

UNIVERSITATEA DE MEDICINĂ ŞI FARMACIE " GR. T. POPA IAŞI"
FACULTATEA DE MEDICINĂ
DISCIPLINA DE HISTOLOGIE
Rezumatul tezei de doctorat
IMPLICAREA METALOENZIMELOR (SOD) ÎN
REDUCEREA SAU NEUTRALIZAREA EFECTELOR
STRESULUI OXIDATIV LA NIVEL PULMONAR
Conducător ştiinţific,
Prof. Univ. Dr. Carmen Lăcrămioara Zamfir
Doctorand,
Oana Raluca Temneanu
-IAŞI 2011-


Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
1
CUPRINS
LISTA DE ABREVIERI
INTRODUCERE
STADIUL ACTUAL AL CUNOAŞTERII
CAPITOLUL I STRESUL OXIDATIV - CONCEPTE ACTUALE
I.1. Caractere generale ale stresului oxidativ
I.1.1. Concepte actuale, mecanisme de acţiune, organe ţintă I.1.2. Alterări ale ADN induse de stresul oxidativ
I.2. Specii reactive de oxigen (SRO)
I.2.1. Mod de formare, particularităţi de reacţie I.2.2. SRO - efecte epigenetice
I.2.3. SRO - molecule semnal
I.2.4. Surse exo - şi endogene de SRO
CAPITOLUL II
MECANISME DE PROTECŢIE ÎMPOTRIVA STRESULUI OXIDATIV II.1. Criterii de clasificare a antioxidanţilor
II.2. Principalele tipuri de antioxidanţi II.2.1.Enzime antioxidante
II.2.2. Compuşi antioxidanţi cu greutate moleculară redusă, cu acţiune directă
II.2.3. Compuşi antioxidanţi cu greutate moleculară redusă, cu acţiune indirectă
CAPITOLUL III
SUPEROXID DISMUTAZELE (SOD)
III.1. Evoluţia conceptului de apărare superoxid dismutazică III.2. Profilul genic al SOD
III.3. Activarea şi mecanismul de acţiune a SOD
III.4. Principalele tipuri de SOD III.5. Evoluţia SOD pe parcursul dezvoltării organismului

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
2
CAPITOLUL IV
IMPLICAŢII ALE STRESULUI OXIDATIV LA NIVEL PULMONAR
IV.1. Surse pulmonare de SRO IV.2. Efecte ale stresului oxidativ la nivel pulmonar
IV.3. Stresul oxidativ de natură fizică şi intervenţia SOD
IV.4. Interrelaţia SOD-melatonină şi adaptarea la stres
IV.5. Stresul oxidativ de natură chimică
CONTRIBUŢII PERSONALE
CAPITOLUL V
MOTIVAŢIA ŞI OBIECTIVELE STUDIULUI PERSONAL
CAPITOLUL VI MATERIAL DE STUDIU ŞI METODE DE CERCETARE
VI.1.Material
VI.2.Metode de lucru VI.2.1. Investigaţii histopatologice şi imunohistochimice
VI.2.2.Metode de determinare a hemoleucogramei şi a parametrilor
stresului oxidativ VI.2.3.Evaluarea morfometrică a parametrilor stresului oxidativ
VI.2.4. Analiza corelaţiilor statistice între parametrii evaluaţi
CAPITOLUL VII REZULTATELE STUDIULUI PERSONAL
CAPITOLUL VIII
DISCUŢII
CAPITOLUL IX
CONCLUZII
BIBLIOGRAFIE

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
3
INTRODUCERE
Viaţa aerobă presupune o implicare promptă şi o intervenţie
absolut indispensabilă a oxigenului şi a compuşilor săi. Desfăşurarea
în condiţii normale a oricărui proces fiziologic este astfel strict
condiţionată de un aport permanent de oxigen. Extragerea oxigenului
din mediul ambient şi distribuţia sa nu numai pentru fosforilarea
oxidativă, dar şi ca substrat pentru reacţii biochimice a fost
conservată pe parcursul evoluţiei speciilor prin dezvoltarea unui
sistem avansat care menţine homeostazia oxigenului şi supravieţuirea
celulară.
Stresul oxidativ, apărut ca o consecinţă a dezechilibrului
dintre verigile pro- şi antioxidante este în prezent atât de frecvent
încât face parte din existenţa cotidiană. În condiţiile în care efectele
sale asupra organismului se însoţesc de nenumărate tulburări grave
ale funcţiilor vitale ale organismului, mecanismele de producere a
stresului oxidativ, dar mai ales cele implicate în protecţia
antioxidantă, deşi intens studiate, nu sunt pe deplin elucidate.
Superoxid dismutazele (SOD) reprezintă o categorie
particulară de metaloenzime, cu rol esenţial în apărarea organismului
împotriva stresului oxidativ. Cu o evoluţie de aproximativ două
bilioane de ani, ce coincide cu proliferarea treptată a organismelor
fotosintetice care încep să producă oxigen, SOD au progresat
continuu drept verigi antioxidante fundamentale ale unui proces care
a permis supravieţuirea. Prin catalizarea reacţiei de dismutare a
radicalilor superoxid în oxigen şi peroxid de hidrogen, aceste
metaloenzime protejează aproape toate celulele expuse la oxigen,
prin îndepărtarea superoxidului şi reducerea concentraţiei tisulare a
oxigenului.
Ideea care a stat la baza acestei teze a urmărit realizarea unei
corelări validată experimental între diversele tipuri de stres oxidativ,
efectele lor asupra plamânului şi respectiv tipul de SOD care se
descarcă masiv şi se distribuie la nivelul ţesutului pulmonar, în aşa

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
4
fel încât să faciliteze identificarea celei mai bune variante terapeutice
bazată pe utilizarea SOD, raportată direct la stresul oxidativ care
agresează organismul.
Lucrarea de faţă este structurată în două mari părţi: o parte
generală - stadiul actual al cunoaşterii şi o parte personală -
contribuţii personale. În partea generală am inclus o trecere în revistă
a celor mai noi date, ipoteze de lucru şi consideraţii generale din
literatura de specialitate despre stresul oxidativ şi efectele sale
generale, precum şi despre SOD.
Primul capitol evaluează cele mai noi concepte despre stresul
oxidativ, cu evidenţierea alterărilor ADN şi a implicării directe a
SRO.
Cel de al doilea capitol abordează diversele mecanisme de
protecţie antioxidantă cunoscute până în prezent, cu menţionarea
tuturor tipurilor de antioxidanţi care susţin aceste procese.
Cel de al treilea capitol furnizează date recente despre
structura şi funcţiile SOD, precum şi despre evoluţia conceptului de
apărare antioxidantă asigurată de familia SOD.
Capitolul patru trece în revistă suita implicaţiilor pe care
activarea SOD le exercită la nivel pulmonar, ţinând cont de faptul că
plămânul este direct şi masiv vizat de stresul oxidativ.
Cea de a doua parte a cercetării rezervată contribuţiilor
personale a inclus un prim capitol-capitolul cinci, care se referă la
motivaţia şi obiectivele acestui studiu, după care capitolul şase
descrie materialul şi metodele utilizate în acest studiu experimental.
Capitolul şapte prezintă rezultatele obţinute în urma investigărilor
histopatologice, biochimice, imunohistochimice, susţinute de analiza
statistică şi de interpretarea morfometrică.
Capitolul opt prezintă o serie de discuţii asupra rezultatelor
obţinute, iar capitolul nouă a fost destinat concluziilor acestui studiu.
Efectuarea acestui studiu experimental complex, care a
implicat un număr mare de animale de laborator şi multiple
investigaţii, a fost posibilă printr-o colaborare şi o conlucrare strânsă

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
5
între domenii care aparent nu au neapărat un numitor comun:
histologie, imunologie, farmacologie clinică şi biochimie.
Un studiu amplu, fie şi cel al unei teze de doctorat, nu poate
acoperi şi argumenta coerent şi complet o temă atât de generoasă şi
de importantă cum este cea a terapiei antioxidante. Pe parcursul
derulării acestei teze, în timpul efectuării studiului experimental, s-au
desprins numeroase verigi colaterale care pot servi cu siguranţă drept
repere ale unei noi etape în această cercetare ştiinţifică. De aceea ne
permitem să afirmăm că această teză reprezintă doar un prim pas în
descifrarea complexelor mecanisme antioxidante bazate pe
participarea familiei SOD.
MOTIVAŢIA ŞI OBIECTIVELE STUDIULUI
PERSONAL
Plămânul reprezintă, datorită localizării şi suprafeţei întinse
de contact cu oxigenul, unul din organele cele mai vulnerabile la
acţiunile stresului oxidativ. Speciile reactive de oxigen produse
intracelular de celulele parenchimului pulmonar sau extracelular, de
către macrofage sau neutrofile, reprezintă elemente cheie ale unei
ample patologii pulmonare. Mecanismele antioxidante, în cadrul
cărora superoxid dismutazele (SOD) sunt considerate cele mai
importante enzime implicate în protecţia antioxidantă, intervenind
prompt în orice formă de instalare a stresului oxidativ, prezintă
numeroase necunoscute, deşi în prezent asistăm la o adevărată
explozie de date şi teorii, furnizate de numeroase studii de
specialitate.
SRO intervin în geneza a numeroase afecţiuni inflamatorii,
degenerative şi tumorale, dar şi în evoluţia unor procese fiziologice,
cum ar fi procesul de îmbătrânire sau dezvoltarea embrionară.
Dualitatea acţiunilor SRO, cu efectele lor benefice, dar şi cu cele
toxice, este parţial explicată de complexitatea funcţiilor lor specifice

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
6
corelate cu particularităţile situsului biologic asupra căruia
acţionează. În acelaşi timp, este una din principalele cauze care ridică
dificultăţi în stabilirea celei mai eficiente metode de abordare a
afecţiunilor cu substrat patologic oxidativ.
Mecanismele antioxidante, în cadrul cărora SOD sunt
considerate cele mai importante enzime implicate în protecţia
antioxidantă, intervenind prompt în orice formă de instalare a
stresului oxidativ, prezintă numeroase necunoscute, deşi în prezent
asistăm la o adevărată explozie de date şi teorii, furnizate de
numeroase studii de specialitate.
Scopul acestei teze de doctorat este acela de a stabili o
corelaţie distinctă între diversele forme de stres oxidativ care pot
perturba structura şi funcţiile pulmonare şi tipul de SOD care
intervine ca trigger antioxidant în fiecare formă de stres; identificarea
enzimei antioxidante care este produsă masiv ca răspuns la
agresiunea O2 poate servi nu numai la monitorizarea intensităţii
stresului oxidativ, ci şi la conturarea unei noi strategii terapeutice
vizând familia de SOD mimetice care ar putea fi o alternativă la
terapiile clasice (administrarea de SOD de natură exogenă a
întâmpinat numeroase probleme şi nu este o soluţie aplicabilă pentru
combaterea efectelor stresului oxidativ).
În acest sens, am realizat un studiu experimental în care am
indus animalelor de laborator o paletă largă de forme de stres
antioxidant : crono-biologic (foto-biologic), chimic, fizic şi respectiv
mixt. La aceste forme de stres am asociat şi un antioxidant clasic,
reprezentat de vitamina C, pentru a urmări modul în care se modifică
amplitudinea stresului oxidativ, precum şi elaborarea pulmonară de
SOD.
Pentru monitorizarea instalării stresului oxidativ şi a efectelor
sale, precum şi a intervenţiei enzimelor antioxidante, am urmărit
evoluţia lor pe trei coordonate: histopatologică, biochimică şi
imunohistochimică. Sudiul morfometric ulterior a investigat pe de o
parte nivelul alterărilor tisulare şi vasculare şi pe de altă parte

