Curs 1 NUCLEOPROTEIDE-ACIZII NUCLEICI Nucleoproteidele -subclasa de heteroproteidecare pe langa componenta de natura proteica au ca componenta de natura neproteica acizii nucleici(ADN si ARN) In functie de componenta neproteica,nucleoproteidele sunt de 2 feluri:-deoxiribonucleoproteide - ribonucleoproteide Acizii nucleici sunt biomacromolecule informationale care constituie baza moleculara a unor procese biochimice fundamentale ca:stocarea,transmiterea si exprimarea informatiei genetice ereditare a biosintezei proteinelor si a diviziunii celulare. Rolul lor in organism este: -pastreaza ,transmit si exprima caracterele ereditare -pastratorii informatiei genetice -baza moleculara a biosintezei proteinelor si enzimelor -baza moleculara pentru diferntierea si reglarea celulara -baza moleculara pentru constanta replicarii celulare Structura chimica a acizilor nucleici
Transcript
Curs 1
NUCLEOPROTEIDE-ACIZII NUCLEICINucleoproteidele-subclasa de heteroproteidecare pe langa componenta de natura proteica au
ca componenta de natura neproteica acizii nucleici(ADN si ARN)
In functie de componenta neproteica,nucleoproteidele sunt de 2 feluri:-deoxiribonucleoproteide
-ribonucleoproteide
Acizii nucleici sunt biomacromolecule informationale care constituie baza moleculara a unor procese biochimice fundamentale ca:stocarea,transmiterea si exprimarea informatiei genetice ereditare a biosintezei proteinelor si a diviziunii celulare.
Rolul lor in organism este:
-pastreaza ,transmit si exprima caracterele ereditare
-pastratorii informatiei genetice
-baza moleculara a biosintezei proteinelor si enzimelor
-baza moleculara pentru diferntierea si reglarea celulara
-baza moleculara pentru constanta replicarii celulare
Structura chimica a acizilor nucleici
Atat AND-ul cat si ARN-ul sunt de fapt,polimetride nucleotide,sunt polinucleotide.Unitatea structural de baza a ADN-ului sau ARN-ului este nucleotidul care este alcatuit din nucleozid(format din baza azotata si pentoza)si acid fosforic(radical de acid fosforic)
Schema de structura a acizilor nucleici
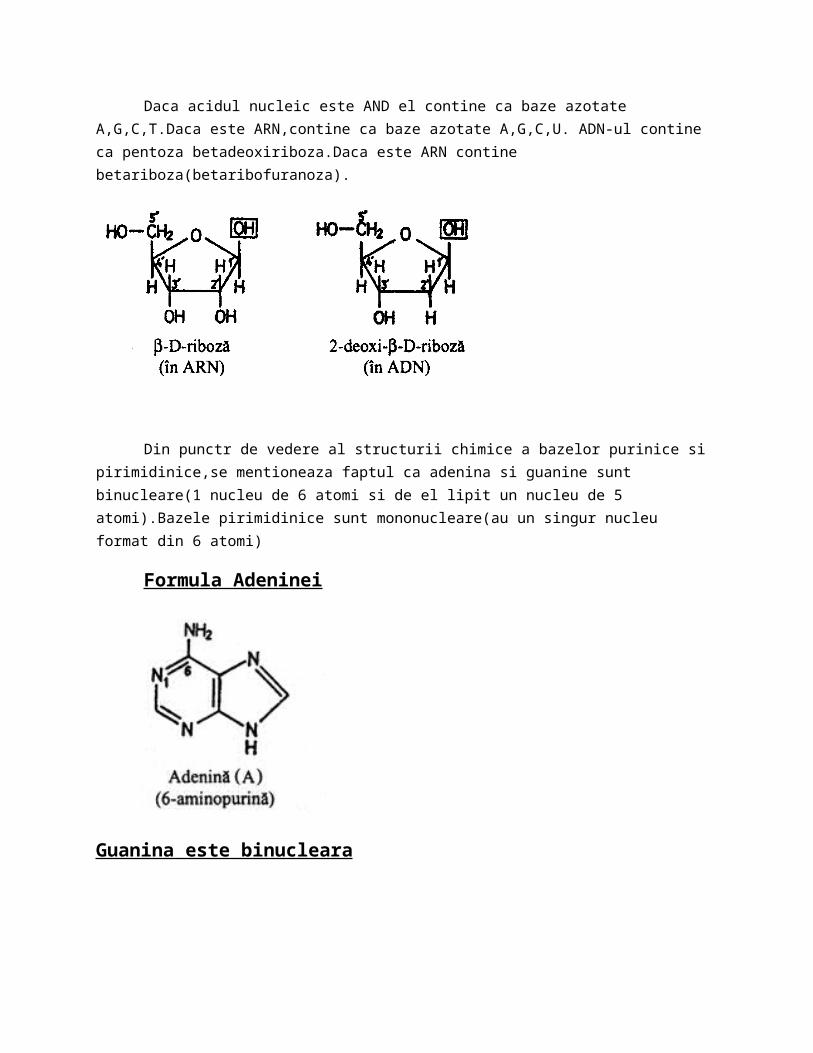
Daca acidul nucleic este AND el contine ca baze azotate A,G,C,T.Daca este ARN,contine ca baze azotate A,G,C,U. ADN-ul contine ca pentoza betadeoxiriboza.Daca este ARN contine betariboza(betaribofuranoza).
Din punctr de vedere al structurii chimice a bazelor purinice si pirimidinice,se mentioneaza faptul ca adenina si guanine sunt binucleare(1 nucleu de 6 atomi si de el lipit un nucleu de 5 atomi).Bazele pirimidinice sunt mononucleare(au un singur nucleu format din 6 atomi)
Formula Adeninei
Guanina este binucleara
In functie de natura pentozei constituent,mononucleotidele pot fi: deoxiribonucleotide(care contin betadeoxiriboza0 si ribonucleotide(betariboza).De aceasta pentoza se va lega o molecula de PO3H2 in pozitiile 3’ si 5’ ale deoxiribozei sau in pozitiile 2’-3’-5’ ale betaribozei=> esteri fosforici care in functie de numarul de grupari de PO3H2 pe care le leaga pot fi: monofosforici,difosforici sau trifosforici.
Un nucleotid esterificat la C5’ al pentozei cu 1-2-3 molecule de PO3H2 formeaza nucleozid 5 fosfati(mono-di sau trifosfati)=> un nucleozid 5 fosfat(5’) este foarmat din urmatoarele parti:
-baza azotata (A,G,C,T,U)legata de OH-ul din pozitia β a deoxiribozei sau ribozei si de aceasta mai departe in pozitia 5 sau 5’ prin eliminare de apa se leaga 1,2 sau 3 grupari de PO3H5=> nucleozidele 5 si 5’
fosfati pot fi:
ADN- d-AMP d-GMP d-CMP d-TMP
-d-ADP d-GDP d-CDP d-TDP
-d-ATP d-GTP d-CTP d-TTP
ARN-r-AMP r-GMP r-CMP r-UMP
r-ADP r-GDP r-CDP r-UDP
r-ATP r-GTP r-CTP r-UTP
Structura ATP
ATP-ul ca si di si mononucleozidele,respective ADP si AMP sunt cei mai importanti compusi din organism deoarece participa la procesele de conservare si utilizare a energiei care se elibereaza in procesele catabolice(de degradare).Sunt donori de grupari fosfat in reactiile metabolice din organismal animal=> se numesc compusi macroergici.Schema de structura a ATP,ADP si AMP este:
ATP-ul reprezinta pentru organismal animal principal rezerva energetica care ia nastere in procese de catabolism si care prin scindare furnizeaza cantitati de energie celulei ca sa traiasca,respewctiv o molecula de ATP furnizeaza celulei 8000-10.000 calorii.
Curs 2-Sructura chimica a ADN si ARN
ADN-ul are 2 tipuri de structure:
1)Structura primara este data de tipul ,de numarul si de segventa bazelor azotate care compun mononucleotidele.
-cele 4 mononucleotide care pot intra in structura primara a ADN-ului sunt:d-AMP
d-CMP
d-GMP
d-TMP
-legarea unui mononucleotid de celelalte se tralizeaza prin eliminarea de apa intre OH-ul din pozitia 3(3’) al deoxiribozei unui mononucleotide si OH-ul de la acidul fosforic de la C5(5’) trxultand astfel asa numita legatura 3-5(3’-5’) fosfodiester.
-s-a demonstrate ca intr-o molecula de AND indifferent de specie suma bazelor purinice(A+G) este egala cu cea a bazelor pirimidinice(T+C) iar rapoartele A/T si G/T sunt aproximativ egale cu 1.De aceea s-a emis ipoteza ca exista o complementaritate intre A si T( A dintr-un mononucleotide dintr-un lant de AND se va imperechea/lega prin 2 legaturi de H cu T din lantul perechie de AND) si acest lucru se simbolizeaza astfel: A::::::T iar G se imperecheaza prin legaturi de H cu C: C⁞⁞⁞⁞⁞G; apar astfel legaturi care stabilizeaza cele 2 catene antiparalele de ADN.
2) Structura secundara: ea reprezinta organizarea in spatiu,respective configuratia tridimensionala a ADN-ului.
-pe baza datelor experimentale obtinute prin metoda difractiei cu raze X,2 fizicieni americani,Watson,Crik au descoperit modelul dublu nelicoidal(dublu helix) al molecule de ADN
-acest model se caracterizeaza prin:
a)o molecula de ADN este formata din 2 catene polinucleotidice rasucite in jurul unui ax comun capatand astfel forma unei spirale orientate spre dreapta.
b)aceste 2 catene sunt orientate antiparalel;o catena este orientate pe directia 5’-3’iar alta pe 3’-5’
c)intre cele 2 catene se realizeaza multiple legaturi de H cu respectarea legii complementaritatii bazelor azotate(A-T;G-C)
d) un tur complet al dublului helix are diametrul de 34 amstroni (Å)si contine 10 perechi de baze azotate
e)datorita perchilor de baze azotate fiecare din cele 2 catene A si B dintr-o molecula de AND devin replica complementara a celeilalte.
f)rezulta deci ca succesiunea de baze azotate din cele 2 catene de ADN inmagazineaza informatia genetica care se transmite nealterata din generatie in generatie.
Localizarea si rolul AND-ului
-la procariote exista un singur ADN dublu catenar,circular, supra infasurat si care constituie defapt un cromozom circular asociat cu membrane plasmatica si acest lucru reprezinta aparatul genetic al procariotelor considerat ca fiind cel mai simplu aparat genetic din biologie.
