G E N E T I C A G E N E R A L A Date de contact: Coordonator curs: Lector Dr. Mirela Mihaela Cimpeanu e-mail: [email protected]Tutore: Lector Dr. Mirela Mihaela Cimpeanu e-mail: [email protected]Obligatii minimale ale studentilor in vederea promovarii examenului la disciplina GENETICA GENERALA Parcurgerea intregii materii incluse in suportul de curs; Definirea principalelor notiuni de genetic si caracterizarea ultastructural – functionala a materialului genetic si a proceselelor si fenomenelor prezentate in suportul de curs; Comunicarea permanenta cu tutorele disciplinei in vederea elucidarii unor aspecte particulare si a intelegerii generale a materiei de curs; Prezenta directa la toate orele de lucrari practice de laborator; Definirea principiilor de baza si a scopului metodelor de laborator parcurse in cadrul orelor de lucrari practice; Insusirea etapelor de lucru ale metodelor de laborator parcurse; Dobandirea unei experiente generale practice de laborator; Corelarea cunostintelor dobandite din suportul de curs cu aplicatiile practice de laborator. Modul de stabilire a notei finale Nota finala la disciplina Genetica generala va fi acordata astfel: evaluarea activitatii desfasurate in cadrul orelor de lucrari practice: 50% examenul final la disciplina: 50% Teme de licenţă ce pot fi abordate la disciplina GENETICA GENERALA 1. Caracterizarea complementului cromosomial la diferite specii din flora spontana 2. Intocmirea de cariotipuri la diverite specii vegetale de interes alimentar, medical etc. 3. Caracterizarea complementului cromosomial la diferite specii din fauna piscicola 4. Studii comparative asupra efectelor unor mutageni fizici si/sau chimici asupra materialului genetic, la diferite organisme model 5. Studiul efectelor poluarii asupra materialului genetic la diferite organisme model
Transcript
G E N E T I C A G E N E R A L A
Date de contact: Coordonator curs: Lector Dr. Mirela Mihaela Cimpeanu e-mail: [email protected] Tutore: Lector Dr. Mirela Mihaela Cimpeanu e-mail: [email protected] Obligatii minimale ale studentilor in vederea promovarii examenului la disciplina GENETICA GENERALA
Parcurgerea intregii materii incluse in suportul de curs;
Definirea principalelor notiuni de genetic si caracterizarea ultastructural – functionala a materialului genetic si a proceselelor si fenomenelor prezentate in suportul de curs;
Comunicarea permanenta cu tutorele disciplinei in vederea elucidarii unor aspecte particulare si a intelegerii generale a materiei de curs;
Prezenta directa la toate orele de lucrari practice de laborator;
Definirea principiilor de baza si a scopului metodelor de laborator parcurse in cadrul orelor de lucrari practice;
Insusirea etapelor de lucru ale metodelor de laborator parcurse;
Dobandirea unei experiente generale practice de laborator;
Corelarea cunostintelor dobandite din suportul de curs cu aplicatiile practice de laborator.
Modul de stabilire a notei finale Nota finala la disciplina Genetica generala va fi acordata astfel:
evaluarea activitatii desfasurate in cadrul orelor de lucrari practice: 50%
examenul final la disciplina: 50%
Teme de licenţă ce pot fi abordate la disciplina GENETICA GENERALA
1. Caracterizarea complementului cromosomial la diferite specii din flora spontana 2. Intocmirea de cariotipuri la diverite specii vegetale de interes alimentar, medical etc. 3. Caracterizarea complementului cromosomial la diferite specii din fauna piscicola 4. Studii comparative asupra efectelor unor mutageni fizici si/sau chimici asupra materialului genetic, la diferite organisme model 5. Studiul efectelor poluarii asupra materialului genetic la diferite organisme model
Unitatea de invatare 1 1. Din istoricul cercetarilor de genetica
Termenul "genetică" a fost propus, la cel de-al III-lea Congres Internaţional de Hibridare şi Ameliorare a Plantelor (Londra), de către W. Bateson, cel ce avea să devină titularul primei catedre de profil din lume (catedra de Genetică de la Cambridge - Anglia, înfiinţată în 1909).
Civilizaţia Greciei antice ne-a lăsat mărturii, de o deosebită profunzime şi corectitudine, relative la fenomenul eredităţii. În privinţa omului, de pildă, grecii au reluat idei ale filosofiei indiene, idei de sorginte eugenică, în conformitate cu care omul poate fi îmbunătăţit dacă generaţiile succesive sunt asigurate de indivizi sănătoşi, inteligenţi, frumoşi etc.
Un punct de reper în orice analiză a antichităţii greceşti îl constituie momentul Aristotel (384-322 î.e.n.). Aristotel a pus bazele embriologiei, urmărind ontogenia puiului de găină - proces în care se pot face magistrale demostraţii relative la fenomenul eredităţii. In Roma antică şi-a etalat şi dezvoltat concepţiile Galenos Claudios (129-199). Renumitul medic a efectuat experimente şi observaţii, a adunat şi prelucrat date referitoare la procese de încrucişare la animale şi a ajuns la concluzia că cele două sexe au rol egal în transmiterea caracterelor ereditare.
Dupa Evul Mediu, toate acumulările de până atunci, combinate cu noile date furnizate de dezvoltarea fără precedent a agriculturii (producţia vegetală şi animală), a industriei (solicitantă a noi cantităţi şi a noi materii şi materiale) şi a relaţiilor interumane, au fundamentat noua concepţie ştiinţifică, noua poziţie a omului în cadrul naturii.
În anul 1651, W. Harwey, cel ce a descoperit şi a descris circulaţia sângelui, stabileşte un adevăr incontestabil şi incomensurabil ca valoare - toate vieţuitoarele provin din ouă fecundate. In 1665, în premieră mondială, R. Hooke (1635-1703) descrie celula. Simultan, Antony van Leeuwenhoeck (1632-1723) descoperă (cu microscopul perfecţionat de el însuşi) şi descrie multe microorganisme şi spermatozoizii de om, câine şi iepure.
La mai puţin de 50 de ani, adică în 1735, apare opera suedezului Carol Linné (1707-1778), Systema nature, operă fundamentală pentru teoreticieni şi practicieni întrucât, în numai 12 pagini,
oferea criterii clare de delimitare şi caracterizare a speciilor, obiectul concret de lucru pentru cei ce investigau anatomia, fiziologia, reproducerea şi comportamentul organismelor vegetale şi animale.
Un punct nodal în demersurile pentru o concepţie biologică modernă îl reprezintă descrierea nucleului celular de către Schneider, Schwann, Schleiden, Virchow. Mai precizăm că dacă diviziunea celulei animale fusese observată în 1827, iar cea a celulei vegetale (la o algă) în 1832, meioza a fost descrisă abia în 1870.
Dar, pentru o istorie a geneticii, este obligatorie prezentarea descoperirilor efectuate de Ch. Naudin care, în anul 1859, a publicat lucrarea Nouvelles recherches sur l'hibridité dans les vegetaux. În respectiva lucrare, Naudin sublinia aspecte importante ale transmiterii caracterelor ereditare, şi anume: uniformitatea hibrizilor din prima generaţie, diversitatea lor în generaţiile următoare, segregarea "esenţelor" (aşa numeşte Naudin factorii ereditari), revenirea la formele parentale etc. Naudin a lucrat cu specii ale genurilor Papaver, Primula, Datura, Nicotiana. Din păcate, el nu a reuşit să surprindă şi să enunţe legităţile transmiterii caracterelor, legităţile eredităţii. Acest merit îi va reveni lui Mendel.
2. Legile ereditatii 2.1. Johann Gregor Mendel (1822-1884) - fondatorul geneticii
Pregătit de o multitudine de descoperiri, observaţii şi experimente, diseminate pe întregul
parcurs al istoriei umane, momentul naşterii geneticii ca ştiinţă este momentul Mendel, mai precis, anul 1865, an în care opera sa Versuche über Pflanzenhybriden (Experienţe asupra
hibrizilor la plante, tradusă şi la noi, abia în 1945, de A. Piescu, în Buletinul Cultivării şi Fermentării Tutunului) a primit bun de tipar şi a fost publicată într-o modestă revistă provincială - "Revista Naturaliştilor din Brünn”.
Pentru a nu exista neclarităţi, precizăm: în 1900 copilul minune al biologiei a fost regăsit şi înscris în actele oficiale. Naşterea lui, însă, avusese loc în 1865.
2.2. Experimentele efectuate de Johann Gregor Mendel
Mendel a experimentat pe diverse specii de plante aparţinând genurilor Hieracium, Phaseolus şi Pisum, atenţie deosebită acordând însă mazării (Pisum sativum).
În primul rând, Mendel a ales ca obiect de studiu o specie autogamă - Pisum sativum. A
avut, astfel, posibilitatea de a urmări, în descendenţă, comportarea hibrizilor pe care i-a obţinut, în ritm şi în proporţii strict determinate.
În cel de al doilea rând, Mendel a făcut ceea ce nu au gândit predecesorii săi - a verificat puritatea genotipică a plantelor luate în studiu. Pentru a fi sigur, Mendel a verificat descendenţa fiecărui individ, timp de 3-4 generaţii.
Se impune, cu această ocazie, încă o remarcă. Alegerea mazării ca obiect pentru investigaţii a fost benefică deoarece:
specia este autogamă, alterarea rezultatelor prin polenizări necontrolate fiind exclusă; perioada de vegetaţie este scurtă; trăsăturile fenotipice contrastante (dominante-recesive) sunt distincte; castrarea plantelor este relativ uşoară; sunt multe soiuri în cadrul speciei, hibrizii dintre diferitele soiuri sunt fertili;
2.2.1. Monohibridarea
În multe manuale de biologie se afirmă că monohibridarea presupune încrucişarea între două organisme deosebite într-o singură pereche de caractere. Întrucât se pot naşte confuzii, grave uneori, supunem atenţiei următoarele aspecte ale problemei. Este vorba de manifestări diferite ale aceluiaşi caracter, nu de caractere diferite. Culoarea verde sau galbenă nu reprezintă caractere diferite, ci manifestări diferite (fenotipic) ale aceluiaşi caracter (caracterul culoare, în cazul de faţă). Prin urmare, monohibridarea reprezintă procesul de încrucişare (hibridare) a doi indivizi aparţinând aceleiaşi specii, genotipic puri (adică homozigoţi), deosebiţi între ei prin manifestarea fenotipică a unui singur caracter.
Mendel a utilizat, pentru monohibridare, două soiuri de mazăre: unul cu boabe de culoare galbenă şi altul cu boabe de culoare verde. Reamintim că, înainte de a efectua hibridarea, Mendel a perpetuat timp de 3-4 ani fiecare soi, spre a se convinge că este stabil (că ester genotipic pur, în terminologia genetică actuală). Recurgând la castrare şi fecundare artificială, în prima generaţie hibridă, notată cu F1 (adică prima generaţie filială - în latină filius, filia = “fiu, fiică”), a obţinut numai
plante cu boabe de o singură culoare - galbenă, în experimentul de acest tip. Întrucât şi în privinţa acestui aspect apar, deseori, confuzii, recomandăm revederea
noţiunilor de botanică şi precizăm: plantele ce au fost supuse castrării şi polenizării artificiale reprezintă generaţia parentală (desemnată în tabele şi grafice cu simbolul P), iar boabele apărute
Johann Gregor Mendel (1822-1884)
pe ele, în urma dublei fecundări prin care a luat naştere embrionul (diploid) şi endospermul (triploid), reprezintă generaţia filială, hibridă (desemnată prin F1).
Prima constatare a lui Mendel a fost: toţi hibrizii din prima generaţie aveau o singură culoare a boabelor. Experimentând cu alt caracter (pe aceeaşi specie), de pildă forma boabelor - netede şi zbârcite, a constatat că în F1 toţi indivizii aveau boabe netede. Pe baza acestui experiment, Mendel a enunţat principiul uniformităţii hibrizilor în F1.
Perpetuând respectivii hibrizi, prin autofecundare, în cea de-a doua generaţie Mendel a constatat reapariţia fenotipurilor parentale (reapariţia expresiilor fenotipice etalate de cei doi părinţi), în raport de aproximativ 3:1. În consecinţă, a fost enunţat cel de-al doilea principiu - segregarea hibrizilor în F2. Deci, Mendel a admis şi precizat că în F1 se manifestă fenomenul de dominanţă şi recesivitate, în sensul că este etalată doar una dintre posibilităţile de exprimare a caracterului (cea dominantă), cealaltă rămânând ascunsă (recesivă). Prin simboluri, experimentul efectuat de Mendel poate fi reprezentat astfel:
Părinţii Galben(●●) x Verde(○○) Hibridare
Gameţii (●) x (○)
F1 Galben(●○) Autofecundare
Gameţii ♂ (●) şi (○)
♀ (●) şi (○)
F2 1 galben (●●)+2 galben (●○)+1 verde(○○)
Segregare fenotipică 3:1
Segregare genotipică 1:2:1
Întrucât, la acea vreme, nu se cunoştea nimic despre cromosomi şi gene (substratul
material al informaţiei ereditare), Mendel a folosit termenul de factori ereditari şi a presupus, cu o
extraordinară intuiţie, că în fiecare celulă a organismului factorul ereditar determinant al unui caracter se află în doză dublă, în timp ce în celulele sexuale se află în doză simplă. Schematic, situaţia poate fi redată şi astfel:
Părinţii AA X aa
Gameţii A X a
F1 Aa Autofecundare sau hibridare de tipul frate x soră
Gameţi Produşi în F1
♀ A şi a
♂ A şi a
F2 AA 2Aa Aa
Segregare genotipică 1 2 1
Segregare fenotipică 3 : 1
Evident, prin A se desemnează factorul ereditar dominant şi prin a factorul ereditar recesiv.
Apoi Mendel a precizat că, după fecundare şi formarea zigotului, factorii ereditari nu se contopesc, ci doar se alătură. În consecinţă, când se formează din nou gameţii, ei se vor despărţi, fiecare trecând într-un gamet. Astfel, conchide Mendel, gameţii sunt puri din punct de vedere genetic. Aceasta este considerată prima lege elaborată de Mendel, supranumită: LEGEA PURITĂŢII GAMEŢILOR.
2.2.2. Dihibridarea
În a doua etapă a experimentului, Mendel a recurs la încrucişarea de indivizi care se deosebeau prin expresia fenotipică a două caractere - culoarea şi forma boabelor. El a luat soiul de mazăre cu boabe netede şi galbene, pe care l-a încrucişat cu soiul caracterizat prin boabe zbârcite şi verzi. În F1, ca şi la monohibridare, toţi indivizii au fost uniformi - boabe netede şi galbene. Deci, s-au manifestat fenomenele de dominanţă şi recesivitate. În F2 s-a manifestat segregarea, dar s-a constatat un lucru interesant: fiecare caracter segregă independent şi, în consecinţă, ester posibilă recombinarea caracterelor şi apariţia de noi fenotipuri. Forma boabelor a segregat în raport de 3:1. La fel şi culoarea boabelor. În plus, au apărut şi noi tipuri de plante - cu
boabe netede şi verzi, pe de o parte, şi cu boabe zbârcite şi galbene, pe de altă parte. Iată cum poate fi redat, schematic, procesul:
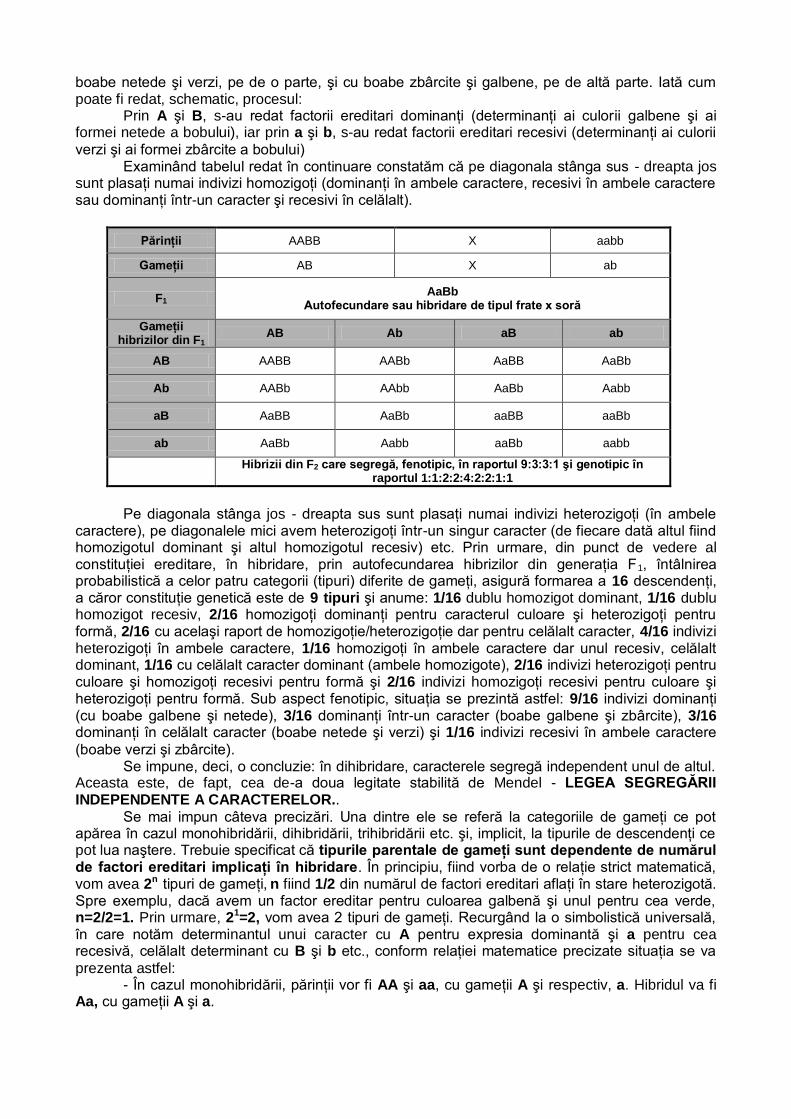
Prin A şi B, s-au redat factorii ereditari dominanţi (determinanţi ai culorii galbene şi ai formei netede a bobului), iar prin a şi b, s-au redat factorii ereditari recesivi (determinanţi ai culorii
verzi şi ai formei zbârcite a bobului) Examinând tabelul redat în continuare constatăm că pe diagonala stânga sus - dreapta jos
sunt plasaţi numai indivizi homozigoţi (dominanţi în ambele caractere, recesivi în ambele caractere sau dominanţi într-un caracter şi recesivi în celălalt).
Părinţii AABB X aabb
Gameţii AB X ab
F1 AaBb
Autofecundare sau hibridare de tipul frate x soră
Gameţii hibrizilor din F1
AB Ab aB ab
AB AABB AABb AaBB AaBb
Ab AABb AAbb AaBb Aabb
aB AaBB AaBb aaBB aaBb
ab AaBb Aabb aaBb aabb
Hibrizii din F2 care segregă, fenotipic, în raportul 9:3:3:1 şi genotipic în
raportul 1:1:2:2:4:2:2:1:1
Pe diagonala stânga jos - dreapta sus sunt plasaţi numai indivizi heterozigoţi (în ambele
caractere), pe diagonalele mici avem heterozigoţi într-un singur caracter (de fiecare dată altul fiind homozigotul dominant şi altul homozigotul recesiv) etc. Prin urmare, din punct de vedere al constituţiei ereditare, în hibridare, prin autofecundarea hibrizilor din generaţia F1, întâlnirea probabilistică a celor patru categorii (tipuri) diferite de gameţi, asigură formarea a 16 descendenţi, a căror constituţie genetică este de 9 tipuri şi anume: 1/16 dublu homozigot dominant, 1/16 dublu homozigot recesiv, 2/16 homozigoţi dominanţi pentru caracterul culoare şi heterozigoţi pentru formă, 2/16 cu acelaşi raport de homozigoţie/heterozigoţie dar pentru celălalt caracter, 4/16 indivizi heterozigoţi în ambele caractere, 1/16 homozigoţi în ambele caractere dar unul recesiv, celălalt dominant, 1/16 cu celălalt caracter dominant (ambele homozigote), 2/16 indivizi heterozigoţi pentru culoare şi homozigoţi recesivi pentru formă şi 2/16 indivizi homozigoţi recesivi pentru culoare şi heterozigoţi pentru formă. Sub aspect fenotipic, situaţia se prezintă astfel: 9/16 indivizi dominanţi (cu boabe galbene şi netede), 3/16 dominanţi într-un caracter (boabe galbene şi zbârcite), 3/16 dominanţi în celălalt caracter (boabe netede şi verzi) şi 1/16 indivizi recesivi în ambele caractere
(boabe verzi şi zbârcite). Se impune, deci, o concluzie: în dihibridare, caracterele segregă independent unul de altul.
Aceasta este, de fapt, cea de-a doua legitate stabilită de Mendel - LEGEA SEGREGĂRII INDEPENDENTE A CARACTERELOR..
Se mai impun câteva precizări. Una dintre ele se referă la categoriile de gameţi ce pot apărea în cazul monohibridării, dihibridării, trihibridării etc. şi, implicit, la tipurile de descendenţi ce pot lua naştere. Trebuie specificat că tipurile parentale de gameţi sunt dependente de numărul de factori ereditari implicaţi în hibridare. În principiu, fiind vorba de o relaţie strict matematică, vom avea 2n tipuri de gameţi, n fiind 1/2 din numărul de factori ereditari aflaţi în stare heterozigotă. Spre exemplu, dacă avem un factor ereditar pentru culoarea galbenă şi unul pentru cea verde, n=2/2=1. Prin urmare, 21=2, vom avea 2 tipuri de gameţi. Recurgând la o simbolistică universală, în care notăm determinantul unui caracter cu A pentru expresia dominantă şi a pentru cea recesivă, celălalt determinant cu B şi b etc., conform relaţiei matematice precizate situaţia se va
prezenta astfel: - În cazul monohibridării, părinţii vor fi AA şi aa, cu gameţii A şi respectiv, a. Hibridul va fi
Aa, cu gameţii A şi a.
- În cazul dihibridării, părinţii AABB şi aabb vor avea gameţi de tipul AB şi ab, hibridul fiind AaBb. De data aceasta însă, având în vedere legea segregării independente a caracterelor, gameţii vor fi AB, ab, Ab şi aB (deci, 2n=22=4 tipuri de gameţi).
2.3. Universalitatea legităţilor descoperite de Mendel
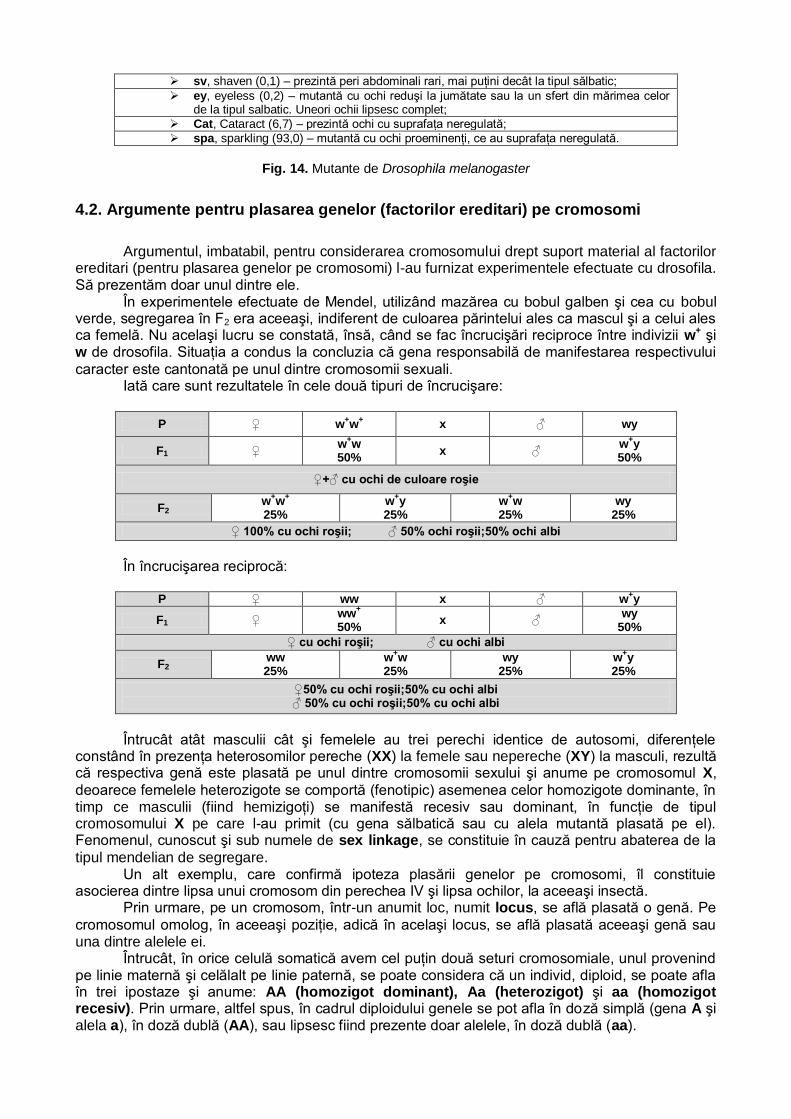
Evident, se pune problema dacă legităţile sesizate şi formulate de Mendel sunt universale, adică dacă sunt aplicabile totalităţii speciilor existente. În consecinţă, cercetări similare celor efectuate de Mendel au vizat şi alte specii, inclusiv omul. Un exemplu în acest sens îl constituie încrucişările dintre cobai de culoare diferită: tipul sălbatic, de culoare gri şi tipul albinos (apărut prin mutaţie).
În hibridarea dintre o femelă de tip albinos (cu blana de culoare albă), cu un mascul sălbatic (cu blana de culoare gri), indivizii din generaţia F1 (puii din prima generaţie) aveau în totalitate blana de culoare gri. Indivizii din F1, încrucişaţi între ei, au dat generaţia F2, în care s-a produs segregarea celor două fenotipuri în raportul de 3 gri: 1 alb, adică 3: 1.
În F1 s-a manifestat dominanţa fenotipului unui părinte (şi recesivitatea celuilalt), fenotipizarea genei estompând fenotipizarea alelei. În consecinţă hibrizii erau uniformi (sub aspect fenotipic). În F2 s-a produs segregarea, fenotipică, în raportul de 3 dominant la unu recesiv Evident, ca şi la mazăre, fenomenul a fost explicat pe baza legii purităţii gameţilor.
Fără a intra în alte detalii care converg spre aceeaşi interpretare, în concluzie, se impune precizarea: legile descoperite de Mendel au caracter de universalitate, fiind aplicabile şi, deci, valabile pentru plante şi animale, inclusiv pentru om.
2.4. Abateri de la principiul dominanţei şi recesivităţii din F1 şi de la raportul de segregare de tip mendelian din F2
Aşa cum am precizat şi demonstrat în paginile anterioare, pe baza experimentelor de hibridare între diferite soiuri ale speciei Pisum sativum L., Mendel a legiferat segregarea
caracterelor în descendenţa hibrizilor de F1 (perpetuaţi prin autofecundare sau reproducere de tipul frate x soră, în cazul speciilor alogame) şi, în plus, segregarea independentă a caracterelor în
cazul hibrizilor pentru mai mult decât un caracter. Ulterior, prin experimente şi observaţii efectuate în hibridări la alte specii de plante sau
animale, s-a ajuns la concluzia că există abateri de la principiul dominanţei şi recesivităţii fenotipurilor parentale în F1 şi de la raportul de segregare din F2 sau, altfel spus, că există alte tipuri de segregare (faţă de tipul mendelian).
Este evident că abaterile de la tipul mendelian de segregare se datorează manifestării altor tipuri şi raporturi de dominanţă/recesivitate. În mod normal, determinismul profund al unui caracter se datorează activităţii unei gene. Dar, sub impactul factorilor de mediu, o genă se poate transforma într-o alelă (prin mutaţie). De obicei, genele domină asupra alelelor, în sensul că la heterozigoţi se manifestă fenotipul datorat genei, şi numai la homozigoţii recesivi, la haploizi şi în cazul hemizigoţiei se manifestă fenotipul determinat de alela recesivă. Abaterile de la aceste reguli induc modificări ale fenomenului dominanţă/recesivitate şi schimbări în raporturile de segregare.
2.4.1. Dominanţa incompletă (semidominanţa)
Recurgând la o simbolistică generală şi unificatoare, putem nota o genă oarecare cu A şi alela ei, rezultată prin mutaţie, cu a. În cazul hibridării AA x aa va rezulta individul Aa, heterozigot. Aşa cum precizam, de obicei A domină pe a, fenotipul heterozigotului Aa fiind identic celui al homozigotului AA. Sunt însă şi cazuri în care AA ≠ Aa. Un exemplu, devenit clasic de acum, este cel al hibrizilor dintre varietăţi de Mirabilis jalapa, observat în experimentele efectuate de C. Correns (în 1912). Încrucişând varietatea cu floare de culoare roşie, cu varietatea având flori de culoare albă, în F1 s-au obţinut indivizi uniformi dar toţi cu flori roz. Încrucişând aceşti indivizi între ei, în F2 s-au obţinut indivizi cu flori roşii (25%), cu flori roz (50%) şi cu flori albe (25%) (deci un raport de segregare de 1:2:1, heterozigoţii fiind fenotipic diferiţi de ambii homozigoţi). În F3 indivizii
din F2 care aveau flori roşii, interîncrucişaţi, au dat doar descendenţi cu flori roşii, cei cu flori albe, doar descendenţi cu flori albe, iar descendenţa celor cu florile roz a segregat în continuare, în raportul de 1:2:1.
R x A ↓
r x r ↓
R + r + A 25% 50% 25%
R = roşu r = roz
A = alb
În acest caz (al moştenirii culorii florii), absenţa dominanţei este explicată prin mecanismele metabolice ale formării pigmentului roşu. Pigmentul se formează printr-o succesiune de reacţii enzimatice. Gena, notată cu I în cazul de faţă (de la ivory = ivoriu, alb), determină producerea pigmentului roşu, iar alela i determină absenţa pigmentului (nu determină producerea lui). În heterozigotul Ii se asigură o concentraţie redusă a enzimei ce determină sinteza pigmentului. La o concentraţie redusă a enzimei (probabil 1/2 din concentraţia normală întâlnită la homozigotul II), sinteza pigmentului este, de asemenea, redusă în timpul dezvoltării florii, deoarece pigmentul este produs în cantităţi fixe şi limitate per gena I.
Părinţii II x ii Gameţii I x i ↓ F1 Ii ↓
Gameţii I şi i ↓
F2 II + Ii + ii 25%+50%+25%
Un asemenea efect nu apare însă în toate sistemele, deoarece 1/2 din cantitatea de
enzimă este suficientă pentru a asigura sinteza unei cantităţi adecvate de pigment, determinând manifestarea fenotipului dominant, în cazul heterozigoţilor.
Exemple de manifestare a semidominanţei pot fi date pentru multe alte specii de plante sau animale. De pildă, în încrucişări între soiuri de Zea mays cu bob albastru şi cu bob galben, în F1
rezultă indivizi cu bob violaceu. În F2 segregarea este de 1 albastru: 2 violet: 1 galben. În literatura de specialitate, pentru segregarea de acest fel se foloseşte expresia "segregare de tip Zea" (1:2:1), tocmai spre a o deosebi de segregarea tipică (tip Pisum, 3:1).
În lumea animală, un exemplu bine cunoscut de semidominanţă este cel referitor la rezultatul hibridării între o rasă de găini cu penajul negru şi o rasă cu penajul alb, hibridare în urma căreia rezultă tipul Andaluzia, cu penajul de culoare albăstruie. În F2, din încrucişarea între indivizi ai tipului Andaluzia, iau naştere indivizi cu penaj negru (25%), indivizi tip Andaluzia (50%) şi indivizi cu penaj alb (25%). Evident, tipul Andaluzia nefiind constant, nu se poate vorbi de rasa Andaluzia (rasele la animale şi soiurile la plante trebuie să fie stabile ereditar).
2.4.2. Codominanţa
Codominanţa reprezintă un fenomen genetic complex datorat în esenţă, polialeliei. Cel mai elocvent exemplu este furnizat de situaţia grupelor sanguine umane, mai ales cele din sistemele ABO şi MN. În codominanţă se constată un comportament diferenţiat al alelelor, în sensul că unele
dintre ele se află în raport de dominanţă şi recesivitate, în timp ce altele sunt codominante, în sensul că au "tărie" egală şi contribuie în mod egal la fenotipizarea unui nou caracter.
În 1900, K. Landsteiner a remarcat că prin amestecul globulelor roşii prelevate de la o persoană cu serul sanguin prelevat de la o altă persoană, se produc două fenomene diferite - în unele cazuri, amestecul nu are repercusiuni, iar în altele se soldează cu aglutinarea globulelor roşii. Prin aprofundarea cercetărilor s-a constatat că în serul sanguin uman există două tipuri de anticorpi (notaţi cu anti A şi anti B, sau α şi β) şi două tipuri de antigeni (notaţi cu A şi B) pe
globulele roşii. Pe criteriul prezenţei sau lipsei celor două categorii de molecule s-a ajuns la concluzia că oamenii aparţin la patru grupe sanguine, notate cu A, B, AB şi 0 (zero). De fapt alelele determinante ale grupelor sanguine, notate în unele nomenclaturi cu IA, IB şi I0, în altele cu LA, LB şi l (în onoarea lui Landsteiner), determină sau nu sinteza unor mucopolizaharide (proteină identică + grup terminal de zahăr, care diferă) localizate pe suprafaţa globulelor roşii. Alela IA determină sinteza polizaharidului A, alela IB determină sinteza polizaharidului B, iar alela I0 recesivă nu determină sinteza nici unui polizaharid. În consecinţă, indivizii IAIA şi IAI0 produc antigenul A, indivizii IBIB şi IBI0 produc antigenul B, în timp ce indivizii I0I0 nu produc antigeni (globulele lor roşii sunt fără polizaharide). Interesant este faptul că descendenţa cuplului IAIA x IBIB (care va fi IAIB) produce ambele tipuri de polizaharide, adică ambii antigeni (A şi B). Acesta este tocmai fenomenul de codominanţă, adică alelele IA şi IB sunt dominante (fiecare) asupra alelei I0, dar sunt codominante una faţă de cealaltă, adică ambele contribuie în mod egal şi plenar la determinarea unui nou fenotip (AB, în cazul de faţă). În consecinţă, grupele de sânge din sistemul AB0 pot fi redate schematic, ca în tabelul de mai jos.
Cunoaşterea grupei sanguine a unui individ este extrem de importantă, din două considerente - medical şi juridic. Sub aspect medical, cunoaşterea grupelor sanguine este obligatorie în cazul transfuziilor. În situaţia în care antigenul A vine în contact cu anticorpul α (adică, globulele roşii de la un individ din grupa A cu serul sanguin al unui individ din grupa B), anticorpul (proteina) se leagă la antigen (polizaharid) şi îl inactivează. Concomitent, se produce agregarea (aglutinarea) globulelor roşii. În consecinţă, transfuziile de sânge pot fi efectuate doar în sensul redat în schema care urmează.
Fenotip Genotip Antigen
(polizaharid de pe globulele roşii)
Anticorp (proteina din serul
sanguin)
A IAIA şi I
AI0 sau
LAL
A şi L
Al
A Anti B (β)
B IBIB şi I
BI0 sau
LBL
B şi L
Bl
B Anti A (α)
AB IAIB sau L
AL
B A + B -
0 I0I0 sau ll - anti A(α)+anti B(β)
0 ↔ 0 / ↓ \
A ↔ A ↓ B ↔ B \ ↓ /
AB ↔ AB
Grupa 0 este donator universal, iar grupa AB este primitor universal. Grupa A poate dona
grupei A şi AB şi poate primi de la A şi 0. Grupa B poate primi de la B şi 0 şi poate dona grupei B şi AB. Fiind genic determinate, evident, grupele sanguine sunt ereditare. Cele trei alele LA, LB şi l ocupă acelaşi locus pe cromosomii omologi. Alela LA prezintă mai multe mutaţii, două dintre ele fiind mai frecvente - LA1 şi LA2, între ele fiind un raport de dominanţă (LA1 domină pe LA2). În consecinţă, fenotipic, grupele sanguine din sistemul AB0 sunt A1, A2, B, A1B, A2B şi 0, conform
tabelului:
Fenotip Genotip
A1 LA1
LA1
, LA1
LA2
, LA1
l
A2 LA2
LA2
, LA2
l
B LBL
B, L
Bl
A1B A2B
LA1
LB
LA2
LB
0 Ll
Fără îndoială, ţinând cont de raportul de dominanţă, codominanţă şi recesivitate dintre
alelele precizate, este simplu să se prevadă grupele de sânge posibile ale descendenţilor. În cazul în care se cunosc grupele sanguine ale părinţilor (şi invers), se pot stabili relaţiile de paternitate şi filiaţie, cu repercusiuni pentru demersuri medico-legale şi juridice. De pildă, doi părinţi cu fenotipul
0 nu vor putea da naştere decât la copii cu acelaşi fenotip (ll x ll = ll). Doi părinţi din grupa B, dacă vor fi heterozigoţi, vor putea da naştere la copii din grupele B sau 0 (LBl x LBl = LBLB, LBl şi ll) etc.
2.4.3. Supradominanţa
Reprezintă fenomenul datorită căruia heterozigoţii depăşesc ambii părinţi în fenotipizarea unor caractere (mai ales cantitative, cum ar fi habitusul, productivitatea etc., inclusiv viabilitatea), ca o componentă a capacităţii de a da răspunsuri adecvate la presiunea selecţiei. Schematic, supradominanţa poate fi redată astfel:
AA < Aa > aa
Se apreciază că în supradominanţă sunt implicate mai multe gene cu calităţi aditive. În mod
frecvent, supradominanţa reprezintă rezultanta interacţiunii a două alele, rezultantă ce face ca heterozigoţii să se situeze în afara amplitudinii de variabilitate a fiecăruia dintre cei doi homozigoţi. Selecţia favorizând heterozigoţii, asigură frecvenţa constantă a unei gene. Într-o populaţie care nu este consangvinizată, cel mult 50% dintre indivizi pot fi heterozigoţi pentru un anumit locus. Numai atunci când există supradominanţă pentru un anumit caracter (pentru o anumită pereche genă-alelă), consangvinizarea şi încrucişarea pot realiza ceea ce selecţia fără consangvinzare nu poate. În condiţii “normale” de dominanţă şi recesivitate, cel mai bun genotip este homozigotul dominant, toţi indivizii unei populaţii putând fi aduşi, prin selecţie, în stare homozigotă.
2.4.4. Gene letale
O altă cauză a modificării raportului mendelian de segregare în descendenţa hibrizilor din F1 o constituie prezenţa genelor letale.
Noţiunea de genă letală a fost introdusă de L. Cuénot (1911). El a observat că încrucişând şoareci de culoare galbenă, întotdeauna în descendenţă rezultă un amestec de indivizi de culoare galbenă şi de altă culoare, în raport de 2: 1. Rezultatele au impus două concluzii. În primul rând,
faptul că descendenţa segregă înseamnă că şoarecii galbeni sunt heterozigoţi. În al doilea rând, segregarea fiind continuu în raport de 2: 1 şi descendenţa indivizilor de culoare galbenă segregând mereu, înseamnă că o parte dintre descendenţii indivizilor de culoare galbenă şi anume cei homozigoţi (a căror descendenţă ar trebui să nu segrege), nu iau naştere. Prin urmare, gena dominantă în stare homozigotă este letală. Aceasta a fost presupunerea lui Cuénot. Sacrificarea femelelor gestante a confirmat această presupunere, deoarece s-au identificat embrioni morţi care corespundeau ca număr procentului indivizilor necesari pentru asigurarea raportului de segregare de 3 : 1.
Dacă notăm gena pentru culoarea normală a blănii cu a şi mutanta care determină apariţia culorii galbene cu Ay (Y de la cuvântul englezesc yellow = galben), situaţia se va prezenta astfel:
- indivizii galbeni heterozigoţi au genotipul Aya. Prin încrucişarea lor rezultă: Aya x Aya = AyAy (25%) + 2Aya (50%) + aa (25%)
Înseamnă că, în mod continuu, indivizii AyAy mor în stare embrionară.
2.4.5. Interacţiunea genelor (epistazia)
Este un alt fenomen care determină modificarea raportului de segregare şi chiar apariţia de noi fenotipuri în descendenţa hibrizilor din F1.
Interacţiunea genelor a fost constatată la încrucişarea dintre două rase de găini, ambele albe: Leghorn şi Wyandotte. Prin experimente şi observaţii multiple s-a demonstrat că la rasa Leghorn culoarea albă este dominantă, în timp ce la rasa Wyandotte este recesivă. În consecinţă,
dacă rasa Leghorn este încrucişată cu o rasă de culoare închisă, F1 va fi de culoare albă. Dacă se încrucişează indivizi ai rasei Wyandotte cu indivizii unei rase colorate, indivizii din F1 vor fi, în totalitate, coloraţi. Prin urmare, la cele două rase, Leghorn şi Wyandotte, culoarea albă a penajului este determinată de gene diferite, nealele (Fig.1).
Se consideră că Leghorn la origine avea penajul colorat (CC; C = simbolul pentru gena determinantă a culorii, de la cuvântul englezesc colour = culoare). Exprimarea genelor determinante ale culorii este blocată însă de altă genă inhibitoare (II; I = simbolul pentru respectiva genă, de la cuvântul englezesc inhibitory = inhibitor). Deci, genotipic, rasa Leghorn este CCII.
Rasa Wyandotte de culoare albă (ca şi rasele White, Plymonth etc.) dar recesivă, are genotipul ccii.
Prin încrucişarea dintre cele două rase CCII x ccii, rezultă hibrizii din F1, de culoare albă, cu genotipul CcIi. Aceştia formează gameţii CI, cI, Ci, ci şi prin încrucişare dau hibrizii din F2,
conform tabelului.
Gameţii CI Ci cI ci
CI CCII Alb
CCIi Alb
CcII Alb
CcIi Alb
Ci CCIi Alb
CCii Colorat
CcIi Alb
Ccii Colorat
cI CcII Alb
CcIi Alb
ccII Alb
ccIi Alb
ci CcIi Alb
Ccii Colorat
ccIi Alb
ccii Alb
Prin urmare, raportul de apariţie a indivizilor cu penaj colorat este de 3/16. Genotipul lor conţine gena inhibitoare în stare recesivă homozigotă, iar gena pentru culoare, în stare dominantă, homozigotă sau heterozigotă.
Fenomenul se numeşte epistazie, iar gena inhibitoare, a cărei acţiune împiedică manifestarea acţiunii genei pentru culoare, se numeşte epistatică. Gena inhibată se numeşte hipostatică.
