OBIECTIVELE DE AMELIORARE LA SPECIILE DE LEGUME – VIE – POMI
90
OBIECTIVELE DE AMELIORARE LA SPECIILE DE LEGUME – VIE – POMI AMELIORAREA TOMATELOR Tomatele sunt legumele cele mai consumate din lume. Tomatele se cultivă în toate ţările, pe toate latitudinile, de la ecuator până aproape de cercul polar. Fructele pot fi consumate proaspete sau prelucrate. Industria alimentară propune produse foarte diverse: concenstrate, suc, ketchup, întregi, mărunţite, deshidratate. Şi modul de cultivare poate fi foarte variat. Astfel, tomatele destinate industrializării sunt cultivate în câmp, fără palisare, mecanizarea fiind foarte avansată, de la semănat la recoltat într-o singură fază. În California, mecanizarea culturii a fost pusă la punct odată cu selecţia unor soiuri adaptate din acest punct de vedere. Tomatele destinate consumului proaspăt pot fi cultivate în câmp în cultură palisată sau nu, sau chiar în spaţii protejate prin palisare. Adăposturile pot fi din plastic sau sticlă., încălzit sau nu, cultivarea făcându-se pe sol sau pe alţi suporţi. Tehnologiile cele mai performante pentru cultivarea în sere au fost puse la punct în Ţările de Jos, astfel, că producţiile la hectar pot depăşi 400-450 t. (PHILOUSE şi LATERROT, 1992) Extensia culturii în noi zone geografice, evoluţia tehnologiilor, diversificarea destinaţiei producţiei face necesară crearea unor soiuri cu adaptări speciale. În cursul ultimilor ani, cercetările s-au îndreptat asupra adaptării materialului vegetal la condiţiile de mediu, în special la cele stresante care au rol limitativ. Pe de altă parte, experienţele consumatorilor în privinţa calităţii fructelor sunt tot mai sporite, atât în ceea ce priveşte aspectul şi consistenţa fructelor, însuşirile organoleptice, nutritive, dar şi cele legate de industrializarea ia sfârşit, cu extensia culturii se asistă la o sporire a problemelor datorate bolilor, probleme care pot deveni cruciale în unele regiuni. Obiective de ameliorare
Transcript
OBIECTIVELE DE AMELIORARE LA SPECIILE DE LEGUME – VIE – POMI
AMELIORAREA TOMATELORTomatele sunt legumele cele mai consumate din lume. Tomatele se cultivă în toate ţările, pe toate latitudinile, de la ecuator până aproape de cercul polar. Fructele pot fi consumate proaspete sau prelucrate. Industria alimentară propune produse foarte diverse: concenstrate, suc, ketchup, întregi, mărunţite, deshidratate. Şi modul de cultivare poate fi foarte variat. Astfel, tomatele destinate industrializării sunt cultivate în câmp, fără palisare, mecanizarea fiind foarte avansată, de la semănat la recoltat într-o singură fază. În California, mecanizarea culturii a fost pusă la punct odată cu selecţia unor soiuri adaptate din acest punct de vedere. Tomatele destinate consumului proaspăt pot fi cultivate în câmp în cultură palisată sau nu, sau chiar în spaţii protejate prin palisare. Adăposturile pot fi din plastic sau sticlă., încălzit sau nu, cultivarea făcându-se pe sol sau pe alţi suporţi. Tehnologiile cele mai performante pentru cultivarea în sere au fost puse la punct în Ţările de Jos, astfel, că producţiile la hectar pot depăşi 400-450 t. (PHILOUSE şi LATERROT, 1992)
Extensia culturii în noi zone geografice, evoluţia tehnologiilor, diversificarea destinaţiei producţiei face necesară crearea unor soiuri cu adaptări speciale. În cursul ultimilor ani, cercetările s-au îndreptat asupra adaptării materialului vegetal la condiţiile de mediu, în special la cele stresante care au rol limitativ. Pe de altă parte, experienţele consumatorilor în privinţa calităţii fructelor sunt tot mai sporite, atât în ceea ce priveşte aspectul şi consistenţa fructelor, însuşirile organoleptice, nutritive, dar şi cele legate de industrializarea ia sfârşit, cu extensia culturii se asistă la o sporire a problemelor datorate bolilor, probleme care pot deveni cruciale în unele regiuni.
Obiective de ameliorareAmeliorarea productivităţii. Productivitatea, la tomate, ca şi la alte specii horticole, este o însuşire
deosebit de complexă. Ea depinde de elementele de productivitate, care sunt caracteristici fixate în baza ereditară a soiurilor, şi de interacţiunea acestora cu condiţiile de mediu.