-la eucariote aproximativ 98% din ADN se gaseste localizat in nucleul celulelor unde se gaseste asociat cu o serie de protein bazice(histonele) si acest lucru formeaza cromatina. S-au pus in evidenta si cantitati foarte mici de AND mitocondrial.
-dpdv al proprietatilor generale al ADN-ului la temperature de 80-90oC ADN-ul sufera fenomenul de denaturare deoarece se rup legaturile de H, bazele azotate nu mai sunt cuplate,creste vascozitatea si creste absorbtia in ultraviolete. Daca insa se raceste lent o molecula de ADN denaturant se constata ca cele 2 catene se pot recombine,fenomen numit renaturare,reconstituind aproape perfect structura bicatenara initiala si se pastreaza proprietatile biologice ale And-ului.
Rol
-AND-ul constituie baza moleculara a conservarii si transmiterii informatiilor genetice din generatie in generatie
-asigura si controleaza biosinteza proteinelor
-asigura diferentierea si reglarea celulara
-reprezinta baza moleculara a tuturor mutatiilor genetice naturale sau induse
Structura,localizare si rol ARN
-spre deosebire de ADN,ARN-ul are numai structura primara reprezentata de o singura catena polinucleotidica formata dintr-un numar variabil de mononucleotide
-cele 4 nucelotide care intra in structura catenei de ARN subt: AMP,CMP,GMP,UMP
Localizare
-este preponderant in citoplasma dar la organismele animale diferitele tipuri de ARN se mai pot intalnii localizate si in nuclei. La celulele bacteriene insa, ARN-ul este localizat numai in citoplasma.
Tipuri de ARN
-mesager-acea molecula care reprezinta 2-4% din ARN celular si este mesagerul informatiei genetice stocate in ARN-ul cromozomial si este cel care copiaza informatia genetica stocata in ARN si o duce la ribozomi
-ribozomal-acel tip de ARN localizat la ribozomi unde se gaseste combinat cu ribozomi si cu care formeaza asa numitele complexe ribonucleo-proteice. La aceste complexe va fi locul de biosinteza al unei niu protein.
-de transfer-acel tip de ARN care va cara aminoacizii din citoplasma,ii va duce la ribozomi si pe baza informatiei genetice adusa de ARNm ii va insiruii intr-o anumita ordine si vor forma o proteina.
Compusi macroergici
-acei compusi biochimici bogati in energie care contin legaturi fosforice macroergice in care se stocheaza energia
-cel mai important este ATP-ul
-creatin fosfatul(CP)-se mai numeste fosfocreatina si se obtine din creatina sub actiunea ATP-ului cand ia nastere creatin fosfatul cu o singura legatura macroergica. Enzima care catalizeaza aceasta reactie se numeste creatin fosfokinaza(CPK); acest compus este rezerva energetica a muschiului cardiac dar si a muschilor striati si netezi
-al 3-lea compus are de asemenea legatura macroergica si reprezinta punctual central comun al tuturor celor 3 tipuri de metabolism( glucidic, lipidic,proteic) poarta numele de acetilcoenzima A: =>
=>H3C-CO~SCoA . Se obtine prin urmatoarea reactie(examen) :
Curs 3-VitamineSunt compusi biochimici de importanta majora in organismal viu deoarece au rol determinant in
desfasurarea normal a proceselor de crestere,de dezvoltare si de multiplicare a organismului si totodata au rol hotarator in mentinera functiilor vitale.
Sunt sintetizate in general de organismele vegetale si intr-o masura mica de catre speciile animale si om.
Omul isi procura vitaminele din hrana si intr-o masura foarte mica le biosintetizeaza la nivelul tractului intestinal sub actiunea microbiana a florei din colon.
Clasificare
Criteriul major de clasificare este solubilitatealor si din acest motive le se clasifica in:
a)vitamine liposolubile(solubile in lipide)-A,D,E,K
b)vitamine hidrosolubile(solubile in apa)-vitaminele din gr B(B1,B2,B6,B16),vitamin PP(miocina),acidul folic,biotina,vitamin C,acidul paraaminobenzoic,acidul pantothenic,vitamina P,inozitolul
Caracteristicile vitaminelor
In afara de solubilitate vitaminele liposolubile se deosebesc de cele hidrosolubile prin:
-cele liposolubile se depoziteaza in ficat in timp ce cele hidrosolubile nu se depoziteaza si in momentul in care au atins un prag optim se elimina pe cale renala
-vitaminele liposolubile presupun nu numai existenta lipidelor in care se solubilizeaza dar au nevoie si de prezenta lipazei pancreatice,au nevoie de prezenta sarurilor biliare care sa emulsioneze si care sa rupa complexele lipovitaminice formate
-cele hidrosolubile se absorb usor,singurul care le distruge este acidul clorhidric din mucoasa gastrica.
-vitaminele liposolubile pot actiona sic a hormone de unde denumirea de hormono-vitamine in timp ce vitaminele hidrosolubile participa in structura unor enzime si se numesc enzimo-vitamine.
-vitaminele liposolubile participa mai mult in procese de anabolism(biosinteza) in timp ce cele hidrosolubile participa la reactii catabolice adica la acelea in care se elimina energie.
EnzimeleNotiuni generale
Totalitatea reactiilor de biosinteza/anabolism sau de degradare(catabolism),reactii care alcatuiesc defapt metabolismul oraganismului viu au loc sub actiunea unor molecule specializate care au rol de catalizatori si care poarta numele de enzime.
Enzimele sunt biocatalizatori adica o clasa speciala de molecule proteice dotate cu activitate catalitica.
Caracterisiticile enzimelor
-sunt cei mai eficienti catalizatori cunoscuti deoarece desi actioneaza la concentratii foarte mici desfasoara o activitate extreme de intense
-nu se consuma si nu se transforma in timpul reactiilor biochimice pe care le catalizeaza
-micsoreaza energia de activare a moleculelor asupra carora actioneaza(si care poarta numele de substrat)si pe care le transforma in produsi de reactie
-nu modifica starea finala de echilibru a reactiilor pe care le catalizeaza ci numai grabesc viteza de reactive cu acre se realizeaza acest echilibru.
-au specificitate de substrat(actioneaza numai asupra unui anumit substrat pe care il transforma in produsi)dar au si specificitatede reactive adica catalizeaza un anumit tip de reactive biochimica sau un grup restrains de reactii biochimice determinand astfel mecanismul de producer al acelei reactii
-unele au proprietati reglatoare(allosterice)adica decid initierea,desfasurarea si stoparea unei anumite reactii sau unui grup de reactii biochimice
Structura si conformatia enzimelor(subiect de examen)
-deoarece enzimele sunt macromolecule de natura proteica ele vor fi alcatuite dintr-un numar foarte mare de aminoacizi care vor fi dispusi intr-o configuratie spatiala determinata gentic,configuratie care va fi specifica pentru activitatea catalitica a enzimei
-fiind proteina va avea toate proprietatile fizico-chimice ale proteinelor
-ca structura exista 2 tipuri de enzime:
a)monocomponente-sunt alcatuite umai din componenta proteica care poarta numele de apoenzima sip e care se va afla centrul catalitic al enzimei denumit situs catalitic la care se va lega reversibil substratul
b)bicomponente-alcatuite din 2 parti:
1)-apoenzima care este componenta de natura proteica,termolabila si nedializabila,dotata cu activitate catalitica deoarece in structura ei se gaseste localizat situsul catalytic adica locul unde se va lega reversibil substratul de reactive.Totodata pe aponevroza se mai poate gasi in cazul enzimelor reglatoare sau allosterice un alt centru care se va numi situs allosteric la care se vor lega asa numitii efectori allosterici
-este acea componenta care stabileste legaturile cu substratul de reactie si confera specificitate de substrat enzimei
-datorita faptului ca este de natura proteica este pasibila de modificari conformationale in anumite limite permissive
2)-cofactorul-molecule de natura neproteica respective de natura chimica diferita dar sunt indispensabile pentru manifestarea activitatii catalitice a unei enzyme
-este micromolecula cu character neproteic
-este termostabila si dializabila
-este indispensabila desfasurarii activitatii catalitice a enzimei si determina atat mecanismul de reactie a enzimei cat si viteza de reactive
-nu reactioneaza decat in prezenta apoenzimei si se poate lega fie intim de aponevroza fie pot fi usor disociabile de apoenzima
-se caracterizeaza prin-induce o conformatie a molecule de enzima optima pentru atasarea lui la apoenzima,conformatie care va permite interactiunea dintre enzima si substrat
-induce o asemenea dispunere spatiala adegvata apoenzimei si substratului deoarece se va situa in vecinatatea situsului catalitic
-participa la reactia enzimatica propriu-zisa fie prin acceptare si transfer de electroni fie prin acceptare si transfer de atomi de grupari chimice de la un substrat de reactive la altul
-dupa cum se leaga de apoenzima poate fi:
-coenzime-sunt in general micromolecule organice care se leaga labil de apoenzima si la sfarsitul reactiilor enzimatice se desprind si se regasesc in stare initiala putand participa la o noua reactive de transformare a substratului in produsi de reactive
-ionimetalici-sunt ioni de fier,de zinc,de cupru,de nichel,magneziu care sunt indispensabili pentru desfasurarea activitatii catalitice a enzimei si se leaga prin legaturi covalente si covalent coordinative de apoenzima formand strat de chelat.
-grupari prostetice-sunt grupari chimice legate strans de apoenzima prin legaturi covalente
!!!!Centrul catalitic(situsul catalitic al enzimei)
-se numeste situs catalitic o zona din apoenzima la care se ataseaza in mod specific substratul de reactive pentru a fi transformat in produs de reactive
-are rol de a stabili legatura dintre enzima si substrat
-se caracterizeaza printr.o anumita configuratie spatiala astfel incat se va creea o anumita compatibilitate intre el si substratul care se ataseaza la el
-este localizat intotdeauna in portiunea interna a apoenzimei,respective in portiunea hidrofoba astfel incat sa ofere conditii favorabile de transfer de electroni,de atomi,de grupari chimice intre enzima si substratul pe care il transforma
-este alcatuit dintr-un grup de aminoacizi dispusi intr-o ordine specifica si care au anumite grupari chimice ordonate dupa cum urmeaza:SH, grupari OH,grupari NH2,grupari COOH
Imaginativ o enzima bicomponenta ar arata asa:
Curs 4Schema generala a unei reactii catalizata de o enzima bicomponenta
-orice reactie enzimatica catalizata de o enzima bicomponenta se desfasoara in 2 faze:
1) este faza in care substratul se aproprie de enzima,recunoaste situsul catalitic si se asociaza specific cu enzima formand un complex intermediar enzima substrat
E + S [ES] 2) este faza in care complexul in care enzima substrat disociaza, substratul este transformat in produsi de reactie,iar enzima se elibereaza putand cataliza o noua reactie.