Un alt fenomen interesant de interacţiune genică, în care nu se modifică raportul mendelian de segregare şi care determină apariţia de noi fenotipuri, a fost constatat tot la găini de către W. Bateson şi R.C. Punnett. Cei doi au efectuat încrucişări între găini caracterizate prin forme specifice ale crestei: rasa Leghorn cu creasta de formă simplă, rasa Brahmas cu creasta de tip mazăre şi rasa Wyandotte cu creasta de tip trandafir (Fig.2).
Rasa neagră x Rasa Leghorn = F1
Rasa neagră x Rasa Wyandotte = F1
Rasa Leghorn x Rasa Wyandotte = F1
+
Forma crestei este ereditară, fiind determinată de două gene nealele, notate cu R şi P.
Forma crestei rasei Leghorn este recesivă. Prin urmare, genotipic rasa Leghorn se notează rrpp. Rasa Brahmas domină rasa Leghorn, având genotipul rrPP. La fel şi rasa Wyandotte domină asupra rasei Leghorn, având genotipul RRpp. Prin urmare, dominanţa celor două rase asupra rasei Leghorn este datorată altor gene. La această concluzie s-a ajuns deoarece, dacă se încrucişează cele două rase dominante între ele, rezultă hibrizii de F1 cu un nou tip de creastă, tipul nucă, cu genotipul RrPp. În F2 segregarea se produce în raportul:
9/16 indivizi cu creasta în formă de nucă;
3/16 indivizi cu creasta în formă de trandafir; 3/16 indivizi cu creasta în formă de mazăre;
1/16 indivizi cu creasta de formă simplă.
Astfel, ca o concluzie finală, din interacţiunea între cele două gene în stare dominantă
apare un nou tip de creastă (tipul nucă), inexistent la genitori. Din interacţiunea între alelele celor două gene, recesive, apare creasta de tip simplu, tot inexistentă la genitori.
2.4.6. Polialelia sau alelia multiplă
Fără a intra în profunzimea problemei, amănuntele urmând a fi analizate în capitolul de genetică moleculară, precizăm că gena, care este responsabilă de determinarea unui caracter, ocupă o poziţie bine definită pe un cromosom (ocupă un locus). Orice genă, pe parcursul
existenţei ei, a putut suferi una sau mai multe mutaţii. În consecinţă, pentru o genă au putut apărea una sau mai multe alele (în limba greacă, alelon = “altul”). Rezultă deci, cu claritate, că o alelă sau
o serie de alele ale unei gene sunt implicate în determinismul aceluiaşi caracter (evident, asigurându-i fenotipizări diferite) şi ocupă acelaşi locus în cromosomul omolog (adică poziţia din cromosomul în care se găseşte gena). Evident, din acest punct de vedere, o genă poate avea o alelă sau mai multe (A poate avea alelele a1,a2,..,an), dar un individ nu poate avea decât una dintre stările posibile (AA, Aa sau aa, sau alte combinaţii care să desemneze statutul de homozigot
recesiv sau heterozigot). Într-o populaţie însă, genotipic şi, bineînţeles, fenotipic, poate exista o multitudine de indivizi, cu o multitudine de combinaţii, atât între gena sălbatică şi alelele ei, cât şi între alele.
F2
Fig. 1. Rezultatele încrucişărilor dintre o rasă cu penaj de culoare neagră şi rasele Leghorn şi Wyandotte, precum şi ale încrucişării dintre Leghorn şi Wyandotte
Concomitent deci, există o multitudine de izoalele sau alele izomorfe, a căror activitate
determină diferenţe fenotipice minore, aproape insesizabile prin metode normale. Un exemplu în acest sens îl furnizează variabilitatea culorii ochilor la Drosophila melanogaster. În unele cazuri, nuanţele culorii ochilor nu pot fi determinate decât prin analiza spectrofotometrică a cantităţii de pigment.
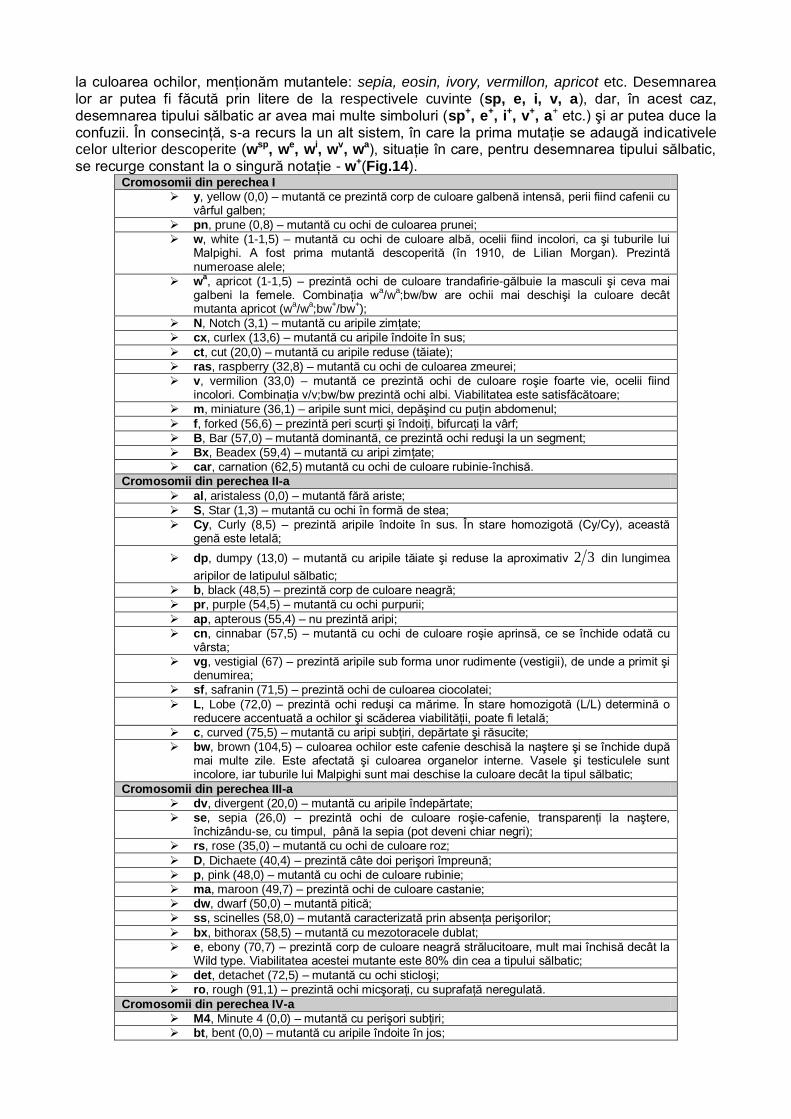
Prin urmare, reluând şi analizând în detaliu problema, rezultă că într-o populaţie, pentru o anumită genă, pot exista mai multe alele. Aşa cum am precizat, acestea provin prin mutaţii succesive, fie ale genei normale, fie ale uneia sau unora dintre alele. Întrucât notaţia alelelor poate duce la confuzii, este bine să redăm câteva dintre modalităţile de desemnare (notare) a lor. La modul general, gena sălbatică se notează cu A, iar alelele cu a1, a2, a3...an. Dar gena normală (sălbatică) se mai poate nota şi cu prima, primele sau un grup de litere din denumirea în engleză a primei mutante (care a fost descoperită şi descrisă) pentru caracterul respectiv, însoţită de indicele +, alelele fiind notate cu aceleaşi indicative, dar fără indicele +. O altă notaţie prevede folosirea unei singure abrevieri, rezultată de la denumirea primei mutante, cu indicele + pentru gena sălbatică şi a aceleiaşi abrevieri, având mereu alt indice format din literele ce desemnează mutantele decelate ulterior, pentru restul alelelor. Să exemplificăm. Culoarea normală a ochiului la Drosophila melanogaster este roşie. Mutanta cu ochi albi se notează cu w (de la cuvântul englezesc white = alb). Gena sălbatică, în acest caz, se notează cu w+. Alelele mutante, altele decât white, se notează cu we (eosin), wa (apricot), wch (cherry), wco (corai).
Prin urmare, culoarea ochilor la drosofila cuprinde o gamă largă de mutante între alb şi roşu. Fiecare nuanţă se datorează unei alele, fapt pentru care se consideră că, la această musculiţă, există un sistem polialelic, o serie alelomorfă. În această serie alelomorfă fenotipul
sălbatic este dominant, toate fenotipurile mutante fiind recesive. Şi între alelele mutante există raporturi de dominanţă şi recesivitate.
Ordinea dominanţei este determinată de intensitatea culorii ochilor, pe care o conferă o alelă. Prin urmare, alela w, care determină culoarea albă a ochilor (deci, lipsa completă a
pigmentului) este recesivă, în raport cu toate celelalte alele. Demn de remarcat este faptul că numărul mutantelor depistate a crescut, concomitent cu punerea la punct a unor metode de laborator pentru determinarea exactă a conţinutului de pigment şi, mai ales, faptul că există mai multe fenotipuri normale (sălbatic tip Steillenbusch w+sw+s, sălbatic tip Couton, w+cw+c, sălbatic tip Graaf-Reinet, w+Gw+G etc.).
Valoarea conţinutului în pigmenţi, spectrofotometric determinată, la ochii mutantelor, variază între 0,0044 şi 0,1636, iar la ochii indivizilor normali între 0,6854 şi 1,2548. Iată de ce heterozigoţii prezintă cantităţi intermediare de pigmenţi şi se poate aprecia că, de fapt, există un fenomen de dominanţă incompletă şi nu de dominanţă/recesivitate de tip mendelian.
Întrucât cantitatea de pigment este variabilă, inducând o variabilitate chiar în cadrul tipului normal, în funcţie de gena concretă pe care o deţine individul, alelele pentru fenotipul normal s-au numit izoalele normale, iar alelele pentru fenotipurile mutante s-au denumit izoalele mutante.
În final, referitor la polialelie putem sublinia câteva particularităţi, şi anume: - polialelele ocupă acelaşi locus în cadrul unui cromosom (grup de linkage); - polialelele fiind, de fapt, mutante ale unei singure gene, afectează întotdeauna acelaşi
caracter. În încrucişare întotdeauna domină gena sălbatică. Concomitent, unele alele sunt dominante
faţă de altele. De aceea, când se încrucişează doi indivizi alelici, în F1 nu apare tipul sălbatic, ci domină unul dintre genitori.
Sunt şi situaţii în care una dintre alele domină asupra tipului sălbatic. Este cazul animalelor agouti şi al unor culori sau forme de ochi la Drosophila melanogaster.
Evident, fiind plasate în acelaşi locus, între polialele nu poate avea loc fenomenul de crossing-over.
2.4.7. Poligenia sau polimeria
Vizează mai ales expresia fenotipică a caracterelor cantitative (înălţime, greutate, producţie de fructe sau seminţe, producţie de alcaloizi, vitamine, uleiuri etc.). Încă din secolul XVIII, prin experimentele sale de hibridare, J. Kölreuter a constatat că hibrizii din F1 rezultaţi din încrucişarea între Nicotiana tabacum pitică şi Nicotiana tabacum înaltă, aveau înălţimea intermediară. În F2
apărea o segregare graduală, în sensul că se identifică o adevărată scară a indivizilor, de la cel pitic, până la cel înalt.
Cercetările efectuate de suedezul H. Nilson Ehle pe grâu şi americanul E. M. East pe porumb, între anii 1910-1913, au permis descoperirea fenomenului poligeniei. În urma acestor cercetări s-a conchis că determinismul caracterelor cantitative este poligenic, în sensul că ele sunt determinate de mai multe gene nealele. Segregarea genelor nealele este independentă (ca şi caracterele independente), doar efectul lor fiind cumulativ.
Un exemplu tipic de efect cumulativ ni-l furnizează manifestarea culorii pielii la om. Culoarea pielii în cazul omului este determinată de conţinutul de melanină, care variază între 0 - 11% pentru rasa albă, 12 - 25% la mulatrii deschişi, 25 - 40% la mulatrii adevăraţi, 41 - 55% la mulatrii de culoare închisă, 56 - 78% la rasa negroidă.
Prin căsătoria între un individ din rasa albă (europoidă) şi unul din rasa neagră (africană), în F1 rezultă indivizi de tip mulatru (relativă uniformitate). Prin căsătoria între mulatri, în F2 se obţine o segregare de 1 alb: 4 mulatri deschis: 6 mulatri propriu-zis: 4 mulatri închis: 1 negru.
Pentru a explica acest raport, C.B. Davenport, care a studiat problema (mai ales, în Jamaica şi insulele Bermude), a considerat că în apariţia culorii pielii sunt implicate două perechi de gene cu alelele lor - P1p1 şi P2p2. Negrii au genotipul P1P1P2P2, iar albii p1p1p2p2. Din căsătoria alb x negru, rezultă:
P1P1P2P2 x p1p1p2p2
↓ F1 P1p1P2p2
Căsătoria între mulatri induce apariţia a mai multe tipuri de indivizi în F2, şi anume:
Gameţii P1P2 P1p2 p1P2 p1p2
P1P2
P1P1P2P2 Negru
P1P1P2p2 Mulatru închis
P1p1P2P2 Mulatru închis
P1p1P2p2 Mulatru
P1p2
P1P1P2p2 Mulatru închis
P1P1p2p2 Mulatru
P1p1P2p2 Mulatru
P1p1p2p2 Mulatru deschis
p1P2
P1p1P2P2 Mulatru închis
P1p1P2p2 Mulatru
p1p1P2P2 Mulatru
p1p1P2p2 Mulatru deschis
p1p2
P1p1P2p2 Mulatru
P1p1p2p2 Mulatru deschis
p1p1P2p2 Mulatru deschis
p1p1p2p2 Alb
Dacă se face un back-cross între un alb p1p1p2p2 şi un mulatru P1p1P2p2, descendenţii vor
fi: 25% P1p1P2p2; 25% p1p1p2p2 şi 50% mulatri, intermediari între părinţi (adică P1p1p2p2 şi p1p1P2p2). Retroîncrucişarea unui negru P1P1P2P2 cu un mulatru P1p1P2p2 induce apariţia a 25%
negri, 50% mulatri închişi, intermediari între cei doi părinţi şi 25% mulatri. Davenport a conchis că acumularea cantitativă a pigmentului în piele este determinată de
efectul cumulat al genelor P1 şi P2 şi al alelelor. Astfel, negrii au genele în stare homozigotă, mulatrii închişi au o genă homozigotă şi celelalte în stare heterozigotă (P1p1P2P2, P1P1p2P2),
mulatrii propriu-zişi sunt heterozigoţi pentru ambele gene sau homozigoţi pentru ambele, dar una în stare dominantă şi cealaltă în stare recesivă (P1p1P2p2; P1P1p2p2; p1p1P2P2), mulatrii deschişi au o singură genă dominantă în stare heterozigotă (P1p1p2p2 sau p1p1p2P2).
Ulterior, au fost elaborate şi alte ipoteze în care s-a presupus existenţa a mai mult de 2 perechi de gene - între 3 (R.R.Gotes, 1953) şi 20 (C. Stern, 1960).
INTREBARI DE VERIFICARE
1. Definiti termenii: hibridare, caracter, factor ereditar 2. Enuntati legile mendeliene ale ereditatii 3. Intocmiti schema monohibridarii 4. Intocmiti schema dihibridarii 5. Care sunt cauzele aparitiei si a altor raporturi de segregare decat cele precizate de catre Mendel 6. Ce reprezinta semidominanta si supradominanta
7. Care este importanta grupelor de sange la om si care este determinismul lor genetic 8. Cum se defineste fenomenul de poligenie 9. Ce reprezinta epistazia 9. Intocmiti schema unei hibridari intre doua persoane apartinand tipului „mulatru propriu-zis”
TEME PENTRU EXAMINAREA FINALA
1. Dobandirea si operarea corecta a notiunilor fundamentale de genetica 2. Intelegerea legitatilor mendeliene si prezentarea schemelor de hibridare in mono- si dihibridare 3. Argumentarea caracterului universal al legilor mendeliene 4. Caracterizarea fenomenelor care produc abateri de la legitatile mendeliene de transmitere a caracterelor
Unitatea de invatare 2
3. Structura si functiile suportului material al ereditatii, variabilitatii si
determinismului caracterelor
3.1. Cromosomii – seiful informaţiei ereditare
3.5.1. Materialul genetic al virusurilor, viroizilor si plasmidelor
Primele forme de viaţă apărute pe Terra erau, probabil, de tip acelular fiind reprezentate de macromolecule de ARN si apoi de ADN şi aveau capacitatea de autoreplicare, autoreglare şi autodezvoltare. Apariţia primelor gene, alcătuitela început din ARN şi apoi din ADN, a insemnat trecerea de la evoluţia chimică la cea biologică. Desigur că primele forme de viaţă cuprindeau in programul lor genetic un număr foarte redus de gene. în care era codificată informaţia necesară realizării structurilor şi funcţiilor caracteristice acestor organisme primitive. Apariţia codului genetic, prin care informaţia ereditară stocată în macromoleculele de acizi nucleici putea fi decodificată şi transformată intr-o secvenţă de aminoacizi, a făcut posibilă asamblarea nucleoproteinelor, mai stabile şi cu o capacitate de evoluţie mai rapidă. Studiul organismelor procariote acelulare actuale, cum sunt virusurile, viroizii şi plasmidele a facut posibilă cunoaşterea mai aprofundată a proceselor care au dus la apariţia vieţii, deoarece ele sunt extrem de simple ca organizare structurală si ca funcţii, fiind oarecum similare cu protobionţii apăruţi in negura timpurilor.
Materialul genetic al virusurilor este reprezentat de un singură moleculă de acid nucleic în care sunt dispuse, in ordine liniară, un număr variabil de gene. Virusurile cu genom ADN, numite si dezoxiribovirusuri, prezintă şapte tipuri principale de structură şi anume: - cu genom ADN mono-catenar. liniar, întâlnit la Parvovirus, fagul M13;
- cu genom ADN mono-catenar circular, ca la fagul X174, la fagul filamentos fd şi la unele virusuri ale plantelor;
- cu genom ADN dublu-catenar liniar repetitiv (redundant), datorită secvenţelor terminale repetate, identic situate la fiecare extremitate a moleculei; ca la fagul T7;
- cu genom ADN dublu-catenar liniar repetitiv (redundant) terminal (ca la Adenovirusuri), fapt care-i conferă proprietatea ca după denaturare, moleculele monocatenare să se poată circulariza prin unirea extremităţilor repetate invers;
- cu genom ADN dublu-catenar liniar redundant terminal, permutat circular, caracteristic fagilor din seria T - pari, care au la fiecare extremitate secvenţe terminale provenite insă din regiuni diferite ale genomului. Genomul liniar al fagilor T - pari se poate circulariza in vivo datorită structurii extremităţilor, care au o secvenţă terminală repetată;
- cu genom ADN dublu-catenar liniar, cu extremităţi monocatenare complementare
(adezive), caracteristic fagului . În particula fagică ADN-ul rămâne totdeauna liniar, dar in interiorul bacteriei gazdă (Escherichia coli) cele două extremităţi adezive complementare
se leagă şi formează o moleculă circulară de ADN dublu-catenar, care prezintă câte o singură breşă pe fiecare catenă;
- cu genom ADN dublu-catenar circular inchis covalent, prezentând aproximativ 20 de răsuciri superhelicale suplimentare sau răsuciri terţiare, care dau naştere unei molecule suprarăsucite. Acest cromosom care nu are nici o extremitate liberă, deoarece toate legăturile dintre nucleotide sunt covalente, este caracteristic pentru virusurile polioma si SV 40.
Virusurile ARN, care au genomul reprezentat de o macromoleculă de ARN, sunt numite ribovirusuri şi prezintă patru tipuri principale de structuri: - cu genom ARN mono-catenar liniar, dispus helical, corespunzător structurii proteice a
capsidei, ca în cazul virusului mozaicului tutunului (VMT);
- cu genom ARN mono-catenar intens pliat, care se impachetează uşor şi se intâlneşte la fagii ARN mici - R17 , MS2, f2;
- cu genom ARN mono-catenar circular, intâlnit la virusul encefalomiocarditei de la
şoareci; - cu genom ARN dublu-catenar divizat, format dintr-un anumit număr de segmente, ca la
Reovirus (10 segmente) sau Myxovirus (8 segmente). Cromosomul viral, constituit din ADN sau ARN, conţine toată informaţia ereditară necesară
pentru formarea particulei virale. Dar, informaţia ereditară a virusurilor devine activă numai atunci când ADN-ul sau ARN-ul lor se află in sistemul celulei gazdă. Când se află în particula virală, toate genele cantonate pe cromosom “tac”, sunt nefuncţionale, inactive. Şi in spermatozoizii animalelor, genele sunt inactive. Şi în diferite ţesuturi vegetale sau animale, unele gene “tac” (sunt inactive), în timp ce altele funcţioneaza intens.
Infecţiozitatea acizilor nucleici virali a fost demonstrată, pentru ADN-ul fagic, de către Hershey si Chase in 1952 şi, ulterior, pentru ARN-ul VMT de către Gierer şi Schramm, în 1956. In
cazul virusurilor animale sunt infectioşi toţi acizii nucleici care sunt transmişi în ARNm de ARN - polimerazele celulei gazdă sau care acţionează ei inşişi ca ARNm (ca de ex. Picorna-, Toga-, Papova-, Adeno si Herpesvirus). Acizii nucleici virali care sunt transcrişi in ARNm de polimeraze codificate de virus (de exemplu ARN proveniţi de la Myxo-, Rhabdovirus si ADN dublu-catenar de la Reovirus, nu sunt infecţioşi).
Acizii nucleici virali izolaţi au o infecţiozitate de 103 - 106 ori mai mică decât aceea a virusurilor din care sunt extraşi, datorită degradării lor rapide de către nucleazele prezente în lichidele extracelulare, ca şi în membranele celulare, precum şi datorită faptului că sunt slab fixaţi de celule (în absorbţia şi fixarea virusurilor pe celule un rol important îl are capsida).
Spectrul de gazde ale acizilor nucleici virali izolaţi este însă mult mărit în comparaţie cu cel al particulei virale mature, la care intervine specificitatea interactiunii dintre capsidă şi receptorii specifici celulari.
Teoretic, acizii nucleici virali, dacă nu sunt degradaţi de nucleaze se pot replica in orice celulă în care au pătruns. De exemplu, în timp ce virionii de Poliovirus infectează numai celulele
umane sau de primate, ARN-ul izolat se replică in celulele de pui de găină sau şoarece, care in mod normal nu sunt permisive pentru acest virus.
O formă acelulară de organizare, mai simplă decât virusurile şi de dimensiuni incomparabil mai mici, este reprezentată de viroizi. Denumirea lor le arată inrudirea cu virusurile, ei fiind in
esenţă molecule mici de acid nucleic neprotejat de inveliş proteic (de capsidă) şi nici de invelişul exterior ce conţine lipide şi proteine, asa cum este cazul la unele virusuri. In anul 1971 au fost evidenţiate, pentru prima dată, nişte boli ale plantelor provocate de un tip special de agenţi patogeni subvirali, denumiţi viroizi. Printre aceste maladii ale plantelor pot fi citate: boala stelării tuberculilor de cartof, cloroza castraveţilor, nanismul şi pătarea clorotică a crizantemelor, boala cadang-cadang a palmierilor de cocos etc. Primul viroid a fost descoperit de Dienner, din USA, pe baza cercetărilor efectuate la cartof. Viroizii sunt un tip special de agenţi patogeni, similari virusurilor, dar cu o organizare mai simplă şi de dimensiuni mult mai mici. Viroizii sunt alcătuiţi din molecule mici de ARN. neprotejate de proteine. Viroidul cartofului, care provoacă deformarea tuberculilor, are un genom reprezentat printr-o moleculă circulară de ARN, cu regiuni mono- şi dublu- catenare, alcatuită dintr-o secvenţă de 359 de nucleotide. A fost pentru prima dată când s-a identificat o astfel de structură circulară, formată din ARN monocatenar.
S-a constatat că viroizii sunt localizaţi în nucleul celulelor, unde se replică cu ajutorul unei enzime de tipul replicazelor din celula-gazdă. La plante s-au identificat secvenţe de nucleotide din ADN-ul nuclear complementare cu ARN-ul viroizilor, dar se pare că replicaţia viroizilor nu se realizează prin intermediul ADN. Date experimentale de ultimă oră arată că în celulele vegetale există o multitudine de enzime care pot asigura replicarea moleculelor de ARN, folosind ca matriţă chiar moleculele de ARN ale viroidului. Originea viroizilor nu este prea bine cunoscută. Pornindu-se de la observaţia că genele plantelor si animalelor sunt alcătuite din regiuni informaţionale, denumite exoni si regiuni non-informaţionale, denumite introni, care sunt eliminaţi in procesul de activare a genelor, s-a emis ipoteza că viroizii ar fi introni nedegradaţi. Caracterul patogen al viroizilor se datorează interreacţiei lor cu genomul celulei-gazdă, probabil prin perturbarea mecanismelor de reglaj genetic al acesteia. Ca urmare, se consideră că viroizii sunt la limita inferioară a vieţii, fiind sisteme autoreplicative, care posedă un program genetic puternic redus. In anul 1952, J. Lederberg, laureat al premiului Nobel, a descoperit plasmidele din celulele bacteriene. Acestea sunt nişte minicromosomi de formă circulară sau lineară, alcatuiţi din ADN bicatenar care se replică independent de cromosomul bacteriei gazdă şi posedă un mic număr de gene. In celula bacteriană se găsesc între 10 şi 200 de copii ale unei anumite plasmide, regula fiind următoarea: plasmidele de dimensiuni mici se pot găsi intr-un număr mai mare de copii, iar cele mari intr-un număr mic de copii. Ulterior s-au descoperit plasmide şi in celulele eucariote, asociate cu nucleul sau cu organitele celulare (mitocondrii şi cloroplaste).
Plasmidele sunt mai simple ca organizare decât virusurile, fiind alcătuite dintr-o moleculă de ADN cu capacitate autoreplicativă. Ele se găsesc la toate speciile de bacterii dar prezenţa lor nu este obligatorie. Până în prezent au fost identificate peste 1000 de tipuri , dintre care 269 numai la bacteria Escherichia coli. De regulă, ADN-ul plasmidial reprezintă 1-3% din cantitatea totală de
material genetic vbacterian. Plasmidele au un rol foarte important în celula bacteriană. Unele dintre ele sunt capabile să
determine conjugarea bacteriilor. Astfel plasmidul denumit factor de fertilitate (F), se poate găsi în celula bacteriană sub formă liberă şi atunci celula este notată F+, sau poate fi integrat în cromosomul bacterian, celula respectivă fiind notată Hfr (high frequency of recombination), sau poate fi absent, situaţie în care celula se notează F- . Bacteriile de tip Hfr sunt capabile să realizeze
recombinarea genetică, adică transferul unui segment cromosomial de la celula masculă (F+) la cea femelă (F-), cu o mare frecvenţă. Există şi plasmide F’ care au inclus un segment din cromosomul bacterian, fapt pentru care acest tip de factor de fertilitate este considerat recombinat genetic. Plasmida F‟ este reprezentată de o macromoleculă de ADN alcatuită din cca. 100.000 perechi de nucleotide. Un alt tip de plasmide bacteriene este constituit de plasmidele pentru rezistenţă (R). Ele sunt alcătuite dintr-o macromoleculă circulară de ADN, pe care se găsesc gene ce determină rezistenţa la compuşii toxici ai unor metale grele (mercurul, cadmiul, bismutul, antimoniul etc.), rezistenţa la unele insecticide şi la unele produse petroliere etc. Plasmidele R pot trece de la o bacterie la alta, cu ajutorul virusurilor bacteriene, printr-un fenomen denumit transducţie.
Plasmidele F si R fac parte din grupa plasmidelor conjugative: favorizează transferul de material genetic, posedând capacitatea de a induce în celulele bacteriene formarea de structuri necesare conjugării (pili de sex). Plasmidele Col (factorii colicinogenici) sunt un alt tip de plasmide, care posedă gene ce determină sinteza unei clase de proteine ce se numesc colinice sau bacteriocine. Acestea,
secretate în mediu, au acţiune bactericidă asupra altor bacterii, care sunt lipsite de astfel de plasmide. Astfel, bacteria Escherichia coli produce colicine, Bacillus subtilis produce subtilizine, Pseudomonas aeruginosa produce pyocine. Fiecare tip de bacteriocină este codificat intr-un
anumit segment de ADN din plasmidul respectiv. Plasmidele sunt cromosomi in miniatură, alcătuiţi din ADN dublu-catenar, circular închis. Circularitatea este o condiţie esenţială pentru supravieţuirea plasmidei in celula gazdă deoarece altfel ADN-ul plasmidial ar fi cu usurinţă atacat de nucleaze. Plasmidele au câteva proprietăţi distincte:
- dimensiunea mică a moleculei: reprezinta 1-2% din mărimea cromosomului bacterian; - secvenţa de baze azotate diferă de cea a ADN-ului cromosomului bacterian; - prezintă proprietăţi structurale aparte;
- reprezintă informaţie ereditară accesorie, fără importanţă vitală pentru bacterie; bacteriile pot să piardă plasmidele;
- reprezintă repliconi - structuri capabile de replicare independentă dar, in general, corelată şi controlată de cromosomul bacterian.
ADN-ul plasmidial poate exista sub mai multe forme: a. covalent închis, circular şi superrăsucit. Această stare facilitează izolarea plasmidelor prin
centrifugare in gradient de densitate; b. circular deschis; c. ADN multimer.
Diferite date experimentale au permis stabilirea hărţilor genetice ale plasmidelor, demonstrându-se existenţa, în ADN-ul plasmidial, a mai multor categorii de determinanţi (gene): - gene necesare pentru replicare; - secvenţe de inserţie (porţiuni de ADN ce au funcţia de a se cupla cu ADN-ul cromosomului bacterian, în cazul în care plasmida se inseră în acesta); - gene implicate in corelarea replicării plasmidei concomitent cu diviziunea celulei şi repartizarea plasmidelor în celulele rezultate după diviziune; - în unele cazuri există un operon de transfer, reprezentat de un grup de gene pentru sinteza pililor de sex si a antigenelor celulare de recunoaştere, obligatoriu prezente în conjugare; - gene structurale ce determină funcţii noi, manifestate în fenotipul gazdei: rezistenţă la antibiotice, rezistenţă la metale grele (mercur, plumb, cadmiu etc), producerea de bacteriocine, inducerea de tumori la plante, capacitatea de a face fermentaţie lactică, capacitatea de a produce anumite antibiotice (cloramfenicol şi metilenomicina), etc.
In ultima vreme au fost identificate plasmide şi în celulele eucariotelor, unele asociate cu nucleul şi altele cu mitocondriile şi cloroplastele. Prima plasmidă asociată cu genomul celular a fost identificată la Saccha-romyces cerevisiae. Aceasta este alcatuită dintr-o secvenţă de 6300 perechi de nucleotide, este de tip circular şi se găseşte în aproximativ 50-100 copii per celulă. Plasmida, cunoscută sub numele 2µm ADN, posedă 3 gene, din care două sunt necesare pentru replicarea sa şi una este implicată în
procesul de recombinare genetică intramoleculară, precum şi două secvenţe inversate de nucleotide, cu ajutorul cărora se poate integra în cromosomii celulei gazdă. O altă plasmidă circulară, asociată nucleului celular, a fost identificată la ciuperca Cephalosporium acramonium, şi are mărimea de 21 Kb.
Un al doilea tip de plasmide, în celula eucariotă, este asociat cu mitocondriile şi cloroplastele. La ciuperca Podospora anserina a fost identificată o plasmidă cu o mărime de 2,4
Kb, care prezintă analogie cu genomul mitocondrial. Această plasmidă se consideră că a provenit prin amplificarea unui segment din genomul mitocondrial. La plantele superioare au fost identificate unele plasmide mitocondriale. La Vicia faba indivizii normali conţin 5 tipuri de plasmide mitocondriale mici (1-2 Kb.). La liniile androsterile, unul din aceste tipuri de plasmide este absent. Plasmidele din celula eucariotă, asociate cu nucleul sau mitocondriile şi cloroplastele, sunt componente normale ale genomului celular şi sunt foarte variate ca structură şi greutate moleculară. Ele contribuie la mărirea variabilitaţii genotipice şi flexibilităţii acestor sisteme genetice. Se speră că va fi posibilă utilizarea lor ca vectori în programele de ameliorare a plantelor, prin transfer de gene.
3.5.2. Structura cromosomului bacterian
Celula bacteriană, spre deosebire de celulele organismelor eucariote, nu are un nucleu adevărat ci un „nucleu‟ cu o organizare primitivă, fiind lipsit de membrană şi inclus direct in citoplasmă, în partea centrală a celulei. Acest „nucleu” nu parcurge procesele de tip mitotic, de aceea el este numit nucleoid sau pur si simplu cromosom bacterian. Genomul bacterian este alcătuit din două categorii de gene: gene esenţiale eucromosomale, localizate în cromosomul bacterian si gene accesorii, prezente in plasmide, elemente genetice transpozabile (SI si TN) si în
unii fagi (Campbell, 1981) Cromosomul bacterian are formă circulară şi este alcătuit dintr-o singură molecula de ADN bicatenar, cu o lungime de 1400 µm şi un diametru de 2,5 nanometri (nm), corespunzând diametrului moleculei de ADN dublu catenar, o lungime de cca. 1360 µm si o masă moleculară de
cca. 2,5 ± 0,3x109 Daltoni. La Escherichia coli şi la majoritatea celorlalte bacterii, cromosomul este
format dintr-o unică moleculă de ADN dublu-catenar, circular închis şi superhelical. Respectiva moleculă constă din aproximativ 4 milioane pb (perechi de baze nucleotidice) care alcătuiesc aproximativ 3000 de gene, dispuse liniar pe cromosomul circular, alcătuind un singur grup de linkage. Lungimea totală a cromosomului bacterian este de cca 1300µ, in condiţiile in care o bacterie are forma cilindrică cu un diametru de 1µ si o lungime de maximum 3µ. Este normal ca, în condiţiile dimensionale menţionate, acest cromosom să fie superrăsucit pentru a putea incăpea în celulă. A.Worcel şi E.Burgi, în 1972 şi Pettijohn in 1974, au reuşit să studieze structura cromosomului la Escherichia coli. S-a demonstrat că el constă din 40-50 bucle care îşi păstrează structura cu ajutorul ARN. Fiecare buclă este bogată în ARN şi poate funcţiona relativ independent de altele. Cele 40-50 de bucle mari au superrăsuciri secundare alcătuite din cca. 400 perechi de nucleotide. Menţinerea acestor bucle de mărimi diferite, se pare că se face nu numai cu ajutorul ARN, ci şi cu ajutorul unor proteine şi enzime de tipul endonucleazelor. Aceste enzime sunt capabile să determine rupturi în una dintre catenele ADN-ului, fapt care impiedică (blochează) derăsucirea buclelor superrăsucite. In acest fel structura cromosomială este stabilă, iar cromosomul bacterian îsi păstrează continuitatea. Enzimele de tipul ribonucleazelor pot hidroliza unele molecule de ARN, fapt care duce la ruperea legăturilor dintre două bucle succesive. Una dintre enzimele care joacă un rol esenţial în replicarea ADN bacterian şi este responsabilă de suprarăsucirea sa, este aşa-numita ADN-giraza. Dacă într-o cultură de E. coli se adaugă, în mediu, coumeromicina, un antibiotic cu rol de inhibitor al funcţiei ADN-girazei, cromosomul celulei respective îşi va pierde constituţia superhelicală. Din motive incă necunoscute, E. coli posedă mecanisme de reglaj al gradului de superrăsucire a cromosomului. Alături de ADN-girază există şi o a II-a enzimă implicată în răsucirea cromosomului - topoizomeraza I. Această enzimă produce relexarea regiunilor
superhelicale. Au fost izolate şi mutante lipsite de topoizomerază sau care produc o formă inactivă a topoizomerazei. Nucleoidul izolat de la astfel de mutante prezintă un grad înalt de superrăsucire, cu cca 32% mai mult decât normalul. Paradoxal însă, aceste mutante nu suferă defecte de creştere şi, în plus, suferă frecvent mutaţii ale genei pentru ADN-giraza, care conduc la producerea unei giraze mai puţin eficiente. În consecinţă cromosomul prezintă un grad normal de superspiralizare. În mod normal, bacteriile aflate în faza de repaus, în culturi staţionare şi vechi, au un singur cromosom, fiind uninucleate. În faza de creştere activă, în culturi tinere menţinute pe medii optime, ele apar ca multinucleate, având 2-4 cromosomi, care sunt însă identici, deoarece provin prin replicare dintr-un singur cromosom parental. De aceea, indiferent de aspectul morfologic al materialului nuclear, din punct de vedere genetic, bacteriile sunt organisme haploide, cu n cromosomi. Atunci când, prin recombinare, o celulă bacteriană primeşte un aport de material nuclear exogen, diploidia nu este decât parţială şi tranzitorie. Această stare se numeşte merozigotă.
Deci, cromosomul bacterian poartă în structura sa toată informaţia ereditară necesară pentru existenţa unei celule, adică poartă toate genele (genomul) necesare pentru desfăşurarea metabolismului şi pentru reglarea activităţii celulare. Cromosomul bacterian determină şi arhitectura celulei care-l conţine, ereditatea şi capacitatea ei de evoluţie. În prezent nu se cunoaste exact mecanismul molecular prin care cromosomul bacterian, mai lung de 100-400 de ori decât lungimea celulei bacteriene, este “împachetat” într-un volum atât de mic, cum este cel al nucleoidului în care se află. Dacă “împachetarea” s-ar face la întâmplare, s-ar produce inevitabil “încurcarea” moleculei de ADN, o parte din informaţia ereditară ar fi prinsă în zonele mai ascunse şi ar deveni inaccesibilă pentru transcripţie. Încă din anul 1966, Maale şi Kjeldgaard, bazându-se pe structura constant limitată la un volum mic din celulă, fără să se răspândească în citoplasmă, a ADN bacterian şi pe faptul că informaţia genetică este totdeauna accesibilă sistemelor de transcripţie şi translaţie, au emis ipoteza menţinerii fibrilelor de ADN învecinate într-o structură organizată, prin intermediul poliaminelor, care ar forma legături tranzitorii cu ADN. Problema modului de împachetare a cromosomului bacterian este deosebit de importantă pentru biologia bacteriană deoarece este strâns corelată cu unele funcţii esenţiale ale materialului genetic (ca replicarea, transcrierea si recombinarea). Fong (1967) a propus un mecanism de împachetare prin suprarăsucire. După ipoteza sa
modificările proprietăţilor dielectrice ale mediului nuclear afectează forţele de atracţie
intermoleculară dintre perechile de baze şi, ca urmare, molecula circulară dublu elicală se răsuceşte intr-o superelice de gradul I (o dublă helice în care fiecare helice este o dublă helice, provenită din molecula iniţială). În felul acesta, molecula circulară, s-ar transforma intr-o structură liniară a cărei lungime este jumătate din circumferinţa iniţială. Procesul ar continua prin formarea de superhelixuri de gradul 2,3,4, s.a.m.d. O superhelice de gradul 10 ar avea o dimensiune liniară de 210 ori mai mică decât lungimea originară a ADN (respectiv de 1024 ori mai mică), ceea ce corespunde aproximativ dimensiunilor nucleoidului de E.coli. După Fong, acest proces ar necesita
introducerea a ~ 1000 ture de suprarăsucire în ADN, schimbare teoretic posibilă, deoarece ar corespunde o tură supraelicală, la fiecare 300-400 ture ale ADN originar. Ulterior, Pettijhon şi Hecht (1974) au elaborat un model de împachetare a ADN cromosomial, bazat pe un proces dublu de pliere şi formare de superelice, în care structura condensată a ADN ar fi menţinută prin acţiunea asociată a ARN şi a proteinelor din nucleoid. Modelul este susţinut de următoarele observaţii:
a. Nucleoidul se derulează daca este tratat cu agenţi care degradează sau denaturează proteinele, ceea ce arată că acestea sunt esenţiale pentru menţinerea stabilităţii cromosomului izolat in stare compactă (Pettijohn 1973, 1979, Drilica si Worcel, 1975);
b. In mod similar, tratarea cu RN-ază sau suprimarea sintezei ARN prin cultivarea în medii cu rifampicină duc la apariţia unor molecule cromosomiale mai mult sau mai puţin depliate şi aproape lipsite de structuri superelicale detectabile (Hecht şi colaboratorii, 1977). Deplierea totală are loc când fie ARN, fie proteinele nucleoidale sunt complet disociate de ADN;
c. Din cercetarile lui Vinograd (1965) este cunoscut faptul că producerea unei incizii monocatenare cu ajutorul ADN-azei, în structura AND, suprimă constrângerile rotaţionale şi, ca urmare, catena sectionată se roteşte liber în jurul celei intacte, determinând suprimarea suprarăsucirilor. Ori, în cazul nucleoidului de E. coli incizia monocatenară elimină brusc
constrângerile şi structura supraelicală numai într-un segment localizat al ADN, ca şi cum anumite restricţii ar împiedica propagarea rotaţiei catenei secţionate în regiunile adiacente. Aceasta demonstrează că ADN cromosomial este impărţit într-o serie de domenii de suprarăsucire, fiecare supus unor constrângeri topologice separate. In conformitate cu aceste fapte de observaţie, modelul propus de Pettijohn şi Hecht (1973) consideră că moleculele de ARN în curs de formare se leagă de două situsuri separate de pe molecula de ADN cromosomial, împărţind-o intr-o serie de domenii de pliere (folding) şi suprarasucire (supercoiling). Un domeniu de suprarăsucire este deci reprezentat de o regiune a
ADN dublu elicoidal delimitată de cele două situsuri de legare ale ARN. Datorită lor, producerea unei incizii monocatenare cu DN-aza desface superelicea numai în bucla respectivă, fără a afecta structura superhelicală a altor domenii. Degradarea a două molecule de ARN, aparţinând unor domenii adiacente, cu RN-aza, uneşte cele două domenii, fără pierderea structurii lor superelicale. Fiecare moleculă de ARN leagă între ele două regiuni indepărtate ale dublei elici cromosomiale în aşa fel incât o serie de interacţiuni de acest gen separă întregul cromosom într-o serie de domenii succesive de-a lungul lui. ARN în curs de sinteză este legat de extremitatea 3‟ a ADN prin intermediul unei molecule de ARN - polimerază, celălalt situs de legare putând fi un hibrid ARN- ADN, o legătură triplu elicală sau mediată de o proteină specifică (Lydersen si Pettejohn, 1977). Numărul de domenii estimat prin tehnica mai exactă de producere de incizii monocatenare cu ajutorul radiatiilor gama este de 100 ± 30 per echivalent genomic. Deoarece nucleoizii conţin în medie 2,2 echivalenţi genomici, Pettijohn si Carlton (1979) apreciau că în medie ar exista ~200 domenii per nucleoid. Ca o consecinţă a legării ARN, cromosomul bacterian circular având Ø de ~ 350 µm, este pliat, formând un număr de ~ 80 bucle care suferă formarea de superelici răsucite spre stânga, după modelul Worcel si Burgi (1972).