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
7
scorizarea studiului imunohistochimic pentru cele trei tipuri de SOD
studiate.
În vederea conturării profilului oxidativ al mediului în care
plămânii îşi desfăşoară funcţiile vitale, am recurs la determinarea
acţiunii individuale a principalelor tipuri de enzime care compun
sistemul enzimatic antioxidant al organismului: SOD, catalaza,
glutazion peroxidaza şi malonildialdehida.
Asocierea hemoleucogramei ne-a permis o interpretare
coerentă a reactivităţii organismului în condiţii de stres. Explorarea
histopatologică a permis surprinderea tipurilor specifice de
modificări tisulare şi vasculare ale plămânilor pe fondul inducerii
stresului pulmonar, cu evoluţia progresivă a inflamaţiei şi fibrozei
pulmonare, care sunt patognomonice pentru agresiunea oxidativă..
Studiul imunohistochimic a urmărit identificarea imunolocalizării
celor trei tipuri distincte de SOD şi corelarea cu forma de stres
oxidativ indus.
MATERIAL DE STUDIU ŞI METODE DE
CERCETARE
Material Lot experimental de studiu
Studiul experimental a utilizat 72 de şobolani rasa Wistar
adulţi, masculi, furnizaţi într-o singură tranşă de Biobaza Institutului
Cantacuzino Bucureşti (aprilie 2010). Animalele au avut o greutate
cuprinsă între 210-230 grame.
Şobolanii au fost divizaţi în 9 loturi distincte, fiecare fiind
format din câte 8 şobolani:
Lot 1 - control;