[ES] P+EImaginativ procesul arata astfel:
In ceea ce priveste mecanismul prin care substratul………………..s-au emis mai multe ipoteze prin care:
-ipoteza “lacat cheie”(este ipoteza care presupune existenta unei complementaritati reciproce intre geometria moleculei de substrat si geometria situsului catalitic al enzimei)
-ipoteza “potrivirii induse”- o ipoteza care sustine existent unei flexibilitati a constatntei spatiale a situsului care se poate modifica ca sa se potriveasca la substrat
--ipoteza catalizei covalente( conform careia legarea reversibila a substratului la situsul catalitic se poate realiza numai prin stabilirea de legaturi covalente cu formarea complexului enzima substrat ca intemeietor instabil si foarte reactiv
*.Enzime Allosterice(sau reglatoare)
-sunt acele enzime care pe langa activitate catalitica exercita si un rol reglator. Aceste enzime au pe apoenzima inafara de situsul catalitic o zona numita situs allosteric la care se pot atasa diverse molecule numite efectori allosterici(modulatori allosterici). Legarea acestor modulatori la situsul allosteric este
selectiva si reversibila si moduleaza (determina) tranzitii ale moleculei de enzima intre 2 conformatii posibile pe care aceasta le poate adapta. Daca enzima poate lua aceste conformatii ea poate permite sau nu accesul sustratului la situsul catalitic astfel:
-daca la situsul allosteric se leaga o molecula care este activatoare(activator allosteric) pentru enzima atunci aceasta va lua in spatiu o conformatie favorabila astfel incat substratul sa acceada rapid la situsul catalitic si enzima sa transforme substratul in produs
-daca la situsul allosteric se fixeaza o molecula inhibitoare pentru enzima(inhibitori allostrici) atunci enzima va adopta in spatiu o asemenea configuratie incat situsul catalitic nu se va mai potrivi la substrat si acesta nu va mai fi tranformat in produsi
Imaginativ procesul arata astfel:( desenul nu este fix ca cel pe care l.a facut profa la curs)
Mecanismul de actiune al enzimelor reglatoare al enzimelor allosterice/ reglatoare este determinat de asa numitii efectori heterotrofici adica de aceste molecule care pot fi activatoare/inhibitoare..
Daca intr-un sir de reactii enzimatice care sunt catalizate de enzime obisnuite iar prima reactie
EA E2 E3
A B C D.
este catalizata de o enzima allosterica atunci produsul final al acestui sir de reactii adica D poate fi pentru enzima allosterica activator/ inhibitor astfel:
-daca celula considera ca s-a format prea mult produs D atunci produsul D se ataseaza la situsul allosteric al enzimei prime si este un efector inhibitor pentru enzima si atunci compusul A nu mai trece in B.
-daca celula vie considera ca produsul final D s-a format in cantitati prea mici atunci D se va fixa la prima enzima care este allosterica si va fi un activator allosteric pentru ea si ii va determina o asemenea configuratie spatiala incat reactia de trecere a lui A in B sa se faca foarte rapid.
Acest mecanism de actiune a compusului de reactive D asupra enzimei allosterice prime se numeste mecanism de feed-back sau retroinhibitie sau inhibitie prin produs final.
Cinetica enzimatica si factorii care influenteaza viteza unei reactii enzimatice
-se numeste cinetica enzimatica/chimica legile dupa care se conduc reactiile biochimice sub raportul vitezelor de reactive respective evolutia in timp a transformarilor caracterelor unei reactii enzimatice
-studiul cinetic al activitatilor enzimatice se bazeaza pe masurarea cantitativa a vitezei de desfasurare a reactiei catalitice.
-se stie ca viteza de reactive se noteaza cu v si poate fi variatie de substrat pe variatie de timp(v=−dSdt )
sau variatia de produs pe variatie de timp v=+dPdt
-viteza de reactive este parametrul cu ajutorul caruia se poate exprima activitatea unei enzime
-dpdv cinetic reactiile enzimatice se diferntiaza prin asa numitul ordin de reactive care se noteaza cu n si
care exprima dependenta vitezei de reactive de concentratia reactantilor : v=K[S]n
-pentru reactiile de ordinal 0 => v=K[S]0 => v=K => v= o constanta => enzima are situsul catalitic saturat cu substrat
- pentru o reactive cu n=1 => v=K[S]1
* Factorii care influenteaza viteza unei reactii enzimatice
1) concentratia substratului- s-a demonstrat ca cresterea concentratiei substratului determina cresterea vitezei de reactive pana la o valoare numita viteza maxima la care concentratia substratului ramane constanta. Jumatate din aceasta viteza maxima corespunde la o concentratie de substrat numita Km = constanta Michaelis : ea este specifica fiecarei enzime si reflecta afinitatea enzimei pentru substrat
2)concentratia enzimei- viteza unei reactii enzimatice este direct proportionala cu concentratia enzimei
3)temperatura- cresterea temperaturii determina cresterea vitezei de reactie pana la o valoare la care enzima fiind proteina se denatureaza iar viteza scade. Fiecarei enzime ii corespunde un optim de temperatura la care activitatea ei catalitica este cea mai mare.
4) influenta pH-ului-fiecarei enzime ii corespunde un pH optim de actiune la care enzima isi manifesta activitatea catalitica maxima.
5) efectorii enzimatici- acele molecule care pot fi activatoare sau inhibitoare pentru enzima(ionii de Cl.Mg,Cu,Ca sunt activatori in general pentru enzime) iar inhibitori sunt ionii metalelor grele –Pb,Cd,Hg
Curs 5-Clasificarea EnzimelorExista o comisie international de clasificare si nomenclatura a enzimelor- se numeste Comisia de
enzime aUniunii Internationale de Biochimie( IUB) care se ocupa cu identificarea fiecarei enzime printr-un numar de cod si un nume systematic.
Fiecare enzima este caracterizata de un cod de forma EC WXYZ unde EC se refera la comisia de enzime, W intotdeauna va insemna clasa enzimatica din care face parte, X sub clasa enzimatica, Y numarul substratului specific sau poate reprezenta numarul specific al coenzimei, Z reprezinta numarul de ordine/serie al enzimei respective.
* Dupa natura reactiilor pe care le catalizeaza enzimele au fost clasificate in 6 mari clase enzimatice.
1)oxidoreductaze- catalizeaza reactiile de oxidoreducere care decurg cu transformare de H , cu transformare de electroni sau cu transformare de O
2)hidrolazele- catalizeaza reactiile de rupere a moleculei de substrat sub actiunea apei
3) transferaze- catalizeaza reactiile de transfer a unor receptori chimici de la un substrat donor la un substrat acceptor
4)izomeraze- catalizeaza reactiile de izomerizare adica de rearanjare intramoleculara a atomilor sau a gruparilor functionale din molecula de substrat
5)liazele-catalizeaza reactiile de rupere/scindare a unor legaturi chimice fara participarea apei
6)ligazele(sintetazele)-catalizeaza reactiile de sinteza a unor macromolecule deci catalizeaza reactiile de formare a noi legaturi chimice,reactii care se petrec cu consum de energie deci cu participarea ATP-ului.
Oxidoreductazele
-sunt enzime care catalizeaza reactiile redox care se petrec cu:
a) transfer de H si se vor numi dehidrogenaze
b)transfer de electroni si se vor numi transelectronaze
c) transfer de O si se vor numii oxigenaze
a)Dehidrogenazele
-acea clasa de oxidoreductaze care catalizeaza reactii de oxidoreducere cu transfer de H adica au niste coenzime care joaca rolul de acceptori intermediari de H intre substrat si produsul final.