3.5.3. Cromosomii la eucariote
Studiul diviziunii celulare indirecte (Francis, Dudith & Inzé, 1997) a evidenţiat atât existenţa cât şi comportamentul cromosomilor. Cromosomii au fost denumiţi astfel, în 1888, de către W. Waldeyer şi, apoi, au fost identificaţi ca purtători (substrat) ai informaţiei ereditare.
Morfologia cromosomilor este variabilă, atât în funcţie de faza din ciclul diviziunii celulare, cât şi în funcţie de specia la care aparţin indivizii (plante sau animale) analizaţi. Încă o precizare,
extrem de importantă, este aceea în conformitate cu care cromosomii eucariotelor sunt cu totul altfel structuraţi în comparaţie cu cei ai procariotelor.
În interfază, cromosomii celulelor somatice normale sunt invizibili la microscopul optic, deoarece au o extensie maximă, care le conferă un aspect filiform. Sunt alcătuiţi din două filamente paralele, alipite, numite cromatide. Examinate la microscopul optic, cromatidele etalează o structură helicoidală. O rotaţie a helixului alcătuieşte un gir. Girii determină o suprapunere, în spaţiu, a spirelor şi, ca o consecinţă, o suprapunere de material cromatic. De aceea, când sunt privite dintr-o anumită direcţie, cromatidele dau impresia că sunt alcătuite dintr-o succesiune de granule, numite cromomere. De aici a pornit o mare confuzie şi anume: un timp s-a considerat că
cromomerele reprezintă o materializare a genelor. În consecinţă, cromatidele erau considerate ca fiind alcătuite din unităţi discrete - cromomerele numite (la acea dată) şi gene - având, deci, o structură discontinuă. Probe de ordin electronografic şi genetic au dovedit că structura cromatidelor este continuă. Cele două cromatide ce alcătuiesc cromosomul se numesc cromatide surori. Ele se unesc într-un singur punct al cromosomului, în zona centromerului - formaţiune cu
diametrul mai mic decât al cromosomului. Din cauza diametrului inferior celui al cromosomului, centromerul realizează constricţia cromosomială primară.
3.5.3.1. Morfologia cromosomilor eucariotelor
Centromerul nu se colorează cu coloranţi bazici. Poziţia acestuia în cromosom este variabilă, împărţind cromosomul în două braţe. În funcţie de lungimea reală a braţelor şi, apoi, a raportului dintre ele, se stabileşte tipul cromosomului.
Dacă centromerul este situat exact în centrul cromosomului, delimitând două braţe perfect egale (raportul dintre braţe - r - fiind 1), cromosomul este metacentric (prescurtat se notează cu M). Când centromerul împarte cromosomul în două braţe inegale, cu r având valori cuprinse între 1 şi 1,7 , cromosomul este median (m), iar când r are valori cuprinse între 1,7 şi 3, cromosomul este submedian (sm). Când raportul braţelor este cuprins între 3 şi 7 cromosomul este subtelocentric (st), iar când raportul este mai mare decât 7 (r > 7) cromosomul este telocentric (T) (Fig.3). În ceea ce priveşte poziţia terminală a centromerului (adică în ceea ce priveşte posibilitatea existenţei cromosomilor telocentrici) părerile sunt contradictorii. Unii autori afirmă că centromerul nu se poate găsi niciodată în poziţie terminală, în timp ce alţi autori admit această posibilitate.
Se cunosc, de asemenea, mai multe ipoteze explicative şi de modelare a structurii moleculare a cromatidei. Orice ipoteză, orice model molecular, trebuie să satisfacă următoarele cerinţe:
- cromatida să fie formată din două subunităţi, care să-i asigure posibilitatea de a se replica semiconservativ,
- cromatida să acţioneze ca o singură unitate, în schimburile reciproce de material genetic. Cromosomii reprezintă un caracter de diagnoză, de delimitare a speciei asemenea oricărui
alt caracter de natură morfo-anatomică, fiziologo-biochimică, fenologică sau reproductivă. În acest sens, trebuie precizat că cromosomii fiecărei specii eucariote, de plante sau de animale, au particularităţi morfologice şi numerice caracteristice speciei.
Pe lângă constricţia primară, determinantă a poziţiei centromerului, cromosomii au şi constricţii secundare, cu rol în formarea nucleolului, fapt pentru care au primit şi denumirea de organizatori nucleolari. Uneori, cromosomii au la unul din capete o constricţie secundară prin care se delimitează un segment numit satelit (trabant). Asemenea cromosomi se întâlnesc la speciile Secale cereale (2n=14 - cromosomii perechii a VII-a), şi Zea mays (cromosomii perechii a
VI-a). Numărul nucleolilor dintr-un nucleu este egal cu numărul cromosomilor cu satelit. Cele
două specii menţionate mai sus au, deci, câte doi nucleoli în nucleu. Centromerul are rolul de a fixa cromosomul pe fibra fusului acromatic, în timpul diviziunii
celulare. Prin diviziunea sa, care precede diviziunea cromosomului şi separarea cromatidelor, în mitoză sau în cea de-a doua diviziune meiotică, se asigură partiţia cromosomului în cele două cromatide surori. În cazurile în care, din diverse motive, cromosomul se rupe, fragmentele fără centromer (numite acentrice) nu se pot reface şi se resorb.
Un fragment acentric se păstrează doar atunci când se ataşează de un cromosom cu centromer. Sunt însă şi cazuri (coccidii, scorpioni, specii de Luzula) în care unii cromosomi, extrem de scurţi, îndeplinesc funcţii centromerice în totalitate sau pe mare parte din lungimea lor. De fapt aceşti cromosomi au centromerul difuz şi poartă denumirea de cromosomi policentrici. Ei sunt apţi, însă, de a se fixa pe toată lungimea lor de fibrele fusului acromatic.
La capetele cromosomilor există o zonă (o porţiune) care le împiedică unirea. Respectiva porţiune poartă numele de telomer (telomere). Dincolo de rolul de a împiedica unirea
cromosomilor prin capetele lor, telomerele au şi un rol, din ce în ce mai bine definit, în determinismul longevităţii organismelor. Fiind zone heterocromatice ale cromosomilor, telomerele au acidul dezoxiribonucleic super condensat (super răsucit). S-a constatat că, la diverse specii, lungimea telomerelor este foarte diferită şi că numărul de diviziuni succesive ale unei celule este în relaţie de directă proporţionalitate cu lungimea telomerelor. Când, în succesiunea de diviziuni, telomerul dispare, cromosomul se autolizează iar celulele se distrug.
Schematic (Fig.4), deci, un cromosom apare astfel: telomer, satelit, constricţie secundară,
braţ scurt, centromer (constricţie primară), braţ lung, telomer (vezi schema anterioară). Fiecare braţ este alcătuit din două cromatide, fiecare cromatidă conţinând două cromoneme. Regiunea cromosomială din vecinătatea centromerului poartă numele de regiune proximală, iar cea de la capetele braţelor poartă numele de regiune distală. Aşa cum am precizat anterior, cromomerele
sunt, de fapt, nişte artefacte. În ultimul timp (detaliile vor fi date în paginile următoare) este contestată şi realitatea cromonemei.
Precizam anterior că tipul cromosomilor reprezintă un caracter de diagnoză, o caracteristică a speciei. Fără a contrazice această afirmaţie, se cuvine precizat că tipul (forma) cromosomilor poate varia între anumite limite, în funcţie de specializarea şi starea fiziologică a celulei. Uneori, în cadrul aceluiaşi individ, se constată şi o variabilitate numerică a cromosomilor, tot în corelaţie cu structura şi funcţiile anumitor ţesuturi.
De regulă, cel mai propice stadiu pentru investigarea numărului şi tipului cromosomilor îl reprezintă metafaza diviziunii mitotice. Puternic spiralizaţi, ataşaţi de fibrele fusului în zona ecuatorială, în această fază cromosomii etalează conformaţia lor specifică, formă caracteristică speciei căreia îi aparţine individul (pot avea formă de bastonaş sau forma literelor V, J ,I ,O etc.)
Fig. 3. Tipurile de cromosomi. 1 - cromosom M (metacentric); 2 - cromosom m (median); 3 - cromosom sm (submedian); 4 - cromosom st (subterminal = subtelocentric); 5 - cromosom T (telocentric). Prin culoarea gri a
coloanei se redã zona în care se poate găsi centromerul. Desigur, braţul de sus se scurteazã cu porţiunea cu care se alungeşte cel de jos, în aşa fel încât raportul dintre ele sã se situeze între limitele precizate anterior (după Băra,
1999)
3.5.3.2. Mărimea cromosomilor eucariotelor
Este o caracteristică de specie, deşi poate varia destul de mult în cadrul unei populaţii, în cadrul unui individ şi chiar în aceeaşi celulă, în funcţie de faza de diviziune în care se află aceasta din urmă. Lungimea cromosomilor variază şi în funcţie de biotopul pe care-l ocupă o populaţie în cadrul arealului speciei, fiind influenţată de temperatură, anumite substanţe chimice etc.. În general, mărimea cromosomilor variază între 1 şi 50 microni pentru lungime şi între 0,1 şi 2 microni pentru diametru.
În principiu se admite că lungimea cromosomilor este direct proporţională cu numărul de gene pe care le conţin. Realitatea este, însă, mult mai complicată.
3.5.3.3. Numărul cromosomilor la eucariote
Este un alt caracter de diagnoză, deşi cunoaşte şi el o amplitudine de variabilitate suficient de largă. În lumea vie numărul de cromosomi variază între 2, la Ascaris megalocephala şi câteva sute, la Amoeba proteus. Pentru caracterizarea unei specii, sub aspectul numărului de cromosomi, se folosesc simbolurile: x, n, 2n şi NF.
- 2n reprezintă numărul de cromosomi din celulele somatice ale indivizilor oricărei specii
eucariote; - prin n se desemnează numărul haploid de cromosomi, caracteristic celulelor gametice
(sexuale) ale indivizilor speciilor eucariote; De regulă, în cadrul unui gen, totalitatea speciilor provine dintr-o specie ancestrală, prin
evoluţie divergentă. Procesul a implicat, printre altele şi modificarea numărului de cromosomi, prin autopoliploidizare, pseudopoliploidizare, aneuploidizare, amfipoliploidizare etc.. Numărul cromosomial de la care s-a plecat, în cazul unei serii poliploide de pildă, reprezintă numărul cromosomial de bază, notat cu x. La diploidul de origine, numărul de bază coincide cu numărul haploid (x = n). Evident, x rămâne acelaşi pentru triploid, tetraploid etc., n modificându-se în concordanţă cu 2n. Spre exemplu, în genul Papaver, secţia Oxytona există speciile Papaver
Fig. 4. Reprezentarea schematicã a morfologiei şi organizăriii cromosomului la eucariote (prelucrare dupã Tudose, 1992)
bracteatum (2n = 14, n = 7, x = 7), Papaver orientale (2n = 28, n = 14, x = 7) şi Papaver pseodo-orientale (2n = 42, n = 21, x = 7). Este clar că n variază în funcţie de 2n, în timp ce x (numărul fundamental de cromosomi) rămâne acelaşi, cele trei specii fiind 2x, 4x şi 6x.
Alteori, o serie poliploidă nu provine printr-o poliploidizare reală, ci prin pseudopoliploidie. De exemplu, dacă o specie are 2n = 8 cromosomi metacentrici, mediani, submediani sau subterminali, prin fisionarea cromosomilor dintr-o pereche, două, trei sau toate perechile, vor rezulta 2n = 10, 2n = 12, 2n = 14, sau 2n = 16 cromosomi. Dacă, însă, se va determina cantitatea de ADN per nucleu, se va constata că rămâne constantă, la toate aceste numere de cromosomi. Pe de altă parte, dacă se vor număra braţele cromosomiale, din cariotipurile efectuate pe baza numerelor menţionate, se va constata că cifra este aceeaşi – 16. Prin urmare NF (numărul fundamental de braţe) a rămas constant, sugerând că speciile din respectiva serie nu sunt
aneuploizi sau poliploizi adevăraţi ci pseudoaneuploizi sau pseudopoliploizi.
3.5.3.4. Structura internă a cromosomilor la eucariote
Un real progres în studiul cromosomilor mamiferelor s-a înregistrat concomitent cu punerea la punct a metodei Giemsa de colorare şi examinare a cromosomilor. Apoi, definitivarea metodelor de bandare a cromosomilor, de către Arrighi şi Hsu, în 1971, urmată de unele modificări aduse metodei Giemsa, au permis stabilirea existenţei diferitelor tipuri de cromosomi, în cadrul unuia şi aceluiaşi genom. Mileniul 3 a debutat cu cea mai mare realizare în domeniu – descifrarea genomului uman. S-a stabilit, cu această ocazie, că omul nu deţine peste 100.000 de gene, cum se afirma frecvent, ci aproximativ 35.000. Prin urmare, extraordinara complexitate a organismului uman nu poate fi pusă pe seama unui număr mare de gene ci pe interacţiuni multiple şi complexe între un număr de gene aproape egal cu numărul genelor deţinute de o muscă.
În ceea ce priveşte compoziţia chimică a cromosomilor eucariotelor, încă din anul 1920 s-a stabilit că principala componentă o constituie substanţa cromatică, denumită cromatină,
alcătuită, la rândul ei, din aciizi nucleici şi proteine, în proporţii egale (câte 50% pentru fiecare dintre ele). Organizarea structurală intimă a acestor două componente a fost descifrată abia în jurul anilor 1960. Un progres considerabil l-a reprezentat identificarea componentelor proteice majore ale cromatinei. S-a constatat că acestea sunt proteine histonice, de cinci tipuri, notate cu H1, H2A, H2B, H3 şi H4. Aceste cinci histone sunt prezente în cromosomii tuturor eucariotelor.
Interesant este faptul că aceste proteine au secvenţă aminoacidică asemănătoare la majoritatea plantelor şi animalelor, ceea ce poate sugera că gena care este responsabilă de biosinteza lor are o origine foarte îndepărtată (este foarte veche) şi nu a suferit prea multe mutaţii, pe întregul parcurs al procesului filogenezei.
Un alt progres important a fost constatarea că patru dintre histonele menţionate (H2A, H2B, H3 şi H4) formează octameri, particule în jurul cărora se înfăşoară ADN-ul. Rezultă o formaţiune care, împreună cu H1 alcătuieşte nucleosomul.
Nucleosomul este considerat, deocamdată, structura de bază a cromosomului. Analizele cu raze X au dovedit că nucleosomii sunt legaţi între ei prin ADN neasociat cu histone (în unele manuale de specialitate sunt şi alt tip de afirmaţii), fiind sub forma unei fibrile de cca 10 nm grosime. Această fibrilă se continuă de la un capăt la altul al cromosomului, pliată (împachetată) foarte compact. Cromosomii sunt, deci, cromatină suprastructurată.
Cromatina este numele conferit substanţei totale din care sunt alcătuiţi cromosomii. În
ciclul mitotic sau meiotic, cromatina se colorează rapid şi este uşor vizualizată. În interfază este vizibilă doar heterocromatina, partea vizibilă în timpul diviziunii primind numele de eucromatină (ambele noţiuni vor fi analizate, pe larg, ulterior). Regiunile heterocromatice ale cromosomilor sunt considerate ca fiind heteropicnotice (în limba greacă pycnosis =dens). Intensitatea colorării lor
poate varia de la o etapă a ciclului celular la alta. Fiecare cromosom dintr-o celulă eucariotă, conţine o macromoleculă lungă de ADN, care
se întinde de la un capăt al cromosomului la celălalt, prin centromer. Această macromoleculă gigantică este superîmpachetată, în cromosom, în numai câţiva microni. La om de exemplu, cel mai mare cromosom conţine o macromolecul de ADN de aproximativ 85 mm (85.000 μm sau 8,5 x 107 nm). Această macromoleculă gigantică este împachetată într-un spaţiu care atinge, în metafază, 0,5 μm în diametru şi 10 μm în lungime, adică o împachetare de aproxomativ 104 ori.
Realizări remarcabile în descifrarea infrastructurii cromosomilor se datoresc cercetărilor efectuate de Hewish & Brugoyne (1973) care au constatat că prin tratarea cromatinei cu
endonucleaze activate cu Ca++ şi prin supunerea produselor rezultate la electroforeză, se obţineau repetiţii de fragmente de ADN de aproximativ 200 pb (perechi de baze), repetiţii ce s-au dovedit a fi un constituient permanent şi ubiquitar al cromosomilor, cunoscut sub numele de nucleosom.
Pe de altă parte, prin microscopie electronică s-au identificat bobiţe elipsoidale de aproximativ 11 nm în diametru şi 6 nm în înălţime, legate între ele prin fire subţiri de acid dezoxiribonucleic. Aceste bobiţe nu sunt altceva decât nucleosomii. Pe de altă parte, digestia cromatinei cu nucleaze a evidenţiat existenţa unor formaţiuni de aproximativ 146 pb care nu sunt disociate prin digestie. Mai mult decât atât, uneori, un set de asemenea formaţiuni rămâne intact. Conform datelor de microscopie electronică, aceste seturi sunt asociaţii de nucleosomi.
Nucleosomii reprezintă peste 95% din greutatea cromatinei şi sunt consideraţi a fi unităţile structurale de bază ale cromatinei. Structura lor poate fi redată schematic (Fig.5).
În celulele eucariotelor cu organizare superioară, nucleosomii conţin aproximativ 195 - 200 pb de ADN, un octamer histonic alcătuit din câte două molecule de H2A, H2B, H3 şi H4 şi o moleculă de H1. Acesta nu este însă un nucleosom complet deoarece în figură nu este precizat şi un fragment de 27 pb, fragment care nu este direct asociat cu octamerul histonic. Acest fragment conectează, între ei, doi nucleosomi consecutivi. În majoritatea cazurilor însă, octamerul histonic are în jurul lui aproximativ 1,7 ture de ADN (adică 146 pb). Octamerul histonic împreună cu cele 146 perechi de nucleotide alcătuiesc miezul, sâmburele, particula centrală a nucleosomului, cunoscut sub numele de nucleosomal sau histone core particle (Richmond et al., 1984). Acesta
este modelul nucleosomal propus pe baza determinărilor efectuate în soluţie cu dispersie de neutroni, dar şi pe baza analizelor efectuate prin difracţia razelor X (Bradbury & Baldwin, 1966; Klug et al., 1985; Burlingame et al,1985). Astfel, în conformitate cu acest model, există un disc de 11,0 nm în diametru şi 5,5 - 6,0 nm înălţime, care are 1,7 ture de acid dezoxiribonucleic, cu o grosime de 3,0 nm, înfăşurat în jurul lui. Cele 146 de pb ale acidului dezoxiribonucleic nu sunt repartizate uniform în jurul octamerului histonic. Tetramerii 2H3 şi 2H4 interacţionează cu ADN-ul şi-l răsucesc în jurul lor. În acest fel se completează structura nucleosomală. S-a demonstrat că partea centrală a octamerului are histonele H3 şi H4, celelalte două histone fiind la exteriorul lor (pe feţele discului) – aşa cum este redat în modelul structurii discului octameric (Fig.6).
Rezumând, putem preciza că există trei etape în structurarea unui nucleosom şi anume:
1. inima nucleosomului, alcătuită din 146 pb şi octamerul histonic (câte 2 molecule de H2A,
H2B, H3 şi H4); 2. inima plus încă 22 pb, dând un total de 168 pb, plus histona H1, alcătuieşte
cromatosomul;
3. cromatosomul plus încă aproximativ 32 pb (ajungându-se la un total de aproximativ 200 pb.), formează nucleosomul. Trebuie precizat că, deocamdată, nu este precis stabilit dacă există o singură moleculă H1
per nucleosom. Lungimea linkerilor dintre cromatosomi variază atât de la specie la specie, cât şi de la un tip celular la altul. S-au descris linkeri în lungime de numai 8 pb,dar şi linkeri de 114 pb. Nucleosomul complet conţine, spre deosebire de inima nucleosomului, două ture complete de ADN. Se mai afirmă că structura şi rolul nucleosomilor diferă în regiunile genetic active de cele din
Fig.5 Diagrama unui nucleosom în jurul căruia se înfăşoarã ADN pe aproximativ două ture (model propus de Brandbury, 1992)
regiunile inactive. Interacţiunile dintre ADN şi histone, în cadrul nucleosomului, sunt de natură electrostatică, implicând părţile aminice, cu bazicitate crescută, din lanţurile de arginină şi lizină, precum şi unele legături slabe ale histonelor bazice, cu părţile fosfat ale ADN-ului (Nieto & Palacian, 1988).
Nucleosomii împreună cu acidul dezoxiribonucleic linker alcătuiesc o fibră cromatică de 11
nm grosime care se spiralizează, la rândul ei, dând un solenoid de 30 nm grosime. Cercetările nu au dat încă un răspuns clar şi definitiv relativ la structurarea solenoidului de 30 nm grosime (Fig.7).
Când concentraţia ionică a mediului creşte, apare aşa numita formă înaltă a cromatinei
(Thoma & Koller, 1977). La concentraţia de 2 mM de electrolit de MgCl2 şi în prezenţa histonei H1 se poate pune în evidenţă această fibră de 25 - 30 nm grosime. În literatura de specialitate sunt menţiuni privitoare la aceste fibre (McGhee & co., 1983; Felsenfield & McGhee, 1986). Analiza acestor fibre, prin microscopie electronică, cu raze X şi cu neutroni, a relevat o structură prezentă la toate metazoarele şi la toate embriofitele, structură existentă în cromosomi - atât în cei interfazici, cât şi în cei metafazici. Se admite că această structură reprezintă un element de bază în cromosomii tuturor eucariotelor, cu eventuale excepţii pentru unele protiste şi pentru unele talofite.
Fig.6 Modelul structurii miezului octameric, cu aranjarea probabilă a histonelor. a - ADN-ul reacţionează cu tetramerii H3, H4 şi formează sâmburele discului nucleosomal, b - discul cu histonele H2A şi H2B, asociate în exteriorul
ADN-ul din componenţa solenoidului este, la rândul lui, constituit din două părţi - o parte
ataşată la miezul histonic şi o altă parte cu rol spaţiator (alţi autori consideră, dimpotrivă, că are rol de linker), care asamblează nucleosomii în solenoid. Aşa cum precizam mai sus, segmentele spaţiatoare (sau linkeri) sunt de lungimi variabile. Se pune doar o problemă - cum sunt organizate aceste segmente? Unii autori (McGhee et al., 1983) apreciază că ADN-ul spaţiator este superrăsucit. Linia AB, care trece prin centrul cromatosomilor adiacenţi, este înclinată cu peste 100 faţă de linia CD, care este perpendiculară pe axa solenoidului. Aceasta determină deschiderea unghiului ADN-ului spaţiator supercondensat care, la rându-i, determină distanţa dintre nucleosomii vecini (unghiul δ).
Pe de altă parte, se cuvine menţionat faptul că structurile aparente la acest nivel de rezoluţie microscopică, reprezintă implicit şi un rezultat al metodei de analiză folosite, metodă care presupune tratamente specifice (metode specifice de preparare a materialului). Prin urmare, se pot aştepta rezultate întrucâtva diferite, de la un laborator (grup de cercetare) la altul. Toate rezultatele, însă, converg spre concluzia că fibra de 30 nm este o realitate (există în realitate), rămânând deschisă doar problema structurii sale.
La data prezentă, structura solenoidului pusă în evidenţă cu raze X, neutroni, microscopie electronică etc., pare a avea un suport faptic suficient de solid.
Structurarea ulterioară a cromatinei (organizarea solenoidului), pentru a forma cromosomii vizibili în metafază (sau regiunile cromosomiale heteropicnotice vizibile şi în interfază) (Fig.8), este
suficient de bine cunoscută şi explicată. Părerile, ca şi argumentele, celor mai mulţi cercetători converg către concluzia în conformitate cu care fibra cromatică de 30 nm grosime se pliază şi formează bucle (laţuri), asigurând astrfel structurarea cromosomilor. Pentru a avea o reprezentare cât mai reală a ratei de compactare a fibrei cromatice în cadrul unui cromosom, recurgem la exemplul dat de Wagner, Maguire & Stallings (1993) şi anume. Cromosomul 16 din genomul uman conţine aproximativ 1,1 x 108 pb, ceea ce reprezintă aproximativ 3,7 x 104 μm lungime (pentru respectiva fibră de cromatină). Dar, în metafază, când condensarea este maximă, lungimea atinge doar 3 μm. Se asigură, deci, o scurtare (datorată compactării) de peste 12.000 de ori (adică 3,7 x 104 μm / 3 μm = 12.000). În prometafaza timpurie, acelaşi cromosom (cromosomu l 16) este de 4 μm lungime (adică s-a scurtat, prin compactare, de 9250 de ori). Pe baza acestui tip de calcule s-a conchis (Manuelidis & Chen, 1990) că diferenţa între rata scăderii lungimii fibrei cromatice de 30 nm, prin compactarea în interiorul cromosomului şi rata scăderii lungimii cromosomului de la interfază la metafază este de 7,5. De aceea cromosomul 16, în interfază, are o lungime de aproximativ 23 μm (corespunzând unei rate de condensare de aproximativ 1600).
Fig. 7. Diagrama solenoidului. Sunt 6 nucleosomi per tură. Nucleosomii au o înălţime de 5,7 nm şi un diametru de 11 nm. ADN-ul se continuă de la un nucleosom la altul (linia întreruptă) (după Widom & Klug, 1985)
Prin urmare, întrebarea care se pune este următoarea: cum se ajunge de la o lungime de
3,7 x 104 μm la una de 23 μm? Cele mai multe imagini, obţinute la microscopul electronic cu scanare, relevă situaţia compactării fibrei de ADN. Se observă plieri şi contorsionări ale fibrei de cromatină, care explică acceptabil scurtarea cromosomilor. Dinamica spiralizării, de la solenoid la cromosomul metafazic, este conformă cu concepţia relativă la structura internă a cromosomului. De aceea, o etapă în compactarea cromosomului este asigurată prin înfăşurarea ADN-ului în jurul miezului nucleosomal şi, apoi, prin spiralizări care duc la formarea solenoidului. Se asigură, astfel, o rată a împachetării de aproximativ 40. Deseori, restul compactării se asigură prin alte procese.
Cromosomii metafazici reprezintă condensarea cromatică maximă (observată la cromosomii eucariotelor). Rolul acestor formaţiuni este acela de a organiza şi împacheta macromolecula gigantică de ADN, permiţând segregarea către nuclei celulelor fiice.
Cromosomii metafazici, care sunt eliberaţi de aproximativ 99% din conţinutul lor histonic, îşi păstrează ADN-ul în formă aproximativ constantă (asemenea cromosomilor iniţiali, intacţi), probabil tocmai din cauză că ADN-ul este ataşat, în numeroase puncte, la scheletul proteic nonhistonic intern (Paulson & Laemmli, 1977). Acest schelet constă, în primul rând, din două proteine, desemnate prin Sc1 şi Sc2, cu greutăţi moleculare de 170 kD şi, respectiv, de 135 kD (Lewis & Laemmli, 1982). Sc1 este, în fapt, Topoizomeraza II (Earnshaw et al., 1985), buclele radiale ale ADN-ului fiind ataşate la această proteină, în punctele numite SAR (în engleză scaffolding attaching regions - regiuni de ataşare la schelet) (Earnshaw & Heck, 1985; Gasser & Laemmli,
1986).
3.5.3.5. Eucromatina şi heterocromatina
Examinând cu atenţie cromosomii, pe lungimea lor, constatăm că nu sunt uniformi sub aspectele chimice, fizice şi genetice. În primul rând, diferă reacţia lor la tratamentul cu coloranţi bazici, în sensul că unele segmente se colorează slab, cunoscute fiind sub numele de eucromatice, iar altele se colorează mult mai intens şi au fost desemnate cu numele de heterocromatice.
Interesant şi important de consemnat este faptul că zonele heterocromatice nu conţin gene în stare funcţională, fiind considerate genetic inerte. În mod artificial, segmentele eucromatice pot fi introduse în segmentele heterocromatice, operaţiune prin care se obţin schimbări comportamentale ale ambelor categorii de segmente.
Fig.48. a. Cromosomi metafazici de câine, văzuţi la microscopul electronic cu baleiaj (SEM); b. Cromosom metafazic de hamster chinezesc (după Wagner et al., 1993)
Eucromatina reprezintă materialul normal, izopicnotic, deţinătorul informaţiei genetice, cu comportament tipic în cazul diviziunii celulare (se spiralizează, se condensează, se decondensează şi se colorează). La rândul ei, eucromatina este de două tipuri: eucromatina activă şi eucromatina permisivă.
Eucromatina activă conţine genele ce vor fi transcrise în ARNm. Eucromatina permisivă este reprezentată de acea porţiune din eucromatină care devine
activă doar după ce acceptă (permite) semnale declanşatoare (din categoria hormonilor, enzimelor etc.). Procesul autoreplicării semiconservative a ADN-ului, în faza S din ciclul diviziunii celulare, începe la nivelul eucromatinei. În consecinţă, replicarea eucromatinei este mult mai timpurie, în comparaţie cu cea a heterocromatinei.
Heterocromatina reprezintă materialul unor regiuni (uneori al unor întregi cromosomi) heteropicnotice, caracterizate prin structură densă şi compactă, inclusiv în telofază, interfază şi profaza timpurie. Din aceste cauze, heterocromatina se colorează intens şi este vizibilă şi în interfaza ciclului celular (în nucleii celulelor în interfază).
Zonele heterocromatice sunt răspândite pe întreaga lungime a cromosomului, dar mai ales în jurul centromerului, unde formează heterocromatina centromerică. Destul de frecvent este localizată şi în apropierea organizatorilor nucleolari şi spre capetele cromosomilor. Cu această ocazie se poate menţiona faptul că în aceste zone cromosomiale (heterocromatice) au loc cele mai frecvente ruperi ale cromosomilor.
Heterocromatina este componenta preferenţială a cromosomilor sexuali şi a celor suplimentari (cromosomii B). Sunt unele specii (broaştele ţestoase, unii viermi) la care întregul set cromosomial este heterocromatic.
Replicarea ADN-ului, în zonele eucromatice şi heterocromatice, se desfăşoară asincron. În ceea ce priveşte heterocromatina, însă, apar şi unele aspecte deosebit de interesante.
De pildă, în stadiile timpurii de dezvoltare embrionară, heterocromatina lipseşte. Deci, ca o concluzie firească, se poate accepta că ea nu se transmite de la o generaţie la alta, ci se formează într-un anumit stadiu al ontogeniei. După ultimele observaţii şi experimente, se pare că orice regiune a cromosomului poate deveni, la un moment dat, heterocromatică. În consecinţă, heterocromatina şi eucromatina trebuiesc privite nu ca unităţi distincte şi discontinue ale cromosomilor, ci stări ale cromatinei, dinamice atât în timp cât şi în spaţiu.
Sunt şi autori care consideră că există o heterocromatină nativă, reprezentată prin
segmente cromosomiale heterocromatinizate, încă de la naşterea organismului şi o heterocromatină funcţională, care se individualizează temporar.
O altă clasificare a tipurilor de heterocromatină a fost propusă de Brown, în 1965. În conformitate cu această clasificare, heterocromatina poate fi constitutivă şi facultativă.
Heterocromatina constitutivă este identificată în cromosomul Y de la Drosophila melanogaster sau de la alte specii şi are ca trăsătură definitorie faptul că se constituie încă de la începutul vieţii individului şi rămâne în această stare (genetic inactivă) pe tot parcursul dezvoltării individuale. Heterocromatina constitutivă poate fi prezentă şi în alţi cromosomi, fiind localizată (de obicei) de ambele părţi ale centromerului.
Heterocromatina facultativă este considerată cea din unul dintre cei doi cromosomi X, de la femelele de mamifere de pildă, care devine genetic inactivă şi se evidenţiază în nucleul interfazic sub forma unui corpuscul intens colorat, cunoscut sub numele de cromatină sexuală sau corpuscul Bar, bastonaşul de tobă (drum-stick) etc. În acest context, unul dintre cromosomii X devine nefuncţional, asigurându-se astfel egalitatea între cele două sexe. Fenomenul este cunoscut sub numele de compensaţie de doză. Important este faptul că heterocromatina facultativă are
capacităţi reversibile, astfel încât, în momentul în care cromosomul X inactivat de la femelă ajunge să fie unicul cromosom X de la mascul, el devine funcţional. Dar el devine activ şi în ovulul în care, din întâmplare, nimereşte. Apoi, dacă prin fecundarea respectivului ovul se ajunge din nou la un zigot femel, iniţial, acesta va avea ambii cromosomi X funcţionali. În stadiul de blastocist, unul dintre ei devine nefuncţional - capătă statutul de cromatină sexuală pentru întreg ciclul ontogenetic.
Iniţial se considera că zonele heterocromatice nu conţin gene funcţionale. Dar, prin studii efectuate pe tomate, în 1961, s-a demonstrat că în heterocromatină există gene funcţionale - în heterocromatina centromerică.
Heterocromatina poate apărea supercondensată, în unele regiuni cromosomiale, cum este cazul la Zea mays. Aici apar un fel de noduli, denumiţi knobi, în care activitatea genică este
complet blocată. Numărul şi topografia knobilor sunt constante pentru anumiţi cromosomi.
Deocamdată nu se cunoaşte rolul concret al knobilor şi nici dacă au sau nu gene cantonate pe ei. Sunt doar, presupuneri cu privire la rolul lor în procesele de gametogeneză.
3.5.3.6. Categorii de cromosomi
a. Cromosomii principali. Sunt cromosomii cu cea mai largă răspândire, altfel spus,
cromosomii a căror prezenţă este considerată normală pentru celulele somatice şi pentru cele sexuale normale şi sunt grupaţi în două clase: cromosomii A şi cromosomii B.
I. Cromosomii A sunt cromosomii tipici, normali ai complementului cromosomial ce
defineşte o specie. Ei se comportă constant (de obicei) atât sub aspect tipologic (M, m, sm, st şi T), cât şi sub aspect numeric.
II. Cromosomii B, supranumiţi şi cromosomi accesorii sau suplimentari, sunt cromosomii aflaţi în exces, putând fi în număr variabil, atât la plante, cât şi la animale. Un exemplu tipic de plantă cu cromosomi B este Zea mays.
Numărul cromosomilor B este extrem de variabil, nu numai de la individ la individ, ci şi între ţesuturile aceluiaşi individ. Cromosomii B nu manifestă fenomenul de omologie, nici între ei şi nici cu cromosomii A. Sunt, de obicei, heterocromatici, mici şi inerţi din punct de vedere informaţional. În diviziunea celulară, distribuţia lor nu este egală către celulele fiice.Nu li se cunoaşte originea, ci se presupune doar că au apărut din regiunile centrale ale unor cromosomi A, prin pierderea de către aceştia a extremităţilor (extremităţi ce s-au putut ataşa la alţi cromosomi A, modificându-le constituţia şi, deci, conţinutul informaţional). În context, trebuie menţionat că nu li se cunoaşte nici rolul, deoarece lipsa lor (parţială sau totală) nu perturbă existenţa individului purtător. Dimpotrivă, uneori, prezenţa lor este dăunătoare.
b. Cromosomii secundari, adică acei cromosomi care coexistă cu cromosomii A şi B,
numai în anumite celule. În această categorie intră cromosomii uriaşi sau politenici din glandele salivare de la
Drosophila melanogaster, Chironomidae şi alte insecte, cromosomii lampbrush (perie de sticlă de
lampă) prezenţi în oocitele vertebratelor, în profaza meiozei etc. 1. Politenicii sau cromosomii uriaşi au fost descoperiţi de către E. Balbiani (citolog
italian), în 1881, în glandele salivare de Chironomus. Ca dimensiune, ei depăşesc de 100-200 ori
lungimea cromosomilor normali, conţinând de peste 1000 de ori mai multe cromoneme. Cromosomii politenici au fost depistaţi şi în unele celule somatice ale larvelor de diptere - cum ar fi tubul digestiv, tubulii lui Malpighi etc., fiind prezenţi chiar şi la unele plante (în sinergide) şi la alte specii de nevertebrate. Evident, din considerente didactice, cromosomii politenici tipici sunt exemplificaţi prin cei din glandele salivare de Drosophilla melanogaster.
Cauza apariţiei acestor cromosomi este relativ simplă - celula în care ei se găsesc, nu se divide, ci doar îşi măreşte dimensiunile. Nucleul respectivelor celule se găseşte într-o perpetuă stare de interfază (pe întreaga durată a stadiului larvar). Pe de altă parte, fenomenul are şi o altă cauză. Cromosomii perechi se apropie şi se unesc. Este fenomenul cunoscut sub numele de conjugare somatică. Concomitent cu creşterea larvei, fiecare cromosom se măreşte, prin replicarea cromatidelor şi, implicit, a cromonemelor, fără migrarea lor la polii celulari. În final, fiecare cromosom ia aspectul unui fascicul de filamente (filamentele reprezentând cromonemele), de unde şi numele lor de politenici.
În esenţă, avem un fenomen tipic de endomitoză - adică reduplicarea cromosomilor fără separarea şi migrarea lor.
De obicei, fiecare cromosom uriaş conţine mai mult de 1000 cromoneme (subcromat ide), care sunt vizibile ca filamente separate. Spiralizarea lor, pe lungimea cromosomului, este inegală. În plus, au şi o structură chimică diferită de la o pereche la alta, ceea ce conferă filamentului un aspect specific. Spiralizarea inegală, local mai compactă, a cromonemelor, induce apariţia unor formaţiuni caracteristice, cunoscute sub numele de discuri. Discurile pot avea dimensiuni şi structuri diferite. Unele discuri conţin mai mult ADN şi, în consecinţă, au o culoare mai intensă, altele conţin mai multă proteină şi, deci, au o culoare mai deschisă. Sunt şi păreri în conformitate cu care toate discurile au aceeaşi compoziţie, doar că reprezintă porţiuni mai puternic sau mai slab condensate (spiralate), de unde şi coloraţia lor diferită (Fig.9).
Cromosomii uriaşi de la Drosophila melanogaster formează o figură tipică, fiind constituiţi din cinci braţe lungi şi unul mai scurt. La punctul de întretăiere formează cromocentrul. Unul dintre braţele lungi se notează cu X şi reprezintă cromosomii X, altele două sunt notate cu 2S şi 2D, celelalte cu 3S şi 3D, iar braţul scurt cu 4.
Lungimea braţelor, însumate, ajunge la 2000 μ, adică de peste 266 ori mai mare decât
lungimea însumată a celor opt cromosomi somatici normali (7,5 μ în total). În cromosomii uriaşi (politenici), numărul cromonemelor dintr-un braţ este de
aproximativ 256, deci cu mult mai mare decât normalul (2 cromoneme / cromatidă). Altfel spus, dacă prin C redăm numărul catenelor de acid dezoxiribonucleic dintr-un cromosom normal, într-un cromosom uriaş acest număr este dublat de 9 ori succesiv, ajungând la 1024 C. Pe de altă
parte numărul de dublări succesive diferă de la un ţesut la altul, atât la larvele de drosofila cât şi la alte diptere.
2. Cromosomii lampbrush sunt tot cromosomi uriaşi. Forma lor se datorează conjugării
cromosomilor omologi într-un singur complex. În plus, cromosomii sunt puternic despiralizaţi şi formează bucle simetrice cu orientare perpendiculară pe axul complexului. Cromosomii de tip lampbrush sunt prezenţi în oocitele amfibienilor, moluştelor, echimodermelor, peştilor, reptilelor, la unele mamifere şi chiar la ceapă. O asemenea conformaţie se găseşte şi la cromosomul Y de la Drosophila melanogaster.
La unele specii, aceşti cromosomi ating lungimea de peste 1000μ, fiind consideraţi cei mai lungi cromosomi. Diametrul lor însă este foarte mic, uneori fiind atât de subţiri încât se situează la limita de rezoluţie a microscopului optic.
Având în vedere despiralizarea lor puternică, s-a emis părerea că respectivii cromosomi sunt deosebit de activi în procesul dezvoltării oocitelor. Învestigaţiile efectuate cu administrare de uridină marcată (3H), o precondiţie a autoradiografiei, au argumentat această supoziţie. În buclele cromosomilor perie de lampă are loc o intensă transcriere a informaţiei materializată în sinteza intensă de ARNm. Deci, ca o concluzie finală a problemelor analizate până aici, se poate afirma că buclele din cromosomii lampbrush, ca structură şi funcţii, sunt similare pufelor din cromosomii uriaşi.
3.5.3.7. Cariotipul şi idiograma
Când am prezentat categoriile de cromosomi, am precizat că se are în vedere existenţa unei categorii de cromosomi principali, grupaţi în clasa A, reprezentând cromosomii cu cea mai frecventă şi normală prezenţă în celulele somatice şi sexuale ale tuturor speciilor.
Detaliind, precizăm că la speciile care posedă un determinism cromosomial al sexelor, cromosomii pot fi sbîmpărţiţi în două grupe - autosomi şi heterosomi (sau alosomi, gonosomi, cromosomi ai sexului etc.). Evident, majoritatea membrilor complementului cromosomial sunt
autosomi. De obicei există doi heterosomi, excepţie făcând speciile al căror determinism cromosomial al sexelor este asigurat de heterosomi multipli şi cele cu determinism sexual numeric cromosomial.
Fig.9 Cromosomul uriaş din glanda salivară de Drosophila melanogaster. Prin X, 2R, 2L, 3R, 3L şi 4 sunt desemnaţi cromosomii X, braţele drept (R, de la englezescul right) şi stâng (L, de la englezescul left) ale perechilor 2 şi 3 şi
perechea 4 (desemnată prin 4) (după Tamarin, 1991)
Cariotipul, în concordanţă cu raportul Conferinţei Internaţionale de la Denver (din 1960), întrunită pentru standardizarea cromosomilor umani, poate fi definit astfel: ordonarea perechilor de cromosomi omologi, provenind dintr-o singură metafază mitotică, în ordinea descrescândă a lungimii lor, cu centromerul plasat pe o linie imaginară (Fig.10, 11).
