76
Patogenia infecţiilor virale Factori virali Factori care tin de gazda Factori genetici
| Date post: | 23-Dec-2015 |
| Category: |
Documents |
| Upload: | anca-ciobanu |
| View: | 1 times |
| Download: | 0 times |

Patogenia infecţiilor virale
Factori virali
Factori care tin de gazda
Factori genetici

Factori virali
doză infectantă,
poarta de intrare,
pasaje anterioare ale tulpinii infectante pe gazde intermediare umane sau animale.
Virulenţa este dependentă de tropismul celular şi mutaţiile pe care le poate suferi un virus

Factori virali
Tropismul celular este dictat de specificitatea de receptor.este implicat controlul replicării virale de către promotori celulari prezenţi la un număr restrâns de celule, care îndeplinesc rolul de co-factori ai iniţierii infecţiei (ex. co-receptorii pentru chemokine necesari ataşării HIV);

Relatia virus-celula - gazda
natura celulei infectate este strâns legată de tropismul virusului pentru o anumită celulă; Exemplu: virusului rabiei, la care proteina G, de înveliş, are o afinitate specială pentru receptorii de acetil-colină de la nivelul ţesutului nervos, permiţând virusului să infecteze celulele neuronale.

Factori virali
Mutaţiile pot duce la amplificarea virulenţei: Ex: virulenţa virusului gripal este dependentă de structura hemaglutininei (HA).
Aceste mutaţii ar favoriza şi creşterea sensibili-tăţii HA la acţiunea enzimelor proteolitice:
- proteoliza, la rândul ei, expune extremitatea N terminală, hidrofobă care permite învelişului viral să fuzioneze cu membranele celulare ale celule-lor endoteliale şi, în acest fel,
- favorizează eliberarea NC la nivelul citoplasmei.

Factori ai gazdei:
- vârstă,
- sex,
- stare de nutriţie,
- responsivitatea sistemului imun;
- determinismul genetic.

Vârsta:
aceeaşi boală virală evoluează diferit la adulţi faţă de copii, adulţii fac mai frecvent complicaţii pulmo-nare secundare infecţiei cu VVZ şi efectul paralitic al virusului poliomielitei este mai frecvent semnalat, comparativ cu copiii.Alte infecţii sunt mai grave la nou-născut: infecţia genitală cu VHS2, de la mamă, se poate manifesta grav la nou-născut, ca en-cefalită herpetică sau ca infecţie generali-zată.

Vârsta:
Rata cronicizării HVB acute este pentru adult de 10-15%; infecţia transmisă intra-partum sau post-partum duce, în cazul mamelor AgHBe pozitive, la cronicizări ale infecţiei în 90% din cazuri la sugar, iar riscul evoluţiei ulterioare spre cancer hepatic este de 10 ori mai mare pentru produsul de concepţie, comparativ cu adultul. Adenovirusurile şi VRS determină infecţii severe la copii mici şi infecţii banale la adulţi şi copii mari.

* Status-ul nutriţional al gazdei:
factorii nutritivi deficienţi: deficitul de vitamina A, este incriminat în infecţia rujeoloasă, în complicaţiile oculare ce pot evolua cu cecitate;infecţia cu HIV la malnutriţi determină o entero-patie severă; există imunodeficienţe induse de acest deficit nutriţional, dar şi imunodeficienţe congenitale, cu diferite consecinţe în funcţie de veriga afectată:
- la agammaglobulinemici, infecţiile cu VHS sau virus rujeolos se vindecă fără complicaţii, dar la cei cu imunodeficienţe ale imunităţii celulare, aceleaşi virusuri determină infecţii letale;

* Status-ul nutriţional al gazdei:
imunodeficienţa la pacienţi cu SIDA, contraindică administrarea de vaccin viu atenuat; s-au semnalat complicaţii locale adenoidie-ne după vaccinarea BCG la copii cu HIV.Pe de altă parte, riscul unor infecţii severe la copiii nevaccinaţi subliniază avantajele vaccinării.In acest context, la copiii seropozitivi, asimptomatici, se pot utiliza vaccinuri inactivate.

Factorii genetici ai gazdei
pot favoriza instalarea unor boli viraleEx.: HIV infecţie:
indivizii HLA-B35 au un risc mai mare de a evolua spre SIDA, comparativ cu cei HLA-B8 şi HLA-DR3;persoanele HLA-DR2 prezintă un risc mai mare de a contracta infecţii cu opor-tunişti, iar cei HLA-DR5, de a dezvolta un sarcom Kaposi.

Sexul:
frecvenţa carcinomului nazofaringian determinat de VEB ca şi rata cronici-zării hepatitei B este mai mare la bărbaţi, în timp ce infecţiile cu VCM sunt mai frecvente la femei;
„constelaţia hormonală” pare să joace rolul esenţial.

Mediu:
prin complexitatea lui şi inter-relaţiile factorilor geografici, demografici, etc. contribuie diferit la evoluţia infecţiei virale: sporadic, epidemic sau chiar pandemic.

Transmisibilitatea:
în lanţul epidemiologic intervin mai ales factori care ţin de gazdă:
- momentul;- durata contactului infectant;- susceptibilitatea;- status-ul imun, etc.
Transmiterea orizontală (aerogenă, digestivă, contact direct, etc.) determină epidemii explozive, Transmiterea de la mamă la făt menţine virusurile în colectivităţi izolate, cu densitate mică.

Poarta de intrare
cel mai frecvent este reprezentată de cele 3 suprafeţe epiteliale:
pielea şi mucoasele;
tractusul respirator;
tractusul digestiv.
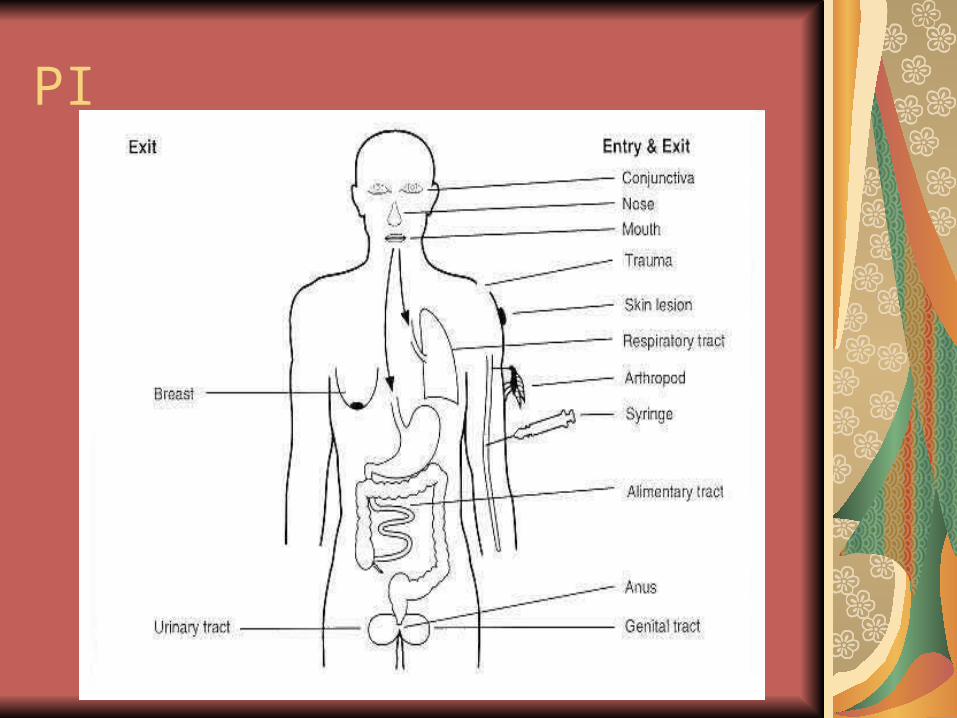
PI

Evolutii:
uneori, infecţia rămâne cantonată la poarta de intrare (PI);
frecvent, aceste infecţii sunt asimpto-matice sau de severitate medie şi, eventual, virusul se propagă spre o nouă ţintă.
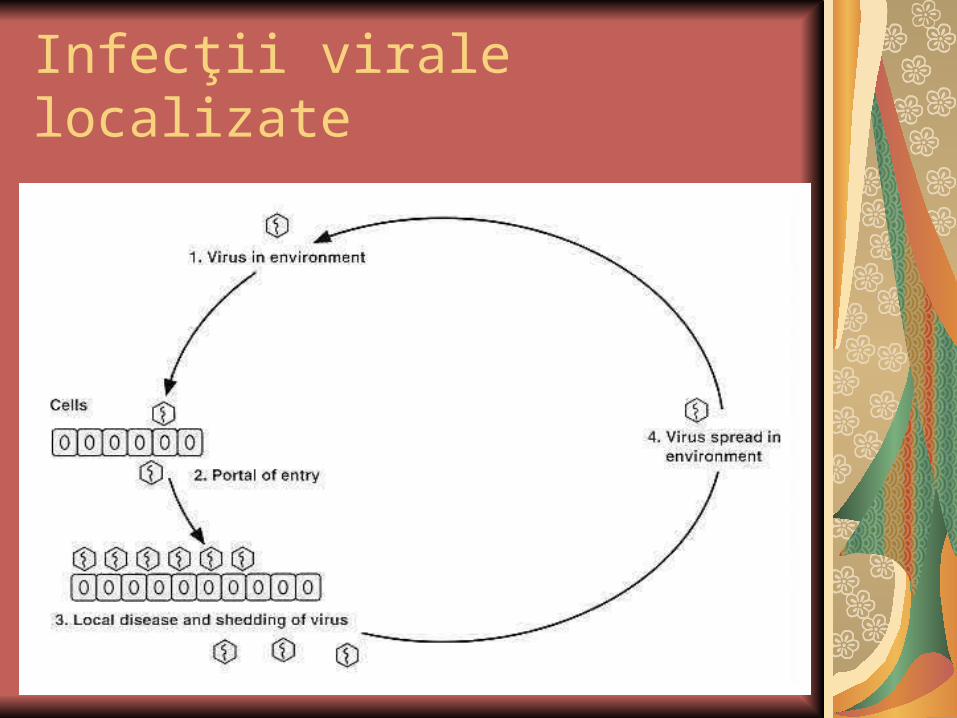
Infecţii virale localizate

Evolutii:
Cel mai frecvent are loc diseminarea, care poate avea loc prin:
- contiguitate
- sistemic.