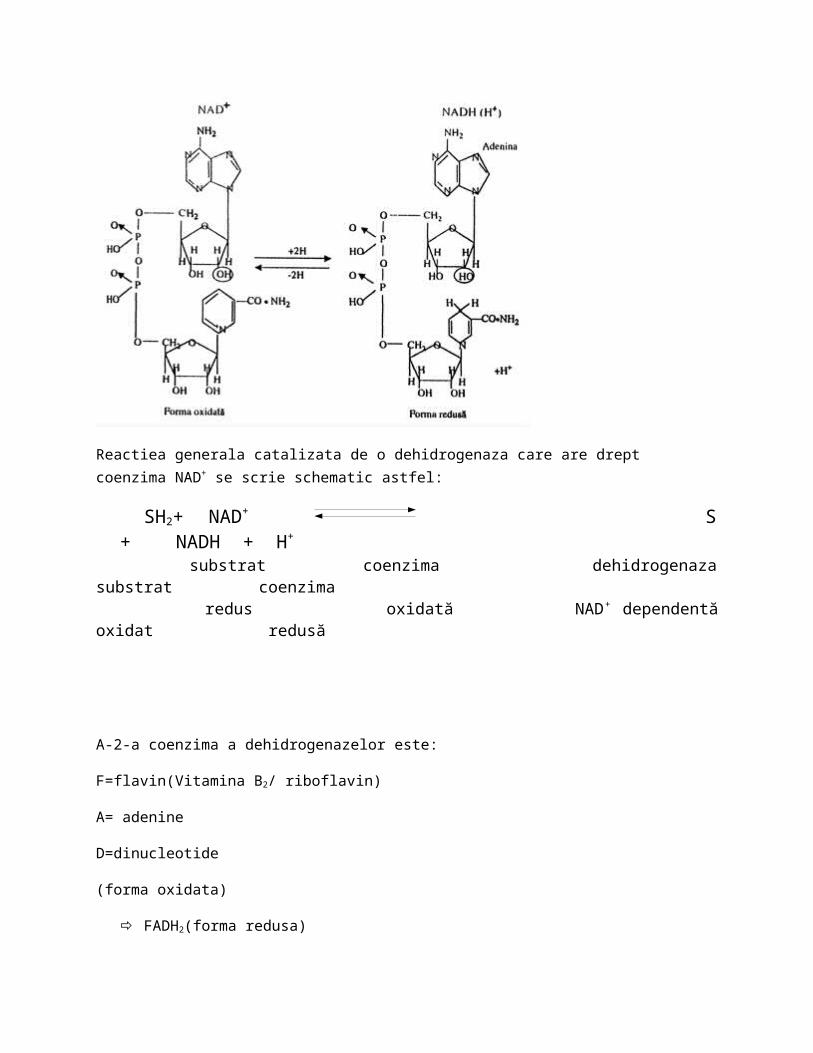
*Coenzimele dehidrogenazelor sunt derivati de la vitamin si contin in plus adenine ca baza azotata si pentoza(riboza) si 2 molecule de acid fosforic. Aceste coenzime sunt:
N=nicotinoamid( vitamin PP/niacina)
A=adenine
D+= dinucleotide (forma oxidata-forma fara H)
‖▼NADH(H+) cand primeste H (forma redusa)
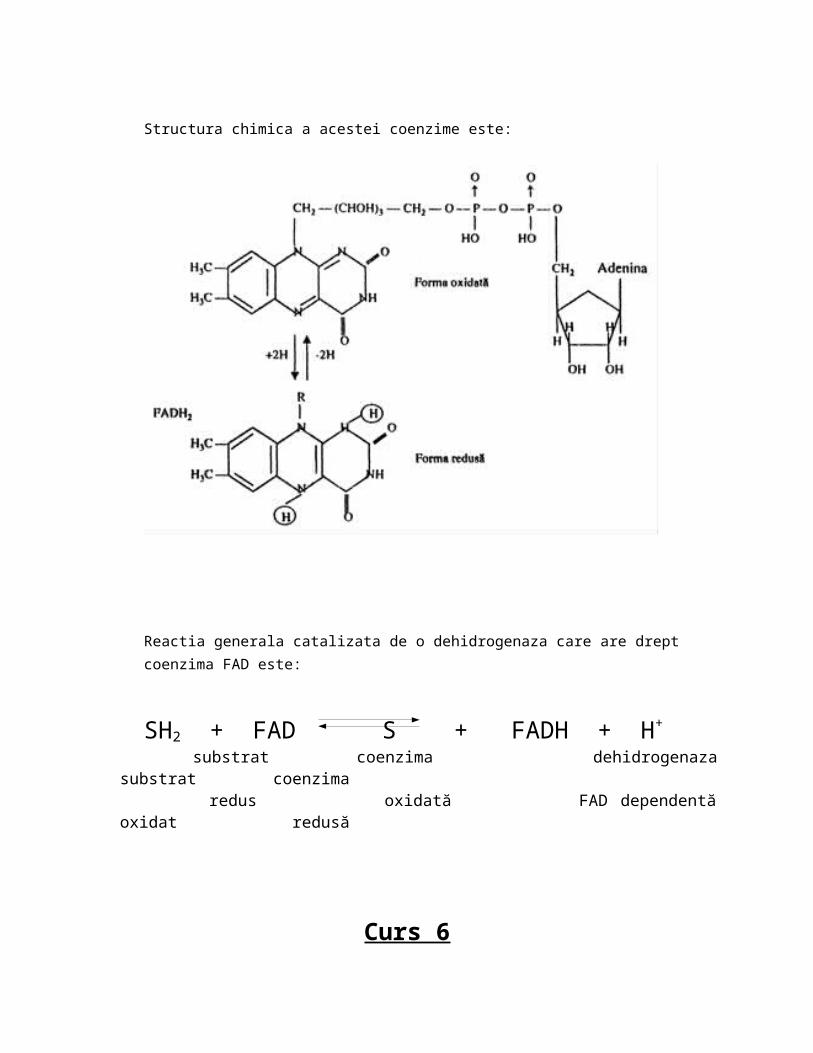
Structura acestei coenzime este:
Amintiri din copilarie:
-In chimia generala orice primire de electroni inseamna reactii de reducere.Orice cedare de electroni inseamna reactii de oxidare.
-In biochimie orice primire de H inseamna reactii de reducere; Orice cedare de H inseamna reactii de oxidare
Reactiea generala catalizata de o dehidrogenaza care are drept coenzima NAD+ se scrie schematic astfel:
-acele enzime care catalizeaza reactii redox cu transfer de electroni-ca structura chimica au drept coenzime derivati ai hemului respectiv se numesc citocromi care au in nucleu ioni de Fe2+ si Fe3+; din acest motiv pot sa treaca prin primire de electoni din Fe3+ in Fe2+(reactie de reducere) sau din citocrom din Fe2+ in citocrom Fe3+(oxidare); acest lucru este important in respirația celulara(lantul transportor de electroni si H) care este ultima parte a catabolismului glucidic lipidic proteic, fenomen la care participa acesti citocromi.
1.C Oxigenazele-sunt enzime care catalizeaza fie incorporarea oxigenului molecular(O2) intr-un substrat si se vor numi oxigenaze sau catalizeaza transferul electonilor direct pe oxigenul molecular, il transforma in O activat si se vor numi oxidaze-ex de oxigenaze-sunt mai multe, cele mai importante sunt 3 care alcatuiesc sistemul de aparare a celulei organismului viu contra fenomenelor de peroxidare lipidica membranara, fenomen cauzat de catre radicalii liberi ai O: O2
- (anion super oxid), H2O2(apa oxigenata) , HO-( anionhidroxil)
1)Superoxiddizmutaza(SOD)-transforma O2- in H2O2
2)Peroxidaza-in hrean dar si lapte- catalizeaza reactii de transfer de apei oxigenate ca donor de oxigenbpe un substrat in apa obisnuita si O2
3)Catalaza- catalizeaza reactii in care apa oxigenata actioneaza atat ca acceptor cat si ca donor de O astfel incat rolul acestei enzime este acela de a detoxifia celula de H2O2
H2O2 + H2O2 O2 + 2H2Ocatalaze
2)Hidrolaze-in aceasta clasa intra enzime care catalizeaza reactii de scindare a moleculelor complexe de substrat in molecule mai mici si mai simple dar cu participarea apei-o astfel de reactie arata astfel:
-exemple: din categoria-glucidaze-amilaza salivara, pancreatica -maltaza -lactaza -esteraze-esteri fosforici-fosfatazele-alcaline(muschi), acide(lapte) -ATP-aza Peptidhidrolazele(proteaze)-ex: pepsina, tripsina, carboxipeptidaze, aminopeptidaze care scindeaza macromolecule proteice la capete
3)Transferaze-enzime care transfera diferite grupari functionale de pe un substrat donor pe un substrat acceptor-in functie de natura gruparii functionale transferate pot fi: -transaminazele(aminotransferaze)-vor transfera gruparea amino(NH2) de la un aminoacid pe un alfa cetoacid, aminoacidul devenind astfel cetoacid iar cetoacidul devenind aminoacid; defapt cea care transfera aceasta grupare NH2 este coenzima acestor enzime care functioneaza ca un mecanism de ping-pong;- reactia generala catalizata de o aminotransferaza este:
-ex de transaminaze:GPT(ALAT), GOT(ASAT)GPT-glutanatpiruvattransaminaza pentru ca aminoacidul 1 va fi acidul glutamic iar alfa cetoacidul 2 este acidul piruvicGOT-glutamatoxalilacetat pentru ca alfa cetoacidul 2 este acidul oxalil acetic-conzima acestor transaminaze(GOT, GPT) se numeste piridoxalfosfat si ea face mecanismul de ping-pong -acetiltransferazele(transacilazele)- sunt enzime care catalizeaza reactia de transfer a gruparilor acil de pe un substrat donor pe un substrat acceptor; coenzima acestor enzime este coenzima A; ca structura chimica este formata din acidul pantotenic(vitamina din grupul B, ADP si cisteamina care este o amina cu sulf); aceasta coenzima transfera radicalul acid prin intermediul gruparii HS(tiol) si una dintre substantele cele mai importante care a primit radical acil este acetilcoenzima A( CH3-CO~SCoA)- este punctul nodal central comun al tuturor celor 3 tipuri de catabolism -kinaze- catalizeaza transferul unui radical fosforic provenit de la ATP pe un substrat acceptor
Curs 7Continuare enzime Izomerazele-catalizeaza reactii de transformare a unei molecule de substrat intr-un izomer al sau.
Liazele-catalizeaza scindarea diferitelor molecule de substrat fara participarea apei iar legaturile chimice care scindeaza sunt lec C-C, C-S, C-P. Un exemplu este aldolaza care scindeaza fructoza 1,6 difosfatul(F-1,6-P)
Ligazele(sintetaze)-catalizeaza reactii de unire a moleculelor mai mici de substrat pentru a obtine o macromolecula, un substrat complex=> se creaza noi legaturi chimice si de aceea reactiile catalizate de aceste enzime sunt cu consum de energie( furnizata de ATP) deci sunt reactii caracteristice anabolismului.
Formarea acetil coenzimei A(CH3-CO~SCoA)
MetabolismulConsideratii generale-totalitatea reactiilor biochimice respectiv a proceselor biochimice de degradare(catabolism) si de sinteza( anabolism), reactii din care rezulta substante chimice necesare vieții, alcatuiesc metabolismul organismului viu. Se spune ca metabolismul reprezinta esenta materiala a vietii.
Procesele metabolice care alcatuiesc catabolismul sunt procese consumatoare de energie respectiv procese endergonice.
Metabolismul este alcatuit din 2 procese unitare si inseparabile respectiv catabolism si anabolism, fiecare cu caracteristicile proprii:-in catabolism(degradare) compusii macromoleculari sunt transformati in molecule simple eliberandu-se totodata energie iar aceasta energie este utilizata apoi in anabolism de catre moleculele simple=> precursori pentru a forma macromolecule proprii organismului.
METABOLISM
CATABOLISM (DEGRADARE): în care compuşii macromoleculari sunt transformaţi în molecule simple
ANABOLISM (SINTEZĂ): în care se formează macromolecule proprii din precursori mai simpli
ENERGIE ENERGIE
Se stie ca oragnismele vii au ca sursa de energie soarele si ca energia solara este transformata in energie chimica in procesul de fotosinteza iar energia chimica formata este stocata in legaturile chimice care formeaza glucide, lipide, proteine si care din planta ajung in organismul animal.
Circuitul energetic din metabolism este comun tuturor formelor de viata de la cea mai simpla la cele superioare.