Idiograma reprezintă redarea grafică a lungimilor medii ale cromosomilor fiecărei perechi de omologi, obţinute pe baza a cel puţin 10 metafaze, din celule diferite provenite de la indivizi diferiţi (Fig.12). Se înţelege că idiograma este mult mai reprezentativă pentru
caracterizarea cromosomologică a unei specii. Modalităţile practice de alcătuire a cariotipului şi idiogramei au fost precizate de lucrările de
laborator (se găsesc în orice manual de lucrări practice), fapt pentru care nu insistăm. Se impun doar, cu această ocazie, câteva precizări.
În stabilirea corectă a perechilor de cromosomi omologi, constricţiile secundare pot juca un rol definitoriu. De obicei, constricţiile secundare nu afectează aceleaşi regiuni la diverşii cromosomi. În plus, s-a constatat că aceste constricţii secundare sunt mult mai uşor observabile dacă se administrează cromosomilor unele tratamente speciale (cu agenţi chimici, fizici sau biologici, cum ar fi: hidrazida maleică, 5-bromdezoxiuridina, virusul SV40sau al herpesului, dacă se cultivă celulele pe mediu fără calciu, dacă se asigură temperaturi scăzute etc.).
Mai recent s-au elaborat şi alte metode de identificare exactă a cromosomilor. Una dintre acestea este cea a bandării. Prin bandare, fiecare regiune a unui anumit cromosom etalează un model caracteristic de benzi transversale, ceea ce permite identificarea cu mare siguranţă a cromosomilor, facilitând observarea şi cuantificarea diferitelor aberaţii structurale sau numerice.
O altă tehnică, extrem de modernă, presupune determinarea conţinutului de ADN în lungul unui cromosom. În acest sens, se recurge la microspectrofotometrie.
Fig. 10. Metafază la Salmo gairdneri Rich (după Minciu şi Băra, 1986)
Fig.11. Cariotipul speciei Salmo gairdneri Rich., efectuat pe baza metafazei anterioare (după Minciu şi Băra, 1986)
INTREBARI DE VERIFICARE
1. Definirea notiunea de cromosom 2. Precizarea caracteristicilor materialului genetic al virusurilor si viroizilor 3. Caracterizarea cromosomului bacterian si a modului sau de impachetare 4. Cromosomul la eucariote – structura 5. Nucleosomul 6. Tipuri de cromosomi 7. Cariotipul si idiograma
TEME PENTRU EXAMINAREA FINALA
1. Intelegerea notiunii de cromosom 2. Prezentarea diferentelor in privinta materialului genetic intre procariote si eucariote 3. Aprofundarea aspectelor legate de structura, ultrastructura si functiile cromosomilor la eucariote 4. Nucleosomii, eucromatina si heterocromatina 5. Tipuri de cromosomi, cariotipul si idiograma
Unitatea de invatare 3
4. Teoria cromosomiala a ereditatii (morganismul)
4.1. Teoria cromosomială
4.1.1. Momentul Morgan
Profesor la Universitatea Columbia, din New York, Thomas Hunt Morgan (1866-1945) a sintetizat într-o teorie unitară multitudinea cercetărilor din domeniul geneticii, elaborând ceea ce a rămas în ştiinţă sub denumirea de "TEORIA CROMOSOMIALĂ A EREDITĂŢII". Răsplata
imediată pentru demersul său a fost conferirea premiului Nobel (1935), răsplata perenă fiind înscrierea sa în galeria somităţilor ştiinţei mondiale.
Dăruit cu geniu, dar şi cu intuiţie, a ştiut să-şi aleagă obiectul de studiu (Drosophila melanogaster) şi colaboratorii (C.B. Bridges, A.H. Sturtevant şi H. Muller), cu care a creat o
adevărată şcoală şi un autentic sistem de cercetare, viabil, atât ca profunzime cât şi ca perspectivă.
Thomas Hunt Morgan (1866-1945), în laboratorul său din Universitatea Columbia
4.1.2. Obiectul studiului
Fără a intra în prea multe detalii, unele dintre ele fiind pe larg analizate în cadrul lucrărilor de laborator, reamintim câteva trăsături ale musculiţei de oţet, care au "recomandat-o" ca material optim pentru investigaţiile de citogenetică.
Această specie a fost descrisă la mijlocul secolului al XIX-lea sub numele de Drosophila ampelophila, ceea ce înseamnă „iubitoare de viţă de vie” (grecescul ampelos = viţă de vie, philos = iubitor). Ulterior i s-a dat numele de Drosophila melanogaster, adică „iubitoare de rouă, cu abdomenul negru” (grecescul drosos = rouă, apă, lichid; philos = iubitor; melas = negru; gastris =
stomac). Conform lucrării „An introduction to the Study of Insects” ediţia din 1964 revizuită de D. J.
Borror şi de D. M. De Long, specia Drosophila melanogaster aparţine genului Drosophila din Familia Drosophilidae, Grupul Acalyptratae, Secţiunea Schizophora, Subordinul Cyclorrhapha; Ordinul Diptera, Diviziunea Endopterigota (Holometabola), Subclasa Pterygota, Clasa Insecta (Hexapoda), Încrengătura Artropoda.
Drosophila melanogaster are aproximativ 2 mm lungime, corpul de culoare gri-gălbuie şi doi ochi compuşi mari, de culoare roşie-cărămizie (la tipul sălbatic). În regiunea capului mai prezintă trei ochi simpli numiţi oceli şi două antene mici, aristate, formate din câte trei segmente; pe cel de-al treilea segment antenal aflându-se arista. Aparatul bucal este adaptat pentru lins şi supt, Drosophila consumă hrană lichidă sau lichefiată, în prealabil, cu ajutorul secreţie glandelor salivare.
Ca toate Dipterele, musculiţa de oţet prezintă două aripi mari, mezotoracice, aripile de pe
metatorace fiind transformate în haltere (organe de echilibru). Pe fiecare segment toracic se află câte o pereche de picioare adaptate la mers.
Numărul segmentelor abdominale vizibile este diferit la cele două sexe – şapte la femelă şi cinci-şase la mascul. Acest număr mic de segmente la masculi este consecinţa fenomenului de telescopare a ultimelor segmente abdominale.
Adulţii de Drosophila melanogaster prezintă dimorfism sexual, manifestat atât prin diferenţe
dimensionale, cât şi morfologice. Femelele sunt mai mari decât masculii. Au abdomenul oval cu extremitatea posterioară puţin ascuţită. La masculi vârful abdomenului este rotunjit şi colorat în maron-negru (Fig.13).
Numărul de omatidii ce intră în alcătuirea ochiului complex este diferit le cele două sexe, şi anume: 780 la femelă, respectiv, 740 la mascul. Masculii prezintă aşa numitul „pieptene sexual” care serveşte la imobilizarea femelei în timpul acuplării. Acesta este format din zece perişori negri, rigizi, ce se găsesc pe primul articol tarsal al picioarelor anterioare (perechea I), mai precis în apropierea articulaţiei dintre tibie şi tars. Evident că diferenţele cele mai însemnate dintre mascul şi femelă ţin de organizarea internă, mai ales de anatomia sistemului reproducător.
Sexul larvelor se determină ţinând cont de poziţia şi mărimea gonadelor. Acestea sunt
localizate în partea postero-laterală a larvei, în corpii graşi. Testiculele sunt mult mai mari decât ovarele (de trei ori mai mari) şi au o formă ovală. Ele pot fi observate cu ochiul liber datorită transparenţei tegumentului larvei.
Ca toate dipterele, Drosophila melanogaster prezintă metamorfoză completă (este o insectă holometabolică), ciclul său vital parcurgând stadiile de ou, larvă, pupă şi adult. Durata ontogeniei variază în funcţie de temperatură, umiditate, compoziţia mediului nutritiv etc.
Stadiul de ou. Este cuprins între momentul depunerii oului de către femela adultă şi cel al apariţiei larvei corespunzătoare. În partea anterioară, dorsal, oul prezintă o pereche de filamente (apofize) cu rolul de a împiedica scufundarea sa în mediul lichid.
Stadiile larvare la Drosophila melanogaster sunt în număr de trei, fiind separate între ele
prin două năpârliri. Larva de stadiul I părăseşte oul prin micropil, este mică, foarte mobilă şi se hrăneşte cu
mare repeziciune. Este o larvă de tip apod (fără apendici), acefală (capsula cefalică este înfundată complet în torace). Aparatul bucal de tip primitiv (adaptat pentru rupt şi mestecat) prezintă mandibule puternice, ca nişte cârlige, cu ajutorul cărora larva pătrunde în materialul nutitiv. Acest prim stadiu larval durează aproximativ 24 ore. Pe măsură ce larva creşte, cuticula sa chitinoasă devine neâncăpătoare, astfel, că larva o părăseşte şi îşi sintetizează o nouă cuticulă (năpârleşte).
Larva de stadiul II apare în urma primei năpârliri. Aceasta este mai mare şi continuă să se hrănească timp de aproximativ 24 ore, după care suferă încă o năpârlire.
Larva de stadiul III apare după cea de-a doua năpârlire (după aproximativ şase zile de la
fertilizare). Aceasta pote atinge 4,5 – 5 mm lungime şi se dezvoltă pe parcursul a 48 ore. Stadiul de pupă are o durată variabilă, fiind profund influenţat de temperatură. La
Drosophila melanogaster pupa este de tip coarctat „pupă butoiaş”, o variantă a pupei adectice (cu
mandibule nesclerificate) de tip liber, caracterizată prin aceea că larva din stadiul III împupează în interiorul ultimei exuvii larvare. La început învelişul pupal chitinos este alb şi moale. Odată cu scurtarea corpului larvei el se întăreşte şi se închide la culoare, fiind numit şi puparium. Are formă ovală, măsoară 2,5 – 3 mm şi prezintă în partea anterioară două prelungiri ce corespund spiraculelor evaginate ale fostei larve.
Stadiul de imago. În urma transformărilor ce au loc în interiorul învelişului pupal, se
formează adultul care eclozează printr-o deschidere circulară din regiunea anterioară a pupariumului. La început insecta este foarte lungă, cu aripile nedesfăcute, lipite de corp şi are o culoare deschisă. Aceste caracteristici ale adulţilor nou eclozaţi constituie criteriile care îi deosebesc de insectele bătrâne.
Indivizii speciei Drosophila melanogaster sun frecvent întîlniţi în natură, pe fructe care au
început să fermenteze (mere, pere, prune, struguri etc.). Vara o întâlnim în vii, grădini, livezi sau în magazii, beciuri cu fructe şi legume (mai ales dacă acestea sunt alterate), în depozite, pe tescovina rămasă de la prelucrarea vinului şi, în general, în locuri insalubre. Această specie nu prezintă diapauză, iernând în magazii, depozite, beciuri etc.
În laborator, musculiţa de oţet poate fi cultivată pe medii artificiale care conţin zahăr şi drojdie – componente ce mijlocesc procesul de fermentaţie. Compoziţia mediului are influenţe asupra ciclulului vital, viabilităţii şi prolificităţii insectei.
Fig.13. Mascul şi femelă de Drosophila melanogaster de tip sălbatic (Wild type)
Temperatura, ca şi compoziţia mediului de cultură, influenţează desfăşurarea ciclului vital la Drosophila melanogaster. La temperaturi mai mari de +310C femela devine parţial sau chiar total
sterilă. În regiunile cu umiditate sporită, musculiţa de oţet poate tolera temperaturi mai ridicate ale aerului. Temperatura optimă pentru dezvoltarea indivizilor de Drosophila melanogaster este de
+24 - +250C. La această temperatură ciclul vital durează 10 zile (stadiului de ou îi revin 24 ore, celui de larvă – 4 zile şi pupei tot 4 zile). Astfel, într-un an se pot obţine 40 de generaţii. La temperaturi mai mici, ciclul de dezvolatre al insectei se lungeşte. Astfel, la +20oC, ciclul vital se prelungeşte până la 16 zile (larva – 8 zile, pupa – 6-7 zile), pe când la +100C durata ontogeniei ajunge la 75 de zile (larva 57 zile, pupa – 13-14 zile).
Umiditatea optimă necesară dezvoltării normale pentru Drosophila este de 75-85%. În unele cazuri acest parametru poate deveni o problemă serioasă. Un nivel al umidităţii sub 70% afectează profund fertilitatea şi viabilitatea insectelor.
Musculiţa de oţet prezintă o prolificitate ridicată. În cursul vieţii, o femelă depune câteva sute de ouă. In majoritatea cazurilor numărul ouălor depuse este de 200-300. Bridges precizează că ameliorarea condiţiilor de viaţă determină o creştere a numărului de ouă depuse, uneori acesta ajungând până la 2000. In medie o pereche de insecte dă naştere la aproximativ la 170 descendenţi.
Adulţii de Drosophila melanogaster, în condiţii de laborator, trăiesc aproximativ 3-4
saptamâni. Longevitatea depinde de condiţiile în care sunt crescute insectele şi anume: temperatura, umiditatea, compoziţia mediului nutritiv, densitatea populaţiei, prezenţa în mediu a paraziţilor etc.
În condiţii optime indivizii de Drosophila pot supravieţui chiar 22 de săptămâni (153 de zile).
Exista unele mutante care au o viabilitate mult mai mare decât a tipului sălbatic însă, de obicei, mutantele sunt mai puţin viabile. În concluzie, putem afirma că sunt argumente care recomandă această specie pentru studii de genetică şi anume:
a. Rapiditatea succesiunii generaţiilor (circa 10-15 zile pentru un ciclu ontogenetic complet, la 24oC).
b. Prolificitate crescută (circa 200 ouă/femelă). c. Pretenţii scăzute faţă de condiţiile de creştere (se dezvoltă bine în condiţii artificiale). d. Număr mic de cromosomi (2n=8) uşor identificabili. Astfel, perechea I este alcătuită din
cromosomi în formă de bastonaşe drepte la femelă şi unul în formă de bastonaş drept şi altul îndoit la un capăt, în cazul masculului. A se reţine acest aspect, ca argument pentru fundamentarea ipotezei plasării genelor pe cromosomi şi pentru considerarea respectivei perechi ca reprezentând cromosomii sexului (pentru femelă, sunt notaţi cu XX, iar pentru mascul, cu XY). Cromosomii din perechile II şi III sunt de forma literei "V". iar cei ai perechii IV sunt punctiformi. Dacă cromosomii sexului mai poartă şi numele de heterosomi, restul sunt denumiţi autosomi.
Principala caracteristică a drosofilei care, pe lângă calităţile amintite anterior, o recomandă drept un material extrem de adecvat pentru investigaţii genetice, o constituie mutabilitatea ei ridicată (rata relativ înaltă de apariţie a mutaţiilor, atât în mod spontan, cât şi sub impactul anumitor tratamente). Practic, pentru fiecare dintre caracterele morfo-anatomice s-au identificat şi descris mutaţii. Sunt stabilite şi caracterizate, până în prezent, peste 500 mutaţii, vizând culoarea ochilor, forma ochilor şi a aripilor, culoarea corpului, pilozitatea şi forma perişorilor etc. Pentru Morgan, mutaţiile au constituit un excelent mijloc de a studia legităţile transmiterii ereditare a caracterelor şi de a stabili mecanismele cromosomiale ale procesului.
În principiu, s-a păstrat metodologia mendeliană a notării genelor (factorii ereditari, în concepţia lui Mendel), dar s-a introdus o inovaţie. În loc să se folosească, în mod aleatoriu, literele alfabetului latin pentru desemnarea fiecărei gene, se foloseşte prima literă, primele litere sau un grup de litere din cuvântul englezesc ce denumeşte prima mutantă descoperită pentru caracterul respectiv. Cel mai utilizat exemplu este acela referitor la culoarea ochilor.
În mod normal, drosofila are ochii de culoare roşie. Prima mutantă identificată, pentru acest caracter a fost cea cu ochi albi. În consecinţă, ea a primit numele white, notarea ei convenţională fiind w. Evident, pentru forma cu ochi normali, dominantă în încrucişările cu mutantele, se putea recurge la desemnarea prin W. S-a preferat, însă, utilizarea simbolului w+, întrucât există şi cazuri
de mutante dominante, a căror desemnare se face prin literă mare. Dar, a apărut o nouă complicaţie. În cazul unor gene (vezi tabelul 4.2) există o serie alelică (polialelia). Referindu-ne tot
la culoarea ochilor, menţionăm mutantele: sepia, eosin, ivory, vermillon, apricot etc. Desemnarea lor ar putea fi făcută prin litere de la respectivele cuvinte (sp, e, i, v, a), dar, în acest caz, desemnarea tipului sălbatic ar avea mai multe simboluri (sp+, e+, i+, v+, a+ etc.) şi ar putea duce la confuzii. În consecinţă, s-a recurs la un alt sistem, în care la prima mutaţie se adaugă ind icativele celor ulterior descoperite (wsp, we, wi, wv, wa), situaţie în care, pentru desemnarea tipului sălbatic, se recurge constant la o singură notaţie - w+(Fig.14).
Cromosomii din perechea I
y, yellow (0,0) – mutantă ce prezintă corp de culoare galbenă intensă, perii fiind cafenii cu vârful galben;
pn, prune (0,8) – mutantă cu ochi de culoarea prunei;
w, white (1-1,5) – mutantă cu ochi de culoare albă, ocelii fiind incolori, ca şi tuburile lui Malpighi. A fost prima mutantă descoperită (în 1910, de Lilian Morgan). Prezintă numeroase alele;
wa, apricot (1-1,5) – prezintă ochi de culoare trandafirie-gălbuie la masculi şi ceva mai
galbeni la femele. Combinaţia wa/w
a;bw/bw are ochii mai deschişi la culoare decât
mutanta apricot (wa/w
a;bw
+/bw
+);
N, Notch (3,1) – mutantă cu aripile zimţate;
cx, curlex (13,6) – mutantă cu aripile îndoite în sus;
ct, cut (20,0) – mutantă cu aripile reduse (tăiate);
ras, raspberry (32,8) – mutantă cu ochi de culoarea zmeurei;
v, vermilion (33,0) – mutantă ce prezintă ochi de culoare roşie foarte vie, ocelii fiind incolori. Combinaţia v/v;bw/bw prezintă ochi albi. Viabilitatea este satisfăcătoare;
m, miniature (36,1) – aripile sunt mici, depăşind cu puţin abdomenul;
f, forked (56,6) – prezintă peri scurţi şi îndoiţi, bifurcaţi la vârf;
B, Bar (57,0) – mutantă dominantă, ce prezintă ochi reduşi la un segment;
Bx, Beadex (59,4) – mutantă cu aripi zimţate;
car, carnation (62,5) mutantă cu ochi de culoare rubinie-închisă.
Cromosomii din perechea II-a
al, aristaless (0,0) – mutantă fără ariste;
S, Star (1,3) – mutantă cu ochi în formă de stea;
Cy, Curly (8,5) – prezintă aripile îndoite în sus. În stare homozigotă (Cy/Cy), această genă este letală;
dp, dumpy (13,0) – mutantă cu aripile tăiate şi reduse la aproximativ 32 din lungimea
aripilor de latipulul sălbatic;
b, black (48,5) – prezintă corp de culoare neagră;
pr, purple (54,5) – mutantă cu ochi purpurii;
ap, apterous (55,4) – nu prezintă aripi;
cn, cinnabar (57,5) – mutantă cu ochi de culoare roşie aprinsă, ce se închide odată cu vârsta;
vg, vestigial (67) – prezintă aripile sub forma unor rudimente (vestigii), de unde a primit şi denumirea;
sf, safranin (71,5) – prezintă ochi de culoarea ciocolatei;
L, Lobe (72,0) – prezintă ochi reduşi ca mărime. În stare homozigotă (L/L) determină o reducere accentuată a ochilor şi scăderea viabilităţii, poate fi letală;
c, curved (75,5) – mutantă cu aripi subţiri, depărtate şi răsucite;
bw, brown (104,5) – culoarea ochilor este cafenie deschisă la naştere şi se închide după mai multe zile. Este afectată şi culoarea organelor interne. Vasele şi testiculele sunt incolore, iar tuburile lui Malpighi sunt mai deschise la culoare decât la tipul sălbatic;
Cromosomii din perechea III-a
dv, divergent (20,0) – mutantă cu aripile îndepărtate;
se, sepia (26,0) – prezintă ochi de culoare roşie-cafenie, transparenţi la naştere, închizându-se, cu timpul, până la sepia (pot deveni chiar negri);
rs, rose (35,0) – mutantă cu ochi de culoare roz;
D, Dichaete (40,4) – prezintă câte doi perişori împreună;
p, pink (48,0) – mutantă cu ochi de culoare rubinie;
ma, maroon (49,7) – prezintă ochi de culoare castanie;
dw, dwarf (50,0) – mutantă pitică;
ss, scinelles (58,0) – mutantă caracterizată prin absenţa perişorilor;
bx, bithorax (58,5) – mutantă cu mezotoracele dublat;
e, ebony (70,7) – prezintă corp de culoare neagră strălucitoare, mult mai închisă decât la Wild type. Viabilitatea acestei mutante este 80% din cea a tipului sălbatic;
det, detachet (72,5) – mutantă cu ochi sticloşi;
ro, rough (91,1) – prezintă ochi micşoraţi, cu suprafaţă neregulată.
Cromosomii din perechea IV-a
M4, Minute 4 (0,0) – mutantă cu perişori subţiri;
bt, bent (0,0) – mutantă cu aripile îndoite în jos;
sv, shaven (0,1) – prezintă peri abdominali rari, mai puţini decât la tipul sălbatic;
ey, eyeless (0,2) – mutantă cu ochi reduşi la jumătate sau la un sfert din mărimea celor de la tipul salbatic. Uneori ochii lipsesc complet;
Cat, Cataract (6,7) – prezintă ochi cu suprafaţa neregulată;
spa, sparkling (93,0) – mutantă cu ochi proeminenţi, ce au suprafaţa neregulată.
Fig. 14. Mutante de Drosophila melanogaster
4.2. Argumente pentru plasarea genelor (factorilor ereditari) pe cromosomi
Argumentul, imbatabil, pentru considerarea cromosomului drept suport material al factorilor ereditari (pentru plasarea genelor pe cromosomi) l-au furnizat experimentele efectuate cu drosofila. Să prezentăm doar unul dintre ele.
În experimentele efectuate de Mendel, utilizând mazărea cu bobul galben şi cea cu bobul verde, segregarea în F2 era aceeaşi, indiferent de culoarea părintelui ales ca mascul şi a celui ales ca femelă. Nu acelaşi lucru se constată, însă, când se fac încrucişări reciproce între indivizii w+ şi w de drosofila. Situaţia a condus la concluzia că gena responsabilă de manifestarea respectivului
caracter este cantonată pe unul dintre cromosomii sexuali. Iată care sunt rezultatele în cele două tipuri de încrucişare:
♀50% cu ochi roşii;50% cu ochi albi ♂ 50% cu ochi roşii;50% cu ochi albi
Întrucât atât masculii cât şi femelele au trei perechi identice de autosomi, diferenţele constând în prezenţa heterosomilor pereche (XX) la femele sau nepereche (XY) la masculi, rezultă că respectiva genă este plasată pe unul dintre cromosomii sexului şi anume pe cromosomul X,
deoarece femelele heterozigote se comportă (fenotipic) asemenea celor homozigote dominante, în timp ce masculii (fiind hemizigoţi) se manifestă recesiv sau dominant, în funcţie de tipul cromosomului X pe care l-au primit (cu gena sălbatică sau cu alela mutantă plasată pe el). Fenomenul, cunoscut şi sub numele de sex linkage, se constituie în cauză pentru abaterea de la
tipul mendelian de segregare. Un alt exemplu, care confirmă ipoteza plasării genelor pe cromosomi, îl constituie
asocierea dintre lipsa unui cromosom din perechea IV şi lipsa ochilor, la aceeaşi insectă. Prin urmare, pe un cromosom, într-un anumit loc, numit locus, se află plasată o genă. Pe
cromosomul omolog, în aceeaşi poziţie, adică în acelaşi locus, se află plasată aceeaşi genă sau una dintre alelele ei.
Întrucât, în orice celulă somatică avem cel puţin două seturi cromosomiale, unul provenind pe linie maternă şi celălalt pe linie paternă, se poate considera că un individ, diploid, se poate afla în trei ipostaze şi anume: AA (homozigot dominant), Aa (heterozigot) şi aa (homozigot recesiv). Prin urmare, altfel spus, în cadrul diploidului genele se pot afla în doză simplă (gena A şi alela a), în doză dublă (AA), sau lipsesc fiind prezente doar alelele, în doză dublă (aa).
În această ordine de idei, se cuvine reamintit faptul că, prin mutaţie, una şi aceeaşi genă sau alela (alele) ei, poate suferi o succesiune de mutaţii, şirul de alele fiind mult mai mare. Fenomenul, cunoscut sub numele de polialelism, contribuie la îmbogăţirea substanţială a variabilităţii intrapopulaţionale, în sensul că genofondul se îmbogăţeşte. Evident, starea probabilă a unui individ poate îmbrăca doar una dintre ipostazele menţionate anterior, cu deosebirea că numai homozigoţii dominanţi vor fi de un singur tip (AA), în timp ce heterozigoţii şi homozogoţii recesivi pot fi de mai multe tipuri: Aa, Aa1...Aan, aa1...aan etc. (pentru heterozigoţi) şi aa, a1a1...anan (pentru
homozigoţii recesivi). Prin urmare, alelele sunt mutante ale genei de origine, plasate în cromosomii omologi, în
acelaşi locus, la indivizi diferiţi şi au rol în determinismul aceluiaşi caracter. În cazul experienţelor lui Mendel, de pildă, alele erau doar A şi a, responsabile de
determinismul culorii bobului, sau B şi b, responsabile de determinismul formei bobului. Între A şi B nu se poate vorbi despre raport de alelism.
Într-o tentativă de definire a genei, nu putem omite calităţile ei de unitate ereditară, mutaţională şi recombinaţională. În timp, noţiunii de genă i s-au atribuit mai multe sensuri. Dintre acestea, le amintim pe următoarele: o genă este responsabilă de determinismul unui caracter; o genă este responsabilă de determinismul sintezei unei enzime, o genă este responsabilă de determinismul sintezei unei proteine; o genă este responsabilă de determinismul sintezei unei biomolecule. Întrucât cercetările în derulare au evidenţiat determinismul poligenic al unor caractere, apoi faptul că o moleculă enzimatică este alcătuită dintr-o proteină şi o grupare prostetică şi implică activitatea a mai mult decât o genă, precum şi faptul că prin activitate genică se sintetizează şi molecule nonproteice (acizii ribonucleici etc.) singura accepţiune valabilă pentru noţiunea de genă a rămas cea din urmă, adică o genă reprezintă secvenţa de codoni responsabilă de determinismul sintezei unei biomolecule.
În sens larg însă, considerăm că atribuţia esenţială a unei gene rezidă în determinarea unui caracter. Dar, aşa cum precizam anterior, structura unei gene (calitatea informaţiei pe care o deţine) se poate schimba, prin mutaţie. În consecinţă, o genă poate avea una sau mai multe alele. Evident, gena şi alelele sale ocupă acelaşi locus (aceeaşi poziţie) pe un anumit cromosom (sau pe omologul său). Prin mutaţie gena îşi schimbă nu numai calitatea informaţională, ci şi "calitatea activităţii", în sensul că, deşi determină acelaşi caracter (şi nu altul), amplitudinea fenotipizării ei va fi alta. Prin intermediul fecundării, într-un zigot (viitor organism) se pot întâlni şi forma o pereche, conform teoriei probabilităţilor, atât genele dominante, cât şi alelele sau o genă cu una din alele, întrucât un genom provine pe linie paternă şi celălalt, pe linie maternă. Este vorba, evident, de perechea de gene sau alele situate în acelaşi locus. Cu această ocazie, mai precizăm încă un aspect. În concepţia noastră, gena este unică, adică cea care s-a constituit iniţial, cunoscută şi sub numele de genă sălbatică. Tot ce a apărut ulterior, prin mutaţie, reprezintă familia de alele ale genei sălbatice. În consecinţă, exprimarea corectă este: o genă şi alelele ei (în limba greacă, allelon = altul). Pe de altă parte, meioza permite asortarea întâmplătoare (dansul aleatoriu al cromosomilor; de la latinescul aleatoriu = întâmplător) a cromosomilor şi, deci, a genelor, în
gameţi. În consecinţă, aşa cum a rezultat din experimentele lui Mendel, indivizii unei descendenţe hibride pot etala o multitudine de recombinări ale caracterelor prezente la părinţi.
4.3. Plasarea liniară a genelor pe cromosomi
În paginile anterioare am demonstrat, fără dubii, că genele sunt plasate pe cromosomi. Dar, un calcul extrem de simplu ne arată că numărul de cromosomi este mult mai mic, faţă de numărul caracterelor etalate de un individ. La Drosophila melanogaster, de pildă, s-au identificat mai multe
sute de caractere şi numai 8 cromosomi. Ţinând cont de omologia cromosomilor, în fapt există patru perechi şi, având în vedere raportul de alelism al genelor, putem vorbi doar de 4 grupe de linkage ca entităţi diferite. Deci, ca o primă concluzie, se impune constatarea că genele responsabile pentru cele câteva sute de caractere sunt cantonate în patru cromosomi. Deci, într-un cromosom există mai multe gene. Dar cum sunt ele plasate, în mod concret, într-un cromosom? Şi ce dovezi avem, totuşi, că mai multe gene sunt plasate în acelaşi cromosom?
Argumentele pentru plasarea mai multor gene pe acelaşi cromosom aparţin tot şcolii lui T.H. Morgan şi au fost susţinute de experimente extrem de simple şi clare, totodată.
S-a efectuat o experienţă de hibridare (tip dihibridare), între indivizi de D. melanogaster ce
difereau între ei prin culoarea ochilor şi forma aripilor. Femela era fenotipic normală (pr+pr+vg+vg+), iar masculul dublu mutant - avea ochii purpurii (prpr) şi aripile vestigiale (vgvg). În F1 s-au obţinut doar indivizi fenotipic normali. Prin încrucişarea lor, în F2, în loc să se obţină segregarea de tip mendelian, de 9:3:3:1, s-au obţinut doar două categorii de fenotipuri - 75% de tip parental dominant şi 25% de tip parental recesiv. Deci, un raport tipic monohibridării (adică 3 : 1). Prin urmare, se impune o primă concluzie. Cele două caractere nu segregă independent. De
aici supoziţia că ele sunt plasate (genele ce le determină) pe acelaşi cromosom. Adică, pentru individul normal vom avea pr+ vg+, şi pr wg pentru cel mutant.
pr+ vg+ pr wg Pentru a elimina confuziile, precizăm că cele două gene sunt cantonate pe cromosomii
perechii a II-a. Indicativul "pr" redă mutanta purple (purpuriu, în engleză). Deci, nu este vorba de cromosomii sexului şi, de aceea, nu am notat masculul mutant prin pr vg. Încrucişarea se derulează după următoarea schemă:
Prin urmare cele două gene sunt localizate pe acelaşi cromosom şi se transmit împreună.
Fenomenul este cunoscut, în genetică, sub numele de linkage (înlănţuire, legare). Este, în fond,
nu numai o abatere, ci o completare a legii segregării independente a caracterelor, demonstrată de Mendel.
4.4. Transmiterea înlănţuită a tuturor genelor plasate pe un cromosom (linkage)
Autenticitatea linkage-ului a fost confirmată şi prin retroîncrucişarea de tipul Test-cross (FT). Adică, un individ din F1 (un mascul) se încrucişează cu unul dublu mutant (o femelă). Se obţin două clase fenotipice şi anume: 50% indivizi normali şi 50% indivizi dublu mutanţi. Precizăm că este obligatorie respectarea sexelor indicate, deoarece masculul nu prezintă fenomenul de crossing-over şi, în consecinţă, rezultatele experimentului sunt constante.
P(F1) ♂ pr
+ vg
+
pr vg
x ♀ pr vg
pr vg
G pr+ vg
+, pr vg x pr vg
FT pr
+ vg
+
pr vg
pr vg pr vg
Heterozigoţi cu fenotip normal
(dominant) 50% Homozigoţi cu fenotip recesiv(dublu
mutant) 50%
Deci, atât genotipic (Sg), cât şi fenotipic (Sf), segregarea este de 1:1. În consecinţă, cele 500 de gene identificate la Drosophila melanogaster au fost repartizate
în patru grupe de linkage, corespunzătoare celor patru perechi de cromosomi omologi. Se cuvine menţionat faptul că mărimea unei grupe de linkage sau, altfel spus, numărul genelor plasate într-o grupă este în concordanţă cu mărimea cromosomului. Astfel, grupa a IV-a de linkage, fiind plasată pe cromosomii perechii a IV-a (punctiformi), conţine doar 12 gene.
Ca o concluzie, generală, menţionăm că la fiecare specie, numărul grupelor de linkage este identic cu numărul cromosomilor unui set haploid (genom). Se ridică o singură problemă doar,
aceea a autopoliploizilor. In mod normal, într-un autopoliploid, numărul grupelor autentice de linkage este acelaşi cu numărul existent în diploidul de origine. La alopoliploizi numărul grupelor de linkage reprezintă suma grupelor diploizilor ce au contribuit la hibridare. In aceeaşi idee, precizăm că la aneuploizi avem grupe de linkage supranumerare sau subnumerare, comparativ cu diploizi i de origine.
In conformitate cu teoria linkage-ului, atunci când două gene segregă împreună, înseamnă că fac parte din aceeasi grupă de linkage, iar când segregă independent (după modelul mendelian) înseamnă că fac parte din grupe diferite de linkage - adică sunt plasate pe cromosomi diferiţi.
4.4.1. Sex-linkage (transmiterea înlănţuită a genelor plasate pe heterosomi)
Aşa cum am precizat deja, specia Drosophila melanogaster are patru grupe de linkage,
adică are patru perechi de cromosomi. Mai exact drosofila are trei perechi de autosomi şi doi heterosomi – identici la femele (notaţi XX) şi diferiţi la masculi (notaţi XY). Prin experimente
proiectate şi efectuate cu deosebită grijă, T.H.Morgan şi grupul său de cercetători au reuşit să demonstreze că una dintre genele responsabile pentru determinismul culorii ochilor este plasată pe heterosomul X (vezi harta genetică), rezultatele încrucişărilor între indivizi normali şi indivizi mutanţi depinzând, întotdeauna, de sexul mutantului. Aşa cum reiese din imaginile redate anterior, rezultatele încrucişărilor sunt dependente de genotipurile masculilor şi femelelor cu care s-a efectuat experimentul.
Figura 4.3. Moştenirea culorii ochilor la Drosophila melanogaster, în descendenţa unei femele normale şi a unui mascul cu ochii albi (După Snustad & co., 1997)
Prin urmare, s-a reuşit să se demonstreze că gena responsabilă de determinismul culorii
ochilor este cantonată pe heterosomul X. În fapt totul a început după ce a fost sesizată prezenţa
unui mascul cu ochii albi. Când respectivul mascul a fost încrucişat cu o femelă normală (de tip sălbatic), toţi descendenţii au avut ochii de culoare roşie, demonstrând recesivitatea culorii albe şi dominanţa celei roşii. Prin încrucişarea descendenţilor între ei s-a constatat că situaţia se schimbă şi se complică – femelele apăreau numai cu ochii roşii în timp ce masculii apăreau atât cu ochi albi (50%) cât şi cu ochi roşii (50%). Este clar, deci, că respectivul caracter (mai exact spus gena care-l determină) este cantonat pe cromosomul X. Altfel spus transmiterea respectivei gene este sex-linkată. Gena pentru culoarea ochilor este plasată pe cromosomul X şi nu pe Y şi, prin mutaţie, apar alele. Tipul sălbatic (normal) este desemnat prin w+, iar alela pentru culoarea albă prin w.
Recurgând la o simbolistică generală şi unificatoare în acelaşi timp, în care notăm genele cu litere latine şi, presupunând că vom avea un grup de linkage din cinci gene, dinamica genotipurilor şi fenotipurilor în descendenţa hibridării va arăta astfel:
P ♀ ABCDE ABCDE
x ♂ abcde abcde
G ABCDE x abcde
F1 ABCDE abcde
x ABCDE abcde
G ABCDE, abcde x ABCDE, abcde
F2 ABCDE ABCDE
ABCDE abcde
ABCDE abcde
abcde abcde
Sg 1 2 1
Sf 3 1
Genele care fac parte din aceeaşi grupă de linkage, sunt gene linkate.
Cromosomii reprezentând substratul (suportul) material al genelor, rezultă că în meioză asortarea întâmplătoare a cromosomilor în gameţi este, în fapt, asortarea întâmplătoare a grupelor de linkage. Altfel spus, o genă şi chiar o grupă de linkage, au comportamente identice cromosomilor.
Figura 4.4 Transmiterea culorii ochilor, la D. melanogaster, prin hibridarea dintre o femelă heterozigotă şi un mascul mutant (sus) şi o femelă mutantă şi un mascul normal (jos) (După Snustad & co., 1997)
În cazul sex-linkage-ului, are importanţă sexul indivizilor pe care-i folosim în încrucişare. Evident, când genele sunt localizate pe autosomi, sexul indivizilor utilizaţi pentru încrucişare nu ar trebui să aibă importanţă. Dar, aşa cum precizam mai sus, la Drosophila melanogaster, între autosomii masculului nu are loc fenomenul de crossing-over. Prin urmare, când se aşteaptă o constanţă a rezultatelor, este bine să se ţină cont de acest lucru.
4.5. Crossing-over (schimbul reciproc de gene)
Aşa cum precizam în rândurile anterioare, nu întotdeauna genele aparţinând aceluiaşi grup de linkage, se transmit împreună. S-au constatat nenumărate excepţii. În tentativa de a da explicaţii adecvate abaterilor de la regulă, Morgan şi echipa sa au constatat că între cromosomii omologi poate avea loc un schimb reciproc de gene. Fenomenul a primit numele de crossing-over.
4.5.1. Crossing-over între cromosomii sexului
Prin încrucişarea unei femele de Drosophila melanogaster, dublu mutantă (cu două mutaţii
recesive, ale genelor localizate pe cromosomul X, determinând culoarea galbenă a corpului şi culoarea albă a ochilor), cu un mascul normal, în F1 s-au obţinut doar indivizi de două tipuri - deci, o segregare de 1:1. Femelele erau cu fenotip normal, dar heterozigote şi masculii cu fenotipizare mutantă, dar hemizigoţi genotipic. Fenomenul a primit denumirea de ereditate în cruciş (criss-cross), deoarece fenotipul mamei se manifestă la fii, iar cel al tatălui la fiice.
Prin încrucişarea indivizilor femeli şi masculi din F1, s-a constatat apariţia, în F2, a patru clase de indivizi - două clase conforme rezultatelor determinate de linkage şi alte două clase cu recombinare de caractere. Proporţia era de 49,25% pentru fiecare dintre primele două clase şi de 0,75% pentru fiecare din clasele cu recombinare.
Precizăm că w reprezintă mutanta pentru culoarea ochilor (ochi albi) şi y reprezintă mutanta pentru corp galben (de la cuvântul englezesc yellow), w+ şi, respectiv, y+ reprezentând
genele normale. Deci, plecându-se de la femele dublu mutante şi masculi normali, în F1 s-a obţinut o
segregare fenotipică (Sf) de 1:1 şi una genotipică (Sg) tot de 1:1, femelele fiind heterozigote, cu
fenotip normal (dominant) şi masculii hemizigoţi, cu fenotip recesiv (mutant). Schematic, încrucişarea este redată astfel:
Părinţi X
yw
Xyw x
Xy+w+
Y
Gameţi Xyw
x Xyw
, Y
F1 X
yw
Xy+w+
X
yw
Y
Ga-meţi
ne-recombinaţi
Xyw
, Xy+w+
x Xyw
, Y
recombinaţi Xy+w
, Xyw+
F2 X
yw
Xyw
Xyw
Y X
y+w+
Xyw
Xy’w+
Y X
y+w
Xyw
Xy+w
Y
Xyw+
X
yw+ X
yw+
Y
49,75% 49,75% 0,75% 0,75%
Teoretic, gameţii indivizilor de F1 trebuiau să fie doar de două tipuri: la femele Xyw şi Xy+w+,
iar la masculi Xyw şi Y. Dar rezultatele încrucişării indivizilor din F1, dând un cu totul alt raport de
segregare în F2 şi, mai ales, dând o categorie de indivizi cu recombinări de caractere, s-a ajuns la concluzia că a avut loc crossing-overul între cromosomii omologi şi apariţia a două noi tipuri de gameţi în cazul femelelor: Xy+w şi Xyw+. Prezenţa acestor gameţi explică apariţia indivizilor cu
fenotipuri recombinate, în F2. Aceştia sunt atât masculi, cât şi femele, cu corp gri şi ochi albi sau cu ochi roşii şi corp galben.
Deci, între genele y+ şi w+ şi alelele lor, situate pe cromosomii X omologi, au avut loc schimburi, rezultând noi grupe de linkage. Adică, din y+w+ şi yw, au rezultat y+w şi yw+, alături de
tipurile parentale care s-au menţinut.
4.5.2. Crossing-over între autosomi
Un alt experiment, efectuat de Morgan şi colaboratorii săi, a demonstrat existenţa crossing-overului şi între autosomi.
Încrucişând o femelă de Drosophila melanogaster cu corpul negru (b - de la black) şi aripi normale (vg+) cu un mascul cu corpul gri (b+) şi aripi vestigiale (vg), în F1 s-au obţinut indivizi heterozigoţi b+bvg+vg, cu fenotipul normal.
Încrucişându-se indivizii din F1 cu indivizi dublu mutanţi (încrucişare de analiză genetică), s-a constatat că în FT, fenotipul indivizilor diferă în funcţie de sexul lor.
Astfel, dacă se efectuează o retroîncrucişare între un mascul din F1, heterozigot (b+bvg+vg) cu o femelă mutantă (bbvgvg), în FT se obţine segregarea de 1:1, ca în experimentele pentru
demonstrarea fenomenului linkage (adică reapar indivizii cu fenotip parental - corp negru şi aripi normale şi corp gri cu aripi vestigiale). Deci, linkage-ul este total.
P b vg
+
b vg+
x b
+ vg
b+ vg
G b vg+ x b
+ vg
F1 b vg
+
b+ vg
x b vg b vg
G b vg+, b
+ vg x b vg
FT b vg
+
b vg
b+ vg
b vg
Sf 1 1
Sg 1(bbvg+vg) 1(b
+bvgvg)
Precizam anterior că, în acest caz, este vorba de autosomi - mai concret, de perechea a II-
a de omologi. Atunci, cum se explică diferenţa de comportament dintre masculi şi femele? S-a ajuns la concluzia că, în cazul masculilor de Drosophila melanogaster, cromosomii din perechea a II-a nu prezintă fenomenul de crossing-over. Lipsa crossing-overului este valabilă şi pentru cromosomii sexului (la mascul), în sensul că între cromosomii X şi Y nu se cunoaşte existenţa acestui fenomen.