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
8
Lot 2 - au fost expuşi la lumină continuă, (200 lux), pe tot
parcursul experimentului (stres cronobiologic/fotoperiodic);
Lot 3 - au fost menţinuţi în întuneric pe tot parcursul
experimentului (stres cronobiologic/fotoperiodic);
Lot 4 - au fost puşi să înoate zilnic, 20 minute, în apă rece
(stres fizic);
Lot 5 - li s-a administrat zilnic nitrofurantoin, prin gavare, în
doză unică de 125mg/kgc/zi (stres chimic);
Lot 6 - au fost expuşi la lumină continuă, li s-a administrat
nitrofurantoin în doză unică de 125 mg/kgc/zi în paralel cu
vitamina C 110 mg/kgc/zi (stres oxidativ mixt);
Lot 7 - au fost menţinuţi în întuneric continuu şi li s-a
administrat nitrofurantoin în doză unică de 125 mg/kgc/zi, în
paralel cu vitamina C 110 mg/kgc/zi (stres oxidativ mixt);
Lot 8 - au fost expuşi la lumină continuă şi li s-a administrat
nitrofurantoin în doză unică de 125 mg/kgc/zi (stres oxidativ
mixt);
Lot 9 - au fost menţinuţi în întuneric şi li s-a administrat
nitrofurantoin în doză unică de 125 mg/kgc/zi (stres oxidativ
mixt).
Durata întregului experiment a fost de 60 de zile, după care
animalele au fost eutanasiate. La începutul experimentului, în cea de
a 30-a zi şi la sfârşitul experimentului s-au recoltat probe de sânge de
la nivelul plexului retroorbitar, după ce în prealabil animalele au fost
anesteziate cu câte 75mg/kgc de ketamină. De asemeni, animalele au
fost cântărite la aceleaşi intervale de timp, pentru a monitoriza
evoluţia greutăţii pe parcursul diverselor tipuri de stres oxidativ.
La sfârşitul experimentului, animalele au fost eutanasiate
după anesteziere cu Thiopental 0,01%. După constatarea absenţei
semnelor vitale s-au prelevat fragmente tisulare pulmonare de la
toate animalele din loturile luate în studiu.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
9
Metode de lucru
Investigaţii histopatologice şi imunohistochimice
Studiul histologic: coloraţii uzuale si speciale;
Studiul imunohistochimic: s-au utilizat urmatorii anticorpi
- Anticorp monoclonal de iepure anti SOD1;
- Anticorp monoclonal de şoarece anti SOD2;
- Anticorp monoclonal de şoarece anti SOD3.
Metode biochimice de determinare a hemoleucogramei şi a
parametrilor stresului oxidativ:
- Determinare a hemoleucogramei
- Determinarea SOD, GPx, catalazei, malonildialdehidei (MDA)
Evaluarea morfometrica a parametrilor stresului oxidativ
Analiza corelaţiilor statistice între parametrii evaluaţi:
- Testul χ2 de omogenitate
- t-student
- Testul ANOVA
- Coeficientul de dispersie Pearson (r)
REZULTATELE STUDIULUI PERSONAL
Acest studiu şi-a propus să monitorizeze impactul stresului
oxidativ, sub diversele sale forme, asupra plămânilor, cu identificarea
corelaţiilor dintre eliberarea SOD şi localizarea lor specifică la
nivelul structurilor pulmonare.
Validarea studiului experimental a fost realizată statistic,
urmărindu-se o serie de parametri care să ne permită evidenţierea
reactivităţii pulmonare la stres :
I. Evaluarea statusului biologic al organismelor prin monitorizarea
unor constante specifice (parametri sanguini, greutate)
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
10
II. Confirmarea instalării stresului oxidativ, prin determinarea
principalilor parametri ai stresului oxidativ
III. Evaluarea unor modificări tisulare şi vasculare induse de stresul
oxidativ (grosimea peretelui alveolar, diametrul şi grosimea peretelui
vascular)
IV. Imunolocalizarea celor trei tipuri de SOD, cu scorizarea
marcajului imunohistochimic.
Evaluarea statusului biologic al organismelor prin
monitorizarea unor constante specifice
-10
-5
0
5
10
1 2 3 4 5 6 7 8 9
0.73
-7.02 -7.06
5.35
-6.59
4.777.13
-6.33
-3.26
%
Lot
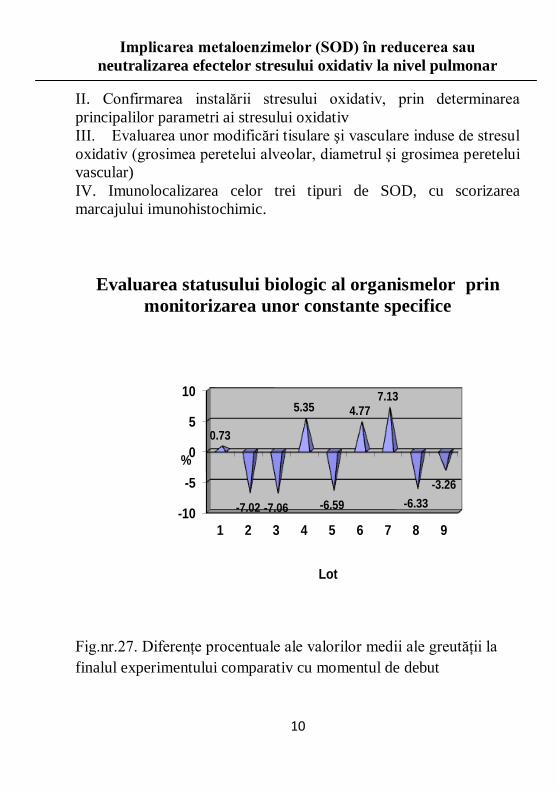
Fig.nr.27. Diferenţe procentuale ale valorilor medii ale greutăţii la
finalul experimentului comparativ cu momentul de debut
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
11
Evoluţia parametrilor sanguini
1.48 0.78 0.54
-10.59
-4.65
0.07
-8.27
25.94
11.71
-15
-10
-5
0
5
10
15
20
25
30
%
1 2 3 4 5 6 7 8 9
Lot
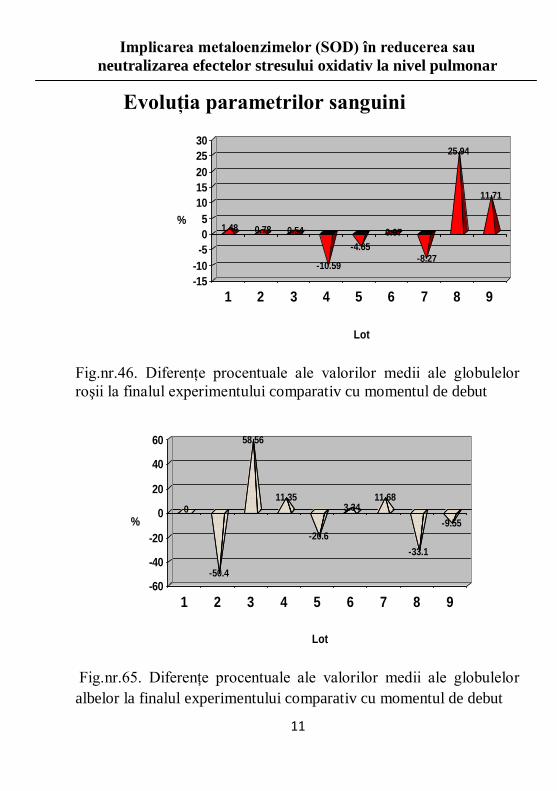
Fig.nr.46. Diferenţe procentuale ale valorilor medii ale globulelor
roşii la finalul experimentului comparativ cu momentul de debut
0
-50.4
58.56
11.35
-20.6
3.3411.68
-33.1
-9.55
-60
-40
-20
0
20
40
60
%
1 2 3 4 5 6 7 8 9
Lot
Fig.nr.65. Diferenţe procentuale ale valorilor medii ale globulelor
albelor la finalul experimentului comparativ cu momentul de debut
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
12
0.67
-45.27-47.06
-9.7-4.56-2.33
-9.23
-66.28
-21.31
-70
-60
-50
-40
-30
-20
-10
0
10
%
1 2 3 4 5 6 7 8 9
Lot
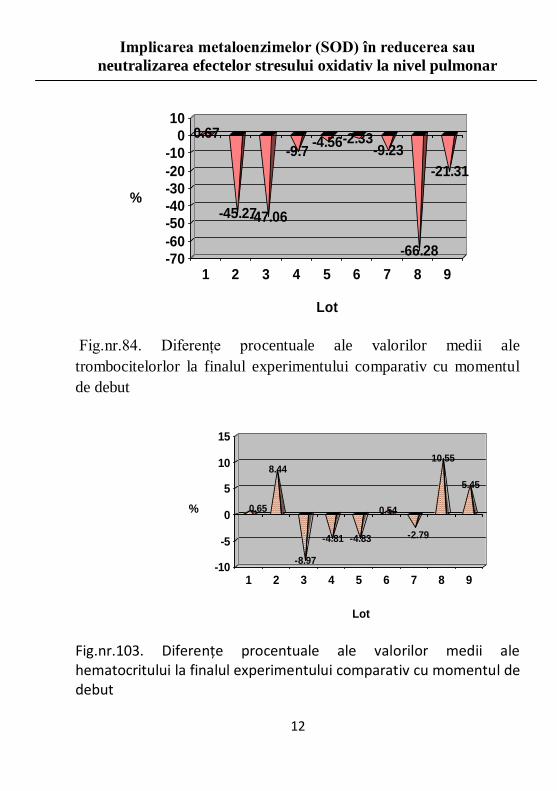
Fig.nr.84. Diferenţe procentuale ale valorilor medii ale
trombocitelorlor la finalul experimentului comparativ cu momentul
de debut
0.65
8.44
-8.97
-4.81 -4.83
0.54
-2.79
10.55
5.45
-10
-5
0
5
10
15
%
1 2 3 4 5 6 7 8 9
Lot
Fig.nr.103. Diferenţe procentuale ale valorilor medii ale hematocritului la finalul experimentului comparativ cu momentul de debut
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
13
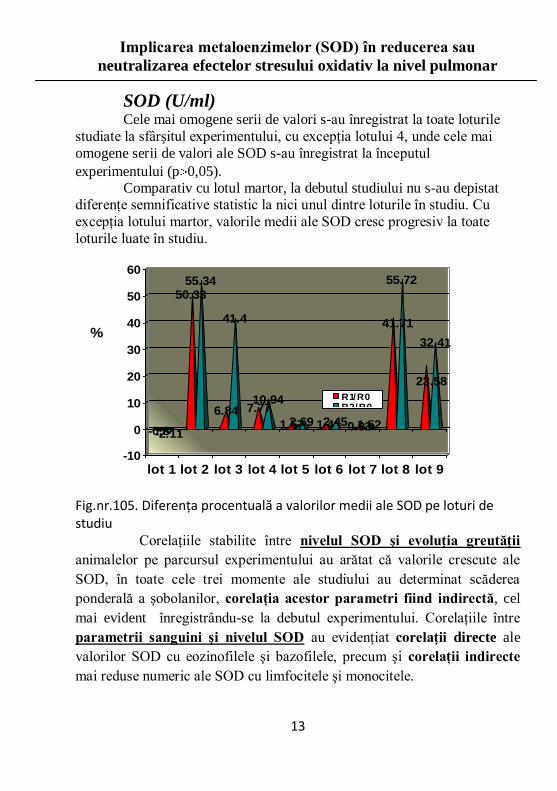
SOD (U/ml) Cele mai omogene serii de valori s-au înregistrat la toate loturile
studiate la sfârşitul experimentului, cu excepţia lotului 4, unde cele mai
omogene serii de valori ale SOD s-au înregistrat la începutul
experimentului (p 0,05). Comparativ cu lotul martor, la debutul studiului nu s-au depistat
diferenţe semnificative statistic la nici unul dintre loturile în studiu. Cu
excepţia lotului martor, valorile medii ale SOD cresc progresiv la toate
loturile luate în studiu.
-0.8-2.11
50.3355.34
6.84
41.4
7.7710.94
1.572.69 1.42.450.931.52
41.71
55.72
23.58
32.41
-10
0
10
20
30
40
50
60
%
lot 1 lot 2 lot 3 lot 4 lot 5 lot 6 lot 7 lot 8 lot 9
R1/R0R2/R0
Fig.nr.105. Diferenţa procentuală a valorilor medii ale SOD pe loturi de studiu Corelaţiile stabilite între nivelul SOD şi evoluţia greutăţii
animalelor pe parcursul experimentului au arătat că valorile crescute ale
SOD, în toate cele trei momente ale studiului au determinat scăderea
ponderală a şobolanilor, corelaţia acestor parametri fiind indirectă, cel
mai evident înregistrându-se la debutul experimentului. Corelaţiile între
parametrii sanguini şi nivelul SOD au evidenţiat corelaţii directe ale
valorilor SOD cu eozinofilele şi bazofilele, precum şi corelaţii indirecte
mai reduse numeric ale SOD cu limfocitele şi monocitele.
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
14
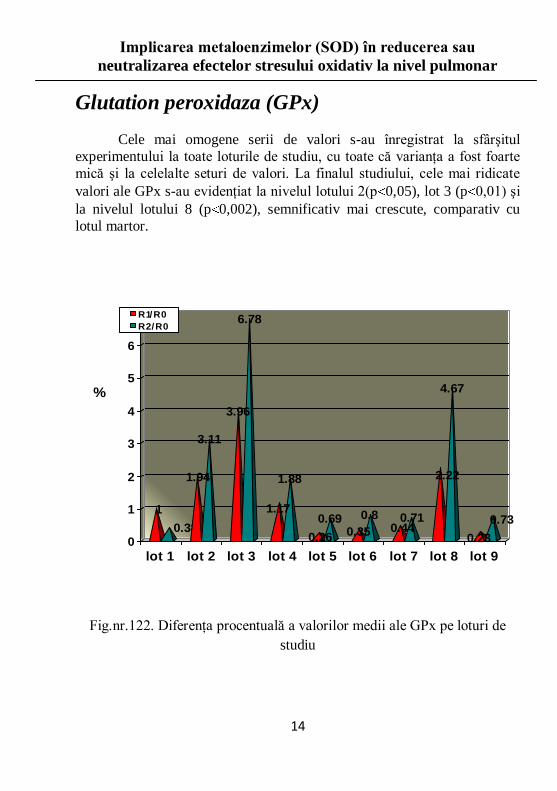
Glutation peroxidaza (GPx)
Cele mai omogene serii de valori s-au înregistrat la sfârşitul
experimentului la toate loturile de studiu, cu toate că varianţa a fost foarte
mică şi la celelalte seturi de valori. La finalul studiului, cele mai ridicate
valori ale GPx s-au evidenţiat la nivelul lotului 2(p 0,05), lot 3 (p 0,01) şi
la nivelul lotului 8 (p 0,002), semnificativ mai crescute, comparativ cu
lotul martor.
1
0.38
1.94
3.11
3.96
6.78
1.17
1.88
0.26
0.690.35
0.80.44
0.71
2.22
4.67
0.28
0.73
0
1
2
3
4
5
6
7
%
lot 1 lot 2 lot 3 lot 4 lot 5 lot 6 lot 7 lot 8 lot 9
R1/R0
R2/R0
Fig.nr.122. Diferenţa procentuală a valorilor medii ale GPx pe loturi de
studiu
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
15
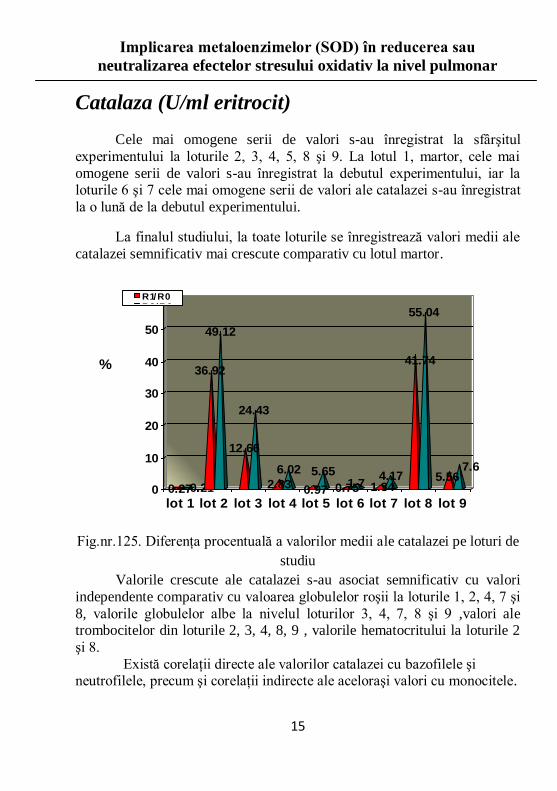
Catalaza (U/ml eritrocit)
Cele mai omogene serii de valori s-au înregistrat la sfârşitul
experimentului la loturile 2, 3, 4, 5, 8 şi 9. La lotul 1, martor, cele mai
omogene serii de valori s-au înregistrat la debutul experimentului, iar la loturile 6 şi 7 cele mai omogene serii de valori ale catalazei s-au înregistrat
la o lună de la debutul experimentului.
La finalul studiului, la toate loturile se înregistrează valori medii ale
catalazei semnificativ mai crescute comparativ cu lotul martor.
0.270.21
36.92
49.12
12.66
24.43
2.83
6.02
0.97
5.65
0.751.7 1.344.17
41.74
55.04
5.567.6
0
10
20
30
40
50
60
%
lot 1 lot 2 lot 3 lot 4 lot 5 lot 6 lot 7 lot 8 lot 9
R1/R0R2/R0
Fig.nr.125. Diferenţa procentuală a valorilor medii ale catalazei pe loturi de
studiu
Valorile crescute ale catalazei s-au asociat semnificativ cu valori
independente comparativ cu valoarea globulelor roşii la loturile 1, 2, 4, 7 şi
8, valorile globulelor albe la nivelul loturilor 3, 4, 7, 8 şi 9 ,valori ale trombocitelor din loturile 2, 3, 4, 8, 9 , valorile hematocritului la loturile 2
şi 8.
Există corelaţii directe ale valorilor catalazei cu bazofilele şi neutrofilele, precum şi corelaţii indirecte ale aceloraşi valori cu monocitele.
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
16
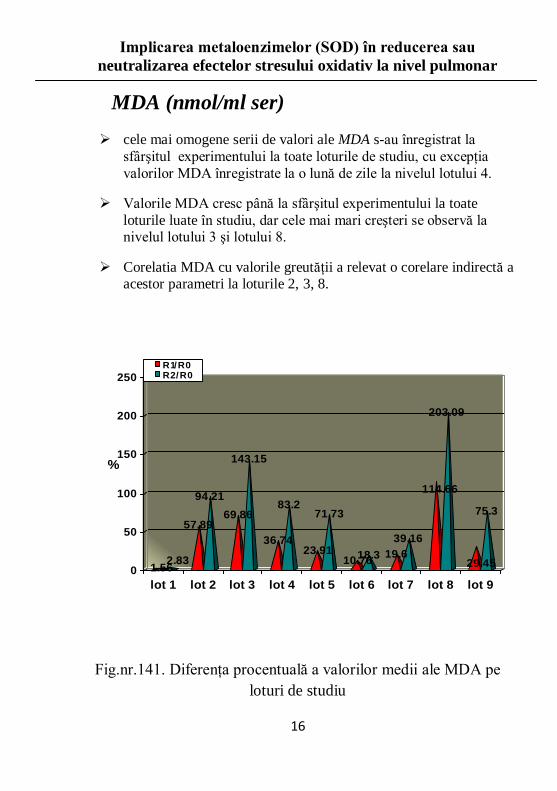
MDA (nmol/ml ser)
cele mai omogene serii de valori ale MDA s-au înregistrat la
sfârşitul experimentului la toate loturile de studiu, cu excepţia
valorilor MDA înregistrate la o lună de zile la nivelul lotului 4.
Valorile MDA cresc până la sfârşitul experimentului la toate
loturile luate în studiu, dar cele mai mari creşteri se observă la nivelul lotului 3 şi lotului 8.
Corelatia MDA cu valorile greutăţii a relevat o corelare indirectă a acestor parametri la loturile 2, 3, 8.
1.562.83
57.89
94.21
69.86
143.15
36.74
83.2
23.91
71.73
10.7618.3 19.6
39.16
114.66
203.09
29.45
75.3
0
50
100
150
200
250
%
lot 1 lot 2 lot 3 lot 4 lot 5 lot 6 lot 7 lot 8 lot 9
R1/R0R2/R0
Fig.nr.141. Diferenţa procentuală a valorilor medii ale MDA pe
loturi de studiu
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
17
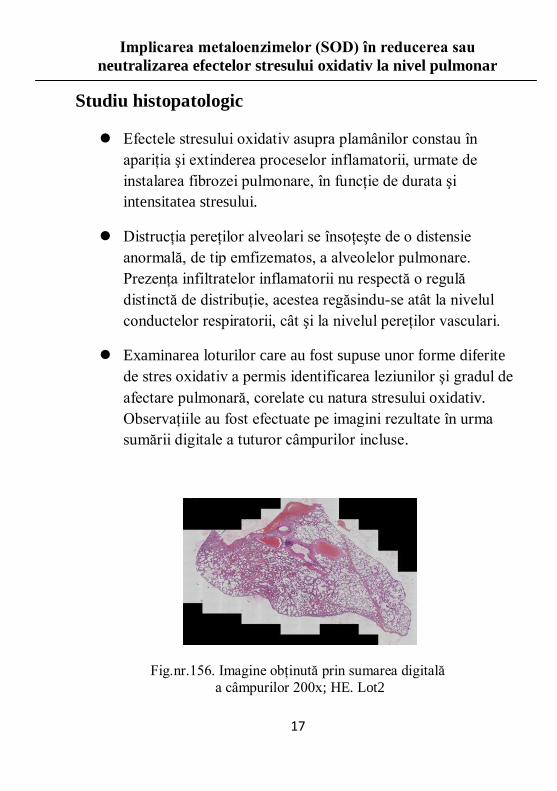
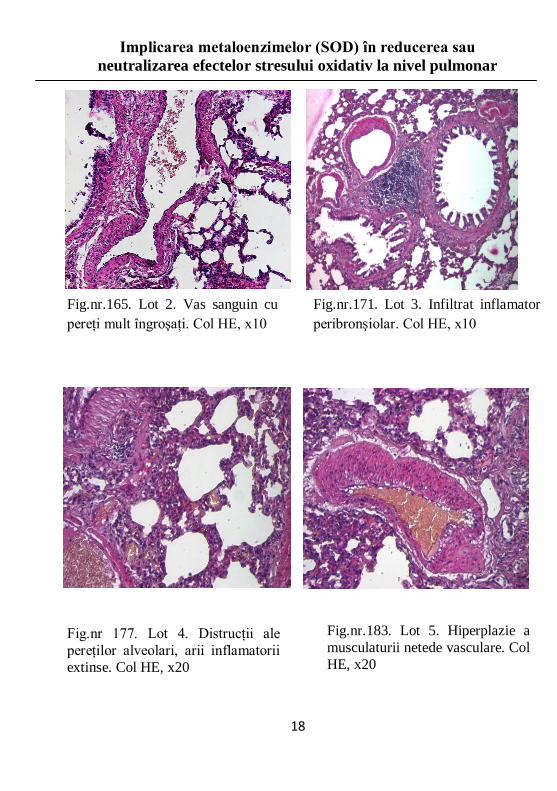
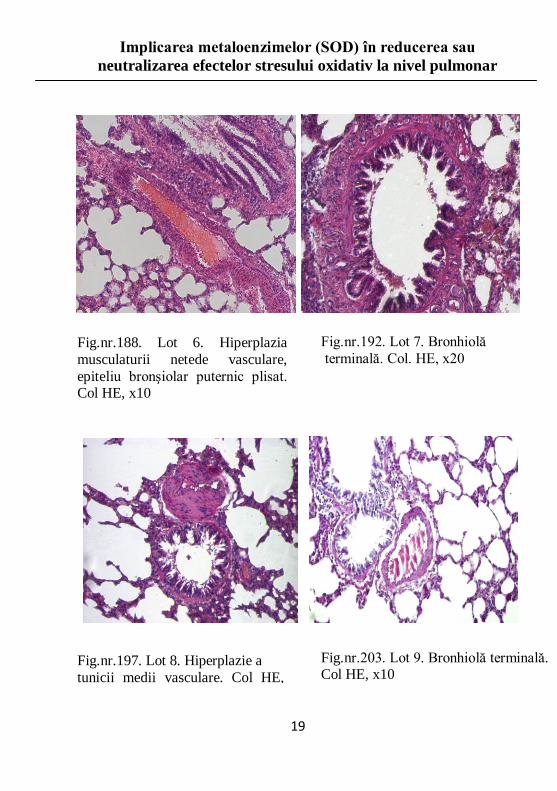
Studiu histopatologic
Efectele stresului oxidativ asupra plamânilor constau în
apariţia şi extinderea proceselor inflamatorii, urmate de
instalarea fibrozei pulmonare, în funcţie de durata şi
intensitatea stresului.
Distrucţia pereţilor alveolari se însoţeşte de o distensie
anormală, de tip emfizematos, a alveolelor pulmonare.
Prezenţa infiltratelor inflamatorii nu respectă o regulă
distinctă de distribuţie, acestea regăsindu-se atât la nivelul
conductelor respiratorii, cât şi la nivelul pereţilor vasculari.
Examinarea loturilor care au fost supuse unor forme diferite
de stres oxidativ a permis identificarea leziunilor şi gradul de
afectare pulmonară, corelate cu natura stresului oxidativ.
Observaţiile au fost efectuate pe imagini rezultate în urma
sumării digitale a tuturor câmpurilor incluse.
Fig.nr.156. Imagine obţinută prin sumarea digitală
a câmpurilor 200x; HE. Lot2
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
18
Fig.nr.165. Lot 2. Vas sanguin cu
pereţi mult îngroşaţi. Col HE, x10
Fig.nr.183. Lot 5. Hiperplazie a
musculaturii netede vasculare. Col
HE, x20
Fig.nr 177. Lot 4. Distrucţii ale
pereţilor alveolari, arii inflamatorii
extinse. Col HE, x20
Fig.nr.171. Lot 3. Infiltrat inflamator
peribronşiolar. Col HE, x10
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
19
Fig.nr.197. Lot 8. Hiperplazie a
tunicii medii vasculare. Col HE, x20
Fig.nr.192. Lot 7. Bronhiolă
terminală. Col. HE, x20
Fig.nr.188. Lot 6. Hiperplazia
musculaturii netede vasculare,
epiteliu bronşiolar puternic plisat. Col HE, x10
Fig.nr.203. Lot 9. Bronhiolă terminală. Col HE, x10
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
20
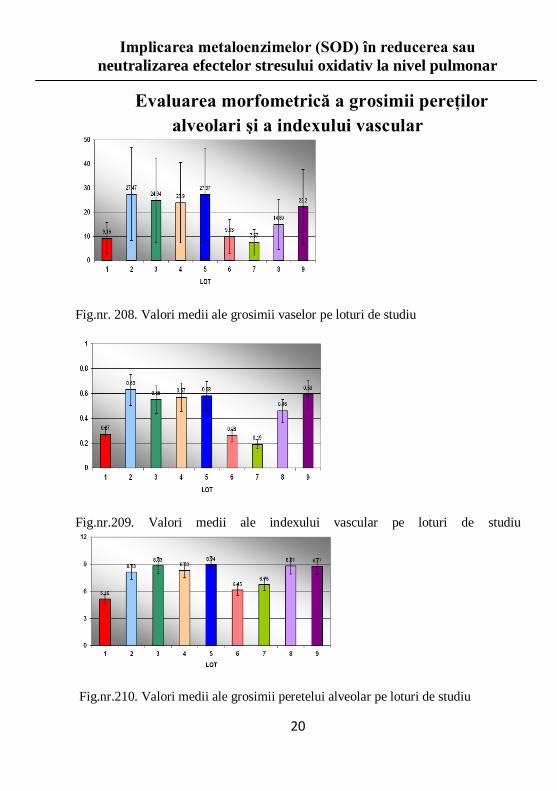
Evaluarea morfometrică a grosimii pereţilor
alveolari şi a indexului vascular
Fig.nr. 208. Valori medii ale grosimii vaselor pe loturi de studiu
Fig.nr.209. Valori medii ale indexului vascular pe loturi de studiu
Fig.nr.210. Valori medii ale grosimii peretelui alveolar pe loturi de studiu
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
21
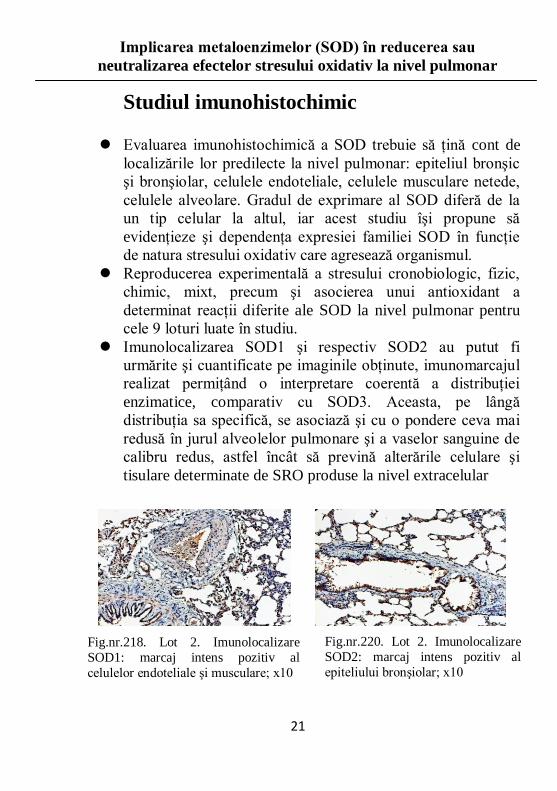
Studiul imunohistochimic
Evaluarea imunohistochimică a SOD trebuie să ţină cont de
localizările lor predilecte la nivel pulmonar: epiteliul bronşic
şi bronşiolar, celulele endoteliale, celulele musculare netede,
celulele alveolare. Gradul de exprimare al SOD diferă de la
un tip celular la altul, iar acest studiu îşi propune să
evidenţieze şi dependenţa expresiei familiei SOD în funcţie
de natura stresului oxidativ care agresează organismul.
Reproducerea experimentală a stresului cronobiologic, fizic,
chimic, mixt, precum şi asocierea unui antioxidant a
determinat reacţii diferite ale SOD la nivel pulmonar pentru
cele 9 loturi luate în studiu.
Imunolocalizarea SOD1 şi respectiv SOD2 au putut fi
urmărite şi cuantificate pe imaginile obţinute, imunomarcajul
realizat permiţând o interpretare coerentă a distribuţiei
enzimatice, comparativ cu SOD3. Aceasta, pe lângă
distribuţia sa specifică, se asociază şi cu o pondere ceva mai
redusă în jurul alveolelor pulmonare şi a vaselor sanguine de
calibru redus, astfel încât să prevină alterările celulare şi
tisulare determinate de SRO produse la nivel extracelular
Fig.nr.218. Lot 2. Imunolocalizare
SOD1: marcaj intens pozitiv al
celulelor endoteliale şi musculare; x10
Fig.nr.220. Lot 2. Imunolocalizare
SOD2: marcaj intens pozitiv al
epiteliului bronşiolar; x10
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
22
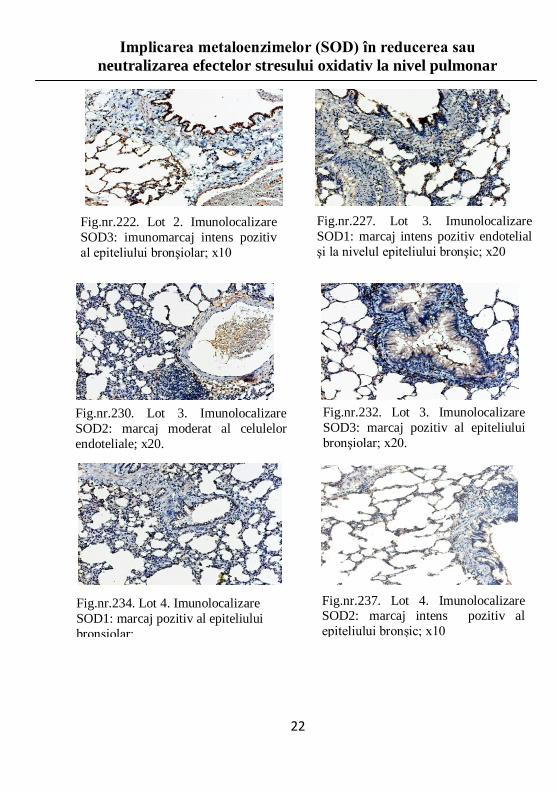
Fig.nr.222. Lot 2. Imunolocalizare
SOD3: imunomarcaj intens pozitiv
al epiteliului bronşiolar; x10
Fig.nr.227. Lot 3. Imunolocalizare
SOD1: marcaj intens pozitiv endotelial
şi la nivelul epiteliului bronşic; x20
Fig.nr.230. Lot 3. Imunolocalizare
SOD2: marcaj moderat al celulelor
endoteliale; x20.
Fig.nr.232. Lot 3. Imunolocalizare
SOD3: marcaj pozitiv al epiteliului
bronşiolar; x20.
Fig.nr.234. Lot 4. Imunolocalizare
SOD1: marcaj pozitiv al epiteliului
bronşiolar; x10
Fig.nr.237. Lot 4. Imunolocalizare SOD2: marcaj intens pozitiv al
epiteliului bronşic; x10
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
23
.
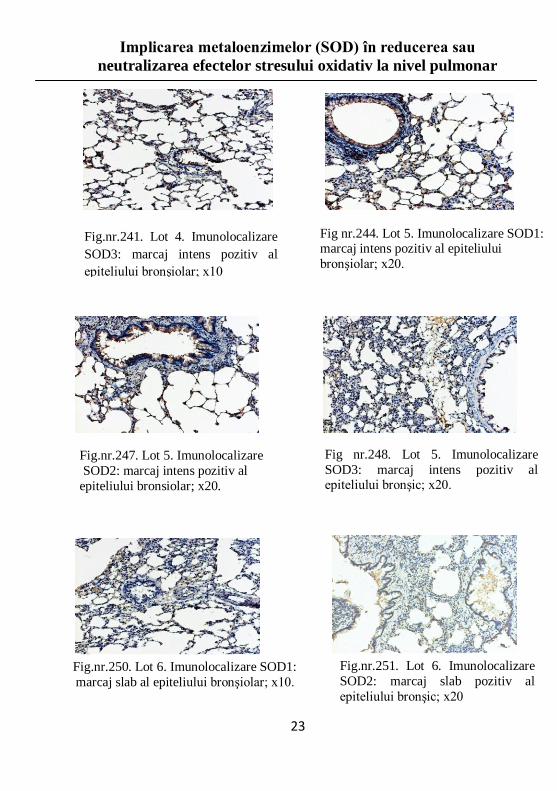
Fig.nr.241. Lot 4. Imunolocalizare
SOD3: marcaj intens pozitiv al
epiteliului bronşiolar; x10
Fig nr.244. Lot 5. Imunolocalizare SOD1: marcaj intens pozitiv al epiteliului
bronşiolar; x20.
Fig.nr.247. Lot 5. Imunolocalizare
SOD2: marcaj intens pozitiv al
epiteliului bronsiolar; x20.
Fig nr.248. Lot 5. Imunolocalizare
SOD3: marcaj intens pozitiv al epiteliului bronşic; x20.
Fig.nr.250. Lot 6. Imunolocalizare SOD1:
marcaj slab al epiteliului bronşiolar; x10.
Fig.nr.251. Lot 6. Imunolocalizare
SOD2: marcaj slab pozitiv al
epiteliului bronşic; x20
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
24
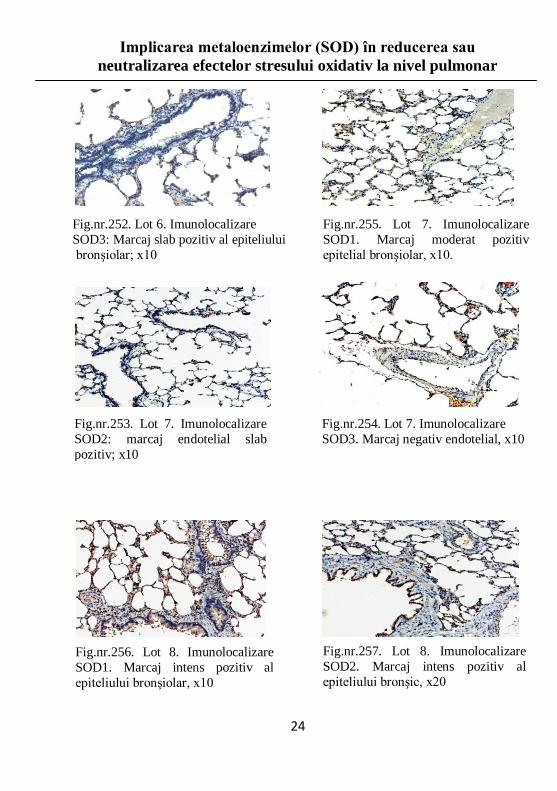
Fig.nr.252. Lot 6. Imunolocalizare
SOD3: Marcaj slab pozitiv al epiteliului
bronşiolar; x10
Fig.nr.255. Lot 7. Imunolocalizare
SOD1. Marcaj moderat pozitiv
epitelial bronşiolar, x10.
Fig.nr.253. Lot 7. Imunolocalizare
SOD2: marcaj endotelial slab
pozitiv; x10
Fig.nr.254. Lot 7. Imunolocalizare
SOD3. Marcaj negativ endotelial, x10
Fig.nr.256. Lot 8. Imunolocalizare
SOD1. Marcaj intens pozitiv al
epiteliului bronşiolar, x10
Fig.nr.257. Lot 8. Imunolocalizare
SOD2. Marcaj intens pozitiv al
epiteliului bronşic, x20
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
25
Evaluarea scorizării marcajului imunohistochimic
al SOD
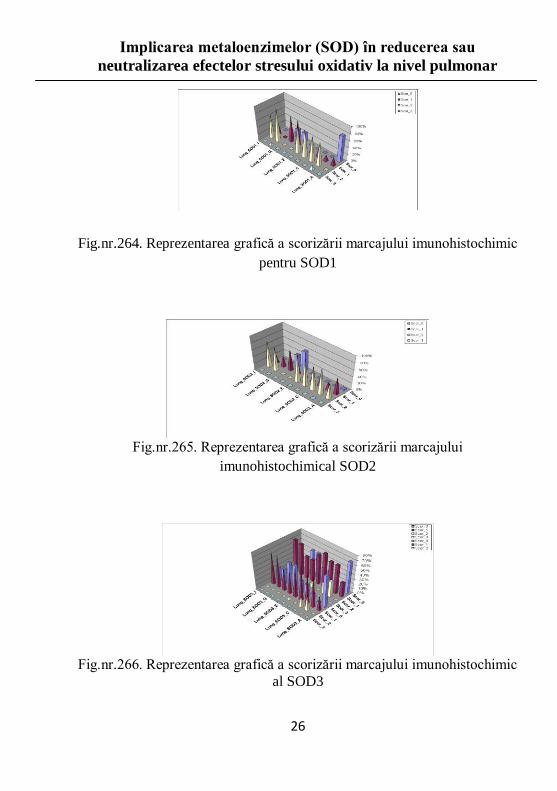
Respectând cele 4 scoruri impuse, respectiv 0, 1, 2 şi 3, am analizat
FOV-urile din imaginile în care am evidenţiat imunolocalizarea celor 3 SOD (Fig.nr.263-266).
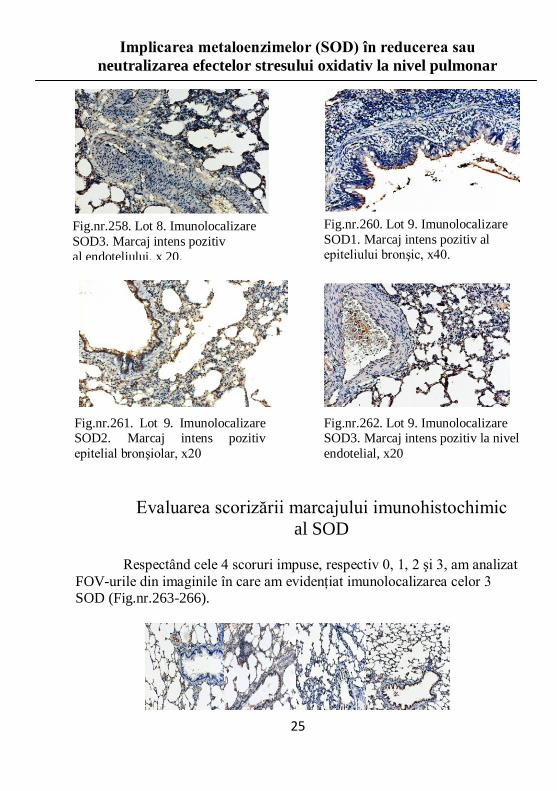
Fig.nr.258. Lot 8. Imunolocalizare
SOD3. Marcaj intens pozitiv
al endoteliului, x 20.
Fig.nr.260. Lot 9. Imunolocalizare
SOD1. Marcaj intens pozitiv al epiteliului bronşic, x40.
Fig.nr.261. Lot 9. Imunolocalizare SOD2. Marcaj intens pozitiv
epitelial bronşiolar, x20
Fig.nr.262. Lot 9. Imunolocalizare SOD3. Marcaj intens pozitiv la nivel
endotelial, x20
Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
26
Fig.nr.264. Reprezentarea grafică a scorizării marcajului imunohistochimic
pentru SOD1
Fig.nr.265. Reprezentarea grafică a scorizării marcajului
imunohistochimical SOD2
Fig.nr.266. Reprezentarea grafică a scorizării marcajului imunohistochimic
al SOD3