SE ELIBEREAZĂ SE CONSUMĂ
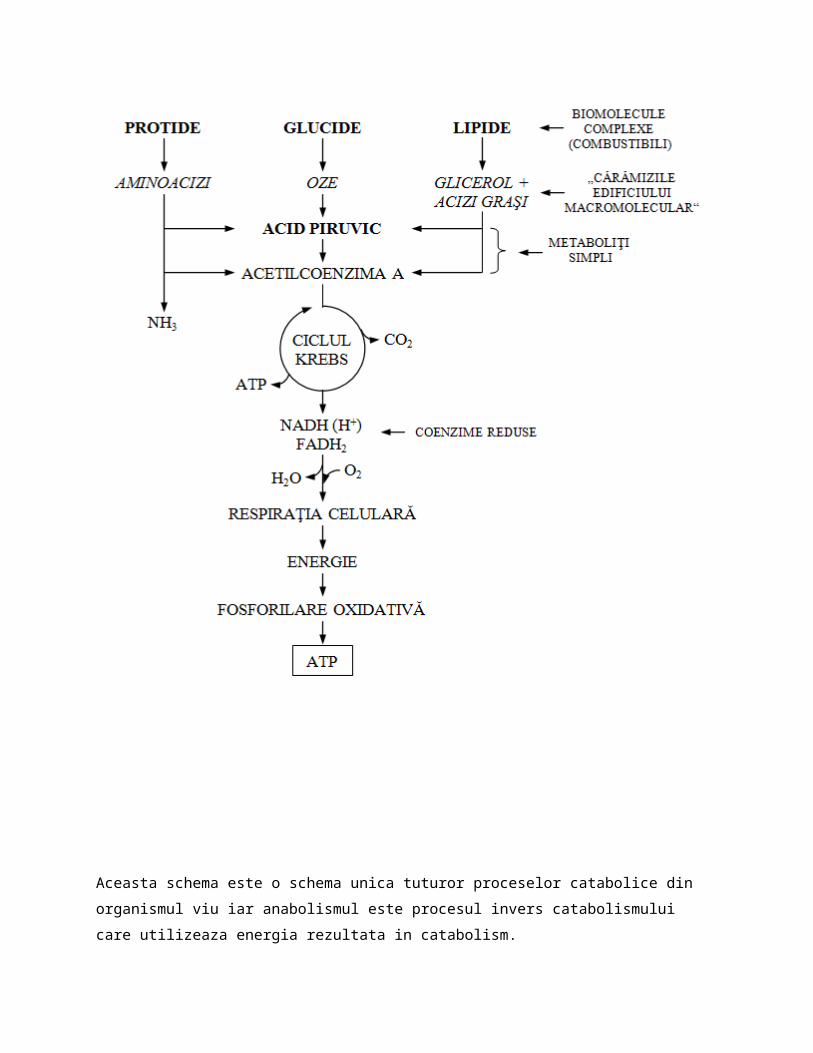
In catabolism au loc reactii de oxidare a enzimelor NADH(H+) si FADH2 si se elibereaza energie sub forma de ATP iar in anabolism au loc reactii de reducere a coenzimelor NAD+ si FAD si se consuma energie.Exista o schema unica a procesului de catabolism care arata ca de la orice clasa de compusi biochimici am porni(glucide, lipide, protide)toate au o cale comuna de degradare pananla produsi finali(CO2, H2O si ATP) . Aceasta schema unica este:
Aceasta schema este o schema unica tuturor proceselor catabolice din organismul viu iar anabolismul este procesul invers catabolismului care utilizeaza energia rezultata in catabolism.
Curs 8Metabolismul glucidicConsideratii generale
Glucidele=macromolecule cu rol structural dar mai ales energetic sintetizate in plantele verzi din compusi simpli(CO2,H2O, si energie solara)
In organismul animal ajung prin hrana si ele asigura o parte din necesarul caloric pentru animal; o parte insa fie se depune iar o mica parte sunt convertite in alte biomolecule
Cuprinde 2 parti( metabolismul glucidic):a)degradarea(catabolismul)b)biosinteza glucidelor(anabolismul)
a)Catabolismula.1) digestia glucidelor
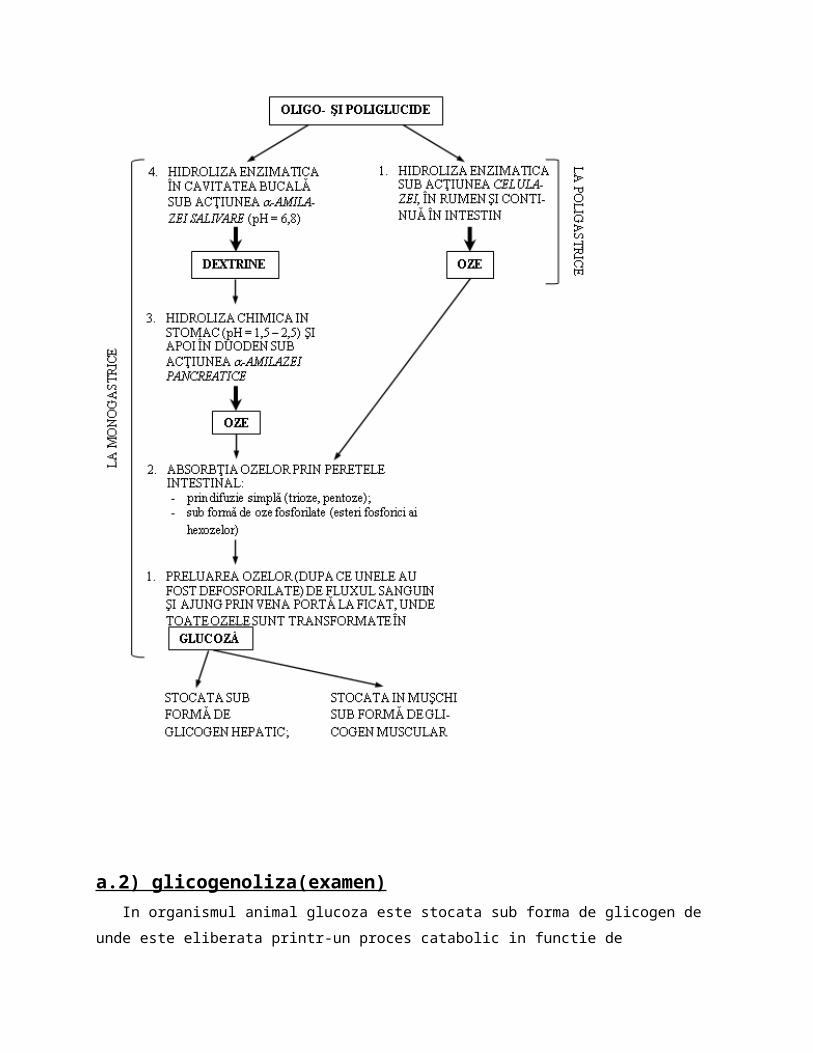
Pentru a putea strabate( a fi absorbite) prin peretele intestinal oligo si poliglucidele sunt supuse unui proces de hidroliza enzimatica(digestie) sub actiunea enzimelor numite glucidaze
Acele oligo/poliglucide care nu pot fi hidrolizate nu pot fi absorbite prin peretele intestinal, trec mai departe in tractul inferior intestinal unde sunt preluate de bacteriile de fermentatie si eliminate prin fecale. Schema generala a digestiei glucidelor este prezentata in mai multe etape astfel:
a.2) glicogenoliza(examen) In organismul animal glucoza este stocata sub forma de glicogen de unde este eliberata printr-un proces catabolic in functie de necesitatile organismului. Principala rezerva de glucoza a oeganismului imediat disponibila mai ales pentru aprovizionarea cu
glucoza a creierului si eritrocitelor este glicogenul hepatic.Deci glicogenoliza este procesul catabolic biochimic enzimatic prin care macromolecula de
glicogen este scindata atat muscular dar mai ales hepatic cu formare de glucozo-6-fosfat(G-6-P) si acesta trece in glucoza.
Prima etapa a acestui proces poarta numele de fosgorilaza deoarece participa si acidul fosforic iar enzima care catalizeaza aceasta reacție poarta numele de fosforilaza "a"*
* Enzima fosforilaza este localizata in citoplasma si se prezinta sub 2 forme:a) o forma activab) o forma mai putin activa si care actioneaza preponderent in ficat
Aceasta forma a) activa catalizeaza defapt ruperea moleculelor de glicogen la nivelul legaturilor C1-C4 glicozilice.
Aceasta forma activa intervine in conditii de efort muscular prelungit atunci cand creste necesarul de glucoza(muschiul in repaos contine fosforilaza b)
In urma procesului de fosforilaza rezulta cate o molecula de glucozo-1-fosfat si molecule mai mici de glicogen care se vor numi dextrine.
Urmatoarea etapa este izomerizarea a G-1-P in G-6-P si acesta sub actiunea H2O si a unei enzime elibereaza acidul fosforic si => glucoza care o parte ramane in sange( formeaza glicemia) iar cea mai mare parte este dusa la muschi/tesuturi si este catabolizata(glicoliza)
Schema glicogenolizei
Curs 9Glicoliza
Este procesul biochimic enzimatic de degradare a glucozei respectiv procesul de oxidare intracelulara a glucozei in scopul obtinerii de energie sub forma de ATP, energie necesara atat celulelor procariote cat mai ales eucariote vegetale si animale.
In functie de conditiile de desfasurare a acestui proces glicoliza este de 2 feluri:
1) Glicoliza anaeroba -are loc in citoplasma, se desfasoara in prezenta de O atmosferic dar fara participarea lui si consta in degradarea glucozei pana la acid lactic ca produs final si o cantitate relativ mica de energie.
2) Glicoliza aeroba - are loc in conditiile participarii directe ale O atmosferic si consta in degradarea glucozei pana la produsii finali: CO2,H2O si o cantitate mare de energie sub forma de ATP.
-cuprinde 3 etape:
a) degradarea glucozei pana la acid piruvic(process citoplasmatic) prin reactii commune cu glicoliza anaeroba
b)acidul piruvic trece in acetilcoenzima A iar aceasta intra intr-un sir ciclic de reactii biochimice enzimatice denumit ciclul Krebs(ciclul acizilor tricarbonilici). Aceasta etapa se desfasoara exclusiv in materialul mitocondrial.
c) lantul respirator(lant transportor de H si O) in care coenzimele reduse NADH(H+) si FADH2 rezultate din ciclul Krebs sunt oxidate de catre O molecular=> H2O si energie care in procesul denumit fosforilare oxidativa este stocata in legaturile macroergice alea ATP-ului.