În încrucişarea reciprocă (retroîncrucişare) dintre o femelă hibridă de F1 (b+bvg+vg) şi un
mascul dublu mutant (bbvgvg), în FT, pe lângă tipurile parentale (corp negru şi aripi normale; corp
gri şi aripi vestigiale), mai apar încă două clase fenotipice (care, în mod normal, nu trebuiau să apară: corp gri şi aripi normale; corp negru şi aripi vestigiale). Într-o populaţie suficient de mare, proporţia indivizilor este de 41,5% pentru fiecare dintre tipurile parentale şi câte 8,5% pentru fiecare dintre tipurile recombinante. Deci, linkage-ul genelor b+ şi vg+ se dereglează şi respectivele
gene "segregă", datorită schimbului reciproc de material cromatic între cromosomii omologi. Rezultă că femelele din F1 pot forma patru tipuri de gameţi - două normale şi două
recombinate, primii fiind numiţi gameţi crossing-overi, ceilalţi fiind gameţi necrossing-overi.
P b vg
+
b vg+
x b
+ vg
b+ vg
G b vg+ x b
+ vg
F1 b vg
+
b+ vg
x b vg b vg
GF1 b vg
+, b
+ vg
şi b+ vg
+, b vg
x b vg
FT b vg
+
b vg b
+ vg
b vg b
+ vg
+
b vg b vg b vg
segregare
41,5% 41,5% 8,5% 8,5%
83,0 % indivizi necrossing-overi
17,0% indivizi crossing-overi
4.6. Crossing-over la plante
4.6.1. Crossing-over simplu la plante
În cel de-al doilea deceniu al secolului nostru, C. Hutchinson, lucrând pe porumb, a pus în evidenţă prezenţa crossing-overului şi în lumea plantelor.
C. Hutchinson a utilizat două linii homozigote de Zea mays - una cu boabe conţinând aleuronă colorată şi endosperm neted (c+c+sh+sh+), iar cealaltă cu aleuronă incoloră şi endosperm zbârcit (ccshsh). Hibridarea între cele două linii a permis obţinerea, în F1, heterozigoţilor c+csh+sh, cu expresie fenotipică normală. Prin retroîncrucişarea unui astfel de hibrid, din F1, cu un părinte dublu mutant, în FT nu s-a obţinut o segregare de 1:1, cum ar fi fost normal în linkage, ci s-au obţinut patru clase fenotipice - două asemănătoare cu părinţii (adică fără crossing-over) şi două recombinante (cu crossing-over), care nu erau aşteptate în mod normal.
P c
+ sh
+
c+ sh
+
x c sh c sh
G c+ sh
+ x c sh
F1 c
+ sh
+
c sh x
c sh c sh
G
c+ sh
+, c sh,
gameţi necrossing-
over x c sh
c+ sh, c sh
+
gameţi crossing-over
FT c
+ sh
+
c sh c sh c sh
c+ sh
c sh c sh
+
c sh
48,2% 48,2% 1,8% 1,8%
Indivizi necrossing-overi Indivizi crossing-overi
În acest caz ne aflăm în faţa unui proces de crossing-over simplu, deoarece între
cromosomii omologi a avut loc un singur schimb, între o singură pereche de gene (gena sălbatică şi alela sa mutantă).
4.6.2. Crossing-over dublu la plante
Dacă în crossing-overul simplu a avut loc o singură ruptură cromatidică, s-au constatat şi cazuri în care se produc recombinări între mai multe gene şi alelele lor - schimburile fiind duble, triple sau multiple.
M.M. Rhoades a descoperit un crossing-over dublu, tot la Zea mays. Încrucişând plante cu fenotip normal verde (v+), având nervura centrală a frunzei de culoare verde (bw+), boabele cu grăuncioare de aleuronă incoloră (pr+) şi plante mutante pentru cele trei caractere, adică având fenotipul verde deschis (v, de la virescent), nervura centrală de culoare brună (bw, de la brown) şi aleurona purpurie (pr), în F1 a obţinut plante heterozigote cu fenotipul normal. Precizăm că cele
trei gene sunt localizate pe cromosomii din perechea a V-a. Hibrizii din F1, retroîncrucişaţi cu indivizi triplu mutanţi, nu s-au comportat în FT conform
legilor mendeliene ale segregării, adică nu au dat 50% indivizi cu fenotip normal şi 50% cu fenotip recesiv (mutant). Rezultatul a fost neaşteptat, obţinându-se opt tipuri de indivizi: două tipuri parentale şi şase tipuri recombinate.
Facem precizarea că procentele sunt întrucâtva ajustate, pentru a rezulta 100%, cele
obţinute în mod concret de Rhoades fiind 42,11%; 14,52%; 35,62% şi 7,75%, reprezentând cîte 467, 161, 395 şi 86 de plante din totalul celor ce au supravieţuit în experiment.
Ca o concluzie generală, menţionăm că la fiecare specie, numărul grupelor de linkage este identic cu numărul cromosomilor unui set haploid (genom). Se ridică o singură problemă doar - aceea a autopoliploizilor. In mod normal, într-un autopoliploid, numărul grupelor de linkage autentic, diferite între ele, ar trebui să fie acelaşi cu numărul existent în diploidul de origine. In aceeaşi idee, precizăm că la aneuploizii hipoploizi (la nulisomici) grupele de linkage sunt subnumerare.
In conformitate cu teoria linkage-ului, atunci când două gene segregă împreună, înseamnă că fac parte din aceeasi grupă de linkage, iar când segregă independent (după modelul mendelian) înseamnă că fac parte din grupe diferite de linkage - adică sunt plasate pe cromosomi diferiţi.
La Drosophilla melanogaster a fost posibilă şi localizarea concretă a fiecărei gene într-un
anumit cromosom. Evident, au fost necesare nenumărate experimente de hibridare, în care se luau, succesiv, tot alte perechi de caractere (gene) şi se nota cu meticulozitate comportamentul lor (dacă segregau independent sau nu).
Recurgând la o simbolistică generală şi unificatoare în acelaşi timp, în care notăm genele cu litere latine şi, presupunând că vom avea un grup de linkage din cinci gene, dinamica genotipurilor şi fenotipurilor în descendenţa hibridării va arăta astfel:
P ABCDE ABCDE
x abcde abcde
G ABCDE x abcde
F1 ABCDE abcde
x ABCDE abcde
G ABCDE, abcde x ABCDE, abcde
F2 ABCDE ABCDE
ABCDE abcde
ABCDE abcde
abcde abcde
Sg 1 2 1
Sf 3 1
Genele care fac parte din aceeaşi grupă de linkage, sunt gene linkate. Cromosomii, reprezentând substratul (suportul) material al genelor, rezultă că în meioză
asortarea întâmplătoare a cromosomilor în gameţi este, în fapt, asortarea întâmplătoare a grupelor de linkage. Altfel spus, o genă şi chiar o grupă de linkage au comportamente identice cu ale cromosomilor.
În cazul sex-linkage-ului, are importanţă sexul indivizilor pe care-i folosim în încrucişare. Evident, când genele sunt localizate pe autosomi, sexul indivizilor utilizaţi pentru încrucişare nu ar trebui să aibă importanţă. Dar, aşa cum precizam mai sus, la Drosophila melanogaster, între
autosomii masculului nu are loc fenomenul de crossing-over. Prin urmare, când se aşteaptă o constanţă a rezultatelor, este bine să se ţină cont de acest lucru.
Se cunosc şi cazuri de înlănţuire falsă, supranumită şi înlănţuire intercro-mosomială. Aceasta apare datorită dereglării ce intervine în procesul combinării libere a cromosomilor neomologi în meioză. Fenomenul se întâlneşte la încrucişarea între linii de şoareci de laborator,
între tulpini de Saccharomyces etc. Fenomenul este explicat prin tendinţa cromosomilor neomologi
de a se asorta neîntâmplător în procesul diviziunii meiotice. Transmiterea linkată a genelor cantonate pe cromosomi neomologi se constată, de asemenea, în cazul hibridărilor interspecifice, mai ales atunci când combinaţiile cromosomilor parentali sunt favorizate fiziologic (sunt fiziologic compatibile).
Uneori este posibil ca toţi cromosomii unui genom (set haploid) să migreze către un po l. Apreciem că aceste fapte, de observaţie şi experiment, se constituie în argumente pentru susţinerea ipotezei organizării supracromosomiale (pe care am prezentat-o la capitolul dedicat studiului cromosomilor).
4.7. Mecanismele crossing-overului
Pentru explicarea modalităţilor concrete prin care se realizează crossing-overul s-au emis mai multe ipoteze, care pot fi grupate în două mari categorii, şi anume:
I. Cele ce consideră că fenomenul se datorează ruperii cromatidelor şi schimbului reciproc de segmente (breakage theory) (Fig.14).
II. Cele ce consideră drept cauză a fenomenului replicaţia selectivă a cromatidelor în timpul dublării materialului cromosomial (copy-choise theory) (Fig.15).
Schematic, situaţia se redă astfel:
I
II Din cadrul primului grup de ipoteze, cea mai cunoscută este teoria chiasmatipiei (Fig.16).
Ea a fost elaborată de către F.A. Jannsen (1909, 1924), reluată apoi şi dezvoltată de C.D. Darlington şi K. Mathei (1932), fiind ulterior verificată de către J.H. Taylor (1957, 1958, 1963) prin metoda izotopilor radioactivi.
Conform acestei ipoteze, în profaza meiozei, între leptoten şi pachiten, între cromosomii omologi (unul de provenienţă paternă şi celălalt, de provenienţă maternă) se produc sinapse. Între pachiten şi diploten, fiecare cromosom apare format din două cromatide. Între cromatidele unei perechi de omologi se menţin puncte de contact, numite chiasme. În ipoteza chiasmatipiei,
anterior diplotenului, se produce un schimb de segmente cromatidice (egale) - fenomen cunoscut sub numele de chiasmata. Prin urmare, doi gameţi rămân nerecombinaţi, iar ceilalţi doi vor avea
cromosomii recombinaţi. O chiasmă nu poate afecta decât două dintre cele patru cromatide ale unei perechi de
omologi. În principiu, deci, crossing-overul între doi cromosomi omologi nu poate depăşi 50%. Fără îndoială, mai ales între cromosomii lungi, se pot forma mai multe chiasme, explicându-se astfel, crossing-overul dublu, triplu, multiplu.
În explicaţia crossing-overului, pe baza acestor ipoteze, intervin şi unele precizări. Una dintre ele priveşte repartiţia chiasmelor. Astfel, prima chiasmă apare la o anumită distanţă de centromer, denumită distanţă diferenţială, care este mai mică la cromosomii mai scurţi. Distanţele dintre chiasme, pe aceeaşi cromatidă, se numesc distanţe de interferenţă şi reprezintă caracter de specie.
Fig. 14 Schimb reciproc de segmente prin ruperea cromatidelor
Fig. 15 Schimb reciproc de segmente cromatidice prin copiere selectivă
Pentru a explica mecanismul ruperii şi schimbului de material cromatidic, C.D. Darlington a
propus o ipoteză conform căreia ruperea cromatidelor se datoreşte răsucirii foarte strânse a cromosomilor, unul în jurul celuilalt, în zigoten. Prin aceasta, s-ar crea o tensiune mare asupra cromatidelor externe. Ca urmare, acestea se rup, se derulează parţial şi, apoi, se resudează, fie în cadrul aceleiaşi cromatide, fie în cromatidele surori. Evident, în primul caz nu se poate vorbi de crossing-over, în cele de-al doilea, da. De aceea se apreciază că probabilitatea maximă de a se manifesta crossing-overul este de 50%.
Teoria copierii selective (replicaţiei selective) consideră că fenomenul de crossing-over
se realizează în stadiul S din interfaza ciclului celular. Plecând de la faptul că sinteza ADN şi, deci, a materialului cromosomial în ansamblu, se
realizează după modelul semiconservativ, s-a presupus că replicaţia cromosomului nu are loc după propria matriţă, ci după cea a cromosomului pereche şi vice-versa. În consecinţă, cromatidele nou formate ar copia o parte a materialului genetic de pe un cromosom şi o parte de pe celălalt cromosom. Aşa ar rezulta recombinările cromatidiene.
Verificarea experimentală a acestor ipoteze s-a efectuat prin marcarea unor bacteriofagi cu 15N. S-au infectat bacterii cu un astfel de virus, după care s-au cultivat pe un mediu cu 14N. Ipoteza copierii selective s-a dovedit neviabilă.
4.8. Chiasmata şi crossing-overul
Dovezile citologice ale crossing-overului pot fi obţinute pe parcursul profazei târzii a primei diviziuni meiotice, când chiasmata devine vizibilă. Este timpul în care cromosomii omologi menţin contacte strânse, atât în zona centromerului cât şi la chiasme. Este situaţia în care se pot număra, foarte exact, toate chiasmele. Cromosomii lungi, aşa cum am precizat cu altă ocazie, au mai multe chiasme decât cei scurţi. Altfel spus, cromosomii lungi au mai multe posibilităţi de crossing-over decât cei scurţi.
Apariţia chiasmelor în profaza primei diviziuni meiotice poate induce concluzia că atunci se produce crossing-overul. Sunt, însă, numeroase experimente care demonstrează că fenomenul crossing-over apare mult mai devreme. În unele dintre aceste experimente s-au utilizat şocuri de temperatură şi s-a constatat o modificare a frecvenţei recombinărilor. Când şocurile de temperatură erau administrate în profaza târzie, efectele erau mai mici decât atunci când au fost administrate mai devreme. De aceea s-a concluzionat că fenomenul crossing-over apare în profaza timpurie.
Dovezi suplimentare pentru această concluzie au fost furnizate de studiile la nivel molecular, studii efectuate în timpul autoreplicării semiconservative a ADN-ului. Deşi aproape toată cantitatea de ADN se sintetizează în timpul etapei S din interfază, se apreciază că o mică parte din ADN se sintetizează în timpul primei profaze meiotice. Acest proces întârziat de sinteză a fost interpretat ca având rolul de a repara rupturile ce apar în cromatide, în timpul crossing-over-ului. Analize de mare fineţe au stabilit că acest proces de sinteză apare anterior trecerii în cea de a doua jumătate a profazei şi nu mai târzziu. Prin urmare, crossing-over-ul apare, la rândul său, tot în prima jumătate a profazei, cu mult înainte de a fi vizibilă chiasmata.
Ce este, atunci, chiasmata şi ce înseamnă ea? Unii specialişti consideră că chiasmata reprezintă un vestigiu al procesului de schimb între cromosomii omologi. Cromatidele între care s-a produs schimbul de material cromatic rămân, probabil, ataşate una de alta în timpul profazei.
Fig. 16 Formarea chiasmelor şi producerea gameţilor recombinaţi (50%) şi de tip parental (50%)
Ulteerior punctele de contact între cromatide se desfac şi cromatidele se îndepărtează îndreptându-se, fiecare, pe fibrele fusului acromatic, spre câte unul dintre polii celulei. De aceea, probabil, fiecare chiasmă reprezintă un punct de sudură apărută printr-un crossing-over ce s-a derulat, probabil, în pachiten.
4.8. Crossing-over intragenic
Cunoscându-se mecanismul crossing-overului şi frecvenţa sa, multă vreme s-a considerat că el este mult mai uşor de explicat pentru genele ce se află plasate în loci îndepărtaţi. În cazul genelor apropiate, frecvenţa sa este mai mică şi posibilităţile de decelare scad. În experienţele cu D. melanogaster de pildă, nu se poate lucra cu mai mult de 50.000 indivizi deodată. Într-un asemenea lot experimental însă, uneori, este imposibil de decelat un crossing-over. Pe de altă parte la bacterii şi la virusuri se poate lucra simultan pe milioane de indivizi. Astfel, s-a constatat că fenomenul crossing-over poate apărea şi în interiorul genei.
INTREBARI DE VERIFICARE
1. Care au fost avantajele folosirii Drosophilei melanogaster in cercetarile de genetica 2. Ce reprezinta linkage-ul 3. Ce reprezinta si cand are loc crossing-overul 4. Intocmiti schemele hibridarilor care sa ilustreze fenomenele de linkage si crossing-over 5. Care sunt mecanismele moleculare care conduc la aparitia crossing-overului
TEME PENTRU EXAMINAREA FINALA
1. Definirea si explicarea fenomenului de linkage. 2. Intelegerea si aprofundarea aspectelor citologice si moleculare in cadrul fenomenului de crossing-over
Unitatea de invatare 5 5. Mutatiile
5.1. Definire, clasificaţie, generalităţi
Noţiunea de mutaţie a fost introdusă în ştiinţă de către Hugo de Vries (unul dintre descoperitorii legităţilor elaborate de Mendel) în anul 1901.
Mutaţia (de la latinescul mutatio = schimbare, modificare) reprezintă o modificare a conţinutului informaţiei ereditare a unui individ, prin alterări produse la nivel molecular, cromosomial, genomic sau celular, sub impactul unor factori de natură fizică, chimică şi biologică sau, pur şi simplu, prin hazard.
Mutaţiile, alături de recombinările intra- şi intercromosomiale, constituie principala sursă de asigurare a variabilităţii individuale - câmpul de acţiune al selecţiei naturale. Dar, în funcţie de transmiterea sau nontransmiterea lor în descendenţă, variaţiile produse în conţinutul informaţiei individuale, mai corect spus, variaţiile fenotipizate de individul purtător, sunt de două categorii:
- modificaţii; - mutaţii propriu-zise. Spre a înţelege ce reprezintă fiecare dintre ele, reamintim schema generală a circulaţiei
informaţiei (inclusiv a informaţiei ereditare) în şi între sisteme:
Canal
Sursă Perturbaţii Destinaţie
Figura 6.1 Schema generală a circulaţiei unui flux informaţional
În principiu, modificările produse în conţinutul informaţiei se pot datora intervenţiei
perturbaţiilor direct în sursă, pe canal sau la destinaţie. În ceea ce priveşte circulaţia informaţiei ereditare în sistemele biologice, se impun unele
precizări. Sursa informaţiei ereditare este asimilată, în cea mai mare parte a cazurilor, cu ADN-ul. ARN-ul mesager nu reprezintă altceva decât canalul pe care circulă informaţia ereditară, ulterior transcrierii sale din molecula ADN. Biomolecula sintetizată în final (prin procesul translaţiei mesajului conţinut în ARNm, în ordinea şi numărul aminoacizilor din respectiva biomoleculă), reprezintă destinaţia circulaţiei informaţiei. Conform cu dogma centrală a geneticii, sensul
circulaţiei informaţiei este univoc: ADN ARN Proteină (precizările şi corecţiile necesare le vom aduce la capitolul de genetică moleculară). Cu alte cuvinte, o perturbaţie poate acţiona, în cazul de faţă, în ADN (echivalat cu sursa), în ARN (echivalat cu canalul) sau în proteină (destinaţie). Evident, modificările produse în ARN sau proteină, deşi manifeste fenotipic, nu se transmit ereditar (dispar odată cu individul purtător). Acestea sunt modificaţiile.
Mutaţiile propriu-zise vizează exclusiv macromolecula de ADN şi, în măsura în care permit supravieţuirea individului purtător, se menţin definitiv în zestrea ereditară a populaţiei din care face parte acesta şi se transmit ca atare în succesiunea de generaţii.
Ca orice alt fenomen sau proces biologic şi mutaţiile sunt clasificate în funcţie de diverse criterii sau interese.
Fără a intra în detalii (care pot fi găsite în orice tratat de genetică), reamintim câteva dintre clasificaţii:
a. Mutaţii: - naturale; - artificiale.
b. Mutaţii produse de factori: - fizici; - chimici; - biologici.
Apreciem că cea mai adecvată clasificare, din punctul de vedere al biologului, este cea în care se ţine cont de mărimea substratului ereditar afectat, de cantitatea informaţiei afectată în sursă.
În conformitate cu acest criteriu putem vorbi despre mutaţii: - genice; - cromosomiale; - genomice.
Calcule efectuate de H.J. Muller au precizat că rata mutaţiilor (per locus) este de aproximativ 1/100.000. S-au constatat corelaţii destul de evidente între rata mutaţiilor şi gradul de
evoluţie (poziţia în arborele filogenetic) a speciilor. La om, de exemplu, mutaţia care provoacă retinoblastomul are o rată de 0,000023. Alte calcule estimează că rata mutaţiilor, per genă, la om este de 4/100.000.
Dacă notăm cu U rata mutaţiei per genom, cu u rata mutaţiei per genă şi cu N numărul total de gene, atunci N=U/u. La drosofila, de pildă, s-a ajuns la estimarea existenţei a 5.000 gene (N = 0,05/0,00001 = 5.000). Estimări mult mai recente ridică la 10-15.000 numărul genelor la Drosophila melanogaster.
5.2. Mecanismele producerii mutaţiilor
5.2.1. Mecanismele mutaţiilor genice
a) Deleţia reprezintă pierderea uneia sau mai multor perechi de nucleotide. b) Adiţia (adăugarea sau inserţia) uneia sau mai multor perechi de nucleotide.
c) Substituţia uneia sau mai multor perechi de nucleotide. d) Inversia unui fragment de ADN cuprinzând cel puţin două perechi de nucleotide.
Pentru exemplificarea celor patru mecanisme menţionate, imaginăm o situaţie ipotetică în care redăm o succesiune aleatorie de perechi de nucleotide (din catena unei macromolecule de ADN), perechi pe care le numerotăm cu cifre arabe. În funcţie de tipul mutaţiei (în funcţie de mecanismul prin care se produce mutaţia), vom dubla, vom anula sau vom înlocui cifrele respective. numărul nucleotidelor 1 2 3 4 5 6 7 8 9 10 nucleotidele unei catene A G C G T A G T C G punţi de hidrogen II III III III II II III II III III nucleotidele complementare T C G C A T C A G C
Catenă (secvenţă) normală
numărul nucleotidelor 1 3 4 5 6 7 8 9 10 nucleotidele unei catene A C G T A G T C G punţi de hidrogen II III III II II III II III III nucleotidele complementare T G C A T C A G C
Deleţia perechii a II-a de nucleotide
numărul nucleotidelor 1 2‟ 2 3 4 5 6 7 8 9 10 nucleotidele unei catene A C G C G T A G T C G punţi de hidrogen II III III III III II II III II III III nucleotidele complementare T G C G C A T C A G C
Adiţia unei perechi (2’) de nucleotide numărul nucleotidelor 1 2 3 4 5 6 7 8 9 10 nucleotidele unei catene A C C G T A G T C G punţi de hidrogen II III III III II II III II III III nucleotidele complementare T G G C A T C A G C
Substituţia perechii a 2-a de nucleotide
numărul nucleotidelor 1 2 3 6 5 4 7 8 9 10 nucleotidele unei catene A G C A T G G T C G punţi de hidrogen II III III II II III III II III III nucleotidele complementare T C G T A C C A G C
Inversia secvenţei 4 5 6
Deci, se schimbă fie numărul, fie ordinea perechilor de nucleotide. În consecinţă se schimbă numărul şi tipul (sau ordinea) codonilor, ceea ce înseamnă, în fapt, o schimbare a conţinutului informaţiei ereditare. Transcripţia se efectuiază după o informaţie modificată, translaţia având ca rezultat sinteza unei biomolecule schimbate (alt număr şi altă succesiune a aminoacizilor).
e) Mutaţiile moleculare (genice) mai pot avea drept cauză şi unele erori în procesul de autoreplicare semiconservativă a moleculei de ADN.
Respectivele erori se pot datora fenomenelor: e.1. Tranziţie - înlocuirea unei baze purinice sau pirimidinice cu o altă bază, tot purinică
sau pirimidinică. A G şi T C
e.2. Transversie - înlocuirea unei baze purinice cu una pirimidinică şi invers.
A T şi A C
G T şi G C
Schematic, transversiile şi tranziţiile se pot reda astfel:
Tranziţie
A G Purinice
Transversie Transversie
T C Pirimidinice Tranziţie A T G C T G G T II II III III II III III II catena iniţială de ADN T A C G A C C A A C G T T G G T II III III II II III III II tranziţie T G C A A C C A
A T G C A C T G II II III III II III II III transversie T A C G T G A C
Judecate prin prisma schimbărilor ce le produc în final (la sfârşitul procesului de translaţie), mutaţiile genice pot fi considerate:
Nonsens, acele care duc la degenerarea codului genetic şi nu se reflectă în schimbarea
succesiunii aminoacizilor în molecula proteică. Spre exemplu, codonii UAU şi UAC codifică acelaşi aminoacid - tirozina. Pe de altă parte, mutaţiile survenite în codonii stop (terminatori ai sintezei unei catene proteice) - UAA, UAG şi UGA, nu au nici o semnificaţie.
Mutaţii cu sens greşit, care transformă un codon funcţional (din cei 61) într-un codon
terminator. În acest caz, molecula polipeptidică sintetizată va fi mai scurtă.
Mutaţiile cromosomiale sunt, în fapt, de două mari categorii - structurale şi numerice. Mutaţiile structural cromosomiale presupun alterări ale materialului cromosomial,
determinate de rupturi şi pierderi sau realipiri (în altă ordine şi în alte poziţii) de segmente cu lungimi variabile. În funcţie de localizarea restructurării, mutaţia structural cromosomială poate fi:
- intracromosomială; - intercromosomială. Cele intracromosomiale pot fi, la rândul lor, intraradiale şi interradiale (adică în cadrul
aceluiaşi braţ sau între cele două braţe ale cromosomului. Indiferent dacă afectează un singur braţ sau ambele braţe cromosomiale, mutaţiile intracromosomiale sunt: deleţii, duplicaţii, inversii, translocaţii.
Deleţiile reprezintă o pierdere de material cromosomial. În funcţie de zona cromosomială pe care o afectează, ele sunt: terminale şi intercalare,
acestea din urmă fiind paracentrice şi pericentrice. Deleţiile terminale afectează capătul cromosomului şi presupun producerea unei singure
rupturi. Întrucât producerea unei deleţii terminale presupune pierderea regiunii telomerice, repercusiunile ei sunt destul de grave. Deleţia terminală determină scurtarea cromosomului şi formarea unui segment acentric, care, de obicei, se pierde: A B C D • E F G H I A B C D • E F G H I
segment acentric Deleţiile intercalare paracentrice presupun producerea a două rupturi, urmate de
eliminarea unui segment cromosomial acentric (din braţul scurt sau lung) şi scurtarea cromosomului: A B C D • E F G H I G H ABC D • E F I
Deleţiile intercalare pericentrice sunt datorate formării unui segment cromosomial centric prin producerea a două rupturi în imediata vecinătate a centromerului şi a două (sau unul, în cazul reunirii lor) fragmente acentrice.
A B C D • E F G H I A B C D • E F G H I
Deci, ca rezultat direct al producerii deleţiilor, putem avea: - fragmente acentrice; - inele acentrice; - cromosomi inelari; - discontinuităţi cromatice pe lungimea cromosomului; - isocromosomi; - micronuclee. Duplicaţiile reprezintă adăugarea (dublarea) uneia sau mai multor gene, la cele existente
deja în locusul (locii) respectiv. În funcţie de modul concret în care are loc dublarea, duplicaţiile pot fi: în tandem direct sau în tandem invers. Şi unele şi celelalte, în funcţie de zona cromosomială în care se produc, sunt: terminale şi intercalare, acestea din urmă fiind: paracentrice şi pericentrice.
Duplicaţiile în tandem direct pot fi reprezentate schematic astfel: A B C D • E F G H I cromosomul iniţial A B A B C D • E F G H I terminală A B C B C D • E F G H I intercalară paracentrică A B C D D • E E F G H I intercalară pericentrică
Duplicaţiile în tandem invers pot fi redate schematic astfel A B C D • E F G H I cromosomul iniţial A B B A C D • E F G H I terminală A B C C B D • E F G H I intercalară paracentrică A B C E E • D D F G H I intercalară pericentrică
De obicei, frecvenţa duplicaţiilor este mai mare decât cea a deleţiilor şi pentru motivul că duplicaţiile nu au efecte nocive.
Inversiile se datoresc ruperii unui fragment cromosomial, paracentric sau pericentric, rotirii
sale cu 180o în planul longitudinal şi resudării capetelor în noua poziţie. Evident, având în vedere existenţa telomerului la fiecare capăt cromosomial, inversiile terminale sunt imposibile.
Pe de altă parte, dacă inversiile paracentrice nu produc modificări în morfologia (adică în tipul) cromosomului, inversiile pericentrice pot determina schimbări consistente în lungimea braţelor, dacă cele două porţiuni, rupte de o parte şi de alta a centromerului, nu sunt egale. De aceea, inversiile pericentrice pot fi egale sau inegale.
Schematic, situaţiile menţionate pot fi redate astfel: A B C D • E F G H I cromosomul iniţial A C B D • E F G H I paracentrică A B C E • D F G H I pericentrică egală A E • DCB F G H I pericentrică inegală
Translocatiile reprezintă fenomenul prin care o porţiune (un fragment), dintr-un cromosom
se rupe şi se realipeşte, fie în acelaşi cromosom (în acelaşi braţ sau în braţul celălalt), fie în cromosomul omolog, fie într-un cromosom neomolog. Translocaţiile (cunoscute şi sub numele de transpoziţii) pot fi, deci şi intracromosomiale (intraradiale şi interradiale) şi intercromosomiale. Evident şi la translocaţii se pun aceleaşi probleme vis à vis de capătul cromosomului - prezenţa telomerilor împiedică producerea de translocaţii terminale.
Translocaţiile intracromosomiale pot fi schematizate după cum urmează: A B C D • E F G H I cromosom iniţial A B C D • E G H F I intraradială A B F C D • E G H I interradială
În ceea ce priveşte translocaţiile intercromosomiale se cuvin menţionate câteva aspecte deosebite implicate de producerea lor. Aşa cum aminteam anterior, ele pot viza cromosomii omologi sau neomologi. Evident, dacă modalităţile concrete de producere (rupere şi apoi alipire) sunt identice, rezultatele lor sunt diferite.
În cazul în care cei doi cromosomi omologi sunt izogenici (adică nu se află în raport de alelism), translocaţia presupune o duplicaţie (dacă vizează izolocii de pe cei doi cromosomi):
A B C D • E F G H I A D • E F G H I A B C D • E F G H I A B C B C D • E F G H I
Dacă cei doi cromosomi sunt în raport de alelism, situaţia se prezintă astfel: A B C D • E F G H I A D • E F G H I a b c d • e f g h i a B C b c d • e f g h i
Dacă translocaţia va viza schimbul între cele două braţe neomoloage ale cromosomilor omologi, evident că nu vom mai avea duplicaţie. În esenţă (ca mecanism vorbind), fenomenul este similar cu cel al translocaţiei între doi cromosomi neomologi, când vom avea:
A B C D • E F G H I A D • E F G H I M N O P • R S T U V M N B C O P • R S T U V
În cazul translocaţiilor intercromosomiale, dacă porţiunile translocate vor fi intercalare (acetrice), unul dintre cromosomi se scurtează (donatorul) şi celălalt se alungeşte (primitorul).
În situaţia în care porţiunea translocată este pericentrică (adică segmentul translocat conţine şi centromerul), rezultatul va fi apariţia unui cromosom dicentric şi a unor fragmente acentrice care se vor resorbi.
A B C D • E F G H I A B C E G H I M N O P • R S T U V M N D • E O P • R S T U V
Procesul se complică şi mai mult în cazul în care schimburile de segmente cromosomiale sunt bidirecţionale (reciproce). De această dată, putem vorbi şi despre translocaţii terminale. Evident, translocaţiile reciproce, fie că sunt terminale sau intercalare (paracentrice sau pericentrice), pot fi simetrice sau asimetrice (adică vizează un schimb de segmente egale sau inegale):
A B C D • E F G H I A B C D • E F G U V M N O P • R S T U V M N O P • R S T H I
A B C D • E F G H I A B C D • E S T I M N O P • R S T U V M N O P • R F G H U V
A B C D • E F G H I A B C P • R F G H I M N O P • R S T U V M N O D • E S T U V
Rezumând, putem conchide că translocaţiile sunt: A. Intracromosomiale:
- intraradiale; - interradiale.
B. Intercromosomiale: - între cromosomii omologi:
- între braţele omoloage (duplicaţii); - între braţe neomoloage.
- între cromosomi neomologi: - extraradiale directe (unidirecţionale):
- paracentrice; - pericentrice.
- extraradiale reciproce (bidirecţionale): - terminale simetrice sau asimetrice; - intercalare paracentrice simetrice sau asimetrice; - intercalare pericentrice simetrice sau asimetrice.
Fenomenul de fuzionare a cromosomilor telocentrici presupune existenţa a cel puţin doi
cromosomi telocentrici. Aceştia pot fuziona (prin fuzionarea centromerilor) dând, astfel, naştere unui cromosom cu centromer median (metacentric sau submetacentric, în funcţie de lungimea celor doi cromosomi).
5.2.3. Mecanismele mutaţiilor genomice
Mutaţiile genomice presupun o modificare a numărului seturilor cromosomiale fundamentale (de bază), notate în mod convenţional cu x. Reamintim ceea ce am descris şi elucidat pe deplin în capitolul dedicat structurii ţi funcţiilor cromosomilor şi anume: x = n numai în
cazul speciilor diploide şi, mai mult decât atât, într-un anumit gen, unde avem mai multe numere cromosomiale de bază (adică mai multe valori pentru x), unul a provenit din altul, concomitent cu diversificarea speciilor. Deci, nu întotdeauna există echivalenţă între genom (notat cu n) şi numărul fundamental de cromosomi (x). Altfel spus, între evoluţia pe orizontală (diversificarea specifică) şi
variabilitatea numărului fundamental de cromosomi din cadrul unui gen, se pot stabili interrelaţii precise.
Mulţi autori plasează aneuploidia în cadrul mutaţiilor genomice. Din punctul nostru de vedere, aneuploidia reprezintă o mutaţie cromosomială şi anume, o mutaţie numeric cromosomială, întrucât ea afectează unul sau doi cromosomi (rareori trei sau patru, aşa cum vom preciza la momentul potrivit) dintr-un genom şi nu multiplicarea numărului genomurilor sau a numărului fundamental de cromosomi.
În consecinţă, mutaţiile genomice includ: haplodia, poliploidiile şi pseudopoliploidiile. Haploidia starea firească în care se găseşte un individ eucariot este starea diploidă.
Această stare se reconstituie în urma fiecărui act de fecundare, act în urma căruia, din fuziunea a doi gameţi de sex opus, ia naştere un zigot ce întruneşte suma cromosomilor existenţi în fiecare din cei doi gameţi. Tot prin definiţie, gameţii sunt haploizi, adică au jumătate din numărul de cromosomi caracteristic părinţilor. Deci, n♀ + n♂ = 2n ♀ sau ♂.
Rezultă, cu claritate, că dacă (din anumite motive), un viitor individ se va dezvolta având ca sursă un singur gamet (androgeneză sau ginogeneză, pseudogamie, embrionie adventivă etc.), în celulele sale se vor găsi n cromosomi şi nu 2n. Fenomenul poartă numele de haploidie.
Prin fuziunea unui gamet ♀ cu unul ♂ rezultă un zigot (punctul de start al noului organism)
care poate fi homozigot dominant, recesiv sau heterozigot (AA, aa sau Aa). Fenomenul este imposibil la haploizi, întrucât aceştia au un singur set din cromosomii omologi (un singur genom), deci au toate genele într-o singură doză (A sau a). Prin urmare, fiind exclus fenomenul de alelism, implicit dominanţa şi recesivitatea, un haploid fenotipizează totalitatea genelor pe care le conţine. Evident, lipsind dominanţa şi recesivitatea, alelele profund dăunătoare, mai ales alelele letale, induc moartea indivizilor haploizi. De aceea, la acest nivel, selecţia este mult mai dură şi mult mai eficientă.
În acest context, se cuvin menţionate şi unele avantaje, din punctul de vedere al practicii, oferite de haploidie. Prin dublarea setului haploid de cromosomi (diploidizarea unui haploid) se obţin indivizi izogenici (izocromosomici), sau aşa zisele linii pure (genetic pure), a căror producere necesită multe eforturi şi mult timp pe căile convenţionale.
Fiind parţial sterili, haploizii au o mică incidenţă în mod natural. Se întâlnesc, pe anumite perioade şi în anumite condiţii, la grupe de insecte (albine, viespi, afide), reprezintă o stare quasinormală la ciupercile cu spori, algele unicelulare, briofite şi sunt starea normală a procariotelor.
Pe de altă parte, diploidia poate reprezenta o stare normală a unui individ sau poate fi rezultanta unei autopoliplodizări sau alopoliploidizări. Evident, haploizii proveniţi din cele trei categorii de "diploizi" nu vor avea comportamente similare. De aici şi posibilitatea clasificării haploizilor în:
a. monoploizi; b. polihaploizi. a. Monoploizii provin prin dezvoltarea autonomă a unei celule sexuale (androgeneză sau
ginogeneză) de la indivizi aflaţi într-o stare de diploidie adevărată. În cazul plantelor, un monoploid se poate obţine şi în urma dezvoltării unei alte celule decât cea gametică, dar aparţinând tot fazei haploide (gametofitului), cum ar fi antipodele şi sinergidele.
Printre modalităţile practice de obţinere a haploizilor (indiferent de sorgintea şi tipul lor) putem menţiona: îndepărtarea staminelor (la florile hermafrodite) anterior maturizării lor, polenizarea cu polen distrus prin iradiere, polenizarea cu polen străin etc.
La animale, pentru inducerea partenogenezei ovulelor nefecundate, se utilizează spermatozoizi iradiaţi, tratamente cu şocuri termice, soluţii hipotonice şi hipertonice etc.
b. Polihaploizii provin. prin mecanisme identice celor ce determină apariţia
monohaploizilor, din poliploizi. Poliploidia, evidenţiată pentru prima dată de către G. Winkler (1916), reprezintă mutaţia
genomică datorată multiplicării numărului de genomuri (numărului fundamental de cromosomi). Poliploidia reprezintă un fenomen cu o incidenţă destul de mare în natură - 47% dintre angiosperme şi 4,6% dintre gimnosperme sunt poliploide.
Poliploizii se dovedesc a fi competitori redutabili în condiţii aspre de mediu, proporţia lor crescând în biocenoze concomitent cu latitudinea şi altitudinea, precum şi în paralel cu deteriorarea drastică a condiţiilor de habitat (deşerturi, soluri sărăturate etc.).
În funcţie de numărul par sau impar al genomurilor întrunite în noul organism, poliploizii pot fi clasificaţi în:
A. Artioploizi - cu număr par de genomuri (2x, 4x, 6x, 8x, 10x).
B. Perisoploizi - cu număr impar de genomuri (3x, 5x, 7x, 9x).
În ceea ce priveşte poliploizii, se cuvin menţionate câteva aspecte. Crescând numărul de seturi cromosomiale per nucleu, evident că va creşte şi cantitatea de AND per nucleu. Poliploidia este un fenomen larg răspândit la plante, animalele etalând cu o frecvenţă mult mai mică această stare. În acest regn poliploidia este mai frecventă la nevertebrate (protozoare, crustacei, lepidoptere, ortoptere), dar şi la unele vertebrate, din clasele peştilor şi amfibienilor. Menţionăm în acest sens ţiparul (Misgurnus fossilis), care este tetraploid în apele noastre (2n=100), în timp ce în apele din Extremul Orient trăieşte o specie diploidă (Misgurnus anquillicaudatus)(2n=50).
Prezenţa poliploizilor a fost semnalată şi la reptile, păsări şi chiar mamifere. Desigur poliploizii sunt prezenţi şi la om dar, în mod firesc, nu sunt viabili.
Conform originii lor (modului în care au apărut), poliplozii au fost grupaţi în: I Autopoliploizi şi II Alopoliploizi. I. Autopoliploidia reprezintă fenomenul multiplicării numărului de genomuri (numere
cromosomiale de bază) proprii. Autotetraploizii se întâlnesc frecvent la Solanusm tuberosum, Medicago sativa, Arachis
hypogea, Coffea arabica, Vitis vinifera etc. În practică sunt deosebit de frecvent utilizaţi triploizii care, pe lângă faptul că manifestă heterozis, prezintă şi avantajul de a nu produce seminţe. Cei mai des întâlniţi şi mai frecvent utilizaţi sunt triploizii bananului, viţei de vie, mărului, pepenelui verde, ananasului, plopului etc.
II. Alopoliploizii (amfiploizii) reprezintă poliploizii a căror apariţie presupune, cu necesitate,
prezenţa unui proces anterior de hibridare interspecifică. În consecinţă, la un amfiploid nu putem vorbi de o multiplicare a numărului de genomuri proprii, ci de o multiplicare a unui număr de genomuri aditive. Spre exemplu, dacă avem specia A, la care notăm genomul cu x1 şi specia B, la care notăm genomul cu x2, hibridul interspecific AB va vea genotipul x1+x2. Dar acest hibrid nu se va putea reproduce, meioza nefiind posibilă. Prin poliploidizarea sa la faza de tetraploid vom avea hibridul AABB, cu genotipul 2x1+2x2, ceea ce nu este tot una cu 4x.
Se apreciază că în natură incidenţa amfiploizilor este destul de mare. Dintre aceştia, menţionăm: Triticum aestivum (2n=42), Gossipium hirsutum (2n=52), Nicotiana tabacum (2n=48), Medicago sativa (2n=32), Prunus domestica (2n=48), Spartina townsendii (2n=126), Brassica napus (2n=38) etc.
Unul din cele mai bine studiate cazuri este acela al speciei Triticum aestivum. Aici avem, în esenţă, rezultanta a două hibridări interspecifice, urmate de poliploidizare. Astfel, Triticum monococcum (2n=14) s-a hibridat cu Aegilops speltoides (2n=14). A rezultat Triticum dicoccoides (2n=28). Acesta, încrucişat cu Aegilops squarrosa (2n=14), a dat specia Triticum aestivum
(2n=42). Iată, schematic, procesul:
Triticum monococcum x Aegilops speltoides 2n = 14 (2xm), genomurile AA 2n = 14 (2xsp), genomurile BB
Rezultă un hibrid steril, 2n = 14, cu genomurile AB (xm + xsp).
Aegilops squarrosa x Triticum dicoccoides 2n=14(2xsq), genomurile DD 2n=28(2xm+2xsp=4x),genomurile AABB
Rezultă un hibrid steril, 2n = 21, cu genomurile ABD (xm + xsp +xsq). Prin alopoliploidizare hibridul devine Triticum aestivum, 2n=42(2xm+2xsp+2xsq=6x), cu genomurile AABBDD
Evident, Triticum aestivum este o specie nouă, sintetică, fertilă şi foarte productivă.