Răspândirea prin contiguitate
este caracteristică pentru acele infecţii care rămân, practic, limitate la PI. Ex.: majoritatea infecţiilor respiratorii (infecţiile cu rinovirusuri sau virusuri gripale) şi unele infecţii cutanate (ex. verucile).Incubaţia acestor infecţii este de scurtă (2-3 zile), viremia survine după debutul bolii şi, secundar ei, nu se supra-adaugă noi simptome. Reinfecţia cu aceste virusuri este frecventă, deoarece Ac care apar nu sunt protectori. Ac secretori IgA pot oferi o protecţie de scurtă durată la nivelul mucoaselor.

Diseminarea sistemică:
după penetrarea PI, virusul ajunge în sânge; viremia este cea care vehiculează agentul etiologic spre alte ţesuturi, anterior debutu-lui clinic. Deoarece viremia precede debutul, perioa-da de incubaţie pentru aceste infecţii este de 2-3 săptămâni. Viremia asociată diseminării sistemice are 2 stadii: primul stadiu se termină când virusul este preluat de sistemul reticuloen-dotelial.

Diseminarea sistemică:
Activarea sistemului RE poate duce la:eliminarea virusului (deoarece apar Ac de tip neutralizant), deci infecţia este abortivă (ex. infecţia poliomielitică);multiplicarea virusului la nivelul SRE, determină o a doua viremie, de mai mare amploare şi care induce manifestarea clinică a bolii.
Reinfecţiile: Ac prezenţi în circulaţia siste-mică au rol protector, reinfecţiile fiind, de regulă, rare.
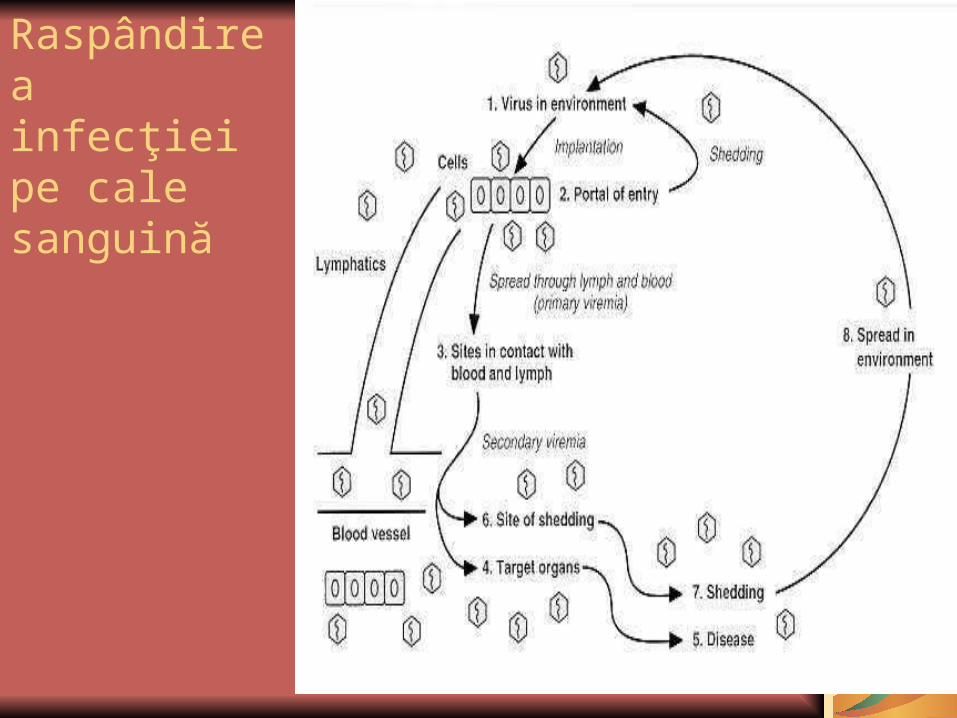
Raspândirea infecţiei pe cale sanguină

Organele ţintă:
multe virusuri au predilecţie pentru anumite organe (ex. rotavirusurile şi agentul Norwalk pentru tractusul gastro-intestinal, virusurile hepatitice primare A-G, pentru hepatocit, etc.).
Această predilecţie nu este absolută: unele virusuri au predilecţie pentru un anumit organ, dar pot infecta şi alte organe/ ţesu-turi (ex. VHS determină atât leziuni orale, cât şi meningoencefalite).

Organele ţintă:
Accesul virusurilor la nivelul pielii sau mucoaselor, secundar unor leziuni mecanice (muşcătura animalului turbat, înţepă-tura insectei care vehiculează un arbovirus sau
acul contaminat), determină infecţii gene-ralizate, de cele mai multe ori fără modificări locale.
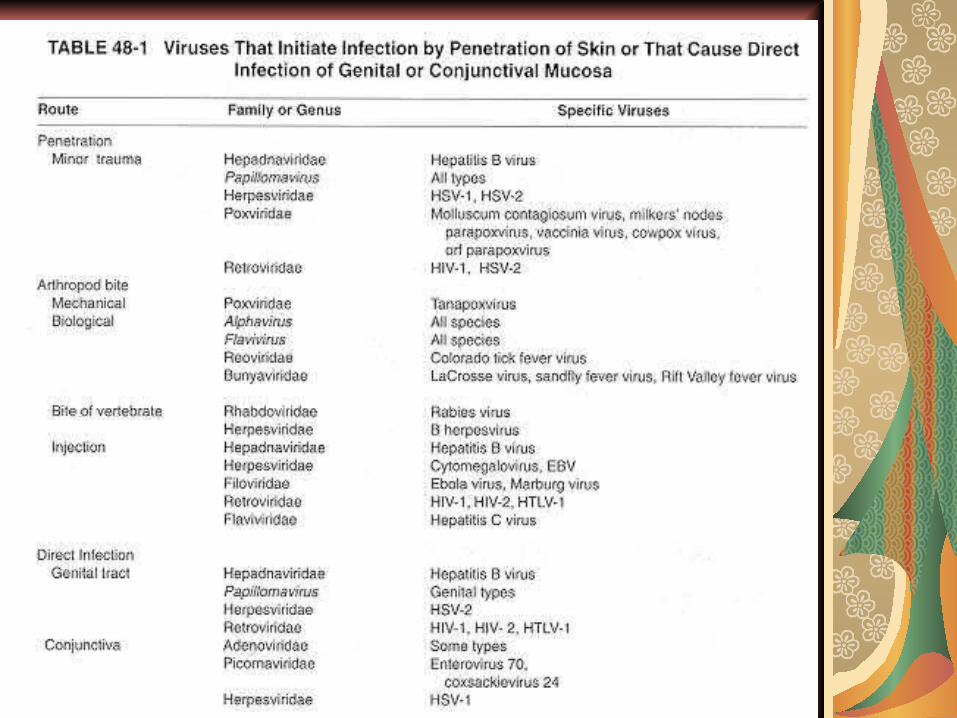

Organele ţintă:
Alte viroze cu aceeaşi poartă de intra-re determină doar manifestări locale:
- negii sau verucile sunt determinate de papillomavirusuri;
- moluscum contagiosum este determinat de un poxvirus (există doar hiperplazia epidermului şi leziuni inflamatorii prolife-rative).

