected at different times from same horses).
Table 6:
Bolfă Pompei Florin – Rezumat teză de doctorat
52
Comparative aspects between AGID and cELISA
AGID cELISA
Antigen p26 p26
Antibodies Titer Increased Low
Specificity Good Raised
Sensitivity Low Raised
Test period 48 to 72 hours 45 minutes
EIA positive (+) 87 82
EIA negative (-) 5 8
EIA suspicious (±) 0 2 Commercial competitive cELISA kit gave equivalent results with the AGID test. The sensitivity
of this test is intentionally kept under observation to avoid false positive reactions. However, using cELISA testing, it is possible to detect lower antibody titters, compared with AGID test, so that sometimes discrepancies could appear between the results obtained from those 2 tests. The reason of this phenomenon could be, also, the presence in serum of antibodies directed against the determinant of interspecific (Gs fraction 2) of the protein p 26 detectable only by cELISA test and not by AGID. Such antibodies may appear from infection with other lentiviruses, and the results must be considered false positive and confirmed by immunoblot test, designed to identify protein p 26 (Paré and Simard, 2004).
The level of information from databases is increasing, related to EIAV the genes and antigens. In the future, EIAV diagnostic methods should include determinants of viral proteins and use stable detection systems. For the interpretation of such tests an easier interpretation would be desirable compared with AGID test, they should be more "user friendly", which could make them more usable and more attractive. In the absence of effective vaccines against VAIE, disease control is dependent on large-scale tests to identify and control the movement of infected animals. The increasing of the efficiency test method could be achived by managing a high number of the animals from the risk populations (Paré and Simard, 2004).
Bolfă Pompei Florin – Rezumat teză de doctorat
53
FIRST GENETIC CHARACTERIZATION OF ROMANIAN EIAV FIELD ISOLATES
Material and methods
For molecular biology tests we used the of blood collected on anticoagulant (EDTA) of 25 horses from Bistrita-Nasaud County, aged between 6 months and 17 years, 6 males and 19 females. The equines were tested as positive for EIA 3 weeks up to 7 years before sample collection. After sampling, the blood was transported in the UASVM Cluj Napoca, Department of Infectious Diseases. Here it was centrifuged (6000 rpm for 10 minutes) plasma was removed, and the buffy coat stored at low temperature (-20°C), then transported to the laboratory Retrovirusologie, Evolutie and Cancer, UMR 754 of the University Claude Bernard, Lyon, France where it was further processed.
Molecular analysis of samples
Extraction of genomic DNA Genomic DNA was extracted from buffy coat samples with the kit "Qiamp®” (Qiagen),
respecting the manufacturers recommendations, with a few exceptions: we doubled the amount of buffer AL, proteinase K and absolute ethanol, due to increased volume (≈ 200μl) of buffy coat. The integrity of DNA extracted was checked by electrophoresis migration (3 μl sample, 3 μl 1 kb ladder and 2 μl media 6X loading dye) on an agarose gel with a concentration of 0.8%, containing "SYBR®Safe", (Molecular Probes, France), a colouring intercalation agent of nucleic acid diluted 1/10.000 in 1X TAE buffer (40 mm of Tris, 20 mM sodium acetate and 1 mm EDTA, pH 8.3). DNA was visualized by ultraviolet transilumination. DNA concentration was measured by reading absorbance at 260 nm using a "SmartSpec ®3000" spectrophotometer (BIORAD). Genetic material purity was checked by 260nm/280nm report.
Amplification by polymerase chain reaction (PCR)
DNA control The quality control of DNA for amplification was carried out after amplification by polymerase
chain reaction (PCR) on part of β-actine gene (Nagarajan and Simard, 2001). PCR mixture contained 500 ng ADN/10μl, 1 μl of 10 mM of each gene specific β-actine primer (GnC5 and GnC6), 1 μl of 10 mM dNTP (dezoxinucleotid-triphosphate), 0.25 μl of Go Taq DNA polymerase 5 U/μl, 10 μl of buffer 5X Green Go Taq Buffer to a final volume of 50 μl.