Se mai cuvine de menţionat faptul că, tot la grâu, japonezul H. Kihara a reuşit să resintetizeze unele specii existente în natură, refăcând astfel drumuri parcurse de procesul speciaţiei în genul Triticum.
Pe de altă parte, s-a reuşit sinteza de novo a unor specii complet necunoscute în natură. Astfel, G.D. Karpecenko, în 1927, a reuşit să hibrideze Raphanus sativus (2n=18) cu Brassica oleracea (2n=18) şi să obţină Raphanobrassica (2n=36). Iniţial, hibridul era steril. Dar, după un
timp de vegetaţie, pe una din ramuri s-au format câteva seminţe. Se admite deci că, în mod natural, s-au produs gameţi diploizi care au asigurat fertilitatea şi perpetuarea hibridului. Noua specie (sintetică) are caractere intermediare celor două specii genitoare - forma păstăilor, de pildă, este cea de la ridiche la partea superioară şi cea de la varză la partea inferioară.
Pseudopoliploidia sau poliploidia falsă reprezintă fenomenul în care multiplicarea
numărului de genomuri se datorează unei fisionări a cromosomilor (perpendicular, la nivelul centromerilor) şi nu unor dereglări în procesul diviziunii celulare. În consecinţă, la un pseudopoliploid, mărirea numărului de genomuri nu se regăseşte în mărirea adecvată a cantităţii de ADN. În altă ordine de idei, precizăm faptul că, în cadrul cariotipului unei specii, pe lângă numărul şi tipul cromosomilor, o caracteristică specifică o constituie numărul fundamental de braţe (NF). De pildă, la o specie cu 2n=10, toţi cromosomii fiind metacebtrici, NF=20. În cazul
în care o pereche de cromosomi din cei 10 (una din cele cinci perechi) sunt telocentrici, restul fiind metacentrici, NF=18 etc.
Evident, în cazurile de pseudopoliploidie, indiferent de numărul de genomuri, NF rămâne constant (ca şi cantitatea de ADN per celulă). De pildă, în cazul menţionat, diploidul are 2n=10, adică 2x, deci NF=20 (toţi cromosomii sunt metacentrici), Prin fisionare la nivelul centromerului, toţi cromosomii devin telocentrici. Prin urmare 2n=20, ceea ce ar reprezenta 4x. Dar NF=20, deci este vorba de un 4x fals.
Endopoliploidia reprezintă fenomenul aşa-zisei poliploidii somatice, adică o poliploidizare
prin cicluri endomitotice repetate (replicarea cromosomilor în interiorul membranei nucleare). Endopoliploidia nu afectează întregul organism ci, de obicei, anumite celule sau ţesuturi ale
acestuia. Fenomenul a fost constatat şi descris, pentru prima oară, de către citologul austriac L.
Geitler, la insectele acvatice Gerris lateralis şi Gerris lacustris, la care s-au identificat celule 512-,
1024- şi chiar 2048 ploide, în ţesutul ovarian. Sunt date în conformitate cu care la specia Chironomus tentans, în glandele salivare,
celulele conţin 32.768 n. Fenomenul de endopoliploidie a fost observat şi la organisme mult mai evoluate, cum ar fi şobolanul (Rattus rattus).
La angiosperme, endopoliploidia este un fenomen comun. La sfecla de zahăr spre exemplu, în endospermul seminţelor se întâlnesc celule cu 24.756 n.
Aneuploidia (heteropoliploidia) reprezintă fenomenul datorat prezenţei în plus a 1-2
cromosomi sau lipsei a 1-2 cromosomi dintr-un genotip. Evident, prezenţa unor cromosomi supranumerari sau lipsa lor, se regăseşte în augmentarea sau diminuarea conţinutului de ADN şi în modificarea corespunzătoare a numărului fundamental de braţe.
Dacă un individ diploid normal mai poartă şi numele de disomic (în sensul că fiecare
pereche de cromosomi conţine doi omologi - unul de origine maternă şi altul de origine paternă), heteroploizii pot fi de două categorii: hipoploizi şi hiperploizi.
Hipoploidia se caracterizează prin lipsa unuia sau a doi cromosomi din setul diploid. a. Nulisomia (2n – 2). În acest caz lipseşte una dintre perechile de cromosomi. De
exemplu, dacă o specie are 2n=10, nulisomicii vor avea 2n=8. În consecinţă, vom avea o serie de 5 nulisomici diferiţi (câte unul pentru fiecare pereche: I, II, III, IV şi V).
b. Monosomia (2n – 1). În acest caz, dintr-o pereche de cromosomi lipseşte unul. Deci,
rămânând la acelaşi exemplu, vom avea 10 monosomici diferiţi (în ideea că cei doi cromosomi ai unei perechi, unul provenit de la mamă şi unul de la tată, nu sunt identici - unul poate avea genele dominante şi celălalt, alelele recesive).
c. Dublumonosomia [2n – (1+1)]. Semnifică lipsa a 2 cromosomi aparţinând la două
perechi diferite. În exemplul nostru, vom avea 40 de dublumonosomici diferiţi. Hiperploidia reprezintă fenomenul datorat unui proces oarecum invers, adică prezenţa a
unul sau doi cromosomi în plus.
a. Trisomia (2n+1). La una (în fiecare caz alta) dintre perechile de omologi, în loc de 2
cromosomi vor fi 3. În consecinţă, vom putea identifica 10 trisomici diferiţi în cazul în care menţinem exemplul speciei cu 2n=10 cromosomi.
b. Tetrasomia (2n+2). Presupune dublarea unei perechi de cromosomi (figurat vorbind,
reprezintă un caz de tetraploidie pentru o pereche de cromosomi). Prin urmare, la 2n=10 vom avea 5 tetrasomici diferiţi.
c. Dublutrisomia [2n+(1+1)]. Reprezintă fenomenul invers dublumonosomiei, rezultând 40
de dublutrisomici diferiţi, posibili. În contextul acestei problematici precizăm faptul că geneticianul E.R. Sears, în anul 1959, a
efectuat studii complete şi complexe asupra tetrasomiei în cazul speciei Triticum aestivum (2n=42), ocazie cu care a identificat şi a descris seria completă de 21 tetrasomici (2n=44).
Hipo-hiperploidia (2n – ? + ?) reprezintă fenomenul rezultat prin combinarea aleatorie a
procesului de hipoploidie cu cel de hiperploidie. Adică simultan, la acelaşi individ, pentru aceeaşi pereche putem sesiza prezenţa unui cromosom în plus, în timp ce la o altă pereche un cromosom lipseşte etc.
Se apreciază că principala cauză a apariţiei aneuploizilor o constituie perturbarea meiozei. În consecinţă iau naştere gameţi cu 1-2 cromosomi în plus sau în minus, fenomen ce duce la apariţia de indivizi modificaţi (cu 1-2 cromosomi în plus sau cu 1-2 cromosomi în minus).
Dacă la multe vieţuitoare, în special la plante, aneuploidia se constituie într-o sursă importantă de variabilitate, fiind şi unul dintre mecanismele asigurării apariţiei de noi numere cromosomiale de bază (x), la animalele superior organizate, mai precis la om, aneuploidia reprezintă o aberaţie incompatibilă cu viaţa normală a individului. Indiferent dacă afectează autosomii sau heterosomii, aneuploidia reprezintă sindroame (boli genetice) deosebit de grave. Spre exemplu, trisomia 21 reprezintă sindromul (maladia) Down (cunoscut şi sub numele de idioţie), indivizii monosomici pentru heterosomi (X0=viabili şi Y0=letali) ca şi trisomicii (XXX sau XXY) reprezintând, de asemenea, boli grave (sindromul Turner şi, respectiv, Klinefelter).
În acest caz, aneuploidia este destul de răspândită în lumea plantelor. Este cunoscut cazul zambilei (Hyacinthus orientalis, 2n=16), introdusă în cultură în anul 1560 în Olanda. Pe parcursul
timpului, în urma selecţiei empirice s-au obţinut forme triploide, tetraploide şi aneuploide foarte diferite între ele din punct de vedere estetic. Cele din urmă (cele aneuploide) se multiplică mai ales pe cale vegetativă.
5.2.5. Pseudoaneuploidia
Dacă la poliploidie sau aneuploidie variabilitatea numerică a genomurilor sau a cromosomilor se regăsea într-o variabilitate a cantităţii de ADN, în pseudoaneuploidie (ca şi în pseudopoliploidie) nu se regăseşte. În consecinţă, pseudoaneuploidia reprezintă un fenomen de falsă aneuploidie, datorat fuzionării sau fisionării cromosomilor.
Spre exemplu, un "monosomic" poate reprezenta, în realitate, rezultanta fuzionării cromosomilor unei perechi, iar un "nulisomic" poate reflecta rezultanta fuzionării cromosomilor din două perechi. În replică, un "trisomic" poate reprezenta, în fond, rezultanta fisionării unui cromosom, iar un "tetrasomic" poate reflecta rezultanta fisionării ambilor cromosomi dintr-o pereche sau a 2 cromosomi din două perechi diferite etc. Desigur, pentru ca cele două procese antagonice (fisiune şi fuziune) să aibă o incidenţă crescută, este importantă prezenţa cromosomilor telocentrici (pentru fuziune) sau metacentrici, submediani şi subtelocentrici (pentru fisiune).
Un element important în stabilirea existenţei unei aneuploidii veridice sau a pseudoaneuploidiei, îl constituie NF (numărul fundamental de braţe). Evident, dacă două specii
înrudite au numere diferite de cromosomi dar totalizează, în fiecare dintre celule, acelaşi număr de braţe cromosomiale, rezultă că ele au luat naştere printr-o falsă aneuploidie (pseudoaneuploidie).
Un exemplu de aneuploidie naturală, în cadrul regnului animal, ni-l oferă genul Drosophila. Specia iniţială, D. virilis, avea 2n=12. Prin fuziuni cromosomiale succesive, din ea au luat naştere speciile: D. pseudoobscura (2n=10), D. melanogaster (2n=8) şi D. willistoni (2n=6).
INTREBARI DE VERIFICARE
1. Ce reprezinta mutatiile si care este rolul acestora 2. Care este mecanismul molecular de aparitie al mutatiilor 3. Ce reprezinta mutatiile cromosomiale 4. Care sunt mecanismele de producere a mutatiilor genomice
TEME PENTRU EXAMINAREA FINALA
1. Precizarea rolului si mecanismelor de producere a mutatiilor 2. Clasificarea mutatiilor in functie de cantitatea de material genetic afectata
Unitatea de invatare 6
6. Acizii nucleici – suport material al informatiei genetice
6.1. Acizii nucleici sunt macromolecule semantice
Dupa 1940, cercetări efectuate pe ciuperca Neurospora au oferit noi argumente (con-
comitent cu noi întrebări), pentru o ipoteză formulată cu ani în urmă de Garrod, în conformitate cu
care genele „controlează” producerea enzimelor specifice. Aceasta a fost prima concepţie relativă la genă, cunoscută în genetică sub forma „o genă - o enzimă”, concepţie în conformitate cu care,
o anumită genă este responsabilă de determinismul sintezei unei anumite enzime. Iniţial, s-a emis o ipoteză simplă: gena determină ordonarea a 20 de aminoacizi diferiţi într-
un lanţ polipeptidic. O asemenea ipoteză presupunea acceptarea prezenţei unui mare număr de enzime, câte una pentru fiecare aminoacid diferit, ce va fi asamblat în proteina ce urma a fi sintetizată. Dar, fiecare enzimă având şi o parte proteică, ar fi fost necesare alte enzime (enzime suplimentare) pentru sinteza enzimelor ordonatoare de aminoacizi şi aşa mai departe. Astfel, o problemă ar genera o alta, la infinit. În consecinţă, concepţia „genei de natură proteică” a trebuit
să fie abandonată. A fost, deci, necesară elaborarea unei alte ipoteze. Aceasta a fost "ipoteza matriţei", adică
ipoteza existenţei unei suprafeţe şablon. In conformitate cu această ipoteză, ar exista unele
con-formaţii cu calitate de matriţă, pe suprafaţa cărora s-ar asigura selectarea unui anumit aminoacid pentru o anumită poziţie (locus) şi, ulterior, asamblarea aminoacizilor între ei. Astfel s-ar fi putut explica o a doua aserţiune şi anume: o enzimă specifică pentru un anumit aminoacid, dar comună pentru sinteza tuturor proteinelor, poate asigura asamblarea aminoacizilor.
Apare, totuşi, o dificultate şi aici. Conform acestei ipoteze, ar fi fost necesar să se accepte că o astfel de matriţă ar trebui să aibă capacitatea de a fi suport pentru propria-i asamblare (adică o matriţă trebuie să fie capabilă să dea naş-tere la noi matriţe). Practic, impasul în care a intrat prima ipoteză s-ar menţine - o enzimă ar trebui să dea naştere unei alte enzime şi, aşa cum s-a demonstrat, enzimele nu pot produce enzime.
Mergând în continuare cu logica, se mai impune o rezolvare şi anume : matriţele trebuie să fie macromolecule cel puţin la fel de mari ca moleculele cărora le dau naştere.
Analiza grupărilor aminoacizilor din proteine nu a adus, însă, confirmări pentru ipoteza enunţată anterior, întrucât nu există, din punct de vedere chimic, nici o posibilitate de a explica de ce inserarea valinei pe o matriţă ar atrage, preferenţial, o altă moleculă de valină pe matriţa complementară. In realitate, nici o grupare de aminoacizi nu are afinităţi pentru una similară. Dimpotrivă, este mult mai uşor să se imagineze molecule cu trăsături opuse sau complementare, care să se atragă reciproc. Spre exemplu, atomii electropozitivi de hidrogen, leagă numai atomi electronegativi, cum ar fi cei de oxigen sau azot. In acelaşi fel, moleculele pot fi atrase, în mod specific, prin forţe van der Waals, numai când posedă forme complementare. In context, se poate imagina că o adâncitură într-o moleculă poate fi complementară unei protuberanţe din molecula
complementară. Aşa stând lucrurile, s-a constatat că nu este posibilă gruparea celor 20 de aminoacizi în 10 perechi constituite din aminoacizi cu forme comple-mentare.
Se poate admite, însă, că există 20 de molecule specifice diferite, pe care le putem numi „conectori”. Fiecare conector ar trebui să posede două suprafeţe identice, dar complementare, unui anumit aminoacid. Respectivele molecule (conectoare) ar putea determina încorporarea unor aminoacizi într-un lanţ, într-o secvenţă identică celei a lanţului polipeptidic ce serveşte ca matriţă. Insă, nu s-au găsit dovezi pentru demonstrarea prezenţei unor asemenea matriţe.
Infirmarea ipotezei matriţei de natură proteică a fost argumentată şi chimic.
Principalul suport al ipotezei respective este acela că matriţa are, în structura sa, acizi aminici.
In acest context, se impune precizarea că o matriţă a cărei specificitate depinde de aminoacizi ca valina şi/sau alanina (aminoacizi extrem de apropiaţi conformaţional - ca structură spaţială), nu poate fi copiată cu suficientă acurateţe. Există riscul de a se asambla alanina în loc de valină sau invers. Un alt exemplu îl constituie valina şi izoleucina care diferă doar prin prezenţa unui grup metil adiţional la izoleucină (Fig.17). De asemenea, glicina şi alanina diferă între ele
printr-o grupare metil etc. Similitudinile structurilor spaţiale pun o problemă simplă - poate exista siguranţa că un
proces de copiere este suficient de sigur (ca acurateţe), adică poate să asigure diferenţa între două structuri aşa de asemănătoare ? Calculele au demonstrat că şansele de copiere nu implică mai mult de o eroare în 108 cazuri. Pe de altă parte, un argument de ordin chimic, nu prea riguros, este acela că o reacţie chimică, nu poate face distincţie între două molecule care diferă printr-o grupare metil, cu o acurateţe care să permită mai puţin de o greşeală în 106 cazuri. Apoi, când s-au realizat experimentele propriu-zise, s-a constatat că leucina, de exemplu, era inserată într-un polipeptid cu o siguranţă de cel mult 99,9%, ceea ce admitea o greşeală la 103 cazuri. Din aceste motive, ipoteza că proteinele ar putea juca rolul de "molecule ereditare" nu a putut fi acceptată. În consecinţă, atenţia cercetătorilor a fost, astfel, canalizată spre o altă componentă celulară - acizii nucleici.
Fig.17 Structura chimică a moleculelor de valină (a), alanină (b) şi izoleucină (c)
Asemenea proteinelor, macromoleculele de acizi nucleici sunt alcătuite dintr-un număr
mare de subunităţi, denumite nucleotide. Acestea stau la baza a două categorii de acizi nucleici - ADN-ul (prin-cipalul constituient al cromosomilor) şi ARN-ul (mai exact acizii ribonucleici).
Iniţial se credea că acizii ribonucleici sunt specifici plantelor şi cel dezoxiribonucleic animalelor.
Ulterior, prin modernizarea metodelor de extracţie şi de caracterizare a acizilor nucleici, s-a constatat că toate celulele (bacteriene, vegetale şi animale) îi conţin pe ambii. Aşa cum am precizat, ambele tipuri de acizi nucleici au drept constituienţi de bază nucleotidele. Fiecare nucleotidă este alcătuită dintr-un radical fosfat (radical al acidului ortofosforic) legat la C5 al moleculei de zahăr (pen-toză), legată, la rându-i, de molecula unei baze azotate (purinică sau
pirimidinică). Purina are structură dublu inelară, iar pirimidina are un singur inel (un singur ciclu) (Fig.
18). Deoarece conţin dezoxiriboză, nucleotidele din ADN sunt denumite dezoxiribo-
nucleotide, iar cele din ARN sunt denumite ribonucleotide. Atât moleculele ADN, cât şi cele de
ARN au câte două nucleotide purinice şi câte două nucleotide pirimidinice, în proporţii variabile. Purinele sunt aceleaşi, atât pentru ADN cât şi pentru ARN (Adenină şi Guanină). In ceea ce priveşte pirimidinele, se cuvin precizate următoarele: Citozina se găseşte în ambii acizi nucleici, în
timp ce Timina se găseşte doar în ADN, fiind înlocuită în ARN prin Uracil (foarte asemănător, ca structură) (Fig. 19).
Fig. 18 Structura purinei şi pirimidinei
In ambii acizi nucleici, nucleotidele sunt asamblate prin legături covalente - se leagă
grupele fosfat ale unei nucleotide, la gruparea hidroxil a zahărului de la nucleotida adiacentă. Aceste legături, numite legături fosfodiester, erau cunoscute ca existând, dar detaliile (atomii care au legat împreună nucleotidele pentru a forma catena de acid nucleic) au rămas multă vreme necunoscute. De asemenea, iniţial, era confuzie cu privire la adevărata mărime a ADN-ului şi a ARN-ului în celule.
Fig. 19 Structura chimică a adeninei, guaninei, citozinei, timinei şi uracilului
Tehnicile utilizate pentru separarea acizilor nucleici degradau puternic macromoleculele,
fapt pentru care doar pe la mijlocul anului 1930 s-a demonstrat că ambii acizi pot apărea într-o multitudine de forme. Cruciale au fost şi experimentele efectuate de Signer şi Casperson, care au
constatat că ADN–ul separat cu atenţie, are greutăţi moleculare mai mari decât multe proteine (mai mare de 500.000 daltoni). Cei doi cercetători au dedus, de asemenea, că ADN-ul este o moleculă foarte alungită - aserţiune complet confirmată prin 1950, cu ajutorul microscopiei electronice, care a relevat faptul că ADN-ul este o moleculă extraordinar de subţire, cu un diametru de numai 20 Å.
Lungimea unei molecule de ADN este de multe mii de angstromi, confirmându-se, astfel, că este alcătuită din multe mii de nu-cleotide (Fig. 20).
Fig. 20 Aspectul electronomicroscopic al unei molecule ADN
În 1928, microbiologul englez Frederick Griffith a efectuat un experiment crucial pentru
genetica moleculară de mai târziu. In linii mari, experimentul a decurs astfel. Griffith a omorât pneumococii virulenţi, de tip S, prin şocuri de temperatură iar resturile acestora, împreună cu pneumococi nevirulenţi de tip R, au fost injectate la un lot de şoareci. Cu surprindere, Griffith a constatat că şoarecii s-au îmbolnăvit de pneumonie şi au murit. Din şoarecii morţi, cercetătorul a izolat pneumococi virulenţi de tip S. Deci, dintr-o anumită cauză, cauză ce se găsea în resturile pneumococilor virulenţi distruşi prin temperaturi ridicate, o parte dintre pneumococii nevirulenţi s-au transformat în pneumococi virulenţi. Altfel spus, componente genetice ale pneumococilor S au trecut în pneumococii de tip R şi i-au transformat. In plus, respectivele
componente genetice trebuiau să fie rezistente la temperatură şi să nu fie foarte mari, pentru a putea penetra prin membranele celulare ale pneumococilor vii, nevirulenţi, unde puteau să ia parte la procese recombinaţionale. Acestea erau, logic, presupunerile. Evident, la acea dată, ţinând cont de stadiul cunoştinţelor, nu se putea da o explicaţie adecvată fenomenului (Fig. 21).
Cercetările ulterioare au confirmat interpretarea dată acestui fenomen. Patogenitatea reflectă acţiunea genelor S (smooth, în limba engleză înseamnă neted) care determină sinteza
unei enzime implicate în elaborarea capsulei (conţinând carbohidraţi) ce înconjoară bacteriile virulente. Când este prezentă alela R (rough, în limba engleză înseamnă rugos, aspru) a acestei gene, nu se formează capsulă şi respectivele bacterii nu sunt patogene.
Câţiva ani după ce Griffith şi-a făcut publice observaţiile, extractele pe bază de bacterii
omorâte s-au dovedit a fi capabile de a induce (determina) transformări ereditare. S-au putut iniţia cercetări pentru identificarea chimică a agentului transformant. Dar, la acea dată, marea majoritate a biochi-miştilor, încă mai credea că genele sunt de natură proteică.
De aceea, surpriza a fost extraordinară, în 1944, când s-a anunţat de către Oswald T. Avery, Colin M. McLeod şi Maclyn McCarty (de la Rockefeller Institute, New York) că agentul genetic activ este ADN-ul. Cei trei au făcut această precizare după o cercetare asiduă de peste
10 ani. Punctul central al cercetărilor lor, a fost acela că au reuşit să se demonstreze că
activitatea transformantă a principiului activ, purificat, este anihilată de către dezoxiribonucleaza pancreatică. Aceasta este, de fapt, o enzimă ce degradează, cu înaltă
specificitate, moleculele de ADN, până la nucleotide şi nu are nici un fel de acţiune asupra integrităţii moleculelor proteice, sau chiar asu-pra celor de ARN. Pe de altă parte, adăugarea de ribonuclează pancreatică sau de enzime proteolitice, nu a avut influenţă asupra principiului activ, transformant.
Mulţi s-au îndoit de valabilitatea universală a descoperirii lui Avery, fapt pentru care rezultatele sale au fost acceptate şi apreciate corespunzător, mai târziu.
Fig. 21 Schema experimentului realizat de Griffith
6.1.2. Acizii nucleici – componentă ubiquitară şi permanentă a cromosomilor
O confirmare, deosebit de importantă, a acestei descoperiri a survenit după ce s-a stabilit
compoziţia chimică a cromosomilor. S-a constatat, iniţial, că ADN-ul este localizat cu prioritate în nucleul celular, apreciindu-se că este absent acolo unde lipsesc cromosomii. Apoi s-au derulat cercetările efec-tuate, tot la Institutul Rockefeller, de către Mirsky şi Ris, precum şi cele realizate, în Franţa, de către Vendrelys, cercetări prin care s-a demonstrat constanţa cantităţii de ADN (per nucleu celular). Pentru indivizii din cadrul unei specii, cantitatea de ADN este dublă în celulele somatice şi simplă în celulele haploide.
6.1. 3. Stabilitatea metabolică a ADN–ului
Un alt argument forte, pentru considerarea ADN-ului ca macromoleculă cu rol informaţional,
a fost furnizat de descoperirea stabilităţii sale metabolice. El nici nu se asamblează, dar nici nu se distruge rapid, asemenea multor altor molecule din celulele animale şi vegetale. Atomii odată încorporaţi în ADN nu sunt scoşi decât atunci când celula se degradează, când încetează de a mai fi o entitate vie.
Confirmări indubitabile ale celor precizate anterior sunt furnizate şi de analizele efectuate pe material viral dar, mai ales, de experimentele prin care s-a elucidat cauzalitatea infecţiilor virale.
Începând cu 1950, a fost posibilă separarea virusurilor în cele două componente - capsida proteică şi „miezul” nucleic. Apoi, injectând separat câte unul dintre cele două componente în
diverse organisme vii, s-a constatat că doar componenta nucleică (acidul nucleic viral) este aptă de a declanşa infecţia tipică, în timp ce capsida proteică („fantoma fagică”) nu are calităţi infectante. Concluzia a fost clară - purtătorul informaţiei ereditare este acidul nucleic. Cu această ocazie, prin analizele efectuate, s-a ajuns şi la o altă concluzie foarte importantă şi anume: toţi viruşii conţin acizi nucleici.
S-a studiat, prin metoda izotopilor marcaţi, multiplicarea bacteriofagului T2 - un fag bacterian care conţine un miez din ADN şi un înveliş alcătuit prin agregarea mai multor molecule proteice diferite. Experimentele au fost efectuate de către Hershey şi Chase, la Cold Spring Harbor, din Long Island (Fig. 22).
Fig. 22 Schema experimentului efectuat de Hershey şi Chase
Proteinele au fost marcate cu 35S şi ADN - ul cu 32P. Virusul marcat a fost utilizat pentru
infecţie şi lăsat să se multiplice în celulele gazdă. Analizele efectuate ulterior nu au detectat, în progeni, proteină marcată ci numai ADN marcat. În plus s-a constatat că învelişul proteic, marcat radioactiv, rămânea ataşat de suprafaţa celulei gazdă. Dacă, prin agitare puternică, fantomele fagice (învelişul proteic) ataşate la suprafaţa celulei gazdă erau îndepărtate, procesul de infecţie nu era afectat cu nimic. Prin aceasta s-a demonstrat, fără echivoc, rolul determinant al acidului nucleic în procesul infecţiei.
Ulterior, prin perfecţionarea tehnicilor de purificare a ADN-ului, a fost posibilă extragerea şi caracterizarea acidului dezoxiribonucleic pur, din virusul polioma prelevat din şoareci. Injectându-se doar acest acid dezoxiribonucleic, la şoareci sănătoşi, s-a determinat apariţia bolii. Din şoarecii astfel îmbolnăviţi s-au putut, apoi, extrage mii de exemplare de viruşi polioma (neoformaţi).
Pe baza tuturor acestor observaţii (completate, în timp, cu numeroase altele) s-a conchis că rolul învelişului proteic viral este acela de a proteja “miezul nucleic”, deţinător al informaţiei virale specifice (concomitent confirmându-se lipsa oricărei încărcături semantice a învelişului proteic ).
6.1.4. Principiile lui Chargraff
Utilizând cromatografia pe hârtie, biochimistul Erwin Chargraff (de la Universitatea Columbia din New York), a analizat componenţa nucleotidică a macromoleculelor de ADN. Investigaţiile sale au acreditat ideea că cele patru nucleotide (Adenină, Guanină, Citozină şi Timină) nu sunt prezente în cantităţi egale (în celulele prelevate de la indivizi aparţinând la specii
diferite) şi că procentul lor variază de la o specie la alta. Aceste constatări au sugerat (idee demonstrată, ulterior, ca valabilă) că aranjarea (ordinea,
succesiunea) nucleotidelor în macromoleculele acidului dezoxiribonucleic, determină specificitatea informaţională a acestuia. Prin urmare, proporţia celor patru nucleotide (în fond, a celor patru baze azotate) în macromolecula acidului dezoxiribonucleic, nu are caracter aleatoriu. În toate probele analizate, numărul de resturi adeninice a fost egal cu numărul de resturi timidinice, numărul de resturi guanidinice fiind egal cu numărul de resturi citozinice. Schematic, aceste constatări pot fi redate astfe:l
A=T şi G=C, de unde A+G = T + C, deci A + G / T + C = 1
Această axiomă este cunoscută, în genetică, sub numele de REGULA CHARGRAFF.
Investigaţii atente au fost efectuate prin difracţia razelor X pe acidul dezoxiribonucleic. Cer-cetările au debutat în 1938 şi i-au aparţinut lui Astbury, care a analizat eşantioane de acid dezoxiribonucleic care i-au fost puse la dispoziţie de către Hammarsten şi Casperson. Peste aproximativ 15 ani, Wilkins şi Franklin, care lucrau la King‟s College din Londra, au obţinut
fotografii de bună calitate. Wilkins şi Franklin au confirmat structura elicoidală a acidului dezoxiribonucleic şi că molecula sa este alcătuită din mai mult decât un lanţ polinucleotidic. Nu s-a putut stabili, la acea vreme, dacă este vorba de asamblarea a două sau a trei lanţuri polinucleotidice.
Aproximativ în aceeaşi perioadă (mai precis în 1952), un alt grup de cercetători, din la-boratorul lui Todd (Cambridge – Anglia) a stabilit că nucleotidele dintr-un lanţ sunt legate între ele
prin punţi fosfodiesterice formate între C5‟ al pentozei dintr-o nucleotidă şi C3‟ al pentozei din nucleotida adiacentă (Fig. 23).
Fig. 23 Structura nucleotidică a ADN-ului monocatenar
6.1.5. Argumente pentru conformaţia dublu elicoidală a ADN-ului
Între timp, Linus Pauling a propus ipoteza unei conformaţii -elicoidale, care a stârnit un imens interes. În consecinţă, Cochran, Crick şi Vand au elaborat teoria conformaţiei elicoidale
(pe baza rezultatelor obţinute prin difracţia razelor X). Teoria finală, în conformitate cu care macromolecula acidului dezoxiribonucleic are o conformaţie dublu elicoidală, cu două catene care se răsucesc elicoidal în jurul unui ax comun (ax ipotetic) a fost elaborată în 1953 de către Watson şi Crick (care lucrau în laboratorul condus de Perutz şi Kendrew, din Londra). Watson şi Crick au reuşit să dea vigoare şi eleganţă teoriei conformaţiei dublu elicoidale , folosind ca argument configuraţia stereochimică stabilită de grupul de cercetători de la King‟s College, pe baza difracţiei razelor X.
În ansamblu, s-a precizat că cele două catene ale ADN-ului sunt menţinute împreună, în dubla elice, prin punţi de hidrogen formate între bazele azotate complementare (punţi duble între A şi T şi punţi triple între G şi C) (Fig. 24).
Fig. 24 Macromolecula de ADN dublu-catenar
De aceea, aşa cum am precizat în rândurile anterioare, în macromolecula dublu elicoidală
numărul de nucleotide cu A (purină) este egal cu numărul de nucleotide cu T (pirimidină) şi numărul de nucleotide cu G (purină) va fi egal cu numărul de nucleotide cu C (pirimidină), conform principiului stabilit de Chargraff. În consecinţă secvenţele bazelor azotate din cele două catene ale unei macromolecule dublu elicoidale sunt perfect complementare. Altfel spus, ordinea bazelor de pe o catenă impune (determină) ordinea bazelor de pe catena complementară.
Concomitent cu definitivarea concepţiei structurii dublu elicoidale s-a produs şi o restructu-rare de profunzime în modul de abordare şi analiză a datelor obţinute de cei mai mulţi geneticieni. Gena a încetat să mai reprezinte o entitate misterioasă, analizabilă doar prin experimente de dihibridare. Destul de rapid gena a devenit o entitate moleculară reală, ce poate fi investigată de chimişti, geneticieni şi evoluţionişti. De acum încolo se punea problema cunoaşterii mecanismelor concrete prin care genele se pot reproduce, se transmit şi funcţionează. S-au înregistrat progrese şi în ceea ce priveşte procesul autoreproducerii acidului dezoxiribonucleic. Era tot mai evident că o catenă serveşte ca suprafaţă specifică (matriţă) pentru procesul de formare (asamblare) a catenei complementare, precondiţie pentru a înţelege cum se autoreplică o genă (Fig. 25).
Dar, pentru a accepta şi, apoi, explica ipoteza “catenei matriţă” necesară pentru asamblarea catenei complementare, s-a impus efectuarea de experimente cruciale, prin care să se realizeze sinteza de acid dezoxiribonucleic in vitro. Merite deosebite în acest domeniu îi revin
şcolii create de Arthur Kornberg care, încă din 1956, a demonstrat că este posibilă neoformarea de acid dezoxiribonucleic în extracte (amestecuri acelulare) din bacterii. Ulterior, Kornberg a demonstrat că, pentru asamblarea precursorilor ADN-ului într-o catenă unitară, este necesară o enzimă polimerizatoare specifică. Nucleotidele utilizate în respectivele experimente au fost : dATP, dGTP, dCTP şi dTTP (adică nucleotide cu bogat conţinut energetic). Enzima utilizată a fost ADN-polimeraza I, prima enzimă care s-a dovedit a fiimplicată în sinteza polinucleotidelor. Enzima este capabilă să lege nucleotidele prin punţi fosfodiesterice între C5’ şi C3’ (cei doi atomi de carbon din două pentoze aparţinând la două nucleotide adiacente) (Fig. 26).
Se impune precizarea că această enzimă este funcţională şi eficientă numai în prezenţa acidului dezoxiribonucleic. Numai în prezenţa unei macromolecule de ADN se poate asigura ordonarea celor patru nucleotide într-o anumită succesiune. De aici şi concluzia că ADN-polimeraza I recunoaşte numai porţiunea regulată zahăr-radical fosforic din cadrul unei nucleotide, fapt pentru care nu poate determina şi specificitatea secvenţei pe care o asamblează (diferenţele dintre nucleotide sunt date de bazele azotate din compoziţia lor). Demonstrarea acestei afirmaţii este relativ simplă. În orice sinteză de novo a ADN-ului, in vitro, proporţia perechilor de baze (A = T şi G = C) este similară cu cea din acidul dezoxiribonucleic care a servit ca matriţă.
Fig. 25 Autoreplicarea ADN-ului
Pe de altă parte se impune menţiunea că în orice experiment de inducere a sintezei in
vitro, în mediu acelular, nu s-a reuşit obţinerea de molecule proteice sau a altor tipuri de molecule. Prin urmare, concluzia este clară : nu există un alt compus, în afară de ADN, care să conţină informaţie genetică.
Capacitatea polimerazei I a ADN de a produce copii complementare matriţei acidului dezoxiribo-nucleic din mediul de cultură, sugerează că ea poate fi enzima majoră la Escherichia coli, implicată în asamblarea nucleotidelor pe parcursul replicării ADN. Desigur, autoreplicarea
semiconservativă a ADN-ului este un proces mult mai complex.
Fig. 26 Biosinteza enzimatică a ADN-ului, catalizată de ADN polimeraza I
6.1.6. ADN-ul se autoreplică semiconservativ
O altă problemă, este aceea a comportamentului celor două catene complementare în
procesul autoreplicării acidului dezoxiribonucleic. Se separă ele sau nu? Răspunsul a fost dat, aproape simultan (în 1958) de Kornberg, Meselson şi Stahl. Cei trei au reuşit să pună la punct metoda de separare a macromoleculelor de ADN, în funcţie de greutatea lor, într-un gradient de densitate. S-a recurs la marcarea macromoleculelor acidului dezoxiribonucleic, cu izotopi grei de
15N şi 13C. S-a reuşit să se obţină macromolecule hibride şi să se demonstreze că, în timpul
replicării, cele două catene de ADN se separă. Iată, pe scurt, cum a fost conceput experimentul. S-a preparat un mediu de cultură, pentru
bacterii, în care se găsea (în compusul azotat specific) izotopul greu 15N. Bacteriile, provenind din aceeaşi sursă, cultivate simultan pe mediul normal şi pe mediul cu 15N, vor produce acizi dezoxiribo-nucleici cu greutăţi moleculare diferite (cele crescute pe mediul normal vor produce acid dezoxiribo-nucleic cu greutate mai mică, adică normală, decât cele crescute pe mediul cu izotopul greu).
Dar, în acest context, se pune întrebarea – cele ce produc acid dezoxiribonucleic greu au molecule grele şi molecule uşoare sau molecule cu o catenă grea şi una uşoară? De răspunsul obţinut la această întrebare depindea un alt răspuns – în timpul autoreplicării ADN cele două catene se separă, sau nu? Adică autoreplicarea este conservativă sau semiconservativă?
Pentru a răspunde la aceste întrebări s-a procedat astfel : extractul de ADN s-a introdus într-o soluţie de CsCl2 şi s-a supus ultracentrifugării. În timpul centrifugării, în lungul tubului de centrifugă, se realizează densităţi diferite – mai mare spre capătul superior şi mai mică spre capătul inferior.
Metoda despre care vorbim are rolul de a “ordona” moleculele de acid dezoxireibonucleic într-un gradient de densitate, conform gradientului de densitate al clorurii de cesiu, din timpul centrifugării.
Prin transferarea bacteriilor de pe un mediu pe celălalt (de pe mediul cu 15N pe mediul cu 14N va fi posibilă obţinerea unui ADN hibrid (cu o catenă grea şi una uşoară), în situaţia în care admitem că biosinteza de noi macromolecule se realizează prin separarea celor două catene ale macromoleculei de start.
Admiţând ipoteza precizată, putem estima densitatea macromoleculelor rezultate după anumite intervale de timp, intervale în care s-au realizat una sau mai multe autoreplicări pe mediul uşor,ale ma-cromoleculelor de ADN ce fuseseră sintetizate pe mediul greu. După prima autoreplicare, fiecare ma-cromoleculă va avea o catenă grea şi una uşoară – dar toate macromoleculele vor fi la fel. După încă o autoreplicare, vor rezulta macromolecule în totalitate uşoare şi macromolecule hibride etc. (Fig. 27).
Fig. 27 Redarea schematică a experimentului prin care s-a demonstrat replicarea semiconservativă a ADN-ului
Prin urmare, aşa cum reiese din schiţa de mai sus, după prima autoreplicare cele două
macromo-lecule rezultate sunt hibride (cu o catenă grea şi una uşoară), după a doua autoreplicare rezultând două macromolecule hibride (cea de sus şi cea de jos) şi două macromolecule normale (numai cu catene uşoare – cele din mijloc, în schiţa prezentată mai sus). Este, în fapt, ceea ce au observat Kornberg, Meselson şi Stahl. Astfel s-a demonstrat că replicarea macromoleculelor de ADN este semi-conservativă – una dintre catene este conservată , rămâne intactă, în şirul generaţiilor succesive.
7.2.Transferul informaţiei ereditare de la acizii nucleici la proteine
O altă dominantă, în preocuparea geneticienilor, a fost aceea a demonstrării modalităţilor prin care informaţia deţinută în macromoleculele de acizi dezoxiribonucleici este determinantă pentru compo-nenţa proteinelor în aminoacizi. Specificitatea macromoleculelor de ADN (diferenţele dintre două macro-molecule provenind din surse diferite) este datorată secvenţei liniare a celor patru nucleotide : A, G, C şi T, din care-i sunt alcătuite catenele. Cele patru nucleotide (mai precis bazele azotate purinice şi piri-midinice din alcătuirea lor), prin repetări aleatorii, constituie un
mesaj lung, cu “semnificaţie precisă”. Cele patru nucleotide pot fi considerate, în fond, patru litere, dintr-un alfabet. Chiar şi
numai cu aceste patru litere, numărul de secvenţe potenţiale (de ADN) va fi 4N, unde N reprezintă numărul de litere (de baze, în cazul de faţă) din cuvânt (din secvenţa de ADN). Prin urmare, se poate admite că există un număr infinit de molecule de acid dezoxiribonucleic, diferite între ele prin mesajele ereditare pe care le conţin. Multe gene bacteriene conţin cam 1500 de baze azotate, într-o anumită succesiune.
Numărul potenţial de gene, de această mărime, este de 41500 – o valoare mult mai mare decât numărul de gene ce au putut exista în cromosomii ce s-au putut constitui de la apariţia vieţii, pe Terra şi până astăzi.
Prin urmare, atunci când ne referim la determinismul profund al caracterelor, ne referim la con-trolul genic al sintezei proteice, la determinismul secvenţei aminoacizilor de către secvenţa nucleotidică, dar şi la determinismul ordinii şi numărului componentelor altor biomolecule.
Primele dovezi, în acest sens, au fost furnizate de studiul hemoglobinei umane, prelevată de la indivizi suferind de anemie falciformă. La respectivii bolnavi, genele care afectează forma globulelor roşii, gene notate cu s (de la cuvântul englezesc sickle = seceră) se pot afla în stare
homozigotă (ss) sau în stare heterozigotă (Ss). Evident, starea individului este diferită în cele două cazuri. În cazul Ss individul este (fenotipic) aproape normal, anemia nefiind prea severă (globulele roşii au formă aproape normală). În cazul indivizilor ss (homozigoţie recesivă) hemoglobina este anormală (fiind caracterizată prin solubilitate diferită de cea a hemoglobinei normale). Moleculele de hemoglobină de tip normal (tipul sălbatic) sunt alcătuite din două tipuri de lanţuri peptidice – tipul alpha şi tipul beta.
Fiecare lanţ are o greutate moleculară de aproximativ 16.100 daltoni. Fiecare moleculă are două lanţuri alpha şi două lanţuri beta, fapt pentru care greutatea ei moleculară atinge 64.600 daltoni. Dar lanţurile alpha1 şi alpha2 sunt controlate de către gene distincte, fapt pentru care o singură mutaţie va afecta fie lanţul alpha1, fie lanţul alpha2, dar niciodată ambele lanţuri simultan.
Prin cercetările efectuate de Ingram (în 1957, la Cambridge), s-a constatat că hemoglobina
s (din hematiile în formă de seceră) diferă de hemoglobina normală prin schimbarea unui singur aminoacid în lanţul alpha, la poziţia 6 (acidul glutamic este înlocuit cu valina). În alte forme de anemie s-au con-statat alte înlocuiri specifice de aminoacizi.
Dar ADN-ul nu poate asigura de unul singur integrarea aminoacizilor în proteine, deşi deţine în-treaga informaţie necesară acestui proces. Altfel spus, acidul dezoxiribonucleic nu serveşte ca matriţă pentru sinteza proteinelor. Iată argumentele pentru această afirmaţie. În primul rând sinteza proteică are loc în zone celulare în care ADN-ul este absent.