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
27
DISCUŢII
Plămânii reprezintă organe puternic afectate de generarea
SRO şi care şi-au dezvoltat un sistem de apărare antioxidantă în care
rolul dominant este deţinut de o familie de metaloenzime care a
progresat continuu odată cu trecerea la viaţa aerobă – SOD.
Produse în celulele aerobe în cursul proceselor oxidative
spontane sau induse, SOD sunt indispensabile protecţiei pulmonare
faţă de agresivitatea SRO generate pe parcursul unui stres oxidativ
datorită intervenţiei lor prompte pentru neutralizarea radicalilor
superoxid. Cum echilibrul dintre atacul SRO şi capacitatea
antioxidanta este extrem de sensibil la nivel pulmonar, orice
perturbare a acestuia determină instalarea unor modificări patologice.
Amploarea şi gravitatea acestora depind de regulă de sumarea
condiţiilor mediului intern şi respectiv extern în care are loc
agresiunea oxidativă.
Ideea care a stat la baza derulării acestei teze de doctorat a
pornit tocmai de la faptul că disponibilitatea antioxidantă enormă pe
care o prezintă SOD nu este cuantificată în strategii terapeutice
eficiente. Un prim pas în realizarea acestui proces îl reprezintă
identificarea tipului majoritar de SOD care se produce de exemplu la
nivel pulmonar în raport de tipul de stres oxidativ care se exercită
asupra organismului. Ulterior, în condiţiile unui stres similar se poate
stimula producţia tipului de SOD, fie direct, fie indirect, utilizând
SOD mimetice, diminuând sau poate chiar anulând astfel impactul
oxidativ.
În studiul nostru experimental am inclus o paletă diversă de
stres oxidativ – cronobiologic sau fotoperiodic, fizic, chimic şi mixt,
pe parcursul cărora am investigat pricipalii markeri ai stresului
oxidativ (SOD, GPx, Catalaza, MDA), pentru a confirma existenţa şi
producerea reacţiilor oxidative, precum şi modul de variaţie a
parametrilor biologici (pentru a evidenţia reactivitatea organismului
la stres), după care am evaluat modificările histopatologice şi am