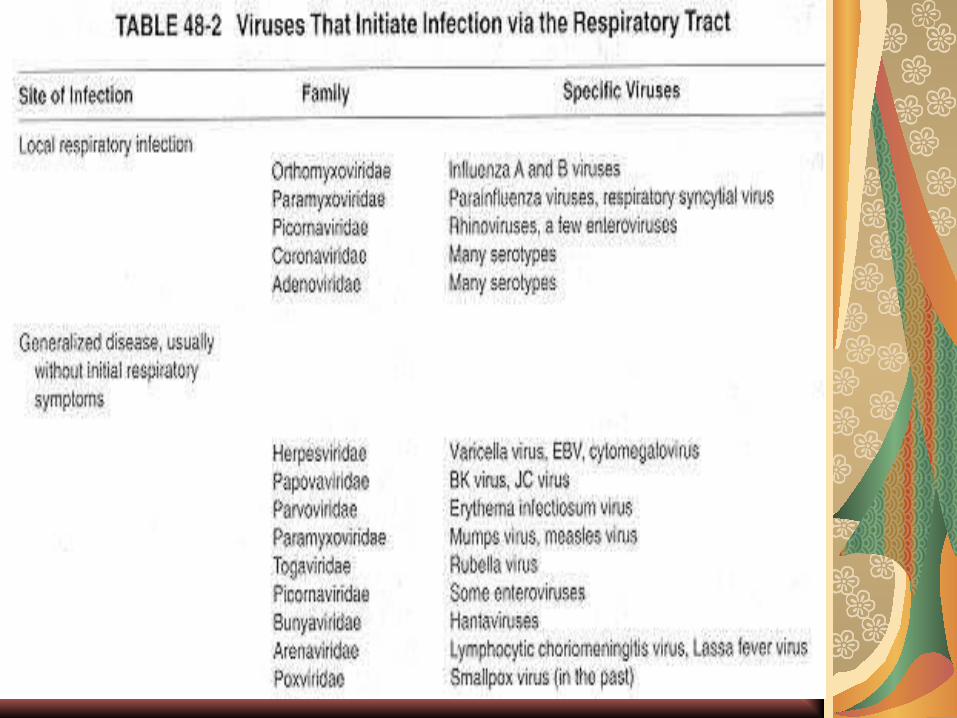
PI respiratorie:
orthomyxovirusuri, paramixovirusuri, picorna-virusuri, reo-, corona- şi adenovirusuri. Frecvent, replicarea iniţială este urmată de generalizarea infecţiei. Alte viroze se caracterizează doar prin localizarea strictă la nivelul mucoasei respi-ratorii:
- virusul gripal infectează iniţial doar câteva celule, virionii progeni rezultaţi, în număr mare, sunt diseminaţi prin intermediul mucusului şi pot determina afectarea bronho-pulmonară.

PI respiratorie:
transudatul care însoţeşte leziunile celulare favorizează diseminarea, iar pe de alta, factorii locali (macrofage, modificări de pH, IgA secretor sintetizat ceva mai târziu) au rol limitativ.
Leziunile iniţiate de virus se pot suprain-fecta bacterian cu germeni condiţionat patogeni (pneumococ, hemofil, stafilococ) care nu ar determina infecţia fără această cauză favorizantă

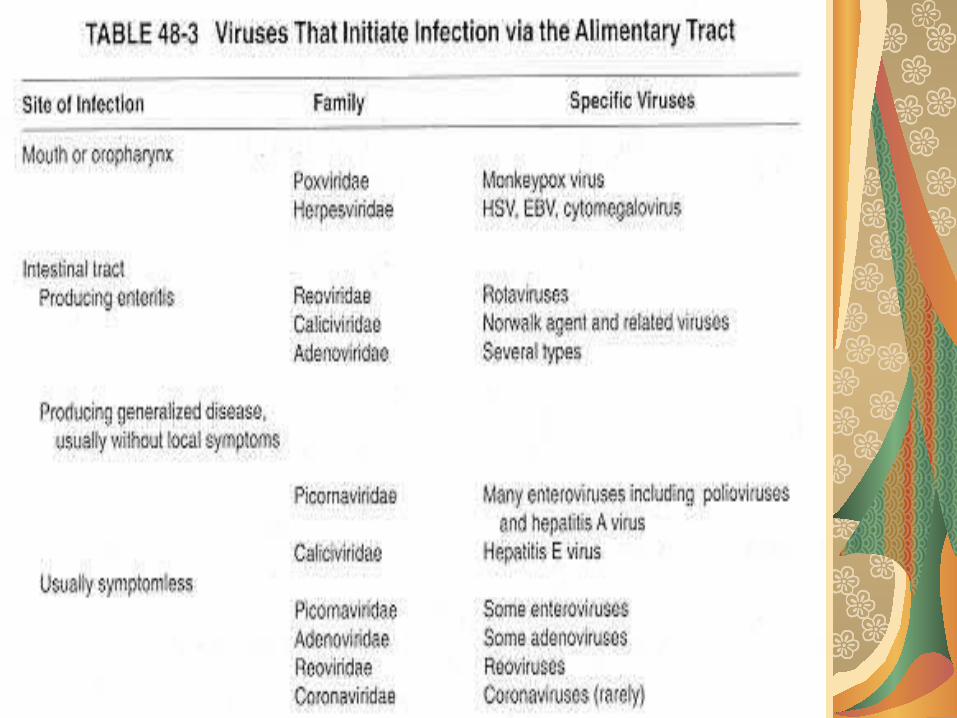
PI digestivă:
preferată mai ales de virusule neînvelite, deoarece virusurile cu înveliş sunt sensibile la pH acid, la detergenţii din bilă sau la proteazele din sucul digestiv. Picornavirusurile, reovirusurile, unele adenoviru-suri (ex. 40, 41, 42, 52) pătrund prin mucoasa digestivă şi determină doar simptome iniţiale locale reduse. Patogenitatea acestora se manifestă secundar generalizării infecţiei. Alte virusuri, asociate gastroetenteritelor endemo-epidemice, nu depăşesc epiteliul digestiv.

PI digestivă:
Rotavirusurile (din familia Reoviridae) şi agenţii asemănători virusului Norwalk sunt patogeni primari şi exclusivi pentru epiteliul intestinal.
Diseminarea de la PI se face, cel mai frecvent, pe cale sanguină, prin viremie. In paralel, există şi o diseminare limfatică; uneori este implicată diseminarea pe cale nervoasă, respectiv pe calea tracturilor nervoase, prin septinevrită.
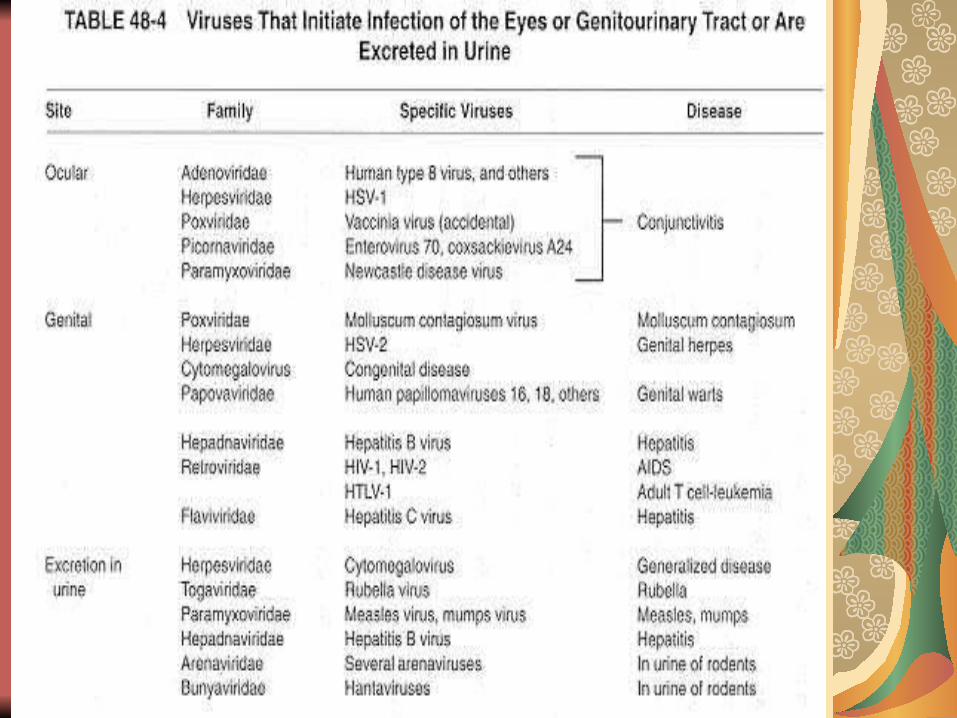
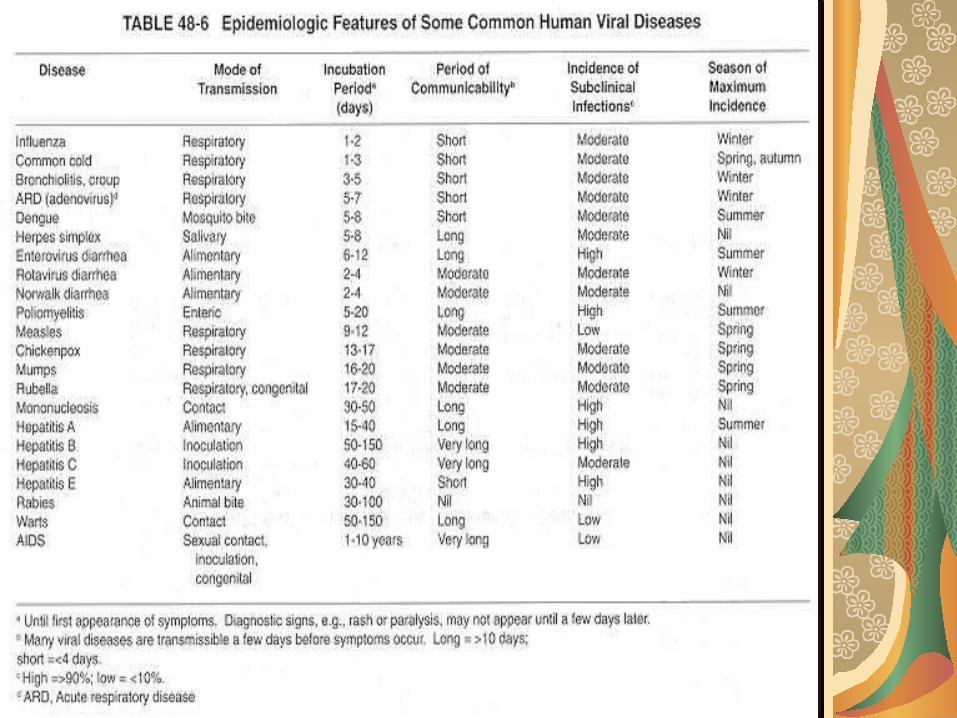