The quality check was performed with primers actine β-5 '(5'-ATG TGC AAG GCC GGC TTC G-3') and actine β-3 '(5'-TTA ATG TCA CGC ACG ATT TCC-3' ) (Nagarajan and Simard, 2001), designed after the human sequence of β-Actine, which generates a fragment of 580 base pairs (bp). PCR conditions were as follows: 5 minutes (min) at 95 ° C for initial denaturing, 35 cycles of 30 seconds (s) at 95 ° C then 30 s at 56 ° C then 30 s at 72 ° C, and 10 min at 72 ° C for final elongation. PCR products were visualized by ultraviolet transillumination after electrophoretic migration on a 1.5% agarose gel containing "SYBR®Safe”, 1/10.000, in 1X TAE buffer. Apart from sample 1279, DNA samples from the experiment were amplified.
EIAV amplification The amplification of the region from gag gene that encodes the viral capsid was performed by a
hemi-nested PCR. For the external PCR, 500 ng of DNA from each sample was mixed with 10 mM of each primer, 10 mM dNTP, 5 units of Go Taq DNA polymerase, buffer 5X Green Go Taq Buffer to achieve a final volume of 50 μl.
Bolfă Pompei Florin – Rezumat teză de doctorat
54
Figure 7. PCR amplification of the EIAV gag gene from PBMC. Electroforesis on 2% agarose gel,
Sybr®Safe stained. The products amplified at 580 pb corespond to the cellular gene β –actine; unrevealed amplicons at 260 pb corespond to the external PCR for the gag gene and the amplicons at 201 pb
corespond to the internal PCR on the gag gene. Positive witnesses corespond to limph-node and hepatic DNA from horse 983. Negative witness is represented by water instead of horse genomic DNA.
Primers used were EIAV3 and EIAV5. For the internal PCR 5 μl from the products of the
external PCR were mixed with 10 mm of each primer, 10 mM dNTP, 5 units of Go Taq DNA polymerase, buffer 5X Green Go Taq Buffer to a volume final 50 μl. Primers used were EIAV4 and EIAV5. PCR reactions generated fragments of 260 and 210 bp. Conditions for reactions were: 5 min at 95 ° C for initial denaturing, 35 cycles of 30s at 95°C - 30s at 56°C – 1 min at 72°C, followed by 10 min at 72°C for final elongation. PCR products were visualized by ultraviolet transillumination after electrophoresis migration on a 2% agarose gel containing "SYBR®Safe”, at 1/10.000 in 1X TAE buffer (figure 7).
Cloning of the gag products Gag gene amplicons were obtained by TA cloning. This technique is based on a property of DNA
Taq polymerase that adds a deoxiadenozin (A) to the end no 3' of PCR product. The products obtained after amplification are inserted at the extremity T (deoxitimidin) 5' level of the plasmid vector. The gag
β -actine gene amplification
Gag gene amplification
Bolfă Pompei Florin – Rezumat teză de doctorat
55
amplicon’s connection with PCR ® 2.1 plasmid vector (Promega, France) was done respecting supplier's work instructions. The ligature products were transformed into the competent bacteria Escherichia coli JM 109. Plasmid PCR®2.1 containing an ampicillin resistance gene, which encodes β-lactamase, allowing selection of transformed bacteria containing the vector, and LacZ gene that encodes the β-galactozidase over the control of the lactose operon promoter, allowing the identification of transformed bacteria which have integrated the plasmid cut-in. Inserting a cut-in in LacZ gene is interrupting his reading frame.
Bacteria were grown on LB agar selective medium (10g/l bacto-tryptone, 5g/l yeast extract, 10g/l sodium chloride, 15g/l agar) containing ampicillin at 50 mg/μl allows selection of bacteria transformed. The medium was supplemented with 10μmol of IPTG (Isopropyl-β-D-Thiogalactopyranozide), inductor for the Operon lactose, and 1μg of X-gal (5-bromo-4-chloro-3-indolyl-β-D-Galactozidase), substrate chromogenic of β galactozidase. In the presence of lactose, the bacteria that have integrated the plasmid were unable to express β galactozidase and have led to white colonies. Blue colonies represent bacteria capable to hydrolyze the X-gal substrate thanks to the presence of integrated plasmid, making Lac-Z gene functional.
Plasmids were extracted using the kit "Wizard®Plus Minipreps DNA Purification System (Promega, France), then digested with EcoRI restriction enzyme whose sites are located to one side of the cloning sites. The digestion products were visualized by UV transillumination following electrophoresis on a 1.2% agarose gel containing SYBR®Safe at 1/10.000 in 1X TAE buffer. Colonies (10 each) were reinserted in an LB broth (Luria-Bertani: 10g/l bacto-Tipton, 5g/l yeast extract, 10g/l sodium chloride) containing ampicillin at 50 mg/μl , and incubated overnight at 37 ° C in a mechanical shaker.