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
28
realizat imunolocalizarea celor trei tipuri de SOD pentru fiecare
formă de stres oxidativ indus.
Evaluarea statusului biologic al organismului prin
monitorizarea unor constante specifice (greutate, parametri sanguini)
ne-a permis următoarele observaţii:
-Analiza greutăţii la nivelul celor 9 loturi luate în studiu a
evidenţiat o disociere pe două direcţii distincte a greutăţii animalelor
investigate. Pe de o parte, asistăm la o scădere drastică a greutăţii la
animalele din loturile 2, 5, 8 şi 9. Pe de altă parte, se observă o
creştere a greutăţii la animalele din loturile 1, 4, 6, 7.
-Analiza parametrilor sanguini a permis evaluarea
următoarelor rezultate:
Hematiile au prezentat o creştere clară la loturile 8 şi 9, în
paralel cu o scădere relativă la loturile 4, 5 şi 7.
globulele albe prezintă o scădere a valorilor medii la loturile
2, 5, 8 şi 9. O creştere se observă la loturile 3, 4, 6 şi 7.
trombocitele, spre deosebire de celelalte elemente sanguine
analizate, cu excepţia lotului martor, au prezentat o scădere
variabila.
hematocritul – analiza sa a evidenţiat o creştere marcată la
loturile 1, 2, 6, 8 şi 9, precum şi o scădere la loturile 3, 4, 5 şi
7. Aceste rezultate se corelează cu cele obţinute în evaluarea
hematiilor, la loturile luate în studiu.
Studiul histopatologic a evidenţiat gradul şi intensitatea
alterărilor tisulare pulmonare în funcţie de tipul de stres oxidat iv
indus. Disfuncţiile pulmonare care se instalează compromit funcţia
de barieră a endoteliului pulmonar, ca o consecinţă directă a
răspunsului inflamator.
Cele mai multe date din literatura de specialitate furnizează
informaţii asupra degradării oxidative a căilor aeriene mari, care au
putut fi relativ uşor investigate şi abordate în studii experimentale.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
29
Studiul nostru experimental cuantifică şi analizează modalitatea de
reacţie a căilor aeriene în ansamblu, insistând pe modificările
survenite în special la nivelul căilor respiratorii de calibru redus.
Acurateţea investigării modificărilor pulmonare produse a
fost asigurată de numeroasele câmpuri investigate şi evaluate atât
histopatologic cât şi morfometric şi imunohistochimic. Într-un
sinopsis al reactivităţii pulmonare faţă de stresul oxidativ menţionat
în literatura de specialitate sunt incluse drept modificări definitorii:
prezenţa infiltratelor inflamatorii pe traseul căilor respiratorii,
cu modificările consecutive, precum şi în interstiţiul
pulmonar;
hiperplazii ale celulelor epiteliului respirator, care vor
delimita lumene bronşice şi bronşiolare puternic faldate;
distrucţia pereţilor alveolari, cu distensia consecutivă a
spaţiilor alveolare;
modificarea calibrului vaselor sanguine adiacente teritoriilor
pulmonare modificate, prin hiperplazia musculaturii netede
de la nivelul tunicii medii.
Aceste modificări se regăsesc într-o manieră cvasisincronă la
majoritatea loturilor examinate, pledând pentru o distribuţie
condiţionată însă de tipul de stres oxidativ indus. În ordinea
frecvenţei de localizare a infiltratelor inflamatorii la nivelul
conductelor respiratorii, lotul cel mai afectat este lotul 2, cel care a
fost expus la lumină continuă pe toată durata desfăşurării
experimentului, urmat de loturile 8 şi 9, supuse unui stres oxidativ
mixt, precum şi de lotul 3 deprivat de lumină, lotul 4, supus la efort
fizic prelungit şi respectiv lotul 5, căruia i s-a administrat
nitrofurantoin.
Studiul imunohistochimic a urmărit identificarea
imunolocalizării celor trei tipuri distincte de SOD şi corelarea cu
forma de stres oxidativ indus.
Scorizarea marcajului imunohistochimic la cele 9 loturi
luate în studiu a relevat o serie de observaţii legate de amplitudinea