Glicoliza anaeroba(examen)
-reprezinta o succesiune de reactii enzimatice care se petrec in conditiile aprovizionarii insuficiente cu O a tesutului muscular si atunci cand tesutul muscular (muschiul scheletic) face un efort muscular de scurta durata dar consumator de energie.
=> glicoliza anaeroba este un proces rapid de generare de cantitati mici de energie dar si de generare de produsi intermediari de metabolism necesar organismului animal in reactii de biosinteza
-este proces exclusiv citoplasmatic si ecuatia generala a acestui proces fiind:
GLICOLIZĂ
ANAEROBĂ
CH3
׀
C6H12O6 2HC OH + 2ATP
glucoză ׀
COOH
acid lactic
Etapele acestui proces se desfasoara in citoplasma, sunt in numar de 10 si sunt urmatoarele(pentru a putea fi degradata glucoza pe cale anaeroba trebuie sa fie transferate in citoplasma de catre o proteina si trebuie activata de catre ATP)
1) etapa de activare care se face cu consum de ATP
2)etapa de izomerizare a G-6-P la fructozo-6-P
triozofosfatizomeraza
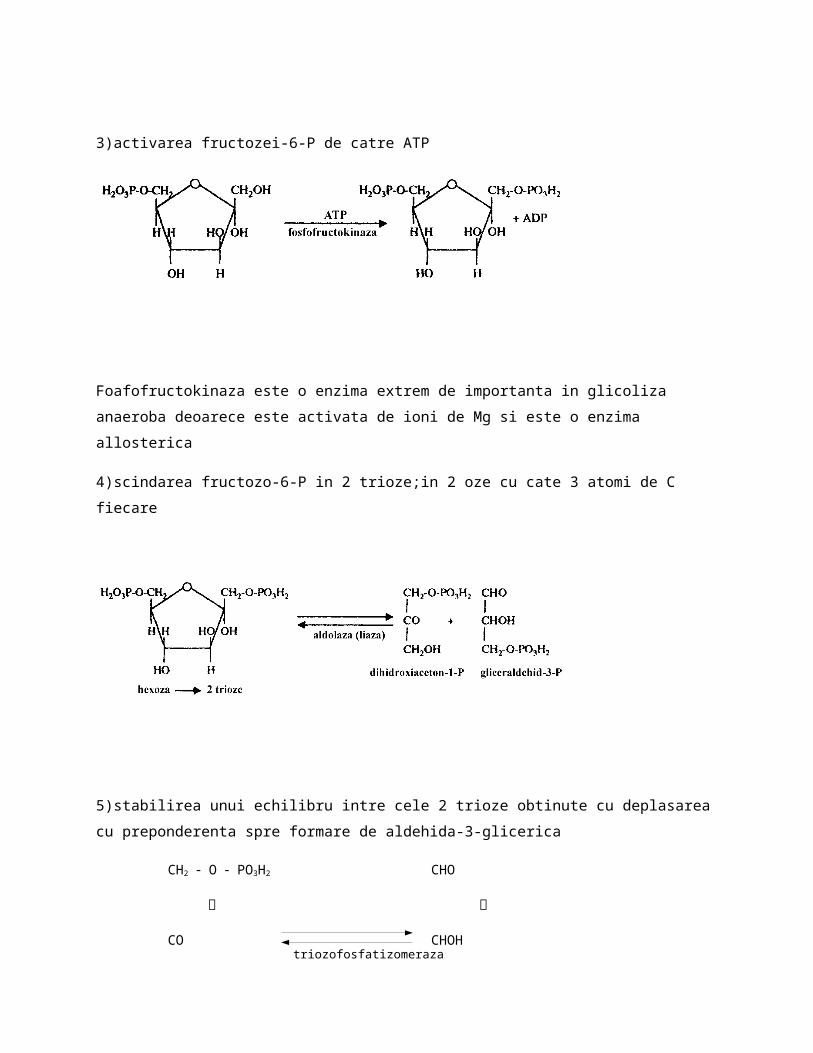
3)activarea fructozei-6-P de catre ATP
Foafofructokinaza este o enzima extrem de importanta in glicoliza anaeroba deoarece este activata de ioni de Mg si este o enzima allosterica
4)scindarea fructozo-6-P in 2 trioze;in 2 oze cu cate 3 atomi de C fiecare
5)stabilirea unui echilibru intre cele 2 trioze obtinute cu deplasarea cu preponderenta spre formare de aldehida-3-glicerica
CH2 O PO3H2 CHO
׀ ׀
CO CHOH
׀ ׀
CH2OH CH2 O PO3H2
(deci in cele 5 etape dintr-o molecula de glucoza s-au obtinut 2 aldehide-3- fosfoglicerice)
6) o etapa pur oxidativa in care aldehida 3 fosfoglicerica suferă un proces de fosforilare oxidativa, proces in care participa acidul fosforic si este catalizat de o enzima din clasa oxidoreductazelor(o dehidrogenaza)care are drept coenzima NAD+. Este formata din 2 subetape.
6.a)
6.b)acidul difosforilat care a luat nastere in etapa 6.a va ceda energia stocata in energia macroergica precum si radicalul fosfat unui mol de ATP=> un mol de ATP
(prima etapa generatoare de ATP. Defapt se produc 2 moli de ATP deoarece in etapa 5 au rezultat 2 aldehide glicerice)
7)o etapa de izomerizare cand acidul-3-Pgliceric trece in acid-2-Pgliceric prin mutarea radicalului P de la C3 la C2. Reactia este catalizata de o izomeraza
8) este o etapa de deshidratare intramoleculara cand acidul-2-Pgliceric sub actiunea enzimei enolaza trece intr-un acid nesaturat: acid-2-fosfoenolpiruvic
9)in aceasta etapa acidul 2 fosfoenolpiruvic cedeaza gruparea fosfat unei molecule de ATP=> ATP iar el trece intr-un cetoacid denumit acid piruvic
(a 2-a etapa generatoare de ATP; 2 moli de ATP deoarece s-a pornit de la 2 molecule de trioza)
In momentul in care celula are nevoie urgenta de energie sau cand tesutul este insuficient aprovizionat cu O acidul piruvic nu mai trece in mitocondrie pentru a fi degradat pana la CO2, H2O si energie, ci ramane in citoplasma si sub actiunea unei oxidoreductaze care foloseste ca, coenzima
LDH
NADH(H+)rezultat din etapa 6.a pentru a fi transformat in acid lactic ca ultim produs al glicolizei anaerobe.
Aceasta ultima etapa este catalizata de enzima lactatdehidrogenaza(LDH)
10)
COOH COOH
׀ ׀
CO + NADH(H+) CHOH + NAD+
׀ ׀
CH3 CH3
acid lactic
Bilantul energetic pentru intreaga glicoliza anaeroba
-din etapa 1 si 3 se consuma in total 2 moli de ATP
-in etapa 6.b si 9 =>in total 4 moli de ATP
-4 ATP formati-2 ATP consumati= 2 moli ATP/mol glucoza
*Daca se porneste nu de la glucoza ci de la glicogen acesta va trece intai in G-1-P, G-1-P in G-6-P fara sa mai consume ATP dupa care urmeaza etapele de la glucoza=> in acest caz nu se mai consuma un ATP si atunci bilantul energetic=> 4moli ATP generati - 1 ATP consumat = 3 moli ATP/ mol glicogen
Schema generala a glicolizei anaerobe
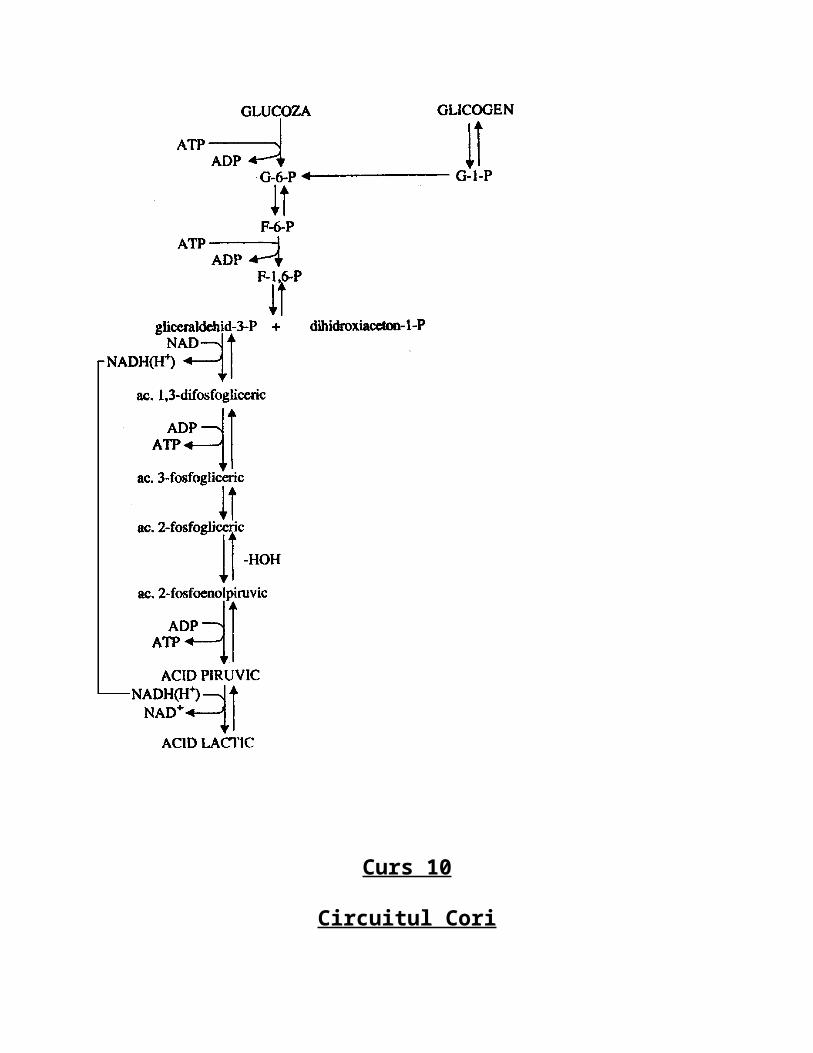
Curs 10
Circuitul Cori
Inafara de muschiul scheletic care in primele faze de efort functioneaza anaerob in organismul animal mai sunt si alte celule care produc acid lactic deoarece utilizeaza glicoliza anaeroba ca cel mai important furnizor de energie. Aceste celule sunt:
-cele din tesutul renal, celulele din retina, eritrocitele mature care neavand mitocondrie folosesc glicoliza anaeroba ca singura cale de producere de energie. In aceasta sitautie se acumuleaza in celula cantitati mRi de acid lactic si acest lucru este nociv pentru celula deoarece acidul lactic ca orice acid va scadea pH-ul citoplasmei si inhiba toate enzimele din citoplasma(inclusiv cele din glicoliza anaeroba) si atunci el este eliminat in sange, este redus/redirectionat la ficat unde este retransformat in acid piruvic si apoi in glucoza(4/5 din acidul lactic iar 1/5 este catabolizat aerob si transformat in CO2, H2O si energie).