PI – organe tinta
în infecţia rujeoloasă, virusul pătrunde pe cale respiratorie, replicarea locală are loc la nivelul ţesutului adenoidian de la nivelul inelului Walda-yer, după care virusul multiplicat în cantităţi sufici-ente este diseminat prin viremie. Limfocitele transportă virusul şi, mai mult de atât, permit replicarea virusului intracelular. Virusul din sânge este permanent îndepărtat de celulele SRE şi astfel sunt afectate organele cu circulaţie bogată: plămânii (bronhopneumonia rujeoloasă), ficatul, pancreasul, SNC, dermul (rash rujeolos).

Evolutie:
Secundar replicării în aceste organe ţintă (unele de mari dimensiuni) survine al doilea val viremic, viremia secundară, limitată de sinteza de anticorpi specifici care duc la vindecarea infecţiei;vindecarea este consecinţa intervenţiei şi a altor mecanisme ale imunităţii celulare.
Descrierea anterioară a diseminării virusului nu ţine cont de unele particularităţi:
virusul poate pătrunde şi la nivelul conjunctivei,diseminarea la nivelul SNC poate avea loc pe căi multiple, fagocitarea virusului de către celulele SRE explică traver-sarea barierei hematoencefalice cu afectarea căilor gliale şi a neuronilor, etc.

Evolutie:
Viremia nu presupune întotdeauna vehicu-larea virusurilor prin intermediul limfoci-telor;
unele arbovirusuri sunt adsorbite de eritrocite, parvovirusurile au fost puse în evidenţă ataşate de trombocite, iar multe picornavirusuri circulă libere în plas-mă.
Alteori, viremia este mixtă: o parte dintre virusuri sunt asociate elementelor figurate, o parte sunt libere în plasmă.

Patogenia infecţiilor cu rash
Erupţiile generalizate sunt consecinţa unui răspuns imunopatologic. Sinteza de Ac faţă de antigene virale, prezente în derm, se soldează cu formarea de complexe Ag-Ac. Acestea determină hiperemie şi acumularea de exsudat - substratul histopatologic al leziunilor maculo-papulare.

Patogenia infecţiilor cu rash
Uneori virusul intervine direct, prin acţiunea lui citopatică;Ex: VHS sau VVZ determină:- hiperplazia dermică, - balonizarea celulelor infectate şi - apariţia incluziilor intranucleare (IN) care conţin
antigen viral, în primele stadii ale infecţiei iar mai târziu,
- pierd antigenul, devenind doar “cicatrici” ale infecţiei.

Patogenia neurovirozelor:
Virusul rabic infectează omul după muşcătura animalului rabigen şi se multiplică iniţial la nivelul ţesutului conjunctiv dermic sau muscular.
Propagarea ulterioară este centripetă, pe traiectele nervoase, către SNC.

Patogenia neurovirozelor:
Leziunile, considerate iniţial patogno-monice, de la nivelul cornului Amon, sugerează că virusul se multiplică mai ales în această zonă. Ulterior, diseminarea periferică se face centrifug, dar tot pe traiectele nervoase. Virusul nu a fost izolat din sânge (la om).
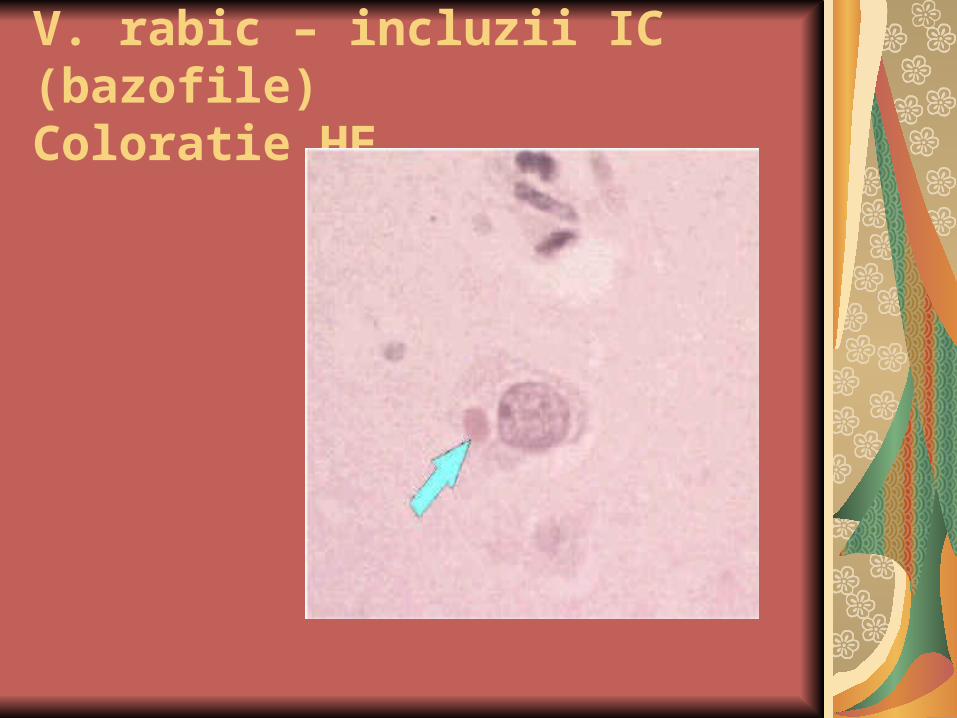
V. rabic – incluzii IC (bazofile)Coloratie HE

Patogenia neurovirozelor:
Perioada de incubaţie depinde de distanţa dintre locul de inoculare şi encefal;
după muşcăturile la faţă sau la membrele superioare, incubaţia bolii este mai scurtă.

Patogenia neurovirozelor:
In alte infecţii, pe lângă invazia SNC pe cale septinevritică, este implicată şi diseminarea sanguină (entero-, arbovirusuri).
Căile pentru traversarea barierei hematoen-cefalice pot fi: - transferul prin endoteliul vascular al vaselor mici
cerebrale;- transferul consecutiv infecţiei meningelui sau
la nivelul plexurilor coroide.

Patogenia neurovirozelor:
Infecţia SNC se caracterizează prin prezenţa a trei tipuri de leziuni:
- necroză celulară;- neuronofagie;- acumularea de infiltrat mononuclear perivascular.
In infecţia cu virus rabic, care nu este citocid, ca şi în infecţiile cu agenţii encefalopatiilor spongiforme subacute există degenerare neuronală cu vacuolizare, leziuni inflamatorii minime sau chiar absente. Afectarea neuronală, deşi puţin evidentă d.p.d.v. morfologic, pare să influenţeze funcţii înalt specia-lizate ale celulelor infectate.
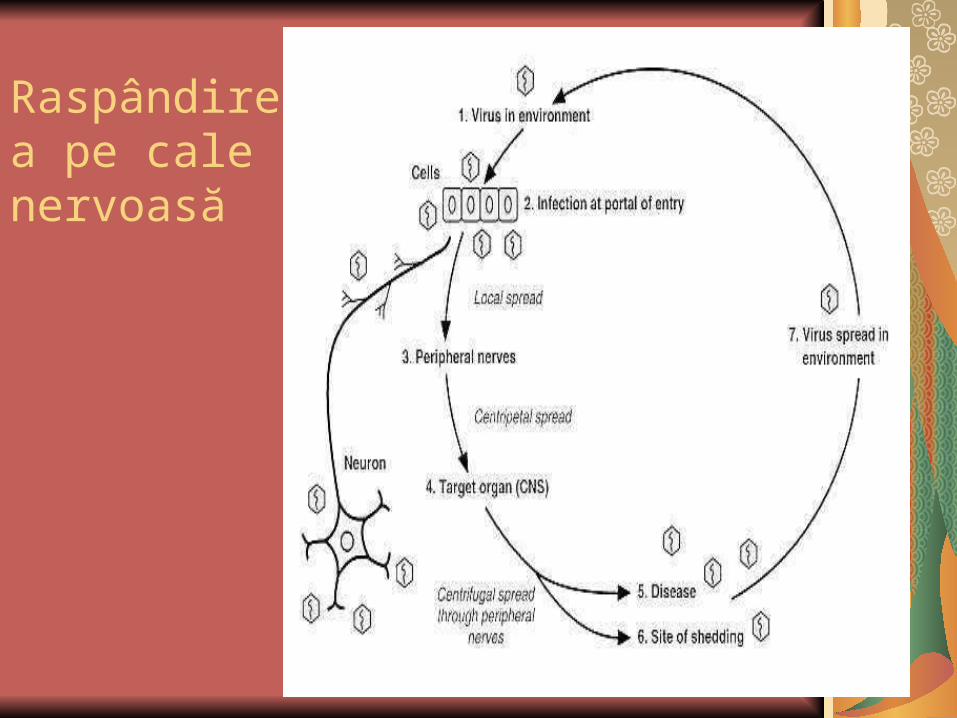
Raspândirea pe cale nervoasă

Evoluţia ciclică a infecţiilor virale
Stadiile succesive ale infecţiilor virale corespund celor din infecţiile bacteriene:
incubaţie: perioada de la contactul infectant la debutul simptoamelor;perioada de debut: asociată, de regulă, viremiei primare;perioada de stare: în care are loc replicarea “în organele ţintă”;perioada de convalescenţă: care duce, de cele mai multe ori, la vindecare.