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
30
imunolocalizării distincte pentru cele trei tipuri de SOD evaluate.
Utilizând tipuri diverse de stres oxidativ, am putut constata faptul că
fiecare dintre cele trei SOD se comportă specific, funcţie de stresul
indus.
Cea mai bine reprezentată a fost SOD1, care a fost localizată
distinct la toate loturile cărora li s-a indus un stres oxidativ. Cea de a
doua izoformă a SOD, SOD2, este mai puţin reprezentată
imunohistochimic, comparativ cu SOD1, dar la parametri care ne
permit o cuantificare şi o interpretare coerentă a rezultatelor obţinute.
SOD3 a fost cel mai puţin detectată prin marcaj imunohistochimic, în
concordanţă de altfel cu cele mai multe date din literatura de
specialitate.
În ciuda eficienţei lor în combaterea stresului oxidativ,
demonstrată de nenumărate studii experimentale, în prezent SOD nu
sunt incluse în arsenalul terapeutic antioxidant curent. Una dintre
motivaţiile prezentate este legată de faptul că SOD nu are o structură
atractivă din punct de vedere farmacologic. Apariţia SOD
mimeticelor ar putea rezolva şi această problemă.
Studiul nostru doreşte ca prin identificarea SOD produsă dominant în
raport cu forma de stres oxidativ la care este supus organismul, să
ofere o alternativă terapeutică modernă, prin stimularea electivă a
situsurilor care descarcă masiv SOD. Se asigură astfel o protecţie
antioxidantă semnificativă, prin mobilizarea resurselor proprii ale
parenchimului pulmonar.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
31
CONCLUZII
1. Studiul de faţă a realizat un model experimental de inducere a
mai multor forme de stres oxidativ, cu evaluarea calitat ivă a
SOD, fiind validat de modificările morfologice pulmonare
instalate, de evoluţia statusului biologic al animalelor de
laborator, de monitorizarea parametrilor stresului oxidativ,
precum şi de analizele statistice, imunohistochimice şi
morfometrice efectuate.
2. Evoluţia procentuală a SOD, GPx, catalazei şi MDA,
considerate drept markeri ai stresului oxidativ a prezentat o
creştere dinamică pe tot parcursul experimentului, comparativ
cu lotul martor, cele mai semnificative valori în dinamică
fiind cele pentru SOD şi catalază.
3. Creşterea valorilor SOD s-a corelat în mod direct cu scăderea
valorii medii a greutăţii animalelor în toate cele trei momente
ale studiului, la loturile 2, 3, 5, 8 şi 9, şi respectiv indirect cu
creşterea valorii medii a greutăţii animalelor la loturile 4, 6 şi
7.
4. Creşterea valorilor SOD s-a corelat în mod direct cu creşterea
valorii medii procentuale a hematiilor la loturile supuse
stresului fizic, asociat cu vitamina C, mixt, a globulelor albe
la loturile 6 şi 8, a trombocitelor la loturile supuse stresului
fotoperiodic, fizic şi stresului chimic şi a hematocritului la
loturile supuse stresului fotoperiodic, mixt şi a celui asociat
cu vitamina C.
5. Creşterea valorilor SOD s-a corelat indirect cu scăderea
valorii procentuale a hematiilor la loturile supuse stresului
fotoperiodic, asociat cu vitamina C şi respectiv mixt, precum
şi cu scăderea valorii medii procentuale a leucocitelor la
loturile supuse stresului fotoperiodic, chimic, mixt, cu
scăderea valorii medii a trombocitelor la loturile supuse
stresului fotoperiodic, asociat cu vitamina C şi mixt şi cu

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
32
scăderea hematocritului la loturile supuse stresului
fotoperiodic şi mixt.
6. Examenul histopatologic a relevat prezenţa la toate loturile
supuse stresului oxidativ a elementelor definitorii ale
agresiunii oxidative la nivel pulmonar: infiltrate inflamatorii
cu distribuţie variabilă, dependentă de tipul de stres, distrucţii
ale pereţilor alveolari cu distensii consecutive ale spaţiilor
alveolare, lumene bronşice şi bronşiolare stelate, neregulate,
datorită hiperplaziei celulelor din epiteliul respirator.
7. Procesele inflamatorii, în ordinea frecvenţei şi amplitudinii
lor, au afectat lotul expus la lumină continuă, urmat de
loturile supuse unui stres oxidativ mixt, lotul deprivat de
lumină şi cel supus efortului fizic şi apoi a celui supus unui
stres chimic. Distrucţiile pereţilor alveolari, în ordinea
frecvenţei şi amplitudinii, au afectat loturile expuse stresului
fotoperiodic, urmat de cel chimic, mixt şi fizic. Hiperplaziile
celulelor epiteliale mucosecretorii au apărut cvasisincron,
moderat, la toate loturile supuse stresului oxidativ. Modificări
de calibru vascular pe seama hiperplaziei tunicii musculare
apar în ordinea frecvenţei la lotul supus efortului fizic,
stresului chimic, stresului mixt şi celui fotoperiodic.
8. Studiul imunohistochimic a relevat imunolocalizarea celor
trei tipuri distincte de SOD la nivelul situsurilor specifice de
la nivelul ţesutului pulmonar şi corelarea acestora cu forma
de stres oxidativ indus asupra fiecărui lot de şobolani.
9. Scorizarea marcajului imunohistochimic realizat pe toate cele
9 loturi de şobolani ne-a permis în primul rând o ierarhizare
per ansamblu a producerii de SOD. Astfel, SOD1 a fost cea
mai bine reprezentată şi mai facil de observat la nivel
pulmonar, urmată de SOD2 şi în final, de SOD3.
10. SOD1 a fost identificată imunohistochimic şi direct corelată
în experimentul nostru, în ordine, cu următoarele forme de
stres oxidativ indus: fotoperiodic, mixt, fizic, chimic.
Localizările de elecţie ale SOD1 raportate la tipurile de stres