-Acest proces ciclic poarta numele de circuitul Cori si schematic arata astfel:
Fermentatia glucidelor
-se numeste fermentatie procesul de degradare enzimatica a glucidelor de catre microorganismele heterotrofe care in felul acesta isi asigura atat hrana cat si energia necesara dezvoltarii si reproducerii lor.
-in functie de modul cum se desfasoara fermentatia ca si in functie de produsul final al fermentatiei ele se impart in 2 mari categorii:
1)fermentatii anaerobe
1.a) fermentatie lactica-acid lactic produs final
1.b) fermentatie alcoolica- alcool etilic produs final
1.c) fermentatie butirica- alcool butiric produs final
2) fermentatii aerobe- produse de microorganisme aerobe
2.a) fermentatie acetica-acid acetic
2.b) fermentatie citrica- acid citric
Fermentatia lactica(cenobioza)
-procesul enzimatic de degradare a glucidelor din lapte sub actiunea bacteriilor lactice, bacterii care transforma glucoza(lactoza) in acid lactic dupa un mecanism asemanator cu glicoliza anaeroba din muschi
-poate fi de 2 feluri
1) homofermentativa-pe langa acid lactic => cantitati mici de alcool etilic, acizi volatili si substante de aroma
2)heterofermentativa- pe langa acid lactic=> cantitati mari de alti produsi secundari precum CO2 si alcool etilic
-in tehnologia alimentara, fermentatia lactica este utilizata in procesul de lactate acide precum iaurt, kefir dar si in producerea de branzeturi
-in zootehnie fermentatia lactica este utilizata in silozarea nutreturilor dar si conservarea si fermentatia legumelor si fructelor
-procesul de fermentatie lactica poate folosi nu numai diglucidul lactoza ca substrat de reactie ci si zaharoza, maltoza, celobioza, glucoza, fructoza, galactoza,
-etapele unei fermentatii lactice sunt:
a) faza preliminara caracterizata printr-o fermentatie violenta insotita de degajare de gaze a carei durata depinde de T si natura produsului supus fermentatiei
b) faza principala a fermentatiei-faza determinanta a fermentatiei deoarece se caracterizeaza printr-o acumulare masiva de acid lactic(in aceasta faza acidul lactic creste de 5 ori)
c) faza finala caracterizata printr-o reducere treptatava acidului lactic deoarece apar microorganisme care incep sa-l consume(drojdiile si mucegaiurile) in acest moment produsul trebuie conditionat pentru ca altfel se altereaza
** Starea PSE a carnii de porc
-aceasta stare in special a carnii de porc este o stare anormala a carnii si se caracterizeaza prin paloare, printr-o moliciune si printr-o stare apoasa, exudativa si acest aspect influenteaza negativ calitatea carnii si deci influenteaza consumul
-aceasta stare se instaleaza dupa sactificare cand inceteaza circulatia sanguina si cand se produce glicoliza anaeroba si se produce acid lactic mult care scade pH-ul muschiului la 5,4 moment in care sunt inactivate marea majoritate a enzimelor din tesutul muscular si permite instalarea acestei stari anormale.
Glicoliza aeroba
Este procesul biochimic, enzimatic, cu preponderenta mitocondrial de degradare a glucozei cu participarea O atmosferic pana la produsi finali CO2, H2 si o cantitate mare de ATP
-acest proces cuprinde 3 etape:
1) formata din reactii comune cu glicoliza anaeroba, se desfasoara in citoplasma si merge pana la acidul piruvic
2) acidul piruvic rezultat in prima etapa trece in mitocondrie si numai este transformat in acid lactic ci este degarboxilat oxidativ in prezenta enzimei piruvatdecarboxilaza si a coenzimeiA si rezulta acetilcoenzimaA. Acest proces fiind un proces oxidativ ia nastere in acetilcoenzima A o legatura macroergica. Reactia de decarboxilare oxidativa este:
3) acetilcoenzima A intra intr-o roata de reactii numit ciclul Krebs(deoarece incepe si se termina cu acelasi compus); se mai numeste si ciclul acizilor tricarboxilici. Din acest ciclu => CO2, H2O, un mol de ATP direct, NADH(H+)+FADH2-> aceste coenzime sunt oxidate intr-un proces numit respiratie celulara(lantul transportor de electroni si H) din care rezulta o cantitate mare de H2O, energie care apoi in procesul numit fosforilare oxidativa=> ATP
Bilantul energetic pe toata glicoliza aeroba este 38 moli ATP/mol glucoza sau 39 moli ATP/mol glicogen
***atentie!!!! Defapt in etapa 2 => 2 acizi piruvici
Curs 11
Glicoliza aeroba continuare; Ciclul Krebs
Acetilcoenzima A care a luat nastere prin decarboxilarea oxidativa a acidului piruvic intra intr-un sir de reactii denumit Ciclul Krebs sau ciclul acizilor tricarboxilici deoarece primii 3 compusi din aceasta roata de reactii sunt 3 acizi cu 3 grupari COOH
Reactiile ciclului Krebs incep si se termina cu acelasi compus si anume acidul oxalilacetic de aceea tot procesul poarta numele de ciclu.
Etapele ciclului Krebs
In etapa 1 acetilcoenzimaA rezultata din decarboxilarea acidului piruvic sufera un proces de hidroliza=> acidul citric acesta se izomerizeaza in etapa 2 sub actiunea enzimei aconitaza si trece in acid cis-aconitic iar acesta in etapa 3 primește apa, sufera hidrolaza si trece in acid izocitric. Acidul izocitric sufera un proces de dehidrogenare (reactie redox) sub actiunea enzimei IDH( izocitratdehidrogenaza) care are drept coenzima NAD+=> prima molecula de NADH(H+) iar acidul izocitric trece in acid alfa-cetoglutaric. Acidul alfa-cetoglutaric in etapa 5 sufera de asemenea in prezenta coenzimeiA un proces de dehidrogenare sub actiunea enzimei alfa -cetoglutaratdehidrogenaza care are drept coenzima NAD+ si => a 2-a molecula de NADH(H+) iar compusul se transforma intr-un compus macroergic numit succinilCoA
SuccinilCoA in etapa 6 sub actiunea acidului fosforic si GDP(guanozindifosfat). Acesta impreuna cu acidul foaforic si cu energia moleculei macroergice din succinilCoA formeaza GTP care impreuna cu o molecula de ADP din mediul de reactie formeaza ATP+GDP(GTP+ADP->ATP +GDP) => directun mol de ATP
In etapa 7 acidul succinic format sufera al 3-lea proces de dehidrogenare sub actiunea enzimei succinatdehidrogenaza care are drept coenzima FAD si => o molecula de FADH2 iar acidul succinic se transforma in acid fumaric.
In etapa 8 acidul fumaric suferă aditie de apa la dubla legatura, sub actiunea enzimei fumaraza=> acidul L- molic
Etapa a 9-a, a 4-a etapa de dehidrogenare, enzima se numeste molatdehidrogenaza are ca si coenzima NAD+ => a 3 - a molecula de NADH(H+) si => acidoxalilacetic care preia inca o molecula din acetilCoA
Bilant energetic numai pe o tura de ciclu Krebs
-un mol de ATP direct din etapa 6
-3 moli de NADH(H+) ( din etapa 4,5,9) care prin oxidare in catena de respiratie celulara furnizeaza 3 moli de ATP pe um mol de NADH(H+)
-1 mol de FADH2(Din etapa 7) care oxidat mai departe in catena de respiratie celulara furnizeaza 2 ATP/mol FADH2
12 moli ATP total pe o tura de ciclu Krebs
Schema generală a ciclului Krebs:
Etapa 3 a glucozei aerobe
Procesul de respirație celulara(lantul transportor de electroni si H) este procesul in care coenzimele reduse NADH(H+) si FADH2 din ciclul Krebs sunt oxidate in acest lant respirator si transformate in coenzime oxidate: NAD+ si FAD
Hidrogenii rezultati din oxidarea lor impreuna cu electronii rezultati din acest proces redox prin intermediul enzimelor transelectronaze( respectiv citocromi care au in interior ioni de Fe2+) sunt transferati pe O2( oxigen atmosferic) adus de Hb(hemoglobina) si transformat in O activat care impreuna cu H formeaza apa si o cantitate mare de energie.
Aceasta energie eliberata este convertita in legaturi macroergice ale ATP-ului.
Reactiile procesului de formare a ATP-ului sunt:
Pertotal glicoliza aeroba(prima +2+3 etapa)
-=> 38 de moli ATP daca se porneste de la glucoza
-39 moli ATP daca se porneste de la glicogen
Pentru fiecare molecula de ATP energia inmagazinata in cele 2 legaturi macroergice este de 10.000 calorii/ mol ATP
Bilantul energetic calculat ca sa iasa 38 moli ATP este:
-pana la acid piruvic-> 2 moli ATP consumati; 4 moli produsi si 1NADH(H+)(3 ATP) care nu se mai consuma pentru ca piruvicu nu mai trece in acid lactic×2
-acidul piruvic trece in acetilCoA=> 1 mol NADH(H+)(3 ATP)×2
40 din care se scade 2 ATP folositi la activarea glucozei => 38 moli ATP total
Curs 12
Catabolismul acizilor grasi
1)Degradarea trigliceridelor
Acizii grasi cu numar mare de atomi de C se exterificau in mucoasa intestinala si formau trigliceride si aceste noi trigliceride, cu proteinele formau complexe lipoproteice ce ajung in ficat pe cale sanguina.
In ficat aceste trigliceride nou sintetizate se degradeaza printr-un proces de hidroliza enzimatica catalizat de enzime din clasa lipazelor cu formare de glicerina/glicerol si acizi grasi dupa o reactie de forma:
2) Degradarea acizilor grasi pe calea beta-oxidarii
Procesul de degradare a acizilor grasi se realizeaza in mitocondrie si pentru a putea fi transportati acizii grasi din citoplasma in mitocondrie ei trebuiesc activati si aceasta activare se face sub actiunea CoA si sub actiunea ATP deoarece procesul este consumator de energie.