Extraction of plasmid From individual cultures, plasmid DNA was extracted using the "PLASMID MIDI KIT®DNA
purification system” (Promega, France) (Sambrook and Rossel, 2001). Extraction of the products digested by the restriction enzyme EcoRI (Promega, France) were viewed after electrophoresis migration on a 0.8% agarose gel containing SyBr ® Safe at 1/10.000 in 1X TAE buffer by ultraviolet transillumination. Plasmid DNA concentration was measured by reading absorbance at 260 nm on spectrophotometer ""SmartSpec ®3000”. DNA purity was checked by reading the report 260nm/280nm. Clones obtained were identified by the following nomenclature: pEIAVX, where "p" is the plasmid origin of DNA, "EIAV" - equine infectious anemia virus and "X" is corresponding to the identification number.
Gag region sequencing Plasmid DNA of clones containing a cut-in gag were sequenced (Genomexpress, Grenoble,
France) using universal primers T7 (5'-TAA TAC GAC TCA CTA TAG GGC GA 3 ') and SP6 (5'-ATT TAG GTG ACA CTA TAG AAT ACT-3 ').
Sequence analysis Sequences aligning, construction of the phylogenetic trees and calculation of the divergence
matrix was carried out using the program Vector NTI Advance (TM) 11.0 (Invitrogen). Sequences used to compare were French sequences pEIAV21_983_Fr, pEIAV40_1028_Fr, pEIAV41_1028_Fr, pEIAV42_1029_Fr, pEIAV44_1029_Fr, pEIAV43_1029_Fr, pEIAV7_860_Fr, America sequence EIAVUK (AF0163 16).
II.5.2.2. Results and discussion
Characterization of isolates EIAV obtained
Characterization of gag gene Nine clones were obtained and analyzed, namely pEIAV107_1271, pEIAV108_1271,
pEIAV114_1282 and pEIAV115_1287, compared with the clone EIAV UK and 7 French clones. It was noted that there is a genetic similarity between the Romanian and the French ones. Most Romanian clones (except 113) showed point mutations distributed into all sequences after the alignment, considering the reference clone EIAVUK and the French isolates (figure 9), characteristic according to the geographical origin of the clone. There is a rapprochement between the Romanian and French isolates, which is more important than the reference strain, against which there are many divergent positions.
Phylogenetic analysis based on nucleotide sequences obtained allowed the presence of different subtypes (subtype 1 and 2), are shown in figure 8. They are distributed, as noted, mostly, according to the geographical origin of the EIAV clone, sequentially and analyzed.
Figure 8. Phylogenetic tree of the cloned gag sequence (obtained by Neigbor joining, wit abootstrap if 1000 iterations, which permits the visualisation of branches). There are 2 major subtypes, the majority
corresponding to geographical origin.
Bolfă Pompei Florin – Rezumat teză de doctorat
57
Figure 9. Alignment of the 210 nucleotides (504 to 714) from the gag gene towards EIAVuk (Cook et al., 1998). We used the French isolates pEIAV7, pEIAV21, pEIAV40, pEIAV41, pEIAV42, pEIAV43, pEIAV44 and the Romanian isolates pEIAV107, pEIAV108, pEIAV109, pEIAV110, pEIAV111,
pEIAV112, pEIAV113, pEIAV114 and pEIAV115. Mutations characteristic for the geographical origin are represented by a blue star.
The present study was performed for a genetic characterizes of the natural EIAV circulating
strains in Romania, based on blood samples from naturally infected horses. It was detected by gene amplification, part of proviral genome integrated gag gene of PMBC. In the future we will be trying the amplification of env gene as well, to culture the virus, from other tissues or organs as well (plasma, liver tissue and spleen tissue).
Bolfă Pompei Florin – Rezumat teză de doctorat
58
None of the infected horses used in the experiment show any clinical signs of EIA at the time of biological sample collection. The present study represents the first EIAV genetic characterization in Romania. It managed to demonstrate the fact that the Romanian virus clones, formed a clearly distinct population from other viruses already known in France or the United-States. Of the nine clones studied, the portion of the gag gene analyzed is very close to eight of them.