Evoluţia ciclică a infecţiilor virale
Durata fiecărei perioade este caracte-ristică virozelor, cu variaţii ce ţin de vârstă, teren, coinfecţii, intervenţii terapeutice, doză infectantă, etc. Diferenţa între infecţiile sistemice şi cele limitate la poarta de intrare se referă mai ales la contagiozitate (excreţie de virus) şi durata replicării virale.

Evoluţia ciclică a infecţiilor virale
Durata perioadei de incubaţie este relativ constantă: scurtă în cazul virozelor în care simptomatologia este în totalitate pusă pe seama virusului care se replică la PI (1-3 zile), sau de 2-3 săptămâni în virozele generalizate.

Evoluţia ciclică a infecţiilor virale
In infecţii determinate de lentivirusuri sau oncornavirusuri, în care integra-rea genomului viral în genomul celulei gazdă determină o perioadă de laten-ţă lungă (ani sau chiar zeci de ani), asimptomatică, infecţia devenind manifestă după intervenţia unor co-factori care activează provirusul din starea dormantă în cea activă, repli-cativă.
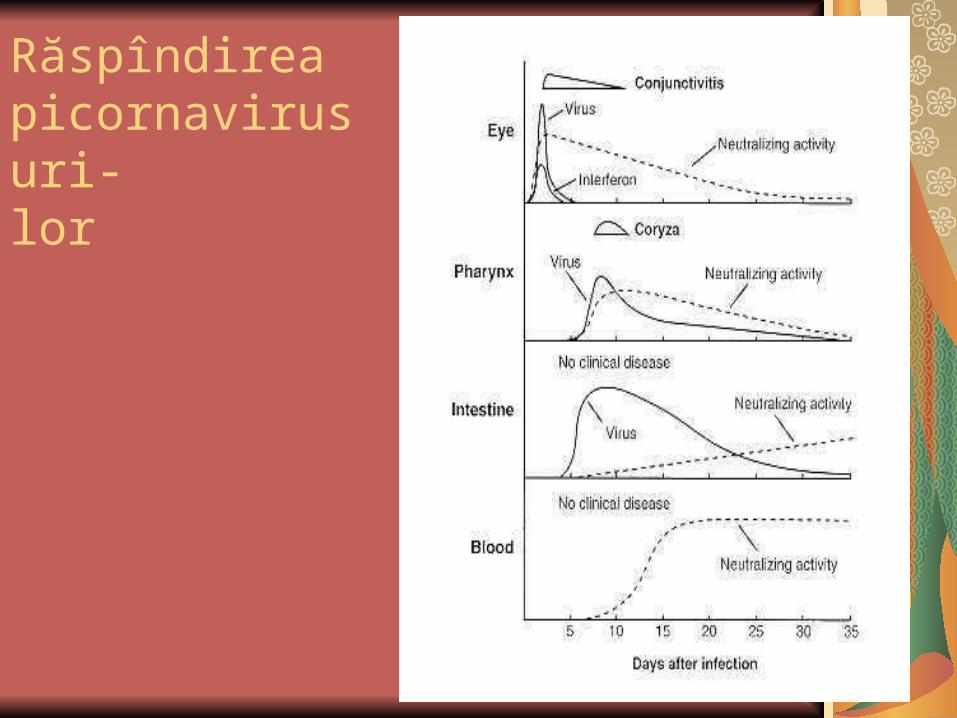
Răspîndirea picornavirusuri-lor

Infectii latente
Infectii subclincie, dar virusul nu este complet eliminat.
Exista o stare de echilibru intre gazdă si virus;
Căi difertite de a se instala:- virus cu genom integrat;- infectie persistentă.

8.05

Infectii latente
Infecţii persistente (replicarea virusului continuă).
Virusul eludeaza mecanismele apărării celulare:- generarea de celule care eludează răspunsul
imun mediat celular;- diminuarea răspunsului/intervenţiei CMH în
celulele infectate (nu sunt recunoscute si distruse de către celulele T);
- infectarea unor celule din sedii “privilegiate” (creier).

Infectii latente:
Sindromul rubeolos congenital
Infectia cu VCM, VEB, VHB, HIV
Infectii latente cu: VHS, VVZ, adenovirusuri, retrovirusuri
PESS, pancencefalita progresiva rubeoloasa
boala Kuru

Infectiile cronice persistente:
Pot fi produse de virusuri invelite: paramyxo-virusuri, herpesvirusuri – VEB, retrovirusuri si arenavirusuri.
Mecanismul de persistenţă: virusurile nu blocheaza functiile esentiale ale celulei.
In rujeola (PESS – panencefalita sclerozanta subacuta), celulele infectate persistent pot fi ajutate de capacitatea anticorpilor de a recunoaste antigenele de pe suprafata celulelor.
Antigenele virale de pe suprafata celulelor sunt distruse, celula fiind eliberata de glicoproteine virale si astfel celula este protejata de celule NK si Limf T.

Infectiile virale latente – oculte
anumite virusuri ADN sau ARN pot deveni nedetectabile dupa o infectie primară;
in timp, reapar şi produc infectie acuta;
aceasta latenţă poate fi îndeplinită prin mai multe mecanisme:

VHSinfectia primara apare la varsta de 6 – 18 luni, dupa care virusul persista si nu poate fi identificat, cu exceptia episodului acut;nu se stie sigur mecanismul prin care virusul persista. Virusul nu poate fi izolat din tesuturi, dar poate fi co-cultivat in culturi de celule –ganglioni senzitivi.VHS a fost identificat in ganglionii: trigeminal, toracic, lombar si sacrat. Studii de hibridizare au detectat genomul viral in creier (fara leziuni), ca si in ganglionii periferici. Aceste date sugereaza ca ADN-ul viral exista in forma lineara, neintegrată, probabil ca episomi
(unitate independentă de material genetic care se poate replica independent sau se poate integra in genomul gazdei).

VHS- continuare
Este posibil ca virusul sa afecteze doar o mica proportie din ganglion (0.01 – 0.1%) datorita anticorpilor, interferenţei virale sau factorilor metabolici.Anticorpii pot neutraliza cea mai mare parte din virus care devine, astfel, nedetectabil. Episodul acut poate fi explicat printr-o modificare:
a nivelului imunităţii sau prin schimbari in susceptibilitatea celulelor neinfectate, care pot fi influenţate de febra, expunere prelungita la razele solare, obosela, menstruatie.
Si alte herpesvirusuri produc infectii latente: VVZ – ganglioni senzitivi, VCM – limfocite si macro-fage, VEB – limfocite B.

Adenovirusuri
Infectiile umane cu adenovirusuri sunt de obicei autolimitate, dar pot stabili o infectie latenta, persistenta, a amigdalelor si ganglionilor.
Desi nu s-a reuşit izolarea virusului din aceste ţesuturi în culturi de celule, fragmente din aproximativ 85% din aceste culturi normale au prezentat, dupa o perioadă de timp variabil, ECP (efect citopatic).

Adenovirusuri
Eşecul initial se poate datora cantităţii mici de virus din produsul patologic, anticorpilor specifici sau absenţei virionilor maturi.
Infectiile latente nu apar probabil prin lizogenie, deoarece ADN-ul viral din limfocitele periferice apare ca o forma lineară, episomală.

PESSlatenţa apare ca rezultat al producerii incomplete de virion;sunt produse virusuri rujeolice imature;infectiile virale latente afectează incidenţa si patogeneza bolii acute virale in câteva moduri:
- un virus reactivat poate răspândi si initia o epidemie la contacti susceptibili. – e.g. VVZ;
- latenţa virala poate fi de asemenea remarcata in cazul dezvoltării infecţiilor cronice dependente de răspunsul imun – e.g. PESS
anumite virusuri latente induc evoluţia spre neoplazii.