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
33
sus menţionate au fost: epiteliul bronşic, epiteliul bronşiolar,
celule endoteliale, celule musculare netede.
11. SOD2 a fost identificată imunohistochimic şi direct corelată
în experimentul nostru, în ordine, cu următoarele forme de
stres oxidativ indus: fizic, chimic, mixt, fotoperiodic.
Localizările de elecţie ale SOD2 raportate la tipurile de stres
induse: epiteliul bronşiolar, endotelii, celule musculare
netede, epitelii bronşice.
12. SOD3 a fost identificată imunohistochimic şi direct corelată
în experimentul nostru, în ordine, cu următoarele forme de
stres: fizic, mixt, chimic. Localizările de elecţie ale SOD3
raportate la tipurile de stres induse au fost: epiteliul
bronşiolar, endotelii, celule musculare netede.
13. Evaluarea SOD nu a putut fi realizată decât în contextul
specific, definitoriu al stresului oxidativ, în condiţiile în care
SOD sunt considerate principalele repere ale combaterii
agresiunii oxidative.
14. Construirea acestui model experimental axat pe studiul
calitativ şi dinamic al SOD raportat la tipul de stres oxidativ
indus ne-a permis să afirmăm că există o deplină corelaţie
între identificarea principalelor situsuri pulmonare implicate
în producţia SOD. şi natura stresului oxidativ indus.
15. Rezultatele obţinute ne îndreptăţesc să considerăm că
observaţiile noastre legate de producerea distinctă a SOD în
funcţie de tipul de stres oxidativ pot constitui un prim pas în
conturarea unei noi abordări terapeutice individualizate a
apărării antioxidante, la nivelul unuia dintre cele mai
vulnerabile teritorii din organism. Stimularea ţintită a
producţiei de SOD în zona în care amplitudinea
imunolocalizării sale, detectată experimental, este maximă ar
avea capacitatea unei intervenţii prompte şi strategic dirijate,
care ar putea deschide în sfârşit perspectivele utilizării
practice a enormului potenţial antioxidant al SOD.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
34
BIBLIOGRAFIE SELECTIVA
1. Kohen R, Nyska A. Review: Oxidation of Biological systems:
Oxidative Stress Phenomena, Antioxidants, Redox Reactions and
Methods for their Quantification. Toxicol Pathol 2002; 30: 620-
24.
2. Rocha M, Esplugues JV, Hernandez-Mijarez A, Victor M.
Mithocondrial-targeted directly Antioxidants and oxidative
stress: a proteomic prospective study. Curr Pharm Design 2009;
15 (26): 3052-62.
3. Galli F, Piroddi M, Annetti C, Aisa C, Floridi E, Floridi A.
Oxidative Stress and Reactive Oxygen Species. Karger 2005;
149: 240-60.
4. Imlay J. Pathways of oxidative damage. Annu Rev Microbiol
2003; 57: 395-418.
5. Mittler R. Oxidative Stress, Antioxidants and Stress Tolerance.
Trend Sci 2007; 7 (9): 405-10.
6. Packer L, Cadenas E. Oxidants and antioxidants revisited. New
concepts of oxidative stress. Free Radic Res 2007; 4 (1): 951-2.
7. Miyata T, Takizawa S, van Ypersele de Strihou C. Hipoxia.
Intracellular sensors for oxygen and oxidative stress; novel
therapeutics targets. Am Physiol Cell Physiol 2011; 300 (2):
C226-C231.
8. Vertuani S, Angusti A, Manfredini S. The antioxidants and pro-
oxidants network; an overwiev. Oxidative balance from
chemistry to chemical medicine. Curr Pharm Design 2004;
1677-94.
9. Constantini D, Verhulst S. Does high antioxidant capacity
indicate low oxidative stress? Funct Biol 2009; 23: 506-9.
10. Fonseca RR, Johnson WE, O'Brien SJ, Vasconcellos V, Antunes
A. Molecular evolution and the role of oxidative stress in the
expansive and functional diversification of cytosolic glutathione
transferases. BMC Evol Biol 2010; 2 (41): 234-39.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
35
11. Dennery PA. Oxidative stress in development: nature or murture
? Free Radic Biol Med 2011; 2 (1): 814-19.
12. Maeda Y, Inoguchi T. Oxidative stress. Nippon Rinsho 2010; 69
(5): 814-18.
13. Dunyapoon T, Lu W, Ogasawara MA, Rivero-Del Vale N,
Huang P. Redox Regulation of Cell Survival. Antiox & Redox
Signalling 2008; 2: 1343-74.
14. Valko M, Leibfritz D, Moncol J, Cronin T, Mazur M, Telser J.
Free radicals and antioxidants in normal physiological functions
and human disease. Int J Biochem Cell Biol 2007; 39 (1): 44-84.
15. Adly AM. Oxidative Stress and Disease: An Updated Review.
Res J of Immunol 2010; 3 (2): 129-145.
16. Klaunig JE, Kamendulis L. The role of oxidative stress in
carcinogenesis. Annual Review of Pharmacol and Toxicol 2004;
18: 564-71.
17. Reiter JR, Duan-xian T, Korkmaz A, Jou M, Manchester LM,
Paredes SD. Biogenic amins In the reduction of oxidative stress.
J of Cell Biol 2008; 12: 98-104.
18. Rahman I, Biswas S, Kode A. Oxidant and antioxidant balance in
the airways and the airway diseases. Eur J Pharmacol 2006; 533
(1-3): 222-39.
19. Kaur A, Van PT, Busch C, Robinson C, Pan M, Lee Pang W,
Reiss D, DiRuggiero J, Balig NS. Coordination of frontline
defense mechanisms under severe oxidative stress. Mol Syst Biol
2010; 393 (6): 156-62.
20. Temple MD, Perrone G, Daves IW. Complex cellular responses
to reactive oxygen species. Trends Cell Biol 2005; 15: 319-26.
21. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Oxidative stress
and cell signaling. Curr Med Chem 2004; 11: 1163-182.
22. Li CY, Jackson RM. Reactive species mechanisms of cellular
hypoxia – reoxygenation Injury. Am J Physiol Cell Physiol 2002;
282: C227-C241.
23. Crapo JD. Oxidative stress as an initiator of cytokine release and
cell damage. Eur Resp J 2003; 22 (44): 936-45.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
36
24. Burton GJ, Jauniaux E. Oxidativ Stress. Best Practice & Res Clin
Obst & Gynecol 2011; 25 (3): 287-299.
25. Zheng J, Zhiping H, Liuwang Z, Wei L, Hainan Z, Li T, Xiao H.
The role of Golgi apparatus In oxidative stress: is this organelle
less significant than mitochondria? Free Rad Biol and Med 2011;
50 (8): 907-15.
26. Tkaczyk J, Vizek M. Oxidative Stress in the Lung Tissue -
Sources of Reactive Oxygen Species and Antioxidant Defence.
Prague Med Raport 2007; 108 (2): 105-114.
27. Morgan J, Liu Z. Crosstalk of reactive oxygen species and NF-
kB signaling. Cell Res 2011; 21: 103-115.
28. Kirkinezos IG, Moraes C. Reactive oxygen species and
mitochondrial diseases. Cell & Dev Biol 2001; 12: 449-56.
29. Taylor-Clark TE, Undem TB. Sensing pulmonary oxidative
stress. Resp Physiol & Neur 2011; 10 (15): 233-40.
30. Forman JH, Maiorino M, Ursini F. Signaling functions of
reactive oxygen species. Biochem 2010; 49: 835-842.
31. Mittler R, Vanderauwera S, Suzuki N, Miller G, Tognetti VB,
Vandepoele K, Gollery M, Shulaev V, Van Breusegem F.B. ROS
signaling: the new wave ? Cell & Mol Biol 2011; 16 (6):300-9.
32. Nathan C. Specificity of a third kind: reactive oxygen and
nitrogen intermediates in cell signaling. J Clin Invest 2003; 111:
769-778.
33. Lambeth J. NOX enzymes and the biology of reactive oxygen.
Nat Rev Immunol 2004; 4: 181-89.
34. Acker T, Fandrey J, Acker H. The good, the bad and the ugly in
oxygen sensing: ROS, cytochromes and prolyl- hydroxilases.
Cardiovasc Res 2006; 71: 195-207.
35. Finkel T. Oxidant signals and oxidative stress. Curr Opin in Cell
Biol 2003; 15 (2): 247-54.
36. Fruehauf P, Meyskens F. Reactive oxygen Species: A Breath of
Life or Death? Clin Cancer Res 2007; 13 (10): 789-95.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
37
37. Watal G, Sharma B, Rai PK, Rai A. LIBS-based Detection of
Antioxidant Elements. A New Strategy. Methods Mol Biol 2010;
594: 275-85.
38. Nesterenko TN, Aly H. Fetal and neonatal programming
evidence and clinical implications. Am J Peri Natol 2009; 26
(3):191-98.
39. Dennery PA. Oxidative stress in development. Nature or
murture? Free Radic Mol Biol 2000; 81: 12-19.
40. Fuji J. Protective Mechanisms Against Oxidative Stress. Roles of
Antioxidant Enzymes and Redox Regulation. Yamaguchi Med J
2001; 19(1): 65-74.
41. Gutteridge J.M, Halliwell B. Free radicals and antioxidants in the
year 2000. A historical look to the future. Ann N Y Acad Sci
2000; 899: 135-47.
42. Nathens AB, Neff MJ, Jurkovich GH, Klotz P, Farver K,
Ruzinski J, Radella F, Garcia I, Maier RV. Randomized,
prospective trial of antioxidant supplementation in critically ill
patients. Ann Surg 2002; 236: 814-22.
43. Halliwell B. The antioxidant paradox. Lancet 2000; 355: 1179-
1180.
44. Rubartelli A. Stress as an intercellular signal: the emergence of
stress associated molecular patterns (SAMP). Antiox & Redox
Signalling 2008; 1003-10.
45. Martin K, Barett JC. Reactive oxygen species as double-edged
swords in cellular processes: low- dose cell signaling versus
high- dose toxicity. Human and Exp Toxicol 2002; 71-75.
46. Black HS. Mechanisms of Pro- and Antioxidation. The J of Nutr
2004; 35 (11): 31695-31705.
47. Kaur A, Van Phu T, Bush C, Robinson C, Wyming PL, Reiss D,
Diruggiero J, Balga NS. Coordination of frontline defense
mechanisms under severe oxidative stress. Mol Biol Syst 2010; 6:
393-405.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
38
48. Kanazawa J. Elucidation of biological defense mechanism for
oxidative stress and drug Research. Science Link J 2004; 1122-
27.
49. Fisher-Wellman K, Bell K, Bloomer J. Oxidative stress and
antioxidant defense mechanisms linked to exercise during
cardiopulmonary and metabolic disorders. Oxid Med and Cell
Long 2009; 2 (1): 43-51.
50. Zamocky M, Koller F. Understanding the structure and function
of catalases: clues from molecular evolution and in vitro
mutagenesis. Prog Biophys Mol Biol 1999; 19-76.
51. Chelikani P, Fita I, Loewen P. Diversity of structures and
properties among catalases. Cell Mol Life Sci 2004; 61 (2): 192-
208.
52. Edeas M. Anti-oxidants, controversies and perspectives: How can
the failure of clinical studies using anti-oxidants be explained? J
Soc Biol 2009; 203 (3): 271-8.
53. Rocha M, Hernandez-MA, Garcia-Halpartida KM, Banuls C,
Bellod L, Victor VM. Mitochondria-targeted antioxidant
peptides. Curr Pharm Res 2010; 16 (28): 3124-31.
54. Zhang YH. Structure-activity relationships and rational design
strategies for radical - Scavenging antioxidants. Pharmacol Ther
2010; 45 (3): 987-95.
55. Noguchi N, Watanabe A, Shi H. Diverse functions of
antioxidants. Free Radic Res 2000; 3 (6): 809-17.
56. Nie Z, Liu KH, Zhong CJ, Wang LF, Yang Y, Tian Q, Liu Y.
Enhanced radical scavenge Activity by antioxidant-
functionalized gold nanoparticles: a novel inspiration for new
artificial antioxidants. Free Radic Mol Biol 2007; 43 (9): 1243-
54.
57. Krinsky NI. Mechanism of Action of Biological Antioxidants.
Exp Biology and Med 2010; 92(5): 399-411.
58. Stohs S, Bagchi D. Oxidative mechanisms in the toxicity of metal
ions. Free Radic Biol Med 1995; 18 (2): 312-36.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
39
59. Chaudiere J, Ferrari Illiou R. Intracellular Antioxidants: From
Chemical to Biochemical Mechanisms. Food and Chem Toxicol
1999; 37 (9-10): 949- 62.
60. Glantzounis GK, Tsimoyiannis EC, Kappas AM, Gakaris D. Uric
acid and oxidative stress. Curr Pharm Design 2005; 11 (32):
4145-51.
61. Meister A. Glutathione-ascorbic acid antioxidant system in
animals. The J of Biol Biochem 1994; 269 (13): 9397-400.
62. Ratman DV, Ankola DD, Bhardwaj D, Sahana V, Kumar RMV.
Role of antioxidants in Prophylaxis and therapy: A
pharmaceutical perspective. J of Contr Rel 2006; 3:189-207.
63. Schwedhelm E, Maas RM, Troost R, Boger RH. Clinical
Pharmacokinetics of Antioxidants And their Impact on Systemic
Oxidative Stress. Clin Pharm 2003; 42 (5): 437-59.
64. Linster CL, Van Schaftingen E. Vitamin C. FEBS J 2007; 274
(1): 1-22.
65. Shanq F. Vitamin C and vitamine E restore the resistance of
GSH-depleted lens cells to H2O2. Free Radic Biol Mol 2003; 34:
521-30.
66. Schalch W. Antioxidants, Pro-oxidants and their Effects Reply.
JAMA 1994; 272-80.
67. Khan MA, Zhang J. Antioxidant enzymes and cancer. Chin J
Cancer Res 2010; 87-92.
68. Duarte TL, Lunec J. Review: When is an antioxidant not an
oxidant?A review of novel Antioxidant actions and reactions.
Free Rad Res 2005; 671-86.
69. Wilcox K, Ash SL, Catignani G. Antioxidants and prevention of
chronic diseases. Crit Rev in Food Sc and Nutr 2004; 44: 275-
295.
70. Mann T, Keillin D. Haemocuprein and hepatocuprein, copper
protein compounds of blood and liver in mammals. Proc R Soc
Ser B 1938; 128: 303-315.
71. Pauling L. The discovery of the superoxide radical. Trends
Biochem Sci 1979; N270-N271.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
40
72. McCord J, Fridovich I. The reduction of cytochrome c by milk
xanthine oxidase. J Biol Chem 1968; 5753-5760.
73. Nozyk-Grayck E, Suliman H, Piantadosi C. Extracellular
Superoxide Dismutase. Int J Cell Biochem 2005; 37 (12): 2466-
71.
74. Johnson F, Giulivi C. Superoxide Dismutases and their impact
upon human health. Mol Asp Med 2006; 340-52.
75. Bannister J, Bannister W, Rotilio G. Aspects of the structure,
functions and applications of superoxide dismutases. CRC Crit
Rev Biochem 1987; 111-80.
76. Fridovich I. The trail to superoxide dismutase. Protein Sci 1998;
7 (120: 2688-2690.
77. Gusev VA, Panchenko L. Superoxide radical and superoxide
dismutase in the free-radical theory of aging. Vopr Med Khim
1982 Jul-Aug; 28(4): 8-25.
78. Lee YM, Friedman DH, Avala V. Superoxide dismutase: An
evolutionary puzzle. Proc Natl Acad Sci USA 1995; 82: 824-28.
79. Brawn K, Fridovich I. Superoxide radical and superoxide
dismutases: threat and defense. Acta Physiol Scand Suppl 1980;
492: 9-18.
80. Cizewski Culotta V. Superoxide Dismutase, Oxidative Stress and
Cell Metabolism. Curr Topics in Cellular Regul 2001; 36: 117-
132.
81. Miller AM. Superoxide dismutases: active sites that save, but a
protein that kills. Curr opin in Chem Biol 2004; 8: 162-158.
82. Landis G, Tower J. Superoxide dismutase evolution and life span
regulation. Mech Ageing Dev 2005; 365-79.
83. Batinic-Haberle M, Reboucas S, Spasojevic I. Superoxide
dismutase mimics: chemistry, Pharmacology and therapeutic
potential. Antiox & Redox Signal 2010; 877-918.
84. Omar A, Flores SM, McCord J. Superoxide dismutase:
Pharmacological Development And Applications. Adv in
Pharmacol 2008; 109-120.
85. Muth CM. Antioxidant enzymes. Free Rad Res 2004; 927-32.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
41
86. Perry J, Shin S, Getzoff D, Tainer A. The structural biochemistry
of superoxide dismutase. Biochim Biophys Acta 2010; 1011-21.
87. Marikovsky M, Ziv V, Nevo I, Harris-Cerruti C, Mahler O.
Cu/Zn Superoxide Dismutase Plays Important Role In Immune
Response. The J of Immunol 2003; 2993-3001.
88. Song Y, Leonard S, Traber MG, Ho E. Zinc deficiency affects
DNA damage, oxidative Stress, antioxidant defenses, and DNA
repair in rats. The J of Nutr 2009; 1626-31.
89. Zelko N, Mariani J, Folz R. Superoxide dismutase multigene
family: comparison between CuZn SOD (SOD1), MnSOD
(SOD2), and EC-SOD (SOD3) gene structures, evolution and
expression. Free Rad Biol & Med 2000; 337-349.
90. Jeney V, Itoh H, Wendt M, Gradek Q, Ushio-Fukai M, Harrison
D, Fukai T. The action of SOD family on superoxide anion. Fre
Rad Res 2007; 1227-34.
91. Chin NH, Pan W. Assay of superoxide dismutase activity by
combining electrophoresis And densitometry. Biol Bul Acad Sci
1996; 107-111.
92. Poberezkina NB, Osinskaia LF. The biological role of superoxide
dismutase. Ukr Biokim Zh 1989; 14-27.
93. Blokhina O, Virolainen E, Fagerstedt VM. Antioxidants,
Oxidative Damage and Oxygen Deprivation Stress: A rewiev.
Annals of Biol 2003; 179-94.
94. Matzinger P. The danger model: a renewed sense of self. Science
2002; 296: 301-305.
95. Kiefer D. Superoxide dismutase-boosting the body's primary
antioxidant defense. Life ext 2003; 170-76.
96. Ames BN, Shigenaga TK, Hagen M. Oxidants, antioxidants and
the degenerative disease of aging. Proc natl Acad Sci USA 1993;
7915- 7922.
97. Fleury C, Mignotte BH, Vaysiere JL. Mitochondrial reactive
oxygen species in cell death Signaling. Biochimie 2002; 131-141.
98. Fisher-Wellman K, Bell I,. Bloomer J. Oxidative stress and
antioxidant defense mechanism Linked to exercise during