Producţia de fructe pe plantă este un caracter polifactorial a cărui manifestare fenotipică este în strânsă dependenţă de tipul de creştere al plantei, de numărul fructelor pe plantă şi de greutatea acestora. Majoritatea soiurilor de tomate produc între 1,5-3,5 kg fructe pe plantă, în funcţie de intensitatea de manifestere a caracteristicilor enumerate anterior. La extragerea de elite, acestea vor trebui să aibă minimum 3 kg fructe pe plantă.
Producţia pe plantă este condiţionată şi de tipul de creştere. La formele cu creştere nedeterminată producţia nei plante poate fi foarte ridicată, dar cultivarea acestor forme necesită sisteme de susţinere. Din acest motiv formele de acest tip se cultivă în spaţii protejate, mai puţin în câmp. Pentru cultira în câmp se recomandă soiurile cu creştere determinată, la acetsea producţia fiind susţinută de desimi mai mari la unitatea de suprafaţă.
Numărul de fructe pe plantă şi greutatea acestora sunt caractere moştenite cantitativ. Numărul fructelor pe plantă este dependent de tipul plantei şi de tehnologia alicată la soiurile cu crestere nedeterminată. Majoritatea soiurilor prezintă fructe rotinde, având un diametru de 57-82 mm şi o greutate de 120-250 g.
La tomatele cultivate prin semănat direct, rezultatele experimentale obţinute de ARDELEAn şi colab. (1984), indică existenţa unor corelaţii fenotipice şi genotipice directe şi pozitive între numărul de fructe pe plantă şi producţia de fructe la unitatea de suprafaţă, fapt ce permite folosirea acestui caracter ca indice de selecţie pentru producţie.
Ameliorarea calităţii. Calitatea tomatelor pentru consumul curent este o însuşire ce trebuie analizată în funcţie de destinaţia soiului sau hibridului. Forma fructelor, culoarea, textura şi fermitatea pulpei, gustul, conţinutul în substanţă uscată, în zahăr şi aciditate, în vitamina C etc. sunt numai câţiva din factorii ce trebuie luaţi în considerare în aprecierea tomatelor destinate diferitelor întrebuinţări.
În general, fructele rotunde, uşor aplatizate. În ultima vreme s-au constatat preferinţe faţă de fructele alungite sau faţă de cele mici, de tip „cireaşă” (10-12 g greutate). Atât forma cât şi greutatea fructelor sunt caractere cu eritabilitate mare.
Culoarea fructelor este prezintă un determinism simplu şi se datorează culorii pulpei. Cele mai multe soiuri au fructele de culoare roşie, fructele mai intens colorate fiind preferate de către consumatori.
Unele mutante au epiderma incoloră astfel că fructul are culoare roz. Alte mutante au fructele galbene sau portocalii. Un alt aspect important este omogenitatea coloraţiei fructului. Aceasta este în mare parte dependentă de generaţia unei gene. Alela “u+” determină prezenţa unui colet (zonă în jurul locului de inserţie a fructului la peduncul) de culoare verde înainte de maturitate. Alela n caracterizează o coloraţie uniformă. Cultivarele care prezintă gena “u+”, la coacere pot avea o coloraţie uniformă sau pot păstra o pată verde la nivelul coletului. De asemenea, ele se pot coace neuniform, mai ales sub influenţa temperaturilor ridicate. Coletul devine galben şi dur, ceea ce duce la deprecierea fructelor. Acest defect este foarte rar la cultivarele care prezintă gena “u”.
Forma fructelor la tomate este destul de variată, cel mai des întâlnite fiind fructele de formă globuloasă, ovală şi piriformă. BARTON şi colab. (1955) consideră forma rotundă (“O”) dominantă asupra celei ovale sau piriforme (“o”). Cum însă între formele ovală şi piriformă se pot obţine câteva forme intermediare distincte, se pare că ipoteza eredităţii monogenice a formei fructului nu este suficientă.
Textura şi fermitatea pulpei sunt caracteristici cu un determinism poligenic şi cu grade de heritabilitate destul de scăzute. Data maturizării, lungimea perioadei de maturizare a fructelor, gradul de maturizare a acestora în momentul recoltării sunt doar câţiva din factorii ce influenţează foarte puternic textura şi fermitatea pulpei fructului la tomate. (CROITORU şi TOMA, 2002)
Pentru ameliorarea formităţii fructului cele mai multe lucrări s-au desfăşurat în SUA unde scopul acestora a fost de a se acumula la număr cât mai mare de gene implicate în realizarea acestui caracter. Au fost create chiar grade pentru măsurarea durităţii fructelor, a rezistenţei la şocuri (lovituri) sau a fermităţii pericarpului (penetrometre).
Gustul este dat, şi la tomate, de raportul dintre diferitele substanţe aflate în fruct , dintre care conţinutul în glucide şi aciditatea joacă un rol esenţial. Ambele componente se moştenesc după modelul cantitativ şi, ceea ce este mai important, independent unul de altul.