CULTIVAREA VIRUSURILOR
EFECTE CITOPATICE
INCLUZII VIRALE
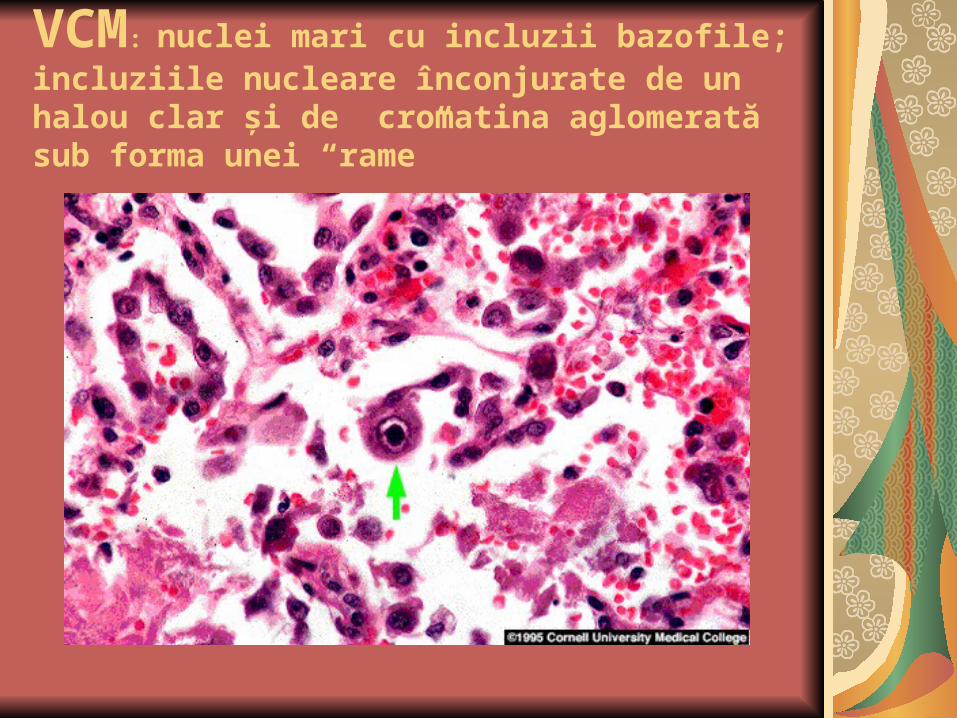
VCM: nuclei mari cu incluzii bazofile; incluziile nucleare înconjurate de un halou clar şi de cromatina aglomerată sub forma unei “rame”
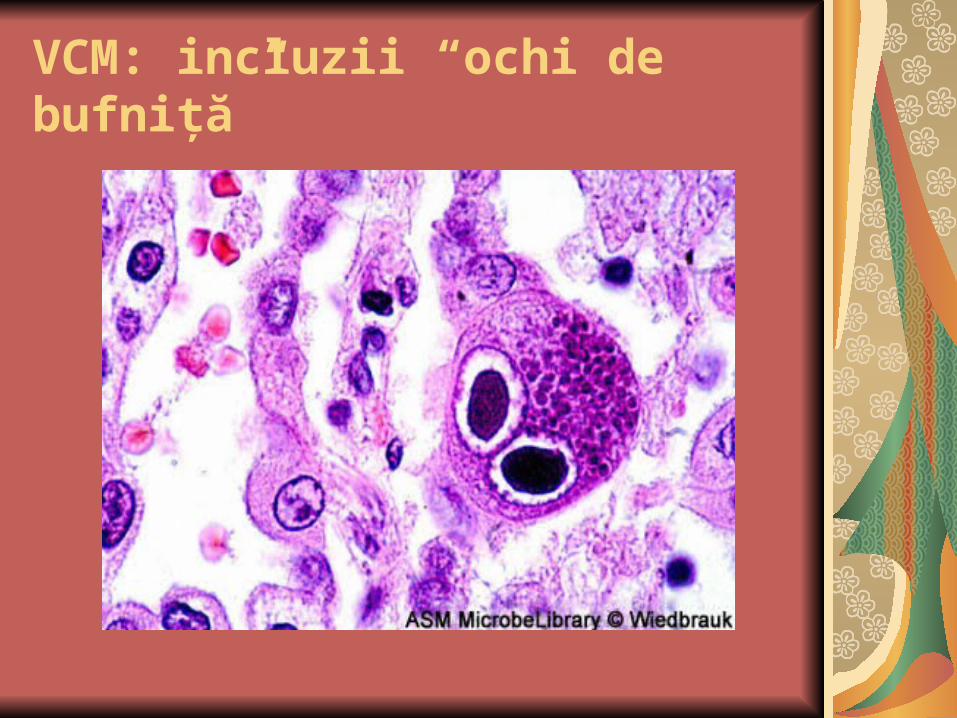
VCM: incluzii “ochi de bufniţă”
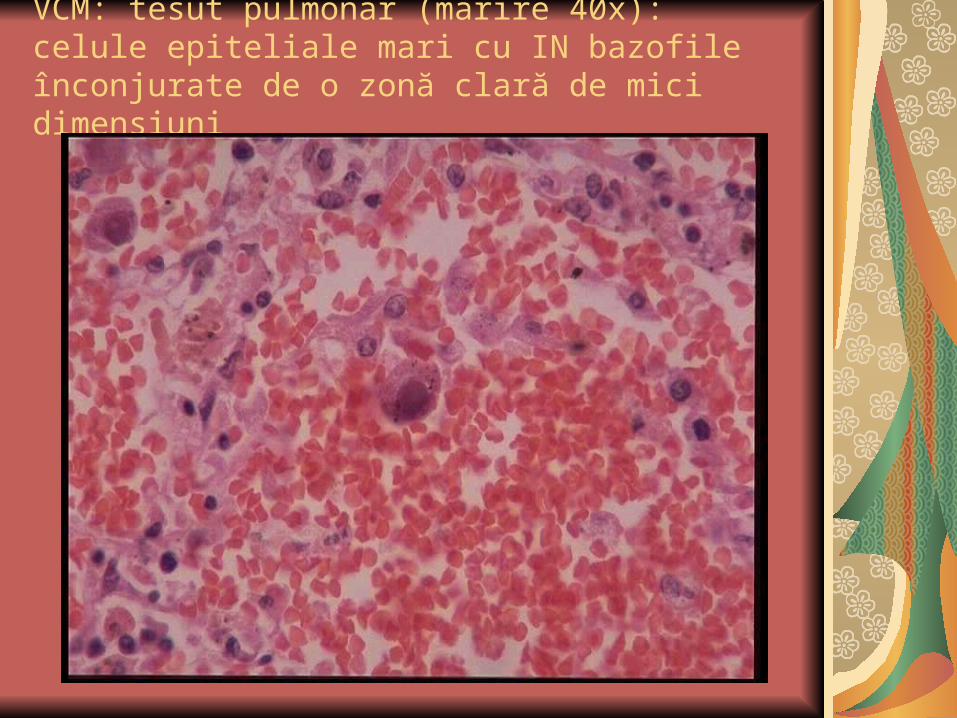
VCM: tesut pulmonar (marire 40x): celule epiteliale mari cu IN bazofile înconjurate de o zonă clară de mici dimensiuni