II.5.3. DISCUSSION ON DIFFERENT EIA DIAGNOSIS TECHNIQUES The test widely practiced today in the routine work of laboratories for EIA confirmation is Coggins test (AGID). It helps in the diagnosis of clinical forms, latent, and certifies movement of EIA-free horses within the state or outside its borders. With the introduction of new serological diagnostic methods, natural range of the EIA could be controlled more effectively, including the total number of horses infected (diagnosed by Coggins test), and others with a lower level of antibodies, showed by granular tests (ELISA, Western Blot or PCR). Of all tests, the most trustworthy and high diagnostic value remains Coggins test, which has however the disadvantage that the same animal is not consistently positive, requiring repeat it. Surprising the animal with EIA on the eclipse phase, in terms of antibodies, in case of a single control, could lead to the interpretation that the animal is EIA free. OIE directive, provided in the Manual of Standards for Diagnostic Tests and Vaccines, 2008 edition, states that tests currently approved for the diagnosis of the EIA are test ID (Coggins) and ELISA, except that, the positive ELISA must be confirmed by the Coggins test. To confirm these tests, the immunoblot test is indicated (OIE, 2009). There are several cases where PCR can be used to detect horses EIAV infected, namely: in case of conflicting results on serological tests; suspected infection, but negative or questionable serological results; complementary test to the serological ones, for confirmation of positive results; confirmation of early infection, before developing EIAV antibodies; ensuring that the needed horses for antiserum production, vaccine or blood donors are EIAV free; confirmation status of a foal from a mare infected (OIE, 2009).
.
Bolfă Pompei Florin – Rezumat teză de doctorat
59
CONCLUSIONS
1. Total immuneglobins and CIC evolve interdependentely in the pathogenesis of EIAV. The increase of these immune indicators is associated with recent viral infection, when the organism has not achieved an equilibrium with the infecting virus and with aging, when the immunological clearance capacity is decreasing.
2. Sera from EIAV infected horses present a more obvious bactericidal effect at a higher concentration,
possibly due to some component activated by the virus, like the complement.
3. Stimulating effects of nettle extract on the phagocytosis in young, recently EIAV infected horses, followed by an inhibitory effect, and could be explained by a possible rapid carbon particle ingestion, followed by cell membrane fragility.
4. As a consequence of a lack of antigenic stimulation in vivo, the value of stimulation indexes are
decreases in healthy animals compared to EIAV infected ones, independently of age or duration of disease progression.
5. The incapacity of response to the used mitogen inductors (in horses over 5 years of age, EIA
positive), is emphasised by the duration of disease evolution, as opposed to the trend seen in young animals, probably due to reduction in immune responsiveness with aging.
6. Young, newly infected animals, show an increased cellular reactivity compared to older animals, due
to infection, on one hand, and general increased immunoreactivity in younger animals, immune recovery effects of the used extracts being more obvious.
7. Changes indicating an imbalance in oxidants/antioxidants are more obvious in EIAV infected horses
versus healthy ones, both at the enzymatic and non-enzymatic level as well as in terms of total antioxidant status.
8. Due to frequent presence of circulating virus in recently infected horses towards equines in
asymptomatic stage, oxidative damage is more obvious in the first category, becoming more severe with age, as the antioxidant defense capacity reduces.
9. Changes in haematological parameters in EIA are associated with febrile episodes, and lack of
circulating virus or fever at the time of sampling, confirms the antigenic "quiescence" period.
10. The present study is the first to demonstrate a correlation between a viral disease of horses (EIAV) and the oxidative stress state.
11. A possible introduction in practice for EIA diagnosis of a cELISA method would allow an earlier
and more sensitive diagnosis, but will not permanently remove AGID testing, which is recommended by the OIE to confirm positive results.
12. Genetic studies of Romanian VAIE clones, from a portion of gag gene, proved that they are clearly
distinguishable from other viruses already known in the U.S. or in France.
13. Analysis of obtained nucleotide sequences, allows us to assert that Romanian and French strains are much closer together than with the North American reference strain.
14. Distribution of most analyzed isolates according to their geographical origin, suggests the circulation
in Europe of different EIAV viral strains.
Bolfă Pompei Florin – Rezumat teză de doctorat
60
SELECTED REFERENCES
21. Andrei S., Pintea A., Catoi C., Sevastre B., Varga A., Baba A.I., Antioxidant enzymes activities, lipids and proteins oxidation – markers of oxidative stress in rats i.p. treated with asbestos, Bull. USAMV CN (2002) 59:598-603.
22. Aquaro S., Scopelliti F., Pollicita M., Perno C.F., Oxidative Stress and HIV Infection: Target Pathways for Novel Therapies? Future HIV Therapy (2008) 2:327-338.