Locul în care s-a stabilit că se desfăşoară sinteza proteică îl reprezintă zona citoplasmatică cu mari aglomerări de ribosomi. Or, se ştie, citoplasma este clar delimitată de carioplasmă (deţină-toarea cromosomilor – seifurile ADN-ului) prin membrana nucleară. Prin urmare, este nevoie ca infor-maţia ereditară, deţinută de ADN -ul din cromosomi, să ajungă, pe o cale sau alta, în citoplasmă, la locul sintezei proteice. În această idee s-a sugerat şi apoi s-a demonstrat, ca reală, existenţa unei alte macromolecule, capabilă să preia informaţia de la ADN şi să o transporte la locul sintezei proteice.
Suedezul Casperson şi belgianul Brachet au constatat că respectiva macromoleculă este ARN-ul, prezent în cantităţi foarte mari în citoplasma celulară. Dar, pentru a admite acest lucru, se mai impune o condiţie. ARN-ul, fiind monocatenar, înseamnă că trebuie să aibă posibilitatea de a prelua, din ADN, doar informaţia ereditară deţinută de una dintre catene. Altfel spus, atunci când nu serveşte pentru autoreplicare, una dintre catenele ADN-ului serveşte pentru asamblarea ARN-
ului. Prin urmare, trebuiau găsite anumite similitudini între compoziţia nucleotidică a unei catene de ADN şi cea a ARN-ului.
ARN-ul, asemenea ADN-ului, este alcătuit din nucleotide. Macromolecula sa este lungă
(de dimensiuni variabile), neramificată şi se formează prin legături fosfodiesterice de tipul 3‟ 5‟ (Fig. 28).
Există, totuşi, două diferenţe între moleculele de ADN şi cele de ARN. Una dintre acestea se da-torează componentei glucidice – reprezentată prin dezoxiriboză în cazul acidului dezoxiribonucleic şi prin riboză în cazul acidului ribonucleic.
O a doua deosebire, mult mai semnificativă, erste aceea că în ADN sunt prezente nucleo-tidele adenină, guanină, citozină şi timină ( A, G, C şi T ), în timp ce în ARN sunt prezente adenina, guanina, citozina şi uracilul ( A, G, C şi U ), adică timina este înlocuită cu uracil.
Dar complementaritatea purină-pirimidină (A = U şi G C) fiind asigurată,
poliribonucleotidul (ARN) este complementar cu polidezoxiribonucleotidul şi, în consecinţă, se formează structuri elicoidale complementare (catenă ADN – catenă ARN) menţinute împreună prin punţile de hidrogen menţionate anterior. ARN-ul este, în mod normal, monocatenar.
Tot ca o normalitate este şi faptul că în macromoleculele de ARN nu se asigură o rată a bazelor complementare. Deseori, cantitatea de adenină nu este egală cu cea de uracil şi
cantitatea de guanină nu este egală cu cea de citozină. Prin urmare, molecu lele de ARN nu au o structură regulată dată de punţile de hidrogen.
Fig. 28 O porţiune dintr-o moleculă de ARNm, cu punctele de start şi de oprire a translaţiei, marcate
7.2.1.Sensul circulaţiei informaţiei ereditare
La începutul deceniului şase al secolului XX (mai exact în 1953), s-a încetăţenit concepţia
în con-formitate cu care ADN-ul cromosomial serveşte drept matriţă pentru asamblare ma-cromoleculelor de ARN, macromolecule care migrează apoi spre aglomerarea de ribosomi din cito-plasmă, zonă în care are loc o intensă sinteză proteică. În 1956, Crick a formulat dogma centrală a geneticii, dogmă în conformitate cu care sensul “curgerii” informaţiei ereditare este de la ADN spre proteină, conform schemei (Fig. 29).
Din această schemă, care la ora actuală nu mai este acceptată ca dogmă, date fiind procesele de reverstranscripţie descoperite ulterior, reiese cu claritate că acidul dezoxiribonucleic
are capacitatea de autoreplicare şi că pe catena sa se asamblează macromoleculele de acizi ribonucleici care părăsesc zona nucleară (totul se petrece în interfaza ciclului celular) şi migrează în citoplasmă. De asemenea, schema sugerează faptul că ARN-ul nu poate fi niciodată sintetizat pe o matriţă proteică şi nici ADN-ul pe o matriţă de ARN.
În ceea ce priveşte sinteza ADN-ului pe o matriţă de ARN, afirmaţia a suferit unele amendamente, în sensul că uneori ARN-ul poate servi ca matriţă pentru sinteza ADN-ului (am menţionat deja existenţa reverstranscripţiei).
Fig. 29 Sensul de circulaţie al informaţiei genetice, de la AND la proteine
7.2.2. Fundamentarea ipotezei adaptorului
Nimic nu părea mai simplu, la început, decât să se accepte cămatriţa de ARN asigură
sinteza proteică pe bază de plieri care determină apariţia de cavităţi specifice fiecăruia dintre cei 20 de aminoacizi. Prin urmare, cavităţile trebuie să fie în aşa fel conformate încât să asigure potrivirea unui singur aminoacid (specificitatea cavitate – aminoacid). Ulterior, însă, Crick a început să se îndoiască de realismul acestei ipoteze. O astfel de matriţă nu-şi putea dovedi funcţionalitatea. În primul rând, grupările specifice de pe cele patru baze ale ARN-ului (A, U, G, C) au capacitatea de a interacţiona cu grupările solubile în apă. Mai mult, grupele laterale ale multor aminoacizi (cum ar fi leucina, valina, fenil-alanina) preferă mult mai mult interacţiunea cu grupele insolubile în apă (cu grupele hidrofobe). Pe de altă parte, chiar dacă unele molecule de ARN pot fi împachetate pentru a expune suprafeţe hidrofobe, nu există nici o cale pentru matriţa ARN-ului de a face deosebirea între aminoacizii foarte asemănători cum ar fi, de exemplu, glicina şi alanina, sau valina şi izoleucina (ambele cupluri diferă doar prin prezen-ţa unui singur grup CH3). De aceea, Crick a precizat că , anterior încorporării în proteină, aminoacizii trebuie ataşaţi la un
adaptor specific, adaptor care să posede o suprafaţă unică, care să se lege specific la bazele de pe matriţa ARN.
7.2.3. Sinteză proteică fără componente celulare
Sinteza proteinelor fiind descoperită şi, parţial, explicată, a fost necesară analiza unor
extracte din care să fie eliminate toate componentele celulare (presupuse a fi implicate în sinteza proteică).
Experimentul a fost imaginat şi derulat, cu succes, în 1953, la Spitalul General Massachusetts din Boston, de către Zamecnick şi colaboratorii săi. Succesul a fost asigurat de folosirea aminoacizilor marcaţi radioactiv, aminoacizi care au permis marcarea proteinelor nou sintetizate. S-a presupus că organitele în care are loc sinteza proteinelor sunt ribosomii, organit care conţin cantităţi mici de ARN.
S-a constatat, totodată, că ATP-ul este necesar ca sursă de energie pentru formarea le-găturilor peptidice. La o scurtă perioadă după aceea, Zamecnik şi Hoagland, au precizat că, înainte de a fi încorporaţi în proteine, aminoacizii sunt ataşaţi la ARNt , cu ajutorul enzimelor aminoacil sintetaze (Fig. 30).
ARNt reprezintă aproximativ 10% din cantitatea de ARN celular. Aceasta a fost o desco-perire neaşteptată, senzaţională.
Crick făcuse deja speculaţia că există un adaptor care, probabil, este un lanţ de ARN care
este capabil de a se împerechea cu moleculele de ARN care servesc drept matriţe pentru sinteza proteică.
Prin urmare, ARNt descoperit de Zamecnik şi Hoagland, nu este altceva decât adaptorul lui Crick. Adaptorul conţine baze complementare celor din ARNm – adică aici (pe ARNt) se găseşte anticodonul (nodoc –ul ) şi pe ARNm se găseşte codonul.
Fig. 30 Molecula de ARNt
7.2.4. Ribosomii nu au specificitate
Prin determinări de înaltă fineţe, s-a demonstrat că peste 85% din ARN-ul celular se gă-
seşte în ribosomi. Pe de altă parte, cantitatea de ARN creşte semnificativ în celulele cu o intensă activitate pro-teosintetică. De exemplu, cantităţile de ARN cresc semnificativ în celulele bacteriene, aflate în diviziune intensă, în celulele pancreatice, hepatice etc. Din aceste considerente, iniţial, ARNr a fost considerat ca fiind matriţa pe care se ordonează aminoacizii. Dar, analizându-se ribosomii de la E. coli, precum şi cei din alte organisme, s-a constatat că aceştia sunt compuşi din două subunităţi inegale, fiecare conţinând ARN, care se unesc sau se despart (în mod reversibi l), în funcţie de concentraţia ionică a mediului. Pe de altă parte, toate lanţurile ARNr, din interiorul subunităţilor mici, au lungime similară -–cca 1.500 baze, cum dealtfel au şi lanţurile din subunităţile mari – cca 3.000 baze. În altă ordine de idei, compoziţia ambelor categorii de lanţuri (cele mici şi cele mari) este aproximativ aceeaşi – bogăţia în G şi C, la bacterii, plante, animale, în pofida variabilităţii ratei AT / GC din ADN-ul aceluiaşi organism. Acest lucru nu ar fi posibil dacă moleculele de ARNr ar reprezenta, în fapt, o colecţie de molecule de ARN care să servească drept matriţe, molecule sintetizate de un mare număr de gene diferite. Dacă situaţia ar fi astfel, componenta bazică a ARNr ar trebui să fie puternic corelată cu aceea a genomului lor. Pe de altă parte, noi nu ne-am aştepta ca lungimea lanţurilor de ARN, care servesc drept matriţă, să fie diferită şi corelată cu mărimea produşilor polipeptidici. Rezultă că nici moleculele mici, nici cele mari, de ARNr, nu pot juca rolul de matriţă.
7.2.5. Schimbările conceptuale produse de descoperirea ARNm
Studiile efectuate pe celule bacteriene infectate cu virusul T4 s-au dovedit deosebit de utile pentru definitivarea conceptului de matriţă. Ulterior infecţiei virale, sinteza ARN-ului specific gazdei în-cetează şi se sintetizează doar ARN-ul în care este transcris ADN-ul specific virusului. Acest ARN, care este de fapt un ARNm, nu formează (nu se asamblează în) ribosomi, ci se ataşează la ribosomii exis-tenţi, ribosomi pe care glisează pînă ce nucleotidele (bazele azotate) sale ajung în poziţiile în care permit legarea complexelor ARNt – AA, în vederea sintezei proteinelor.
Deoarece acest ARN poartă informaţia de la ADN spre ribosomi (locul în care are loc sinteza pro-teică) a primit numele de ARNm (acid ribonucleic mesager).
Această descoperire, făcută în 1960, a fost urmată de multe altele care au demonstrat că există, în realitate, o importantă categorie de ARN şi anume ARN-ul mesager. ARN-ul ribosomal, împreună cu aproximativ 50 proteine diferite (proteine ribosomale, evident, legate de ARN-ul ribosomal), se comportă asemenea unui complex de sinteză a proteinelor, funcţionând pentru a pune în contact componentele complexului ARNt – AA (aminoacizi), în poziţiile în care pot descifra informaţia deţinută de ARNm. Determinări precise au stabilit că numai 4% din cantitatea de ARN dintr-o celulă revine ARNm. Pe de altă parte, acest tip de ARN etalează o variabilitate suficient de mare în ceea ce priveşte lungimea moleculei, în strânsă dependenţă de gena pe care o copie şi de molecula proteică a cărei sinteză o codifică. Întrucât numai un mic fragment din macromolecula de ARNm este ataşat la un ribosom, care glisează pe lungimea sa, doar o singură moleculă de ARN poate fi citită, la un moment dat, simultan de mai mulţi ribosomi. De obicei, ribosomii sunt asamblaţi în poliribosomi în număr destul de variabil – între 6 şi 50 de ribosomi.
Molecula de ARNm este citită şi tradusă, întotdeauna, în aceeaşi direcţie – ribosomii gli-sează de la capătul 5, spre capătul 3,. Prin urmare la ribosomi se ataşează, la început, capătul 5, al macromoleculei de ARNm.
7.2.6. Sinteza enzimatică a ARN-ului pe baza matriţei ADN-ului
După ce a fost descoperit şi caracterizat ARNm, a fost descoperită şi descrisă ş i enzima
responsabilă cu formarea ARN-ului pe matriţa de ADN. Caracterizată şi numită ARN-polimerază, de către biochimiştii americani Hurwitz, Stevens şi Weiss, această enzimă funcţionează numai în prezenţa ADN-ului (care are rolul de matriţă), determinând asamblarea nucleotideluo ATP, GTP, CTP şi UTP în monocatena de ARN.
În cazul bacteriilor, una şi aceeaşi enzimă asigură constituirea tuturor tipurilor de ARN (riboso-mal, transportor şi mesager). Dovezi directe ale faptului că matriţa de ADN dirijează asamblarea corectă a ribonucleotidelor, pentru a forma ARN-ul, sunt furnizate de variaţia compoziţiei nucleotidice a ARN-ului, în concordanţă cu rata AT / GC din macromolecula de ADN, care serveşte ca matriţă. Adică, rata AU / GC, din ARN, este similară ratei AT / GC din ADN.
În timpul transcripţiei, numai una dintre cele două catene ale ADN-ului este transcrisă în ARN. Este normal să fie aşa deoarece, dacă ARN-ul ar transcrie ambele catene, care sunt complementare, prin translaţie ar rezulta polipeptide extrem de diferite. Pe de altă parte, sinteza
ARN-ului se derulează într-o direcţie fixă, mereu aceeaşi - 5‟ 3‟. Prin urmare, transcripţia şi translaţia se derulează, mereu, în acelaşi sens. Între timp a fost evidenţiat procesul migrării ARN-ului de la ADN (care a servit, prin una dintre catenele sale, ca matriţă), prin porii membranei nucleare, spre ribosomi (spre zona citoplasmatică bogată în ribosomi). ARN-ul nou sintetizat părăseşte nucleul cam într-o oră.
7.3. Gena – definitie, structura si functie Gena, ca noţiune, a fost introdusă în ştiinţă de către Johannsen (ca şi conceptul de ge-
notip şi cel de fenotip), în 1909. Conceptul de genă (de la grecescul gennao = a naşte) a cunoscut
o evoluţie, trecând prin câteva etape, pe care vom încerca să le redăm, sintetic, în rândurile urmă-toare.
Iniţial, gena era concepută ca unitatea structurală responsabilă de determinismul fenotipizării unui caracter. Ulterior s-a constatat că sunt multe caractere, cantitative, în fenotipizarea cărora îşi aduc aportul mai multe gene (caractere cu determinism poligenic).
Atunci s-a restrâns conţinutul noţiunii, apreciindu-se că gena este responsabilă de sinteza unei
enzime. Era concepţia o genă = o enzimă. Dar o enzimă are două componente clar diferenţiate – o proteină la care este ataşată o grupare prostetică. Cele două entităţi fiind destul de
deosebite, ca structură şi funcţie, era greu să se accepte că una şi aceeaşi genă este capabilă să determine sinteza a două entităţi atât de diferite. Atunci s-a recurs la restrângerea şi mai accentuată a conceptului, considerându-se că o genă este responsabilă de determinismul sintezei unei proteine. Era conceptul o genă = o proteină. Dar, în acest caz, nu se putea explica determinismul sintezei altor biomolecule, inclusiv a acizilor ribonucleici. În consecinţă, cea mai adecvată definiţie este următoarea : gena reprezintă o secvenţă de codoni (o secvenţă de nucleotide) responsabilă de determinismul sintezei unei biomolecule. Adică, sintetic exprimându-ne, conceptuzl actual este o genă = o biomoleculă.
Cam în aceeaşi perioadă (în jurul anilor 1900) medicul englez Archibald Garrod, studiind
unele maladii metabolice umane (alkaptonuria, albinismul, cistinuria, pentozuria şi porfinuria), a emis ipoteza o genă recesivă = un blocaj metabolic. În anul 1935, Beadle şi Ephrussi, prin cer-cetări efectuate pe Drosophila melanogaster au sugerat şi, apoi, demonstrat că mutaţiile suferite de două gene implicate în determinismul culorii ochilor, duc la blocaje în două etape diferite ale sintezei pigmentului. Ulterior Beadle a experimentat, împreună cu Tatum, pe Neurospora crassa
reuşind să demonstreze, din nou, că genele exercită un control asupra proceselor enzimatice de sinteză. Dealtfel, aceste cercetări au contribuit la fundamentarea conceptului o genă = o enzimă.
Dar gena este nu numai o unitate funcţională ci şi una structurală. Şi concepţia despre structura genei a cunoscut o evoluţie, în timp. Iniţial se considera că genele sunt repartizate în cromosomi ca mărgelele pe o aţă. De aici decurgea concluzia că recombinările sunt posibile doar între gene, ca identităţi şi nu şi între subunităţi ale genelor. Concret, genele erau considerate ca unităţi structurale, funcţionale şi mutaţionale. Dar, în 1940, Oliver a adus prima dovadă că recombinarea poate fi şi intragenic, nu numai intergenic. Apoi, în 1942, Lewis a efectuat testul de
complementaritate, prin care s-a demonstrat efectul de poziţie al unor subunităţi ale genei. Adică, s-a demonstrat că dacă două subunităţi ale genei se află în poziţia cis (sunt una în continuarea
celeilalte, pe acelaşi cromosom), au cu totul alt efect fenotipic decât atunci când se află în poziţie trans (sunt pe ambii cromosomi omologi, una pe unul şi cealaltă pe celălalt dintre omologi). Descoperirea lui Lewis poartă şi numele de alelism de funcţie sau test de complementaritate.
Când două mutaţii ale aceleiaşi gene nu sunt comple-mentare în poziţia trans, concluzia este că ele afectează aceeaşi funcţie. Perin urmare, cele două subunităţi sunt repartizate în acelaşi grup de complementare. Un grup de complementare reprezintă o unitate genetică discontinuă care mai este denumită cistron. Dacă se manifestă comple-mentaritatea şi în poziţie trans, înseamnă că mutaţiile s-au produs în cistroni diferiţi. O excepţie de la această regulă se întâlneşte atunci când genele determină sinteza unor polipeptide ce constituie sub-unităţi ale proteinelor heteromultimere. În celula de tip sălbatic, proteina activă este formată din câteva subunităţi identice. Într-o celulăce conţine două alele mutante, produşii acestora pot interreacţiona, dând naştere la proteine multimere, constituite din ambele tipuri de subunităţi. Uneori, cele două mutaţii se compensează, proteina formată din subunităţile respective fiind activă, în timp ce proteinele constituite dintr-un singur tip de subunitate mutantă sunt inactive. Acest fenomen a primit denumirea de complementaritate interalelică sau intragenică.
Un alt aspect important, care trebuie să fie elucidat când se analizează caracteristicile genei, este acela al colinearităţii dintre genă şi produsul pe care-l determină. La procariote,
colinearitatea este totală. La eucariote, de obicei, genele sunt mult mai complexe şi colinearitatea nu se mai poate realiza. Este ştiut faptul că, în procesul transcripţie-translaţie, molecula de ARNm, care se sintetizează pe matriţa unei gene, prezintă o secbvenţă nucleotidică diferită de cea a genei, dar corespondentă secvenţei aminoacidice a proteinei pe care o determină. Gena conţine secvenţe adiţionale care nu se regă-sesc în molecula de ARNm. Altfel spus, gena este întreruptă. De aceea, secvenţele de ADN ce alcătuiesc o genă întreruptă, aparţin la două categorii – exoni şi introni. Exonii sunt cei copiaţi în ARNm. Intronii lipsesc din ARNm, fapt pentru care au fost considerate ca regiuni noninformaţionale. În realitate, procesul de transcripţie vizează întreaga genă. Astfel ia naştere un ARNm precursor, care suferă procese de rupere, eliminare a intronilor şi resudare a exonilor, procese ce duc la formarea de ARNm matur, care va fi translat. Mutaţiile localizate în diferiţi exoni ai unei gene nu sunt complementare.
Evidenţierea fenomenului de alelism multiplu şi de recombinare interalelică a favorizat con-cluzia că mutaţia şi recombinarea nu respectă graniţele ce delimitează o anumită funcţie a genei.
Gena are, în fapt, trei proprietăţi distincte şi anume: funcţia din lanţul metabolic celular, mutabilitatea şi capacitatea de recombinare. Seymour Benzer a propus o serie de trei termeni,
tocmai pentru a denumi şi delimita cele trei caracteristici ale genei. Astfel el a propus denumirile: MUTONUL – cel mai mic segment de material ereditar care poate suferi o mutaţie; RECONUL – cel mai mic segment genic apt de a participa la recombinare; CISTRONUL – unitatea de funcţie a materialului ereditar, definită pe baza testului de
comple-mentaritate. Aşa cum am precizat anterior, substratul material al informaţiei ereditare, deci implicit al ge-
nelor, sunt acizii nucleici – acidul dezoxiribonucleic pentru majoritatea vieţuitoarelor şi acidul ribo-nucleic pentru ribovirusuri şi viroizi. La nivel molecular, deci, atât mutonul cât şi reconul coincid cu perechea comple-mentară de nucleotide din ADN-ul dublu catenar. Prin urmare gena este constituită din mai mulţi mutoni şi, respectiv, din mai mulţi reconi. Unii autori consideră cistronul sinonim cu gena, în timp ce alţii nu le consideră similare. De exemplu, Arnold Ravin foloseşte noţiunea de genă pentru a desemna secvenţa din catena de acid nucleic care controlează producerea unei macromolecule specifice, din celulă. În a-ceastă accepţie o genă poate fi constituită din mai mulţi cistroni, fiecare dintre ei având rolul de a codifica o subunitate a proteinelor multimerice (de exemplu).
7.4. Codul genetic
În paginile anterioare s-a demonstrat, fără dubii, că informaţia ereditară este stocată în acizii nucleici, acizi responsabili şi de transmiterea ei, de la parentali la progeni, în succesiunea de generaţii. Dealtfel, în lumea specialiştilor, este stabilit faptul că informaţia ereditară, deţinută de un individ, poate circula (poate fi transmisă) pe două canale – unul în cadrul individului, în procesul fenotipizării informa-ţiei deţinute de genotip şi unul în cadrul sistemelor succedente (populaţie-specie, biocenoză), de la un individ la altul. În primul caz este vorba despre procesele de transcripţie şi translaţie, în cel de al doilea caz este vorba, în principal, despre procesul reproducerii. Că informaţia ereditară este stocată în ma-cromoleculele de acizi nucleici nu mai sunt dubii. Se pune, însă, o întrebare. Cum anume este stocată?
Nici vorbă nu poate fi despre o stocare ca atare, sub formă de caractere. Informaţia ereditară reprezintă, în ultimă analiză, un mesaj (în conformitate cu preceptele teoriei informaţiei). Mesajul, la rândul său, reprezintă o succesiune de simboluri convenţionale: litere, cuvinte, sunete, linii, puncte, cifre etc. Ca atare, mesajul informaţional este stocat şi transmis prin intermediul simbolurilor. Unitate de bază a unei simbolizări o reprezintă simbolul. Totalitatea simbolurilor aferente unui mesaj informaţional, alcătuieşte un alfabet.
Cea mai simplă relaţie dintre simbolurile unui alfabet şi un anumit conţinut informaţional este aceea în care unui eveniment, dintr-o diversitate informaţională, îi corespunde un simbol. Aceasta este o corespondenţă biunivocă. Dar, acest sistem de corespondenţă este greoi, fără
flexibilitate. Mult mai eficient este un sistem în care un eveniment, dintr-o mulţime de evenimente, este redat printr-o com-binaţie de simboluri. Este cazul alfabetelor oricăreia dintre limbile comunităţii planetare, alfabete care conţin, în medie, 30 de litere (simboluri) prin combinaţia cărora se poate forma un număr imens de cuvinte. Combinaţia de simboluri prin care se poate defini un eveniment, dintr-o diversitate de evenimente, poartă numele de cuvânt de cod. Prin conbinaţii de cuvinte se pot alcătui pro-poziţii, fraze şi, apoi, texte, în număr nelimitat. Din cele redate mai sus reiese, cu claritate, că orice in-formaţie este stocată şi, apoi, vehiculată codificat, printr-o succesiune de simboluri. Simbolurile, alfa-betul cuvintele de cod, alcătuiesc codul unui tip de informaţie. Altfel spus, codul reprezintă sistemul informaţional prin care este posibilă redarea elementelor unei anumite diversităţi.
Pentru a fi descifrată şi înţeleasă, informaţia trebuie să fie nu numai transmisă şi şi recepţionată şi tradusă, într-un sistem accesibil receptorului. De aceea, teoria informaţiei vizează şi legităţile care guvernează posibilitatea traducerii unui sistem de informaţii în altul.
De obicei, în sistemele tehnice, se foloseşte alfabetul binar, alcătuit din două litere (simbo-luri) – 0 şi 1. 0 nu reprezintă absenţa a ceva ci o alternativă la 1. De exemplu, prin 0 şi 1 putem reda stânga-dreapta, sus-jos, pozitiv-negativ etc. Succesiunea (cuvântul) 00110110, de exemplu, poate reprezenta un element dintr-o diversitate, elementul fiind specificat atât prin numărul, cât şi prin ordinea simbo-lurilor. Mai profund analizată această chestiune, rezultă că sunt posibile doar
două cuvinte alcătuite din câte un simbol fiecare, patru cuvinte alcătuite din câte două simboluri (01, 10, 00 şi 11) etc. Sunt, deci, 2n cuvinte, alcătuite din n simboluri (fiecare).
Un mesaj este alcătuit, în fond, dintr-un număr de cuvinte de cod care reprezintă părţi variabile, organizate şi specificate, din diversitatea informaţiei.
Dar cum se pot interpreta procesele genetice prin prisma teoriei informaţiei? Prezentăm doar câteva argumente, din numeroasa suită a celor existente. În fond, atât în cadrul unui sistem de nivel in-dividual (în procesul ontogeniei) cât şi între sisteme de nivel individual (între parentali şi progeni,în pro-cesul perpetuării speciilor), este vorba de transmiterea, recepţionarea şi materializarea unui mesaj infor-maţional ereditar. Experienţe multiple şi din ce în ce mai sofisticate au demonstrat că suportul material, purtătorul informaţiei ereditare este Acidul dezoxiribonucleic (sau Acidul ribonucleic în cazul ribovirusurilor şi al viroizilor).
Dar, aşa cum am precizat deja, ADN-ul nu conţine, în macromolecula sa, o succesiune de caractere, ci informaţia care determină fenotipizarea acestora, stocată codificat. Diversitatea fenotipică a speciilor de pe Terra şi, în ultimă instanţă a indivizilor unei populaţi i (specii), se realizează pe baza diversităţii proteinelor şi a altor biomolecule. Proteinele, la rândul lor, sunt constituite din cei 20 de aminoacizi existenţi în lumea vie, prin asamblarea lor în ordine şi cantităţi diferite de la o proteină la alta. Dar numărul şi ordinea aminoacizilor dintr-o proteină sunt dictate, pe anumite căi, de mesajele informaţionale deţinute de macromolecula de ADN. Cum anume se realizează această determinare, nu era posibil să se elucideze decât prin cunoaţterea şi aplicarea principiilor teoriei informaţiei. Pe aceste baze s-au născut şi au fost definite noţiunile de codon şi cod genetic.
7.4.1. Codonul
Bazele azotate care intră în compoziţia ADN-ului şi anume A, G, T şi C, constituind
nucleotidele AMP, GMP, TMP şi CMP, reprezintă literele alfabetului pentru codul genetic. Diversi-tatea proteinelor, specificată de combinaţii ale acestor patru baze. Era necesară, doar, găsirea corelaţiilor dintre respec-tivele combinaţii şi combinaţiile de aminoacizi din proteine. Altfel spus, trebuia să se demonstreze, cum patru litere ale alfabetului genetic determină o diversitate de compoziţii proteice prin asamblarea a 20 de aminoacizi. Prin urmare, codul genetic ar trebui să conţină cel puţin 20 de cuvinte, fiecare alcătuit pe baza celor patru litere şi fiecare fiind specific pentru un anumit aminoacid.
Dacă examinăm cu atenţie situaţia, constatăm că, atunci când relaţia ar fi biunivocă, adică o literă să reprezinte un cuvânt, ar rezulta doar patru cuvinte : A, G, T şi C. În acest caz, fiind vorba
de 20 aminoacizi, 16 dintre aceştia ar rămâne nespecificaţi. Următoarea şansă ar fi aceea a cuvintelor alcătuite din două litere. Aşa s-ar obţine 16 cuvinte şi anume :
UU CC AA GG UC CU AU GU UA CA AC GC UG CG AG GA
În această situaţie ar rămâne patru, dintre cei 20, aminoacizi nespecificaţi. Urmîtoarea
şansă este aceea în care, pentru un cuvânt de cod, sunt utilizate trei litere (simboluri). Astfel rezultă 64 de cuvinte de cod şi anume:
Prima nucleotidă
A doua nucleotidă A treia nucleotidă U C A G
U UUU UUC UUA UUG
UCU UCC UCA UCG
UAU UAC UAA UAG
UGU UGC UGA UGG
U C A G
C CUU CUC CUA CUG
CCU CCC CCA CCG
CAU CAC CAA CAG
CGU CGC CGA CGG
U C A G
A AUU AUC
ACU ACC
AAU AAC
AGU AGC
U C
AUA AUG
ACA ACG
AAA AAG
AGA AGG
A G
G GUU GUC GUA GUG
GCU GCC GCA GCG
GAU GAC GAA GAG
GGU GGC GGA GGG
U C A G
Dar, reamintim, în lumea vie există numai 20 de aminoacizi. Prin asocierea a trei litere într-
un cuvânt, rezultă mult mai multe cuvinte, decât ar fi necesar. Mai exact sous rezultă 64 de codoni. Această contradicţie se rezolvă, aşa cum vom vedea în continuare, prin caracterul
degenerat al codului genetic. Se mai impune, odată ajunşi la acest punct, încă o precizare. În paginile anterioare am
menţionat, continuu, că la eucariote informaţia ereditară este stocată pe macromolecula de ADN. De această dată, precizând care sunt cei 64 de codoni, am inserat întotdeauna U şi nu T, adică am
inserat o nucleotidă specifică ARN-ului şi nu una specifică ADN-ului. Atunci se naşte, în mod firesc, întrebarea – genele se află în ADN sau în ARN? Problema îşi va găsi răspunsul atunci când vor fi discutate procesele transcripţiei şi translaţiei.
7.4.2. Caracteristicile codului genetic
Am demonstrat, credem noi, că un codon nu poate fi alcătuit din mai puţin de trei litere de
cod. Totodată am precizat că s-a ajuns la o dilemă: prea mulţi codoni pentru cei 20 de aminoacizi. Această contradicţie se rezolvă prin caracteristica codului genetic de a fi degenerat. Înainte de a discuta această calitate, menţionăm toate caracteristicile codului genetic şi anume: triplet, nesuprapus, fără semne de punctuaţie între codoni, degenerat şi universal.
Că un cuvânt de cod este cu necesitate triplet, s-a demonstrat în rândurile anterioare. Ceea ce nu s-a discutat, însă, este faptul dacă un simbol (o literă de cod) aparţine la doi codoni adiacenţi sau nu. Adică se pune problema dacă codul genetic este cu acoperire sau nu. Acoperirea, la rândul ei, poate fi totală sau parţială. Dacă doi codoni adiacenţi ar avea două nucleotide în comun, acoperirea ar fi totală. Dacă doi codoni adiacenţi ar avea doar o nucleotidă în comun, acoperirea ar fi parţială. Dar, toate verificările efectuate, prin diferite metode, au demonstrat că este vorba despre un cod genetic fără acoperire, adică doi codoni adiacenţi nu au nucleotide în comun. Altfel spus, codul genetic este nesuprapus, adică doi codoni adiacenţi nu se suprapun pe două sau pe una dintre nucleotide. Mai mult decât atât, între doi codoni adiacenţi nu există una sau mai multe nucleotide spaţiatoare. Adică, codul genetic este fără semne de punctuaţie. Dar
să exemplificăm. Să presupunem că, într-o catenă de acid nucleic există următoarea secvenţă de nucleotide : CUUCCUCAUCGUAUUACUAAUAGU. În cazul codului cu acoperire totală, codonii ar fi: CUU, UUC, UCC, CCU, CUC, UCA….etc. În cazul codului cu acoperire parţială codonii ar fi : CUU, UCC, CUC, CAU….etc. Dacă între codoni ar exista semne de punctuaţie, codonii ar fi : CUU, CUC, UCG, AUU ……etc. Dar, în realitate, codonii sunt : CUU, CCU,CAU, CGU, AUU, ACU, AAU, AGU.
În concluzie, putem afirma : codul genetic este triplet, fără acoperire şi fără semne de punctuaţie între codoni. Argumente pentru codul genetic triplet, fără acoperire, au fost aduse prin
experimente de mutageneză. Dacă o nucleotidă ar aparţine la doi codoni adiacenţi, o mutaţie punctiformă care ar afecta respectiva nucleotidă, ar efecta doi codoni şi, în consecinţă, ar semnifica schimbarea a doi aminoacizi în lanţul polipeptidic. Dar, în experimentele efectuate, aşa ceva nu s-a semnalat. Dar ce se înţelege prin cod genetic degenerat? Codul genetic degenerat
presupune existenţa a două categorii de codoni. Unii dintre aceştia, mai precis 3 dintre cei 64, sunt codoni nonsens sau codoni stop, codoni care stopează procesul de translaţie al unei gene. Aceştia sunt UAA (numit ochre), UAG (numit amber) şi UGA (numit opal). Numele codonilor stop
provin de la numele unor clase de mutanţi de la bacteriofagi. Astfel, din cei 64 de codoni, au mai rămas funcţionali 61. Aceştia se caracterizează prin aceea că, mai mulţi dintre ei, pot specifica unul şi acelaşi aminoacid.
Aşa cum rezultă din tabel, unii aminoacizi sunt specificaţi de şase codoni diferiţi, alţii de cinci, patru sau trei, doi ssau numai de un codon (cum este metionina). Acesta este un subiect asupra căruia trebuie reflectat – care este semnificaţia repartiţiei atât de inegale a codonilor per aminoacid?
Degenerarea codului genetic este asigurată prin schimbarea nucleotidei din poziţia II-a sau a III-a din codon. Se pare că nucleotida din poziţia I este inamovibilă, în cadrul unui codon
Universalitatea codului genetic reprezintă o altă caracteristică a sa. Prin universalitate se înţe-lege că, indiferent de specie (microorganism, plantă sau animal), un anumit aminoacid este specificat de aceeaşi codoni. Afirmaţia se bazează pe experimente efectuate atât in vivo cât şi in vitro. De exemplu, in vitro, utilizându-se un mediu acelular alcătuit din ribosomi prelevaţi din reticulocite tinere de iepure şi ARNs de Escherichia coli, a fost posibilă sintetizarea de hemoglobină de iepure. Dacă mediul acelular este alcătuit din ribosomi extraşi din ficat de şobolan şi ARNs de Escherichia coli şi poli U, se sintetizează polifenilalanina. Într-un mediu acelular de Escherichia coli, s-a reuşit sinteza proteinei specifice capsidei virusului poliomelitei, dacă s-a introdus un ARN purificat, din virusul poliomelitei.
Dar cele mai elocvente argumente au fost furnizate de experimente în care s-a introdus ADN specific unei specii, în indivizi aparţinând altei specii, reuşindu-se sinteza proteinelor specifice primei specii. Indiferent de sursa din care este extras ADN-ul şi indiferent de gazda în care este introdus, el are capacitatea de a determina sinteze proteice specifice sursei şi, în plus, are capacitatea de a se replica semiconservativ, în gazdă. Prin urmare codul genetic este universal şi macromoleculele de ADN deţin informaţia ereditară.
7.5. Transcriptia si translatia informatiei ereditare
Am demonstrat, deci, că informaţia ereditară este stocată, codificat, în macromoleculele de ADN (cu excepţia ribovirusurilor şi a viroizilor). Dar indivizii au un fenotip în care-şi etalează trăsăturile specifice. Cum se asigură transferul de informaţie de la genotip la fenotip, prin ce mecanisme se materializează informaţia ereditară stocată, codificat, în ADN? Procesul poartă numele de translaţie şi presupune traducerea ordinii şi numărului codonilor dintr-o genă, în
ordinea şi numărul aminoacizilor dintr-o proteină. Procesul cunoaşte o succesiune de etape în care sunt implicate, pe lângă enzime specifice, macromoleculele de ADN şi macromoleculele de ARN (mai precis cele de ARNm, ARNs şi ARNr – adică acizii ribonucleici mesager, solubil sau de transfer şi cel ribosomal). Pentru a înţelege rolul fiecărei componente în acest complicat proces, este necesară elucidarea unor aspecte esenţiale ale structurării şi funcţionalităţii lor.
Reamintim că ADN-ul are structură bicatenară, fiecare catenă fiind un polinucleotid. Cele două catene, care se răsucesc elicoidal şi jurul unui ax imaginar, sunt menţinute împreună, prin intermediul punţilor de hidrogen, duble între adenină şi timină şi triple între guanină şi citozină (A = T, G = C). Pe una dintre catene, numită convenţional catena sens, sunt cantonate genele,
alcătuind grupele de linkage (o grupă de linkage = un cromosom). Cealaltă catenă poartă numele de catenă antisens. Aşa cum precizam anterior, în definiţia pe care am dat-o pentru genă, gena asigură inclusiv sinteza macromoleculelor de ARN. Evident, în cazul acestor macromolecule, procesul se petrece pe principiul complementarităţii bazelor. Intervine însă, aici, o modificare, în sensul că, în dreptul adeninei nu se mai inseră timina, ci uracilul. O altă diferenţă, în constituţia acizilor ribonucleici, faţă de ce a acidului dezoxiribonucleic, constă în aceea că, în loc de dezoxiriboză este riboza. Reamintim că acizii ribonucleici sunt de trei tipuri: acid ribonucleic mesager (ARNm), acid ribonucleic solubil sau de transfer (ARNs sau ARNt) şi acidul ribonucleic ribosomal (ARNr), cu structuri şi funcţii bine definite în procesele transcripţiei şi translaţiei informaţiei ereditare.
7.5.1. Acidul ribonucleic mesager (ARNm)
Deţine unul dintre principalele roluri în procesele transcripţiei şi translaţiei informaţiei
ereditare. Menţionam că în ADN există o catenă sens şi una antisens şi că pe catena sens se află cantonate genele. În timpul autoreplicării semiconservative a ADN-ului, anterior sintezei catenelor complementare, are loc sinteza macromoleculelor de ARN. Acidului ribonucleic mesager îi revine rolul de a copia, de a transcrie informaţia ereditară conţinută în genele de pe ADN. De fapt nu este o simplă transcriere, o transcriere literă cu literă a respectivei informaţii deoarece, aşa cum am precizat anterior, are loc o înlo-cuire (în toate poziţiile) a timinei cu uracilul.Prin urmare putem vorbi de o primă conversie, de o primă traducere, parţială este drept, a secvenţelor nucleotidice din ADN în secvenţele nucleotidice din ARN. Se pune, firesc, o problemă. Dacă informaţia ereditară este
stocată doar pe una dintre catenele ADN, for-marea ARN foloseşte ca matriţă ambele catene ale ADN, numai una dintre ele şi pe care anume?
Opiniile recente converg către concluzia că, pentru a deţine informaţia ereditară similară celei stocate pe catena sens a ADN, ţinând cont de faptul că ARNm se sintetizează pe principiul comple-mentarităţii bazelor, este obligatoriu ca el să se formeze pe catena antisens. Dacă s-ar forma pe catena sens, informaţia pe care ar deţine-o ar fi similară celei de pe catena antisens. Dovezi concludente în privinţa modului de asamblere a macromoleculei ARN, au fost furnizate de punerea în evidenţă a existenţei hibrizilor ADN-ARN. Numele de mesager i-a fost dat de laureaţii premiului Nobel, francezii Monod şi Jacob, în 1961, datorită asemănării lui puternice cu ADN-ul,
datorită marii sale instabilităţi şi, mai ales, datorită rolului specific în procesul sintezei proteice.
7.5.2. Compoziţia şi organizarea ARNm
Compoziţia nucleotidică a catenei de ARNm este complementară celei a ADN pe baza
căreia s-a asamblat. În consecinţă, compoziţia sa, ca ordine şi cantitate a diferitelor nucleotide, este extrem de va-riabilă, variabilitate legată de sursa din care se extrage, pe de o parte, cât şi de porţiunea din ADN pe care a copiat-o, pe de altă parte.
ARN-ul mesager mai este cunoscut şi sub numele de ARN informaţional, sau ARN trans-locator, deoarece are rolul de a prelua informaţia ereditară conţinută în ADN şi a o transfera în pro-teine. Aşa cum am specificat anterior şi cum reiese din tabelul 3, ARN-ul, indiferent că este mesager, transportor sau ribosomal, conţine nucleotidele A, G, C şi U, neavând niciodată T. Acest
fapt se datorea-ză modalităţii de constituire (sintetizare) a macromoleculelor de ARN, .pe matriţa ADN, respectând prin-cipiul complementarităţii bazelor, dar, complementara A fiind întotdeauna U şi nu T. Astfel, dacă catena ADN-ului va fi AAATCGTTGGACAGCATA, catena complementară de ARN va fi UUUAGCAAC-CUGUCGUAU.
Transcripţia, în cazul formării macromoleculei de ARNm, se petrece după modelul: ADN 5’
Aşa cum am precizat anterior, catena de ADN pe care este stocată, codificat, informaţ ia
eredi-tară, este catena sens (cea de sus, în schiţa anterioară), iar ARNm se formează pe catena nonin-formaţională (catena antisens sau nonsens, catena de jos, în schiţa anterioară). Reamintim că nucleo-tidele din care se constituie molecula ARN sunt ribonucleotide, adică au riboză în componenţa lor şi nu dezoxiriboză, cum au nucleotidele din componenţa ADN.