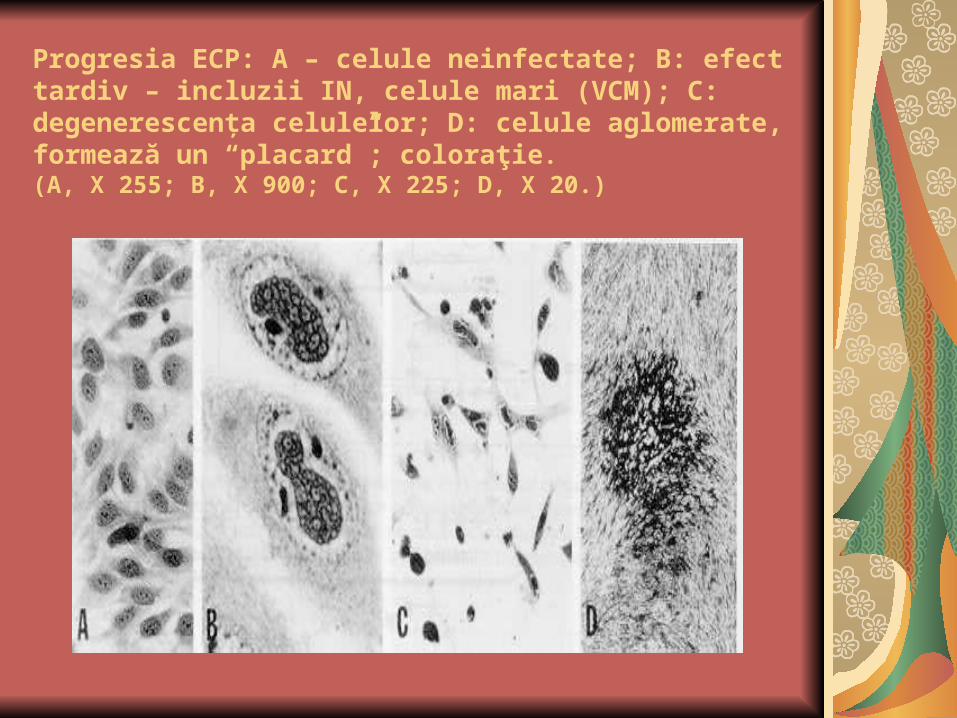
Progresia ECP: A – celule neinfectate; B: efect tardiv – incluzii IN, celule mari (VCM); C: degenerescenţa celulelor; D: celule aglomerate, formează un “placard”; coloraţie. (A, X 255; B, X 900; C, X 225; D, X 20.)
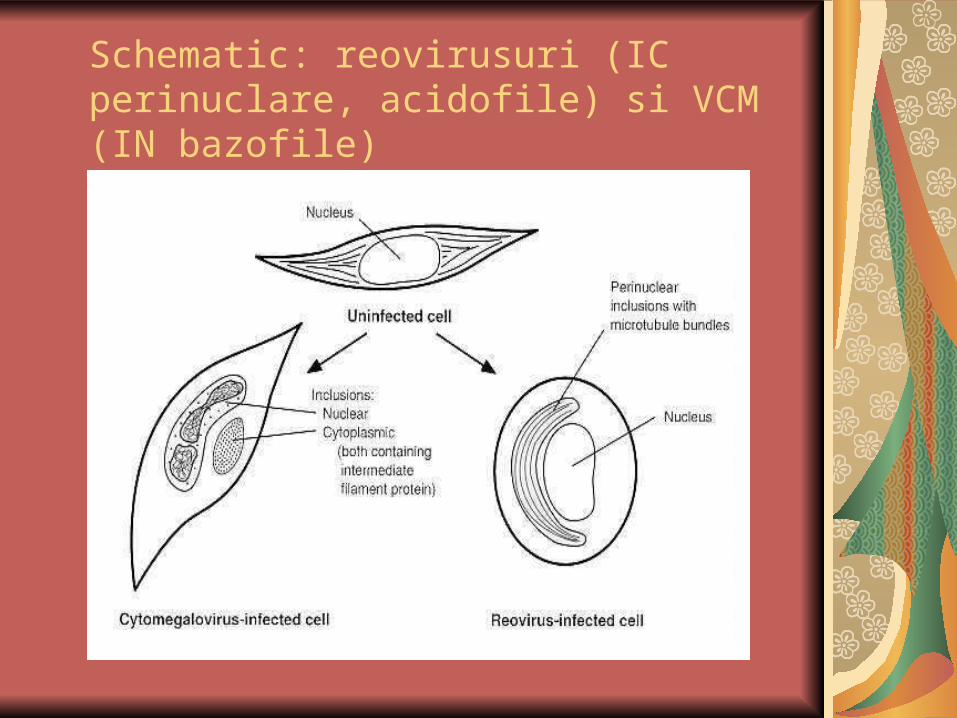
Schematic: reovirusuri (IC perinuclare, acidofile) si VCM (IN bazofile)
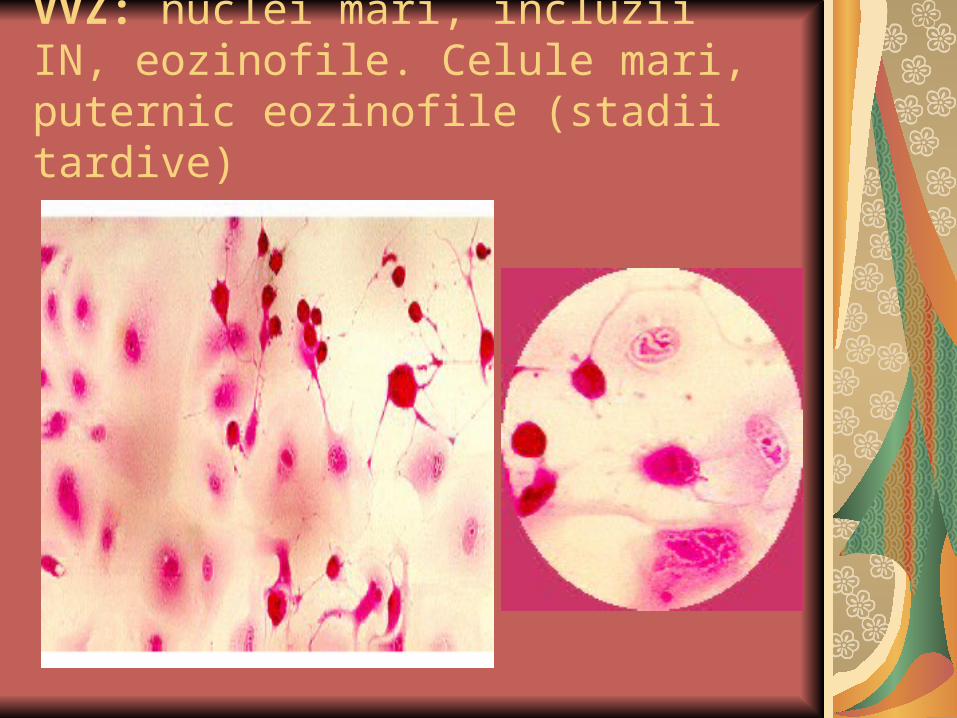
VVZ: nuclei mari, incluzii IN, eozinofile. Celule mari, puternic eozinofile (stadii tardive)