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
42
cardiopulmonary and metabolic disorders. Ox Med and Cell long
2009; 43-51.
99. Kinnula VL, Crapo JD. Superoxide dismutases in the lung and
human diseases. Am J Res Crit Care Med 2003; 1600-19.
100. Cristoufidu-Solomidou M, Muzykantov V. Antioxidant
strategies in respiratory medicine Treatment in Resp Med 2006; 5
(1): 47-78.
101. Piotrowski WJ, Marczack J. Cellular sources of antioxidants
in lung. Int J Occup Med Environ Health 2000; 369-85.
102. Orient A, Donko A, Szabo A, Leto M, Geiszt G. Novel
sources of reactive oxygen species in the human body. Nephrol
Transplant 2007: 1281-1288.
103. Chow WC, Abreu MT, Suzuki T, Downey G. Oxidative
stress and acute lung injury. Am J of Resp Cell and Mol Biol
2003; 427-31.
104. Kuwano K, Nakashima N, Inoshima I, Hagimoto N, Fujita N,
Yoshimi N, Maeyama T, Hamada N, Watanabe K, Hara N.
Oxidative stress in lung epithelial cells from patients with
Idiopathic interstitial pneumonias. Eur Resp J 2003; 230-240.
105. Fuchs B, Sommer N, Dietrich A, Schermuly TW, Ghofrani S,
Grimminger F, Seeger W Gudermann T, Weissman NI. Redox
signaling and reactive oxygen species in hypoxic lung
Vasoconstriction. Resp Physiol & Neurobiol 2010; 282-291.
106. Noor R, Mittal S, Izbal J. Superoxide dismutase-applications
and release to human disease. Med Sci Monit 2007; R210-R215.
107. Caironi P, Langer TM, Gattinoni L. Acute lung injury/acute
respiratory distress syndrome Patophysiology. Curr Opin Crit
Care 2008; 64-69.
108. Shapiro S. The Patophysiology of COPD: What Goes Wrong
and Why? Resp Proc 2003; 593-598.
109. Hautamaki RD, Kobayashi DK, Senior RMW, Shapiro SD.
Requirement of macrophage Elastase for cigarette smoke induced
empnysema in mice. Science 2003; 223S-227S.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
43
110. Chung KF. The role of airway smooth muscle in the
pathogenesis of airway remodeling in COPD. Proc Am Thorac
Soc 2007; 347-349.
111. Larkin EK, Pingsheng W, Gebretsadik TW, Roberts JL,
Gross M, Hartert T. Antioxidant Defense and Adult-Onset
Incident Asthma: The Shanghai Womens Asthma and Allergy
Study. Resp and Crit Care Med 2010; 813-17.
112. Misso NL, Thompson PJ. Oxidative stress and antioxidant
deficiences in asthma: Redox Rep 2005; 247-55.
113. Nadeem A. Oxidant–antioxidant imbalance in asthma:
scientific evidence, epidemiological data and possible therapeutic
options. Ther Adv in Resp Diseases 2008; 4215-23.
114. Cho YS, Moon BY. The Role of Oxidative Stress In the
Pathogenesis of Asthma. Allergy Asthma Immunol Res 2010;
183-187.
115. Sjodin T, Westing I, Apple FH. Biochemical mechanisms for
oxygen free radical formation during exercise. Sports Med 1990;
10: 236-254.
116. Zerba E, Komorowski TE, Faulkner J. Free radical injury to
skeletal muscle of young, adult old mice. Am J Physiol Cell
Physiol 1990; C429-C435.
117. Ji L, La JH. Antioxidant defense: effects of aging and
exercise. Free Rad In Exercise and Ageing. Human Kinetics
2000; 35-72.
118. Dillard JC, Litov RE, Sawin WM, Dumelin EE, Tappel A.
Effects of exercise, vitamin E, ozone on pulmonary function and
lipid peroxidation. J Appl Physiol 1978; 927-832.
119. Quintanilha AT. Effects of physical exercise and vitamin E
on tissue oxidative metabolism. Biochem Soc Trans 1988; 56-69.
120. Niess AM, Simon PW. Response and adaptation of skeletal
muscle to exercise-the role of Reactive oxygen species. Front
Biosci 2007; 4826-4838.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
44
121. Radak Z, Taylor A, Ohno H, Goto S. Adaptation to exercise-
induced oxidative stress; from Muscle to brain. Exerc Immunol
Rev 2001; 90-107.
122. Novelli G, Braciotti G, Falsini S. Spin-trappers and vitamin E
prolong endurance to muscle fatigue in mice. Free Radic Biol
Med 1990; 9-13.
123. Lerch A. Biological rhythma in the context of light at night.
Neuro Endocrinol Let 2002; 23-27.
124. Sangani RG, Ghio AJ. Lung injury after cigarette smoking is
particle related. Int J Chron Obstruct Pulmon Dis 2011; 6: 191-8.
125. Pan L, Fu JH, Xue XD, Xu W, Zhou P, Wei B. Melatonin
protects against oxidative damage in a neonatal rat model of
bronchopulmonary dysplasia. World J Pediatr 2009; 5 (3): 216-
21.
126. Barnes PJ. Mechanisms and resistance in glucocorticoid
control of inflammation. J Steroid Biochem Mol Biol 2010; 120
(2-3): 76-85.
127. Stroescu V. Bazele farmacologice ale practicii medicale,
Editura Medicală, 1999.
128. Olsen KC, Sapinoro RE, Kottmann RM, Kulkarni AA, Iismaa
SE, Johnson GV, Thatcher TH, Phipps RP, Sime PJ.
Transglutaminase 2 and its Role in Pulmonary Fibrosis. Am J
Respir Crit Care Med 2011 Jun 30 .
129. Qi J, Liu Y, Li Q, Chen X. Effect of ginkgo biloba extract
against pulmonary fibrosis and its mechanisms. Zhongguo Zhong
Yao Za Zhi 2010; 35 (22): 3043-7.
130. Martins RR, Marchiori E, Viana SL, Grillo Júnior LS,
Capelozzi VL, Valença LM Chronic eosinophilic pneumonia
secondary to long-term use of nitrofurantoin: high-resolution
computed tomography findings. J Bras Pneumol 2008; 34 (3):
181-4.
131. Babin AL, Cannet C, Gérard C, Wyss D, Page CP, Beckmann
N. Noninvasive assessment of bleomycin-induced lung injury

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
45
and the effects of short-term glucocorticosteroid treatment in rats
using MRI. J Magn Reson Imaging 2011; 3 (3): 603-14.
132. Namagondlu G, Low SE, Seneviratne R, Banerjee A. Acute
renal failure from nitrofurantoin-induced acute granulomatous
interstitial nephritis. QJM 2010; 103 (1): 49-52.
133. Witten CM. Pulmonary toxicity of nitrofurantoin. Arch Phys
Med Rehabil 1989; 70 (1): 55-7.
134. Freise J, Wehmeier P, Welte T. Drug-induced pulmonary
interstitial disease. Dtsch Med Wochenschr 2010; 135 (10): 450-
4.
135. Hargett CW, Sporn TA, Roggli VL, Hollingsworth JW. Giant
cell interstitial pneumonia associated with nitrofurantoin. Lung
2006; 184 (3): 147-9.
136. Targowski T, Jahnz-Rózyk K. Immunomodulatory properties
of macrolides. Pol Merkur Lekarski 2008; 25 (149): 408-11.
137. Hardeland R, Coto-Montes A, Poeggeler B. Circadian
rhythms, oxidative stress, and antioxidative defense mechanisms.
Chronobiol Int 2003 Nov; 20 (6): 921-62.
N.B. Teza de doctorat conţine 355 de titluri bibliografice.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
46
LUCRĂRI ŞTIINŢIFICE PUBLICATE DIN TEMA
TEZEI DE DOCTORAT
IN EXTENSO
1. Raluca Oana Temneanu, Carmen Zamfir. Oxidative stress -
Free radicals and antioxidants. Farmacologia de la
experiment la clinic. Ed. Junimea, 2008; 266-73.
2. Raluca Temneanu, Lelioara Tocan, Elena Cojocaru,
Cătălina Lupuşoru, Carmen Zamfir. Corelaţii între
intensitatea stresului oxidativ şi variabilitatea afectării
parenchimului pulmonar. Revista Română de Anatomie
funcţională şi clinică, macro- şi microscopică şi de
Antropolgie 2009, vol.VIII, nr.1, 98-101.
3. Carmen Zamfir, Raluca Temneanu, Lelioara Tocan, Elena
Cojocaru, Cristina Galeş. Evaluarea imunohistochimică a
SOD eliberate în stresul oxidativ la nivel pulmonar. Revista
Română de Anatomie funcţională şi clinică, macro- şi
microscopică şi de Antropolgie 2010; vol. IX, nr.1, 51-54.
4. Raluca Temneanu, Carmen Zamfir, F. Zugun, Elena
Cojocaru, Lelioara Tocan. Assessment of superoxide
dismutases (SOD) influence on pulmonary morphology
during continuous light exposure of experimental animals.
Revista Română de Anatomie funcţională şi clinică, macro- şi
microscopică şi de Antropolgie 2011; vol. X, nr.1, 22-26.
5. Raluca Oana Temneanu, Carmen Zamfir, F.Eloaie Zugun,
Elena Cojocaru, Lelioara Tocan. Oxidants and antioxidants
relevance in rats pulmonary induced oxidative stress. J of
Med and Life 2011; 2(3): 48-53.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
47
ÎN REZUMAT
1. Raluca Oana Temneanu, Carmen Zamfir, F. Eloaie Zugun,
Elena Cojocaru, Lelioara Tocan. Evaluarea morfometrică a
dispozitivului vascular pulmonar în stresul oxidativ indus prin
deprivare de lumină. Al X-lea Simpozion Naţional cu
participare internaţională de Morfologie Microscopică,
Craiova, 25-28 mai 2011, 236.
2. Carmen Lacramioara Zamfir, F.E.Zugun, Raluca Oana
Temneanu, Lelia Tocan, Elena Cojocaru. Are mesangial
cells reactive to nitrofurantoin when associated with an
oxidative stress ? 23rd
European Congress of Pathology,
Helsinki, 27 aug-7 sep 2011.

Implicarea metaloenzimelor (SOD) în reducerea sau
neutralizarea efectelor stresului oxidativ la nivel pulmonar
48