ARNm reprezintă aproximativ 3% (între 2,5% şi 5%) din totalul de ARN din celule. Greutatea moleculară a ARNm este cuprinsă între 100.000 şi 200.000 daltoni. Locul sintezei sale este, evident, nucleul celular. Asamblarea catenei de ARN, pe baza complementarităţii cu catena ADN, se realizează sub acţiunea enzimei polimeraza ARN dependentă de ADN. Deşi are o durată de viaţă destul de scurtă, pentru a-şi menţine stabilitatea structurală şi funcţională, ARNm este asociat cu o proteină protectoare, numită informozom (care-l protejează de activitatea
enzimelor degradante). S-au constatat corelaţii clare între prezenţa ARNm şi procesul sintezei proteice, atât în
medii acelulare cât şi în celule. Afirmaţia se bazează pe experimente efectuate în mediu acelular, cu un ames-tec de ribosomi, ARNs şi aminoacizi marcaţi, la care s-a adăugat şi ARNm (toate extrase din Escherichia coli). După adăugarea ARNm, viteza încorporării de aminoacizi în lanţul
polipeptidic, a crescut de peste 20 de ori. De exemplu, dacă într-un sistem ribosomal se introduce acid poliuridilic, se induce sinteza unui lanţ de polifenilalanină. S-a demonstrat, apoi, că acidul poliuridilic este un ARNm care, la o greutate mo-leculară de 105, conţine cca. 300 nucleotide şi are o lungime de aproximativ 1.000 Å, cu distanţa dintre nucleotide de 3,4 Å. Concluzia a fost că ARNm serveşte ca matriţă pentru asamblarea lanţului polipeptidic. Pe de altă parte s-a
demonstrat că, în cazul ARNm, distanţa dintre nucleotide poate ajunge chiar la 6,8 Å. Printre caracteristicile definitorii ale ARNm menţionăm:
metabolizarea rapidă în cadrul celulei;
compoziţia nucleotidică complementară uneia dintre catenele ADN, dar similară celeilalte, în cazul în care ambii acizi nucleici provin din aceeaşi sursă;
greutate moleculară extrem de heterogenă;
capacitate de a stimula sinteza de proteine, proces în care are rolul de matriţă pe care se ataşează unităţile ribosomale (formând polisomii sau poliribosomii).
Complexul poliribosomal rezistă la dezintegrarea produsă de scăderea concentraţiei de ioni de magneziu şi sedimentează, la centrifugare, mult mai rapid decât ribosomii singulari. S-a constatat, de asemenea, o corelaţie directă între mărimea complexului poliribosomal şi viteza asamblării lanţului poli-peptidic. Prin marcare cu izotopi radioactivi s-a demonstrat, fără echivoc, că lanţurile polipeptidice năs-cânde sunt legate de poliribosomi. Deoarece polisomii, reprezentând sediul sintezei proteice intra-celulare, consumă cantităţi mari de energie (pe parcursul procesului de sinteză proteică), au mai primit şi numele de ergosomi.
Bazele purinice şi pirimidinice, din macromolecula de acid ribonucleic mesager, sunt dispuse pe o singură parte a axei glucido-fosforice, perpendicular pe aceasta.
7.5.3. Compoziţia şi structura macromoleculei de ARNs
Acidul ribonucleic solubil (numit, în unele tratate, acid ribonucleic transportor - ARNt), este
mult mai puţin variabvil, atât ca număr de nucleotide cât şi, implicit, greutate moleculară, prin comparaţie cu ARNm. În medie, ARNs este alcătuit din 67 de nucleotide (între 67 şi 85 de nucleotide), şi are o masă moleculară de aproximativ 25.000 daltoni. Dar, ceea ce este mai important, acidul ribonucleic solubil poate prezenta şi o structură secundară, în sensul că nu rămâne tot timpul monocatenar. Una dintre explicaţiile apariţiei structurării secundare este aceea că bazele azotate din lungul catenei poli-nucleotidice interacţionează şi dau naştere la porţiuni dublucatenare elicoidale, alternânde cu porţiuni monocatenare. Pe de altă parte, porţiunile monocatenare nu rămân liniare ci se buclează.. Această con-formaţie explică proprietăţile aparte ale acestui acid ribonucleic. Uneori, moleculele de ARNs pot avea şi structurare terţiară. Spre exemplu, moleculele de ARNs de la Escherichia coli, precum şi cele izolate din unele drojdii (Fig. 31), au conformaţii reversibile, care se deosebesc între ele sub aspectul activităţii biologice,
comportamentului cromatografic, proprietăţile optice şi hidrodinamice etc. Rolul ARNs, în procesul sintezei proteice, este acela de a transporta acizii aminaţi la
poliribosomi. A fost descoperit în 1960, de către Allen şi Shweet, fiind unul dintre acizii ribonucleici
cel mai bine studiaţi. S-a stabilit, de pildă, că au în compoziţia lor o cantitate apreciabilă de baze rare, cum ar fi deri-vaţii metilaţi de substituţie : pseudouridina, riboziltimina, N-6-metiladenozina etc. Se apreciază că aceste baze rare contribuie decisiv la asigurarea conformaţiei tridimensionale a ARNs, conferindu-i o mai mare rezistenţă la acţiunea hidrolitică a nucleazelor celulare. Structura primară a acidului ribonucleic trans-portor extras din drojdii, care specifică alanina, a fost stabilită de către Holley şi colaboratorii săi, în 1965. Ulterior, în 1968, s-au descris acizii ribonucleici transportori pentru valină şi serină, de către Zachau şi colaboratorii, precum şi de Başev şi
colaboratorii (în mod independent). ARNs reprezintă aproximativ 10-15% din totalul de ARN celular. Polimorfismul accentuat al
aces-tui tip de acid ribonucleic se datorează faptului că, având rolul de transportor, pentru fiecare aminoacid (din cei 20 cunoscuţi) există un ARNt specific. Diferenţele dintre moleculele de ARNt rezidă, în principal, în componenţa tripletei nucleotidice, plasată în situsul cunoscut sub numele de anticodon sau nodoc.
Fig. 31 Molecula de ARNt de la Saccharomyces cerevisiae (reconstituire)
Pe de altă parte, ca factor uniformizator, macromoleculele de ARNt au, fără excepţie, grupările terminale ale lanţurilor polinucleotidice identice: la unul dintre capete au acid guanilic (G), cu hidroxilul de la C5 esterificat, iar la celălalt capăt au adenozina, înaintea ei f iind două grupări citidilice (CCA). Sunt unele păreri în conformitate cu care gruparea C-C-A este responsabilă de activarea aminoacizilor implicaţi în procesul sintezei proteice, în ultima sa fază – translaţia informaţiei ereditare. Aminoacizii sunt ataşaţi, la macromolecula de acid ribonucleic transportor, prin legături înalt energetice, care se stabilesc între gruparea carboxil a aminoacidului şi hidroxilul terminal 3‟ al acidului ribonucleic. ARNt este activat şi legat cu aminoacidul, în două etape succesive, catalizate de aceeaşi enzimă – aminoacil-ARNt-sintetaza.
Complexul AA~AMP nu este eliberat, de către enzimă, înainte de a se derula şi cea de a doua etapă – cea a legării ARNt. Complexul AA~ARNt reprezintă substratul pentru sinteza
polipeptidică, în complexul poliribosomal.
7.5.4. Compoziţia şi structura ARNr
ARNr este sintetizat pe secvenţele moderat repetitive ale ADN-ului cunoscute şi sub
numele de ADNr. În celulele eucariotelor s-au identificat milioane de ribosomi, fiecare dintre aceştia posedând câ-teva molecule de ARNr, alături de proteine ribosomale. În interfaza diviziunii celulare, clusterii de ADNr formează o structură neregulată, ca formă, numită nucleol, a cărei funcţie este
aceea de a produce ri-bosomi. Porţiunea din cromosom car conţine ADNr este cunoscută sub numele de organizator nucleolar. Pe parcursul diviziunii mitotice nucleolii dispar şi reapar în
nucleii celulelor fiice. La microscopul electronic, masa nucleolului apare formată din particule cu diametrul de 15 – 20 nm, fapt pentru care nucleolul are aspect granulat.
Examinându-se nucleolii din oocitele amfibienilor, s-a evidenţiat existenţa unui miez fibrilar, prin a cărui dispersie se relevă prezenţa unor fibrile circulare. Examinarea acestora la microscopul electronic a dezvăluit asemănarea lor cu un pom de Crăciun (Christmas tree).
Ribosomii eucariotelor conţin patru tipuri de ARNr, trei dintre acestea fiind prezente în sub-unităţile mari, cealaltă fiind prezentă în subunitatea mică. La om, de exemplu, subunitatea ribosomală mare conţine trei molecule de ARNr cu constante de sedimentare de 28S, 5,8S şi 5S, iar subunitatea ribosomală mică conţine cea de a patra moleculă de ARNr cu constanta de sedimentare de 18S. Tipurile de ARNr 28S, 18S şi 5,8S derivă dintr-un transcript primar numit pre-ARNr, în timp ce tipul 5S este sintetizat separat, în afara nucleolului.
Nucleolul nu este doar sediul sintezei ARNr, ci şi locul de asamblare a celor două subunitpţi ribo-somale. Pe parcursul procesului de producere a moleculelor de ARN, s-a identificat prezenţa
a două tipuri de proteine care se asociază cu ele şi anume : un tip care va rămâne ataşat de moleculele ARNr, intrând în structura subunităţilor ribosomale şi un tip aparţinând nucleolului, caracterizat prin relaţii tem-porare cu intermediarii ARNr. Aceste proteine sunt indispensabile pentru prelucrarea precursorilor ARNr, deoarece, împreună cu enzimele, asigură clivajul precursorilor ARNr.
Genele care codifică ARNr 5S sunt separate total de cele care codifică celelalte tipuri de ARNr, fiind plasate în afara nucleolului. Aceste gene sunt organizate în tandemuri, în care porţiunile ce sunt transcrise alternează cu porţiuni netranscrise, porţiuni spaţiatoare, formând unităţi repetitive.
7.5.1.1. Rolul ARNt şi procesul activării aminoacizilor
Am demonstrat, în paginile anterioare, că informaţia ereditară este stocată şi perpetuată, în
şirul generaţiilor eucariote, codificat (în secvenţa nucleotidelor) la nivelul macromoleculelor de ADN.
Întrebarea care se pune este următoarea: cum are loc transferul informaţiei deţinută în ADN, în informaţia specifică proteinelor (secvenţa aminoacidică a acrestora), pe parcursul ontogenezei indivi-duale? Adică se cere elucidarea proceselor prin care informaţia codificată în secvenţă nucleotidică se ma-terializează în informaţie redată prin secvenţă de aminoacizi.
Răspunsul la această tulburătoare întrebare a fost descifrat în deceniul şase al secolului 20, de Crick. Acesta a presupus că transferul de informaţie are loc prin intermediul unei molecule
adaptoare, capabilă să traducă numărul şi ordinea nucleotidelor, în număr şi ordine a aminoacizilor. Crick era de părere că pentru fiecare aminoacid există un adaptor şi că, prin intermediul acestuia, o secvenţă spe-cifică de baze azotate poate fi translată într-o secvenţă definită de acizi aminici. La scurt timp după lan-sarea acestei prezumţii, a fost descoperit un nou tip de ARN şi anume ARNs (sau ARNt), în faza solubilă a citoplasmei. Foarte repede a fost demonstrat rolul de adaptor al acestui tip de ARN.
Apoi s-a demonstrat că ARNt are capacitatea de a recunoaşte două tipuri diferite de molecule : atât ARNm cât şi aminoacizii. Rămânea, doar, să se explice cum au loc aceste recunoaşteri.
Descifrarea structurii ARNt, înb primul rând, a sugerat că aceasta face posibilă existenţa celor două funcţii de recunoaştere. În 1975, după 7 ani de investigaţii, Holley (de la Universitatea Cornell – SUA), prezenta organizarea unei molecule de ARNt – moleculă responsabilă de transferul aminoacidului alanină la locul sintezei proteice. Respectiva moleculă are o secvenţă de 77 nucleotide, 10 dintre acestea fiind baze azotate modificate. După această descoperire au urmat altele şi a devenit evident faptul că toate macromoleculele de ARNt au câteva caracteristici comune şi anume :
dimensiuni mici, date de 73 – 93 nucleotide;
proporţie semnificativă de baze azotate neobişnuite, rezultate prin modificarea enzi-matică a celor patru baze azotate normale – U, C, A, G, după încorporarea lor în catenă;
complementaritatea secvenţei în anumite regiuni, complementaritate ce permite rea-lizarea unei structuri bicatenare între respectivele regiuni, asigurându-se o conformaţie monocatenară, alternantă cu una bicatenară, care îmbracă, în mare, aspectul unei frun-ze de trifoi;
posibilitatea de a lega un aminoacid, întotdeauna, la adenina situată la capătul 3‟-OH al catenei, regiune care poartă denumirea de braţ acceptor;
prezenţa bazelor azotate modificate la nivelul buclelor monocatenare, servesc drept situ-suri de recunoaştere pentru enzime;
capacitatea de a adopta o structură terţiară unică şi foarte bine definită (ca şi în cazul structurii terţiare a proteinelor, prin formarea de legături între diferite regiuni ale moleculei).
Caracteristicile menţionate, mai sus, reflectă participarea moleculelor de ARNs în etapele complexe ale sintezei proteice. Pe de altă parte, moleculele de ARNs trebuie să poată fi diferenţiate de către enzimele care iau parte la sinteza proteinelor, astfel încât fiecare aminoacid
să fie legat la ARNs corespunzător. ARNs are deci responsabilitatea de a citi mesajul nucleotidic
al moleculei de ARNm, asigurând astfel translarea sa în secvenţa polipeptidică. Recunoaşterea între molecula ARNs şi un anumit codon din ARNm, este acompaniată de formarea legăturilor de hidrogen între moleculele de baze azota-te complementare, existente în anticodonul din ARNs şi codonul din ARNm. Prin urmare, în timpul sin-tezei proteinelor, unul dintre momentele de o importanţă majoră este ataşarea corectă a amino-acidului potrivit la molecula ARNs potrivită.
Enzimele care cuplează aminoacidul la capătul 3‟-OH al moleculei de ARNs, prin legături cova-lente, poartă numele de aminoacil-ARNs-sintetaze. Fiecare aminoacid este recunoscut de
către o aminoacil-ARNs-sintetază specifică, capabilă să asambleze complexul AA-ARNs. Aminoacil-ARNs-sinte-tazele reprezintă un exemplu de specificitate a interacţiunii proteină-acid nucleic. Aceste enzime consti-tuie o categorie extrem de heterogenă de proteine, care par să fi evoluat din proteine ancestrale, extrem de diferite la rândul lor. Este posibil ca aceste enzime să fi apărut în stadiile timpurii ale evoluţiei, înain-tea apariţiei multor alte componente ale complexului de sintetizare a proteinelor.
Aminoacil-ARNs-sintetazele catalizează o reacţie în două etape. În prima etapă a acestui proces, desemnată prin denumirea activarea aminoacizilor,
energia deţinută de ATP este utilizată pentru formarea unui aminoacid adenilat, care va fi cuplat la enzimă. Acesta este, dealtfel, primul proces care necesită energie, din complexitatea proceselor de sinteză pro-teică. În etapa următoare, aminoacidul este transferat la molecula de ARNs. Pirofosfatul, PPi, care a fost format în prima etapă, este hidrolizat la fosfat anorganic. Datorită mecanismelor de verificare posedate de enzimă, în cazul în care se constituie un complex AA-ARNs greşit, enzima se reactivează şi hidro-lizează legătura formată între AA şi ARNs.
Părerile referitoare la molecula care este recunoscută de către ARNm, adică dacă este vorba de ARNs sau de aminoacid, au fost deosebit de contradictorii. Dovezile experimentale au fost furnizate de experimentele efectuate de către Lipmann şi colaboratorii săi, experimente în care a fost alterat (chimic) aminoacidul deja cuplat la ARNs specific (Fig. 32).
Fig. 32 Etapa recunoaşterii ARNt cu aminoacidul activat
În experimentul redat în figura anterioară, a fost utilizat ARNt la care s-a cuplat cisteina
(ARNt – cisteină). Cisteina a fost ulterior convertită la alanină şi s-a urmărit eficienţa cu care are loc încorporarea aminoacidului în catena polipeptidică. Trecerea cisteinei în alanină, după ataşarea sa la ARNt specific pentru cisteină, nu a avut nici un efect. Rezultă că secvenţa de recunoaştere a ARNt, reprezentată prin anticodonul său, a rămas nealterată şi, în consecinţă, alanina (obţinută din cisteină) va fi inserată, în lanţul polipeptidic, în locii indicaţi de anticodonul
pentru cisteină, pe baza recunoaşterii codonilor din ARNm. Se va obţine, astfel, un lanţ polipeptidic modificat în consecinţă. S-a demonstrat, astfel, fără echivoc, faptul că molecula recunoscută la nivelul ARNm este ARNt (prin intermediul sec-venţei sale anticodon) şi nu aminoacidul.
Sinteza proteinelor (procesul translaţiei propriuzise) reprezintă una dintre cele mai complexe acti-vităţi ale metabolismului celular. Complexitatea acestui proces este datorată şi faptului că, în el, sunt im-plicate numeroase componente. Astfel, în context, reamintim : diferitele tipuri de ARNt, încărcate cu aminoacizii specifici, ARNm, ARNr şi ribosomii, proteine, enzime cu funcţii specifice, cationi, GTP etc. Întrucât procesul sintezei proteice, deşi urmează în principiu aceleaşi etape, este mai complicat la euca-riotele superioare, vom prezenta principalele lui momente într-o celulă bacteriană. Cea mai importantă diferenţă, faţă de ceea ce se petrece într-o celulă eucariotă superior organizată, constă în faptul că translaţia implică un număr mult mai mic de unităţi proteice ribosomale cu funcţii specifice.
În esenţă, procesul sintezei unui lanţ polipeptidic, poate fi divizat în trei etape, delimitate în funcţie de evenimentele specifice care au loc pe parcursul lor şi anume :
- iniţierea sintezei catenei polipeptidice; - elongarea catenei polipeptidice; - terminarea sintezei. Procesul translaţiei debutează de la codonul de start, de la codonul iniţiator. Aşa cum
rezultă din figurile următoare, este deosebit de important ca ribosomul să fie corect ataşat la nivelul ARNm. Altfel translaţia nu poate debuta de la nivelul codonului iniţiator (Fig. 33).
S-a constatat că interacţiunea dintre ARNm şi ribosom are loc cu mult înainte ca acesta din urmă să fie complet format, aşa încât cuplarea se realizează, în fapt, între ARNm şi unitatea ribosomală mică. Legătura se realizează între ARNm şi respectiva subunitate, la nivelul codonului iniţiator de pe molecula ARNm, codonul AUG.
Ar fi deosebit de interesant să se stabilească modul în care unitatea ribosomală este capabilă să deceleze poziţia codonului iniţiator, între multitudinea de codoni AUG existenţi pe catena ARNm. Cercetările întreprinse în această direcţie au stabilit faptul că ARNm de la procariote conţine o secvenţă specifică de baze azotate, situată cu 5 până la 10 nucleotide înainte de codonul iniţiator AUG. Această secvenţă a primit denumirea de secvenţă Shine-Dalgarno
(după numele celor ce au descoperit-o) şi este complementară unei secvenţe nucleotidice aflată la capătul 3‟ al ARNr 16S, din subunitatea ribosomală mică:
ARNr ……….ACCUCCUUUA3’
ARNm ………GGAGGA…5’
Recunoaşterea codonului iniţiator se realizează prin interacţiunile ce au loc între aceste
secvenţe, complementare, de la nivelul ARNm şi ARNr 16S, aşa că subunitatea ribosomală mică se va putea cupla cu mesagerul. La eucariote, subunitatea ribosomală mică va recunoaşte mai întâi capătul 5‟ al ARNm, capăt “bonetat” cu metilguanozină. Ulterior, această subunitate va “scana” ARNm, până ce va întâlni o secvenţă de nucleotide care va conţine tripleta AUG, care funcţionează drept codon iniţiator. De obicei, secvenţa de nucleotide este 5‟…CCACCAUGC…3‟.
În 90% din cazuri, moleculele de ARNm de la eu-cariote au codonul iniţiator AUG, iar proteina sintetizată nu posedă metionină, în catena sa primară.
Codonul AUG nu este doar codon iniţiator ci şi unicul codon care specifică metionina, în lanţul polipeptidic. Din acest motiv metionina (formil-metionina la procariote) va fi întotdeauna primul aminoacid de la capătul N-terminal al unei catene polipeptidice. În cadrul proteinelor mature, în 50% dintre cazuri, prima metionină este îndepărtată enzimatic.
În celule se întâlnesc două tipuri de ARNt pentru metionină şi anume : ARNtiMet, care va fi
folosit la iniţierea sintezei şi ARNtMet, care va fi folosit ori de câte ori trebuie introdusă metionina în
catena polipeptidică. Un aspect foarte interesant este datorat faptului că, în mitocondrii şi în cloroplaste, iniţierea
sintezei proteice are loc cu fMet, cu toate că acest aminoacid nu este folosit şi pentru iniţierea
sintezei proteice la nivelul ribosomilor citoplasmatici ai eucariotelor. Această constatare se constituie într-un argu-ment forte al ipotezei privind originea procariotă a acestor organite. Subetapele procesului de iniţiere reclamă prezenţa unor proteine ajutătoare solubile, numite factori de iniţiere (notate IF la procariote şi eIF la eucariote). De exemplu, la eucariote, eIF4E
este factorul de iniţiere care se cuplează cu capătul 5‟ al ARNm, în vederea medierii interacţiunii între mesager şi subunitatea ribosomală mică.
Fig. 33 Formarea complexului de iniţiere (A) şi ribosomul complet asamblat (B)
Pe lângă prezenţa factorilor de iniţiere, debutul sintezei proteice este dependent şi de prezenţa GTP-ului. O moleculă de GTP este cuplată cu ARNti
Met înainte de cuplarea acestuia cu subunitatea ribosomală mică. Odată ce ARNt iniţiator este ataşat, subunitatea ribosomală mare se alătură complexului, factorii de iniţiere sunt eliberaţi, iar GTP-ul este hidrolizat. Hidroliza GTP-ului conduce, probabil, la modificarea conformaţională a ribosomului, necesară pentru iniţierea sintezei.
Fiecare ribosom, complet asamblat, va prezenta două situsuri pentru asocierea ARNt (aspect ce iese în evidenţă în figura anterioară). Aceste două situsuri, denumite situsul A (aminoacil) şi situsul P (peptidil), au roluri diferite în procesul de translaţie. Se presupune că mai există încă un situs, situsul E (exit).
Situsul A este locul în care va fi încărcat, de fiecare dată, un nou complex ARNt – AA, specificat dde codonul cantonat pe ARNm, iar situsul P va dona aminoacidul cuplat la ARNt, situat la acest nivel, catenei polipeptidice în creştere. Singurul ARNt care poate fi plasat la început, în situsul P, este ARNti
Met. Din moment ce situsul P este ocupat de ARNt iniţiator, ribosomul este apt şi disponibil
pentru a prelua un al doilea ARNt cuplat cu un aminoacid (al doilea aminoacil-ARNt), la situsul A, declanşându-se, astfel, primul pas în procesul de elongare (Fig. 34).
Înainte ca ansamblul aminoacil-ARNt să fie plasat în situsul A, acest complex va trebui să se combine cu una dintre proteinele din categoria factorilor de elongare (elongation factors, în
limba engleză), cum ar fi : EF-Tu la procariote, sau eEF1 la eucariote. Aceşti factori de elongare sunt cuplaţi, la rândul lor, cu GTP-ul.
Deşi, în principiu, oricare complex aminoacil-ARNt poate fi plasat în situsul A, încărcarea stabilă nu va putea avea loc decât pentru acel complex al cărui anticodon este complementar codonului ARNm aflat în situsul A. Odată ce complexul aminoacil-ARNt-EF-Tu-GTP s-a legat la ARNm, la nivelul unui codon, GTP-ul este hidrolizat şi complexul EF-Tu-GDP eliberat. De asemenea, complexul EF-Tu-GTP-aminoacil-ARNt, va fi regenerat.
Cea de a doua fază, a procesului de elongare, constă în formarea unei legături peptidice. între aminoacizii cuplaţi la cele două molecule ARNt, plasate la situsurile P şi A. Reacţia se realizează prin transferarea metioninei (sau N-formil metioninei, la procariote) de la ARNT iniţiator, plasat în situsul P, la aminoacidul legat la ARNt din situsul A. Reacţia este catalizată de o enzimă numită peptidil-transferază, care se găseşte în subunitatea mare a ribosomului.
Deşi multă vreme a fost considerată o proteină, peptidil transferaza s-a dovedit a fi una dintre moleculele de ARNr ce alcătuiesc, împreună cu proteinele, subunităţile ribosomale. Această constatare este în acord cu evidenţierea rolului catalitic al multor molecule de ARN. Prin urmare, peptidil transferaza este considerată, pe baza datelor experimentale obţinute până la data prezentă, o ribozimă (o moleculă ARN cu proprietăţi catalitice).
În urma formării legăturii peptidice, ARNt de la situsul A va avea anticodonul cuplat cu codonul din ARNm, iar la capătul 3‟ al moleculei va purta o dipeptidă. ARNt din situsul P va fi, în acest moment, eliberat de orice aminoacid. Această etapă a elongării presupune ejectarea ARNt descărcat din situsul P şi deplasarea ribosomului cu trei nucleotide mai departe (cu un codon), în lungul moleculei ARNm, în direcţia 3‟.
Deplasarea ribosomului reprezintă o subetapă a elongării şi a primit numele de translocare, fiind caracterizată de deplasarea dipeptidei, cuplată cu ARNt, la situsul P. Această
subetapă a reacţiei necesită intervenţia EF-G la procariote, sau a eEF2 la eucariote şi hidroliza GTP. Translocarea implică deplasarea ribosomului pe o distanţă de 1 nm. Pe parcursul fiecărui ciclu de elongare, se hidrolizează cel puţin două molecule de GTP – una sau mai multe în procesul selectării aminoacil-ARNt şi una în timpul translocaţiei.
În momentul în care peptidil-ARNt s-a mutat în P, situsul A redevine operaţional pentru încărcarea unui nou complex aminoacil-ARNt, conform informaţiei furnizate de cel de al treilea codon de pe molecula de ARNm care, prin deplasarea ribosomului, se află în dreptul situsului A. În momentul formării legăturii peptidice, ARNt de la situsul A va deveni purtătorul unei tripeptide, în timp ce ARNt de la situsul P va fi descărcat şi, apoi, ejectat. Ciclul de elongare se va putea relua, prin deplasarea ribosomului, de câte ori va fi necesar, până ce se va sintetiza întregul lanţ polipeptidic.
Când am prezentat codul genetic am precizat că trei dintre cei 64 de codoni, îndeplinesc funcţia de codoni STOP (codonii ochre, amber şi opal). Situaţia este explicabilă prin aceea că nu există ARNt care să posede anticodoni complementari acestor codoni. Când ribosomul ajunge cu situsul A în dreptul unuia dintre cei trei codoni (UAA, UAG sau UGA), elongarea se întrerupe şi catena polipeptidică, cuplată la ultimul ARNt, este eliberată.
Terminarea elongării este dependentă de intervenţia factorilor de eliberare, care inter-
acţionează cu codonii stop. Bacteriile, de pildă, posedă doi factori de eliberare – RF1 care recunoaşte codonii UAA şi UAG şi RF2 care recunoaşte codonul UGA. Celulele eucariote au un singur factor de elibe-rare – eRF (în engleză, RF =release factor). Factorii de eliberare sunt, în
prealabil, cuplaţi cu GTP, care va fi hidrolizat.Când translaţia este întreruptă, catena polipeptidică va fi decuplată de la ARNt care, alături de factorul de eliberare, va fi pus în libertate din ribosom.
Fig. 34 Iniţierea translaţiei cu formarea primei legături peptidice (C), şi elongarea (D şi E)
Ribosomul, la rândul său, se decuplează de la ARNm şi se disociază în cele două
subunităţi, conform Fig. 35. Dacă se examinează la microscopul electronic ARNm ce urmează a fi translat, se poate
observa prezenţa mai multor ribosomi, ataşaţi de el. Complexul ARNm-ribosomi (vezi figura următoare) poartă numele de poliribosom sau polisom. Fiecare ribosom se ataşează la codonul de iniţiere şi, apoi, se va deplasa, pe parcursul procesului de translaţie, codon după codon, spre capătul 3‟ al catenei de ARNm, până la primul codon STOP (Fig. 36).
Fig. 35 Terminarea translaţiei
Fig. 36 Schema (dreapta) şi electrono-microfotografia (stânga) unui complex poliribosomal
Imediat ce primul ribosom ataşat la macromolecula ARNm se găseşte la o distanţă
suficientă de codonul de iniţiere, un nou ribosom se va cupla la acest codon şi va începe, la rândul său, deplasarea în vederea translaţiei. Translaţia simultană a ARNm are efecte benefice asupra sintezei proteice celulare.
Spre deosebire de eucariote, la care transcripţia ADN cu formarea de ARNm are loc în nucleu, iar translaţia în citoplasmă, la procariote cele două procese sunt intim cup late. De fapt, la bacterii, sinteza proteinelor debutează înainte ca ARNm să fi fost complet format.
Sinteza ARNm se efectuiază în direcţia 5‟ 3‟, direcţie respectată şi de deplasarea ribosomilor, în procesul translaţiei. În momentul în care capătul 5‟ a l moleculei ARNm se eliberează de la ADN, la acesta va începe ataşarea ribosomilor, de la nivelul codonului iniţiator, având ca efect final formarea poliribosomilor. Proteinele, în stare născândă, vor avea lungimi diferite, fiind cu atât mai mari cu cât ribosomul a fost legat mai de timpuriu la ARNm, deoarece a avut timp să transleze o porţiune mai lungă din mesaj.
INTREBARI DE VERIFICARE
1. Care sunt componentele chimice ale acizilor nucleici 2. Modelul Watson-Crick de structura a ADN-ului 3. Structura si functiile genei 4. ARN-ul – structura si functii 5. Exprimarea informatiei genetice – transcriptia si translatia (sinteza) proteinelor
TEME PENTRU EXAMINAREA FINALA
1. Structura si functiile acizilor nucleici 2. Notiunea de gena 2. Exprimarea informariei genetice – transcriptia si translatia
BIBLIOGRAFIE
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K., Watson, J.D., 1994. Molecular Biology of the Cell.
Garland Publishing Inc., New York, London. Băra, I.I., 1996. Vademecum in genetică. Editura CORSON, Iaşi. Băra, I.I., 1999. Genetica. Editura CORSON, Iaşi. Brown, D.D., David, I., 1968. Specific gene amplification in oocytes. Science, 160, 272. Bucătaru, N., 1993. Genetică. Editura Universitas, Chişinău. Butnaru, G., Nicolae, I., Tămaş, E., 1999. Genetică moleculară. Editura MIRTON, Timişoara Callan, H.G., 1955. Recent work on the structure of cell nuclei. Symposium on the Fine Structure of
Cells.IUBS Publ. Series B., 21, 89. Gavrilă, L., 1986. Genetica. I. Principii de ereditate. Editura Universităţii Bucureşti. Ghiorghiţă, G., 1999. Bazele geneticii. Editura Alma Mater Bacău. Klug, W.S., Cummings, M.R., 1986. Concepts of Genetics. Merill Publishing Company. A Bell & Howell
Company, Columbus-Toronto-London-Sydney. Knippers, R., 1997. Molekulare Genetik. Georg Thieme Verlag, Stuttgart, New York. Latchman, D.S., 1998.Eukaryotic Transcription Factors. Third Edition. Academic Press, San Diego, London,
Boston, New York, Sydney, Tokyo, Toronto. Lewin, B., 1997. Genes, VI. Oxford University Press, Inc., New York, USA. Miller, O.L., Hamkalo, B.A., 1972. Visualization of RNA synthesis on chromosomes. Int. Rev .Cytol., 33,1. Moraru, I., Antohi, Şt., 1966. Introducere în genetica moleculară. Ediţia II-a. Editura Medicală, Bucureşti. Raicu, P., 1997. Genetică generală şi umană. Editura Humanitas, Bucureşti. Snustad, D.P., Simmons, M.J., Jenkins, J.B., 1997. Principles of Genetics. John Wiley & Sons, Inc., New
York, Chichester, Brisbane, Toronto, Singapore, Weinheim. Tamarin, R., 1991. Principles of Genetics. WCB (Wm. C. Brown) Publishers. Toma, O., Băra, I.I., 1996. La sorgintea vieţii – aminoacizi, proteine si acizi nucleici, Vol.1. Editura Corson,
Iaşi. Tudose, I., 1992-1993. Genetică. Editura Universităţii “Al.I.Cuza”, Iaşi. Voiculeţ, N., Puiu, L., 1997. Biologia moleculară a celulei. Grupul editorial ALL. Watson, J.D., Hopkins, N.H., Roberts, J.W., Steitz, J.A., Weiner, A.W., 1987. Molecular Biology of the Gene.
The Benjamin/Cummings Publishing Company, Inc. Menlo Park, California. Reading, Massachusetts. DonMills, Ontario Wokingham, U.K. Amsterdam. Sydney. Singapore. Tokyo. Madrid. Bogota. Santiago. San Juan.
Băra, i.i., 1973. Studiu asupra biologiei populaţiilor unor specii apomictice şi sexuate înrudite.Teză de doctorat, Facultatea de Biologie, Universitatea Bucureşti.
Băra, I.I., Ghiorghiţă, g., 1980. Din enigmele evoluţiei (Apomixia şi rolul ei în evoluţie). Editura Ştiinţifică şi Enciclopedică, Bucureşti.
Băra I., Corneanu G.(sub redacţia), Fraley l., Gottschalk W., Imreh St., Lazanyi A., Nedelcu C., Whicker W., 1989. Elemente de radiobiologie vegetală. Ed. Ceres, Bucureşti.
Băra, I.I., 1989. Reproducerea, factor al evoluţiei plantelor. Editura Academiei, Bucureşti Băra, I.I., 1999. Genetica. Editura Corson, Iaşi Beggs C., Schneider-Ziebert U., Wellmann E., 1986. UV-B Radiation and Adaptativ Mechanisms in Plants.
Stratospheric Ozone Reduction, Solar Ultraviolet Radiation and Plant Life (Edited by R. C. Worrest and M..M. Caldwell), NATO ASI Series, Vol. G.8, Springer-Verlag Berlin Heidelberg, 235-251.
Brookes P., Lawley P., 1961. The reaction of mono- and difunctional alchilating agents with nucleic acids. Biochem J, . 80, 496-502.
Buican, D., 1997. Mendel şi genetica de ieri şi de astăzi. Editura ALL, Bucureşti. Butnaru, Gallia, Nicolae,I., Tămaş, Elena, 1999. Genetică moleculară. Editura Mirton, Timişoara. Cavalli-Sforza, l.l., Bodmer, W.F.,1971. The Genetics of Human Populations. Freeman, San Francisco. Charlesworth, B., 1996. The evolution of chromosomal sex determination and dosage compensation. Curr.
Biol., 6, 149-162.
Ciobotari, Ghe. V., Băra, I.I., 2000. Frecvenţa crossing-overului între cromosomii X, la femelele hibride de Drosophila melanogaster obţinute prin încrucişarea între mutanta Muller 5 şi indivizi din trei populaţii naturale. Genetică şi Evoluţionism (sub redacţia Ion I. Băra, Csilla Iuliana Surugiu, Mirela Cîmpeanu, Marilena Maniu), Editura Corson, Iaşi, 53-78
Ciobotari, G.V., Băra, I.I., 2000. Efecte ale tratamentului cu nicotină, în diferite concentraţii, aplicate indivizilor de Drosophila melanogaster dintr-o populaţie reversmutantă. Genetică şi Evoluţionism (sub redacţia Ion I. Băra, Csilla Iuliana Surugiu, Mirela Cîmpeanu, Marilena Maniu), Editura Corson, Iaşi, 79-84.
Ciupercescu d. , 1986. Curs de Genetică si Eredopatologie, Tipo Agronomia, Cluj-Napoca. Ciupercescu, D.D., Vereskens, J., Mouras, A., Y, D., Briquet, M., Negruţiu, I., 1990. Karyotyping Melandrium
album, a dioecious plant with heteromorphic sex chromosomes. Genome, 33, 556-562. Cîmpeanu, M.M., Cîmpeanu, C.S., Băra,I.I., 2000. ADN – recombinant. Editura Corson, Iaşi. Cîmpeanu, Mirela M., Maniu, Marilena, Surugiu, Iuliana C., 2002. Genetica – metode de studiu. Editura
Corson, Iaşi. Coohill Th ., 1989. Ultraviolet Action Spectra (280 to 380 nm) and Solar Effectiveness Spectra for Higher
Plants. Photochem. Photobiol., 50 ,4 , Printed in Great Britain, 451-457. Cook, l.M., 1976. Population Genetics. Chapman and Hall, A Halsted Press Book London, John Wiley &
Sons, Inc., New York Corneanu, Mihaela, 2001. Genetica. Editura Sitech, Craiova. Creed, E.R., 1971. Ecological Genetics and Evolution. Essays in Honour of E.B. Ford.Blackwell, Oxford. Crow, J.F., Kimura, M., 1970. An Introduction to Population Genetic Theory. Harper and Row, New York. Crow, J.F., 1972. The dilemma of nearly neutral mutations :How important are they for evolution and human
welfare ? J. Heredity, 63, 306-316 Darwin, Ch., Wallace, A.R., 1958. Evolution by natural selection. Cambridge University Press Deering R., 1962. Ultraviolet radiation and nucleic acid, in Organic chemistry of life. Editors: Calvin M., Pryor
A., Freeman and Company, San Francisco, 377-382. Dellaporta, S.L., Calderon-Urrea, A, 1993. Sex determination in flowering plants. Planta Cell, 5, 1241-1251. Dobzhansky, T., 1970. Genetics of the evolutionary process. Columbia University Press, New York. Drake J., 1970. The molecular basis of Mutation, Holden Day, San Francisco, Dumitru, I.F., 2001. Noutăţi în biotehnologie. Editura ILEX, Bucureşti Dumitru, I.F., Toma, N. (coordonatori volum), 2001. Progrese în Biotehnologie. Universitatea Bucureşti,
Centrul de Cercetări în Enzimologie, Biotehnologii şi Bioanaliză. Fairbanks, D.J., Andersen, R.W., 1999. Genetics. The continuity of life. Brooks/Cole Publishing Company.
Wadsworth Publishing company. ITPR an International Thomson Publishing Company. New York, Scottsdale AZ, Singapore, Tokyo, Toronto
Falconer, D.S., 1960. Introduction to quantitative Genetics. Oliver and Boyd, Edinburgh. Fernandez, J.M., Hoeffler, J.P., 1999. Gene expression Systems. Using Nature for the Art of Expression.
Academic Press, San Diego, London, Boston, New York, Sydney, Tokyo, Toronto. Floria F., 1979. Studiu comparativ asupra comportării unor specii de plante superioare in tratamentul cu
agenţi mutagenici chimici alchilanţi, Teza de doctorat, Univ. “Al.I.Cuza”, Iaşi, 1974. Floria F., Gille E., Popescu T., 1984. Influence of the treatmens with alchilating agents and gamma rays on
the cardenolides content of Digitalis lanata, Ehrn. Rev. Roum. Biochim. 21,3, 177-180. Flondor, P., Ionescu, C. (ediţie îngrijită de), 1998. Introducere în algoritmi genetici. Editura ALL, Bucureşti. Ford, E. B., 1964. Ecological Genetics. New York, John Wiley & Sons. Ford, E.B., 1975. Ecological Genetics, 4th Edition. Chapman and Hall, London. Friedberg, E.C.,Walker, G.C., Siede, W., 1995. DNA Repair and Mutagenesis. ASM Press, Washington D.C. Fristrom, J.W., Clegg, M.T., 1988. Principles of Genetics (Second edition). Freeman & co., New York. Gardner, E., Simmons, M., Snustad, D., 1991. Principles of Genetics. John Wiley & Sons, Inc., 288-308. Gavrilă, L., 1986. Genetica principii de ereditate, vol. I. Tipografia Universităţii, Bucureşti. Ghiorghiţă, G., Toth, E., Gille, E., 1984. Some cytogenetic physiological, and biochemical effect induced by
single and combined treatments with gamma rays and ethylmethansulfonate in two-row barley and wheat. Rev. Rum. Biol., Biol. Veget., v. 29,2, 137-145.
Ghiorghiţă, G.I., 1999. Bazele geneticii.Editura Alma Mater, Bacău. Ghiorghiţă, G.I., Corneanu, G., 2002. Radiobiologie. Editura ALMA MATER, Bacău. Glick, B.R., Pasternak, J.J., 1994. Molecular Biotechnology. Principles and Applications of Recombinant
DNA. ASM Press, Washington D.C. Hartl, D., Freifelder, D., Snyder, L., 1988. Basic genetics. Johns and Bartlett Publishers, Boston. Jiang, J., Gill, B.S., 1994. Different species-specific chromosome translocations in Triticum timopheevii and
T. turgidum support the diphyletic origin of polyploid wheats. Chromosome research, Oxford & New York, vol. 2, nr. 1, 59-64.
Kihara, H., Ono, T., 1923. Cytological studies on Rumex L. Bot. Mag., 37, 84-90. Kimura, M., 1968. Evolutionary rate at the molecular level. Nature, 217, 624-626.
Kimura, M., 1968. Genetic variability maintained in a finite population due to mutational production of neutral and nearly neutral isoalleles. Genet. Res., 247, 11.
Klug, W., Cummings, M., 1986. Concepts of genetics, Second edition. Merrill Publishing Company, 177, 197-2000, 306-312.
Lewin, B., 2001. Genes VII. Oxford University Press, Oxford Malling, H., de Serres, F., 1970. Genetic effects of N-methyl N-nitrosoguanidine in Neurospora Crassa.-
“Molec. Gen. Genetics”, v. 106, 165 Manna, G., 1971. Chromosome aberrations induced by living mutagens. Natl. Acad. Sci. Sect. Biol. Sci., 1-
15. Morton, N.E., 1991. Parameters of the human genome. Proc. Natl. Acad. Sci. Usa, 88, 7474-7476 Pelling, C., Allen, T.D., 1993. Scanning electron microscopy of polytene chromosomes (I). Chromosome
research, Oxford & New York, vol. 1, nr. 4, 221-237. Raicu, P., Stoian, V., Nicolaescu, M., 1974. Mutaţiile si evoluţia. Editura Enciclopedică, Bucuresti. Raicu, P., Gorenflot, R., 1980. Cytogénétique et évolution. Editura Academiei Române, Bucureşti. Snustad, P.D., Simmons, M.J., Jenkins, J.B., 1997. Principles of Genetics.John Wiley & Sons, Inc., New
York, Chichester, Brisbane, Toronto,Singapor,Weinheim. Socaciu, C., 1996. Mutageneza chimică. Editura Genesis, Cluj-Napoca, p. 76-84. Soll, D., Rajbhandary, U.L.(edited by), 1995. tRNA – Structure, Biosynthesis and Function. ASM Press,
Washington D.C. Toma, N., Gavrilă, L., 2000. Ereditatea extranucleară. Editura Universităţii din Bucureşti. Trachsel, H. (edited by), 1991. Translation in Eukaryotes. CRC Press, Boca Raton, Ann. Arbor, Boston,
London Tudose, I., 1992-1993. Genetica vol. I, II, Ed. Univ. “Al. I. Cuza”, Iaşi.