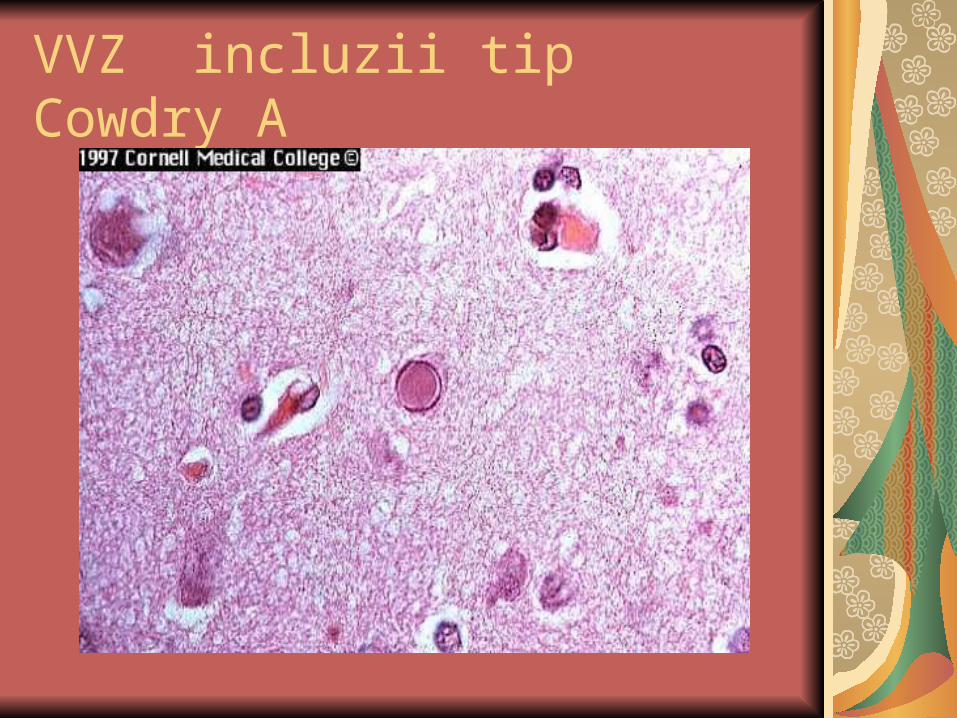
VVZ incluzii tip Cowdry A
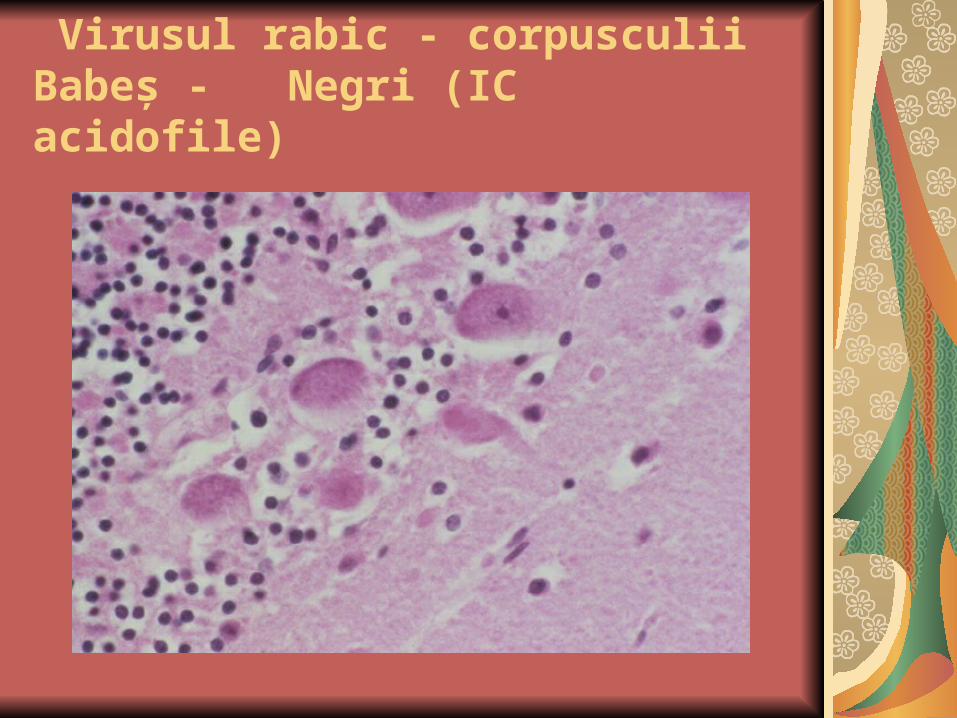
Virusul rabic - corpusculii Babeş - Negri (IC acidofile)
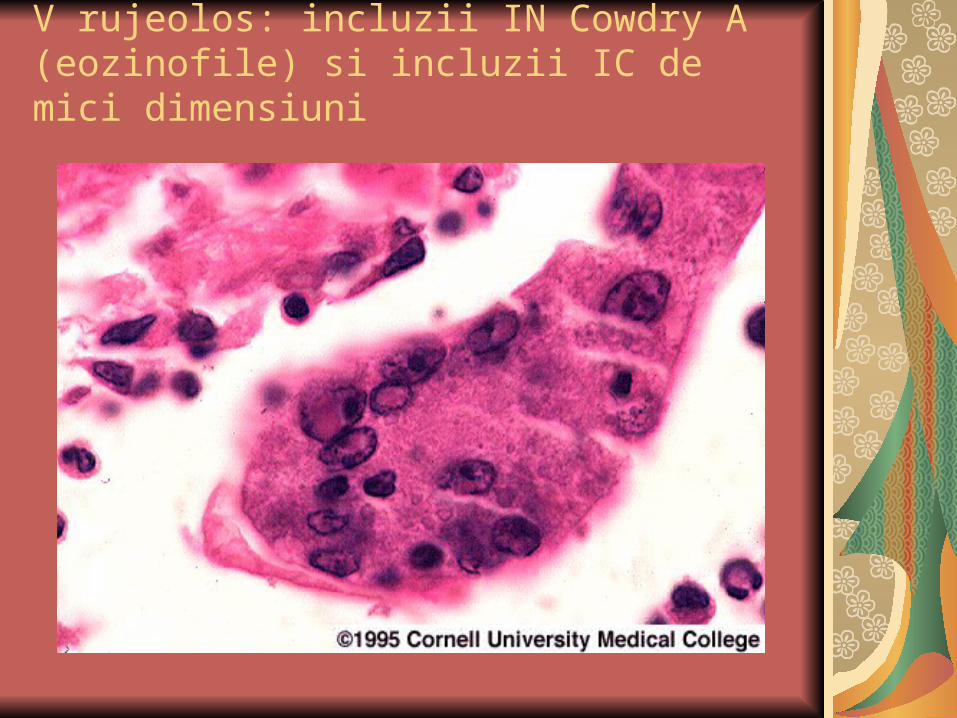
V rujeolos: incluzii IN Cowdry A (eozinofile) si incluzii IC de mici dimensiuni
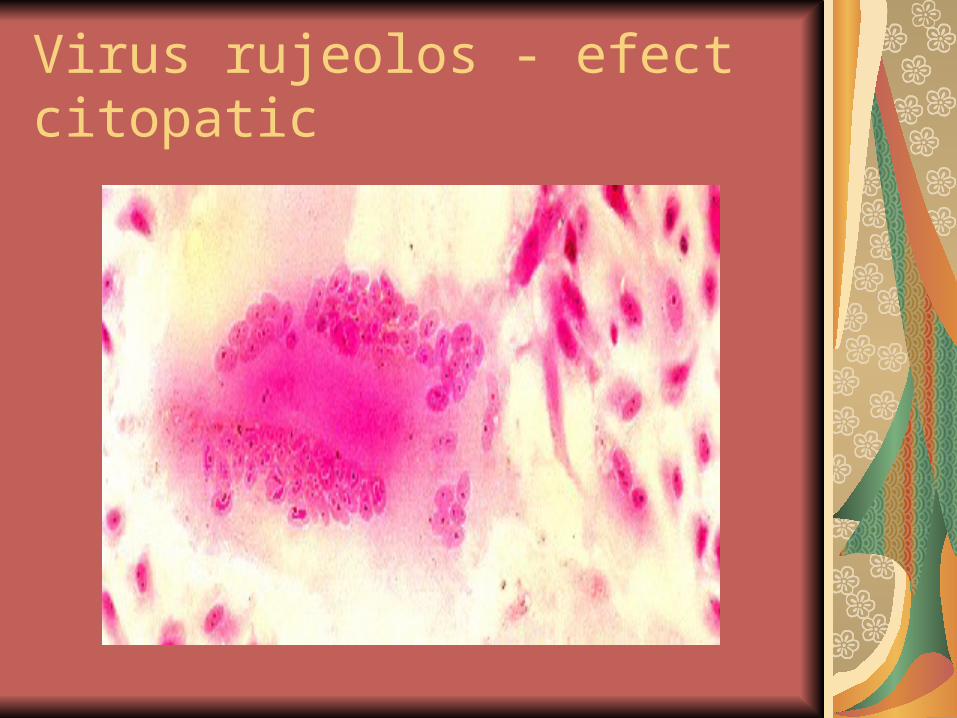
Virus rujeolos - efect citopatic
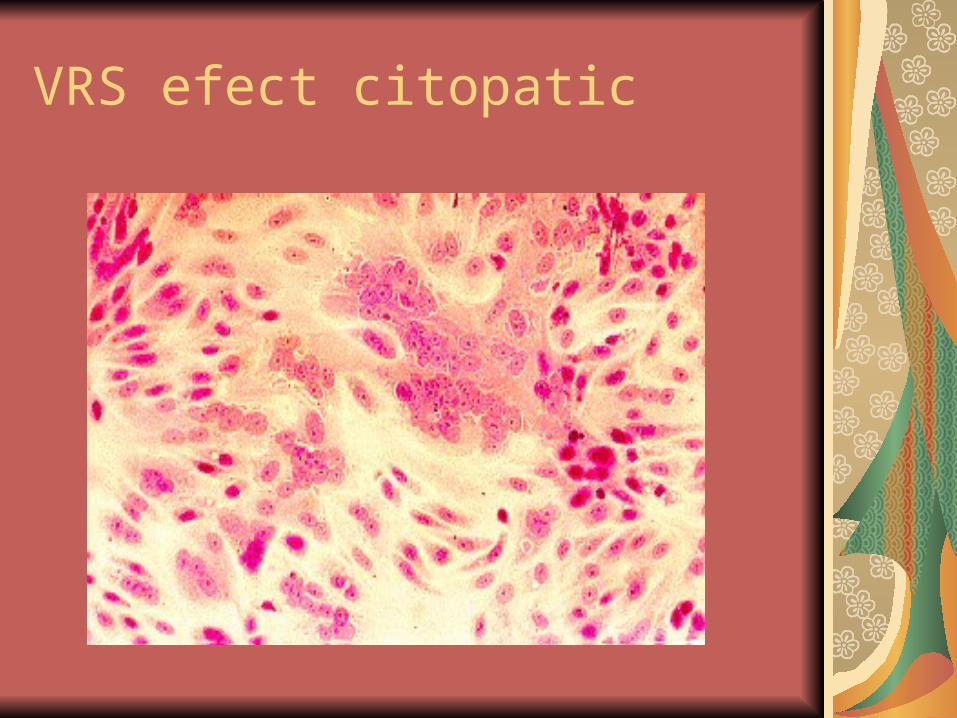
VRS efect citopatic
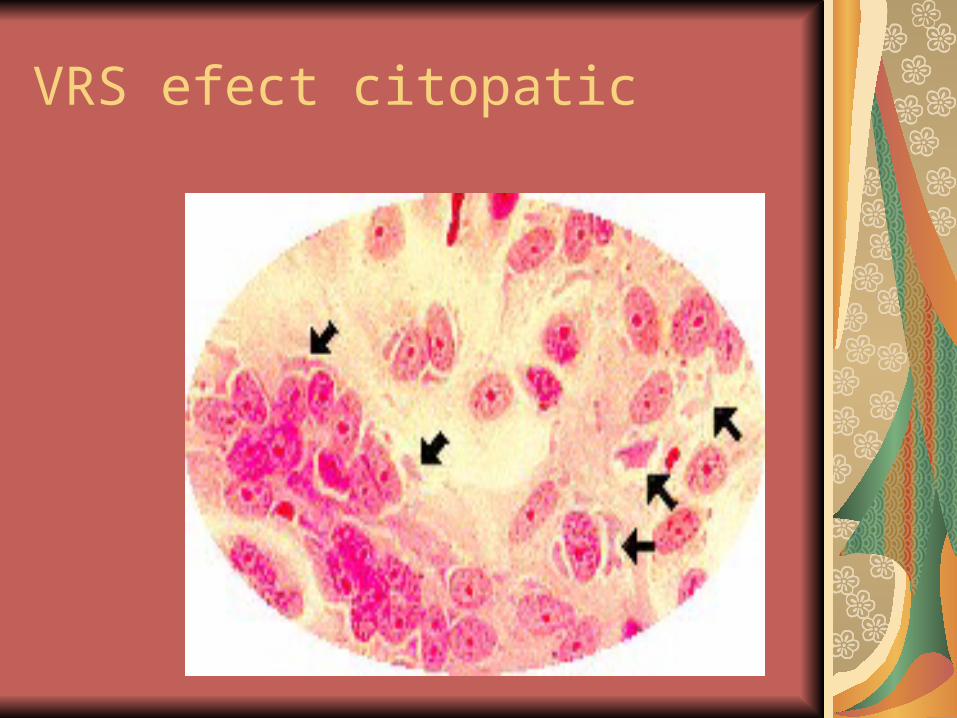
VRS efect citopatic