23. Calamari L., Ferrari A., Bertin G., Effect of selenium source and dose on selenium status of mature horses, J. Anim. Sci. (2009) 87:167-178.
24. Castejón F., Trigo P., Muñoz A., Riber C., Uric acid responses to endurance racing and relationships with performance, plasma biochemistry and metabolic alterations, Equine Vet. J. Suppl. (2006) 36:70-73.
25. Craigo J.K., Montelaro R.C., EIAV Envelope Diversity: Shaping Viral Persistence and Encumbering Vaccine Efficacy, Curr. HIV Res. 2010 Feb 18, [Epub ahead of print].
26. Delguste C., de Moffarts B., Kirschvink N., Art T., Pincemail J., Defraigne J.O., Amory H., Lekeux P., Change in blood antioxidant status of horses moved from a stable following diagnosis of equine motor neuron disease, Can. Vet. J. (2007) 48:1165–1167.
27. Gil del Valle L., Gonzales I., Tarinas A., Alvarez A., Molina R., Tapanes R., Perez J., Evaluation of oxidative stress in AIDS pediatric patients, Acta Farm. Bonaerense (2004) 23:466-471.
28. Gil L., Martínez G., González I., Tarinas A., Alvarez A., Giuliani A., Molina R., Tápanes R., Pérez J., León O.S., Contribution to characterization of oxidative stress in HIV/AIDS patients, Pharm. Res. (2003) 47:217–224.
29. Jaime M, Ramirez-Boscá A., Soler A., Díez A., Carrión-Gutiérrez M.A., Díaz-Alperi J., Quintanilla-Ripoll E., Bernd A., Quintanilla-Almagr E., Increase with age of serum lipid peroxides: Implications for the prevention of atherosclerosis, Mechanisms of Ageing and Development (1998) 100:17-24.
30. Kirschvink N., de Moffarts B., Lekeux P., The oxidant/antioxidant equilibrium in horses. The Veterinary Journal (2008) 177: 178–191.
31. Kirschvink N., Fievez L., Bougnet V., Art T., Degand G., Smith N., Marlin D., Roberts C., Harris P., Lekeux P., Effect of nutritional antioxidant supplementation on systemic and pulmonary antioxidant status, airway inflammation and lung function in heaves-affected horses. Equine Vet. J. (2002) 34:705–712.
32. Leroux C., Cadore J.L., Montelaro R.C., Equine infectious anemia virus (EIAV): what has HIV’s country cousin got to tell us?, Vet. Res. (2004) 35:485-512.
33. Lippi G., Montagnana M., Franchini M., Favaloro E.J., Targher G., The paradoxical relationship between serum uric acid and cardiovascular disease, Clinica Chimica Acta (2008) 392:1-7.
34. Mandas A., Iorio E.L., Congiu M.G., Balestrieri C., Mereu A., Cau D., Dessì S., Curreli N., Oxidative Imbalance in HIV-1 Infected Patients Treated with Antiretroviral Therapy, J. Biomed. Biotechnol. (2009) doi: 10.1155/2009/749575.
35. Marañón G., Muñoz-Escassi B., Manley W., García C., Cayado P., de la Muela M.S., Olábarri B., León R., Vara E., The effect of methyl sulphonyl methane supplementation on biomarkers of oxidative stress in sport horses following jumping exercise, Acta Vet Scand. (2008) 50(1): 45.
36. Moffarts B., Kirschvink N., van Erck E., Art T., Pincemail J., Lekeux P., Oxidative stress in exercising horses: fact or fiction?, Equine and Comparative Exercise Physiology, (2005) 2: 253-261.
37. Pintea A., Andrei S., Bele C., Biochimie medical veterinară:lucrări practice, Ed. AcademicPres, Cluj-Napoca, 2008, pp. 72-74, 91-92, 108-109, 135-136.
38. Van Regenmortel M.H., Mayo M.A., Fauquet C.M., Maniloff J., Virus nomenclature: consensus versus chaos, Arch. Virol. (2000) 145:2227–2232.
39. Waring W.S., Convery A. , Mishra V., Shenkin A., Webb D.J., Maxwell S.R.J., Uric acid reduces exercise-induced oxidative stress in healthy adults, Clinical Science (2003) 105:425–430.
40. Williams C.A., Kronfeld D.S., Hess T.M., Waldron J.N., Saker K.E., Hoffman R.M., Harris P.A., Oxidative stress in horses i n three 80-km races. Equine Nutr. Phys. Soc. Proc. (2003) 18:47- 2.