Reactia de activare a acidului gras inainte de a trece din citoplasma in mitocondrie este:
-transportul acidului gras din citoplasma in mitocondrie este posibil datorita unei proteine purtatoare
numita carn itina care il cara in mitocondrie, il lasa acolo, ea se elibereaza si trece din nou in citoplasma pentru a prelua un nou acid gras activat.
Odata ajuns in mitocondrie acidul gras activat parcurge o succesiune de 4 reactii enzimatice la sfarsitul carora se elibereaza cate o molecula de acetilcoenzimaA si => un acid gras cu 2 atomi de C mai putin decat cel initaial care reintra in procesul beta-oxidarii astfel ca la sfarsit se vor obtine numai molecule de acetilcoenzimaA ce vor intra in ciclul Krebs.
Beta-oxidarea=proces oxidativ mitocondrial al acizilor grasi in urma careia => SCoA si un acid gras cu 2 atomi de C mai putin
Procesul beta-oxidarii cuprinde 4 reactii care sunt catalizate de enzime si care se petrec tot timpul la atomul de C din pozitia beta( adica atom de C vecin grupării COOH)
Cele 4 reactii ale procesului de beta-oxidare sunt:
1) dehidrogenarea acidului gras activat anterior sub actiunea unei dehidtogenaze care are drept coenzima FAD
2) hidratarea acidului gras activat nesaturat sub actiunea enzimei enolaza
3)( o reactie de dehidrogenare) Dehidrogenarea acidului beta hidroxi acid gras activat sub actiunea unei dehidrogenaze care are drept coenzima NAD. Reactia are loc tot la C beta
4) un proces de tioliza(rupere/liza sub actiunea coenzimeiA care contine gruparea tiol (SH)). Acest fenomen se va produce tot la C beta
Aceasta insiruire de 4 reactii enzimatice se desfasoara ca o spirala, a purtat numele de spirala Lynen si imaginistic arata asa
- moleculele de acetilCoA rezultate intra in ciclul Krebs unde vor fi catalizate aerob=> CO2+H2O si o cantitate mare de energie sub forma de ATP
Bilantul energetic pe tot parcursul beta-oxidarii
1) se calculeaza numarul de spire Lynen dintru-un proces de beta-oxidare cu formula urmatoare
nr . de ture din ¿spirala Lynen ¿
= nr . de atomi de C ai acidului2
−1¿
acidul oleic= 18/2 -1=8
-pentru fiecare tura Lynen inseamna: un FADH2=2 ATP si un NADH(H+)= 3 ATP=> 5 ATP
-numarul de ture Lynen inmultit cu 5 ATP => 8×5=40 ATP
2) Aceea prin care se calculeaza numarul de acetilcoenzimeA
nr . moleculelor de ¿acetilCoA eliberate ¿
=nr . de atomi de C ai acidului
2 ¿
Fiecare molecula intra intr-o roata Krebs inseamna 12 ATP
In cazul acidului oleic=> 9×12=108 ATP
108+40=148 ATP rezultati
148 ATP -1 consumat la inceput la activarea acidului gras =147 ATP total
Citogeneza
Este procesul de formare a corpilor cetonici.
Insituatii de infometare, atunci cand organismul pentru a-si produce energie consuma toate rezervele de glicogen si glucoza, el face apel la tesutul adipos si prin catabolismul lipidelor din acest tesut => cantitati foarte mari de acizi grasi; Acesti acizi grasi nu mai pot fi prelevati de ficat pentru a fi degradati ci urmeaza o cale metabolica anapoda in care ei sunt transformati in acetilCoA in exces, aceasta nu mai poate fi catabolizata pe calea ciclului Krebs si atunci este transformata in mitocondriile hepatocitare in 3
compusi organici care poarta numele de corpi cetonici si acestia sunt: acetona , acidul beta
hidroxibutiric si al 3 lea corp cetonic este acidul acetilacetic
Acesti corpi cetonici se acumuleaza in sange, sunt toxici, se elimina prin urina si fenomenul se numeste cetourie iar formarea lor se numeste cetogeneza. Ei se produc in organism in mod fiziologic dar in cantitati mici. Cand cantitatea lor depaseste limitele devin toxici si chiar produc mortea.Fenomenul se intalnea foarte des in grajdurile de vite care consumau furaje prost insilozate, se întâlneste in situatiile de diabet zaharat si chiar in gestație.
Curs 13
Biosinteza lipidelor
-biosinteza lipidelor apare mai ales ca urmare a excesului de glucide atunci cand organismul inlatura glicogenul cu o forma de depozit mai convenabila respectiv lipidele neutre
-apare in special la animle ca urmare a unui consum excesiv de hrana bogat in lipide in schimb la plante biosinteza lipidelor apare ca urmare a necesitatilor reparatorii a membranelor celulare
-biosinteza lipidelor inseamna defapt biosinteza de acizi grasi si de glicerina sau glicerol
Biosinteza acizilor grasi
Atat in regnul vegetal mai ales in cel animal, biosinteza acizilor grasi au drept precursor actilCoA care poate provenii din glucide, prin beta-oxidarea acizilor grasi din hrana , sau poate provenii din asa zisii amioacizi glucoformatori.
In functie de localizarea procesului de biosinteza a acizilor grasi exista 2 cai de biosinteza:
a) calea citoplasmatica( calea malonilCoA)
b) calea mitocondriala( calea elongatiei)
a) Are loc in citoplasma si se formeaza ca prim compus din acetilCoA printr-un proces de carboxilare(+CO2) si in prezenta ATP, asa numita malonilCoA(HOOC-CH2-CO-S-CoA) ; de la ea printr-o succesiune de reactii are loc biosinteza unui acid grasbcu 2 atomi de C mai mult( mai intai se formeaza nutirilCoA si apoi de la ea acidul palmitic)
b) Are loc in mitocondrie si pornește de la un acid gras activat si o molecula de acetilCoA si pe cale absolut inversa beta-oxidarii=> in final un acid gras cu 2 atomi de C in plus
Biosinteza glicerinei/glicerolului
-se porneste de la trioze fosforilate rezultate din glucoza anaeroba(aldehida -3-fosfoglicerica) si printr-un sir de 2 reactii enzimatice in prezenta de ATP=> glicerina/ glicerol, ATP, si NADH(H+)
Biosinteza trigliceridelor
-din acidul gras biosintetizat pe una din cele 2 cai amintite mai sus si din glicerol biosintetizat ca in etapa de mai sus=> printr-un proces de esterificare trigliceride(lipide simple)
Reglarea metabolsimului lipidic
-acest mecaniam de reglare presupune stabilirea unui echilibru intre lipoliza( degradarea lipidelor) si lipogeneza(biosinteza lipidelor)
-acest echilibru este de asemenea sub control hormonal si dereglari de metabolism lipidic se numesc lipodaze si se manifesta prin: tulburari de digestie, tulburari de absorbtie a lipidelor, tulburari de antrenare a lipidelor din tesutul adipos dar si tulburari de catabolism si anabolism lipidic.
Metabolismul protidic
Aspecte generale
Proteinele din hrana sunt preluate si transformate de oragnism in aminoacizi si acestia la randul lor folosesc ulterior la sinteza de proteine proprii oragnismului zi pe care nu le poate procura din hrana sau o parte din ei sunt compusi ai ciclului Krebs care mai departe se degradeaza pana la CO2, H2O si ATP( acest lucru dovedeste ca este o legatura indestructibila intre metabolismul glucidic, lipidic si protidic)
1) Digestia proteinelor
a) la monogastrice-incepe in stomac la pH-ul 1,5-2,5 si a enzimelor proteolitice denumita pepsina cand rezulta fragmente moleulare proteice cu masa moleculara mai mica(polipeptide, proteoze, peptone). Din acestea mai departe in duoden la pH 7,5-8,7 si sub actiunea enzimei tripsina=> molecule si mai mici cu 6-8 aminoacizi denumite oligopeptide. Acestea in intestin sub actiunea enzimelor denumite carboxipeptidaze sau aminopeptidaze=> dipeptide zi in final aminoacizi
b) la poligastrice-digestia proteinelor ingerate incepe in prestomace in rumen care secreta enzime proteolitice si care vor scinda cel putin 50% din proteina vegetala ingerata=> polipeptide a căror hidroliza finala se desavarseste in duoden si => aminoacizi
** In sucul gastric la animalele sugare exista asa numita renina, o enzima care precipita cazeina din lapte astfel incat ea sa stationeze un timp mai indelungat in stomac pentru a fi hidrolizata
2) Absorbtia proteinelor
-se realizeaza numai sub forma de aminoacizi la nivel de intestin subtire printr-un fenomen de competitie sub actiunea unor enzime transportoare numite translocaze care vor transporta pe tipuri de aminoacizi. Odată ajunsi pe suprafata externa a intestinului, aminoacizii sunt preluati de vena aporta, dusi la ficat unde are loc caabolismul lor
Catabolismul aminoacizilor(degradare)
-se realizeaza pe 3 cai:
A)Dezaminarea(eliminare de amoniac)
B)Decarboxilarea(eliminare de CO2)
C) Transamilarea
A) Dezaminarea
-in functie de mecanism este la randul ei de 4 feluri
1) dezaminarea oxidativa-se realizeaza in prezenta oxigenului si a unei dehidrogenaze cu coenzima FAD=> un alfa cetoacid
2) deaminarea hidrolitica care se petrece in prezenta apei=> un alfa hidroxiacid
3) deaminare refuctiva-in prezenta H furnizat de o dehidrogenaza care are drept coenzima NADH(H+)=> un acid carboxilic
4) deaminare intramoleculara-eliminare de NH3 din propria molecula
B) Decarboxilare
-eliminare de CO2 cand rezulta amine biogene
**- aminele biogene au functii specifice in organism de ex histamina este vasodilatator, creste secretia nazala, secretia lacrimala si bronsica, tiramina este prezenta in branzeturile fermentate si are actiune antagonista histaminei, unele insa sunt toxice(cadaverina, putresceina)
C) Tranasaminarea
-reactia de transfer a unei grupari NH2 de pe un aminiacid pe un alfa cetoacid, aminoacidul devenind cetoacid iar cetoacidul devine aminoacid.