Dintre acizii organici, conţinuţi de fructele de tomate, un rol deosebit revine acidului ascorbic (vitamina C). Sporirea conţinutului de vitamina C în fructele de tomate constituie, multă vreme, un obiectiv important de ameliorare la această plantă.
O sursă valoroasă pentru un conţinut ridicat de vitamina C o constituie unele forme ale speciei L.peruvianum (peste 75 mg vitamina C la 100 g fruct). Această specie a fost folosită, încă din 1946 (YEAGER şi PURINTON) ca genitor în vederea obţinerii unor descendenţi bogaţi în vitamina C, primul rezultat palpabil al acestui program fiind soiul Hi C care are un conţinut în vitamina C aproape dublu faţă de celelalte soiuri de tomate cunoscute (62-67 mg/100 g fruct). Din păcate, conţinutul ridicat în vitamina C este corelat negativ cu mărimea fructului, fapt verificat de majoritatea amelioratorilor care au lucrat cu această plantă. Încercările făcute de POLE (1956) prin utilizarea metodei ameliorării convergente au dus la câteva rezultate promiţătoare: selecţii cu un conţinut relativ ridicat în vitamina C (peste 40 mg) şi cu dimensiuni mari a fructelor (70 g). Aceste prime rezultate sugerează că prin aplicarea unor metode de ameliorare adecvate, likage-ul dintre conţinutul ridicat de vitamina C şi dimensiunea redusă a fructelor poate fu rupt obţinându-se descendenţi cu fructe mari şi bogate în vitamina C.
Datorită consumului în cantităţi mari a acestei legume , tomatele nu sunt de neglijat în privinţa
aportului de săruri minerale şi vitamine, mai ales A şi C. În unele ţări, cum este Japonia, ameliorarea este concentrată spre creşterea conţinutului în vitamina C, caracter pentru care în cadrul genului Lycopersicon există o mare variabilitate. În alte ţări, ca Bulgaria, se urmăreşte sporirea concentraţiei în caroten sau vitamina A, folosind mutanta monogenică dominantă B, bogată în β caroten, care provoacă o coloraţie portocalie a fructelor.
Calitatea tomatelor pentru industrializare depinde de conţinutul în substanţă uscată solubilă din fructe este un caracter important pentru cultivarele a căror fructe sunt destinate producerii de concentrate. Acest caracter a fost contra-selecţionat pe parcursul creării cultivarelor foarte productive adaptate la cultivarea şi recoltarea în condiţii de mecanizare totală: dezvoltare vegetativă redusă, maturizarea precoce şi grupată, productivitate mare, fructe mari şi foarte tari cu pereţi îngroşaţi şi cu zonele loculare reduse, toate aceste caractere fiind corelaţie negativă cu un conţinut ridicat de substanţă uscată. În aceste condiţii nu se pot atinge 5-5,5% substanţă uscată, ci doar 4,5%. Progrese substanţiale au fost realizate pornind de la încrucişări cu anumite specii sălbatice, existând astăzi şi cultivare cu 5,5-6% substanţă uscată. În aceste condiţii nu se pot atinge 5-5,5% substanţă uscată, ci doar 4,5%. Progrese substanţiale au fost realizate pornind de la încrucişări cu anumite specii sălbatice, existând astăzi şi cultivare cu 5,5-6% substanţă uscată. Se deschis noi perspective graţie utilizării markerilor moleculari . punerea în evidenţă a unei legături între un caracter cantitativ complex aşa cum este conţinutul în substanţă uscată solubilă, care are şi o ereditate scăzută, ca markeri RFLP şi localizarea unor astfel de caractere în harta cromozomială, trebuie să faciliteze transferul unui astfel de caracter în mai multe cultivare. Astfel de studii au fost efectuate pe un material obţinut din încrucişări interspecifice cu L.chimielewski la care conţinutul de substanţă uscată solubilă este în jur de 10% (OSBORN şi col., 1987; TANKSLEY şi col., 1988)
Pe lângă procentul de substanţă uscată, fructele tomatelor pentru industrializare trebuie să fie bogate în zahăr (4-5%), vitamina C şi caroten, iar aciditatea mia redusă. (GLĂMAN şi col., 2001)
Vâscozitatea este o caracteristică ce interesează pentru produsele ce obţinute în urma transformării, în special pentru concentrate, suc, ketchup. Vâscozitatea este direct legată de conţinutul fructelor în substanţe insolubile în alcool (proteine, celuloză, pectine, polizaharide).
PH-ul produselor realizate în urma industrializării trebuie să fie sub 4,5 pentru a limita timpul de sterilizare necesar şi pentru a se păstra calitatea produsului.
Ameliorarea duratei păstrării fructelor. În privinţa duratei de păstrare a fructelor există diferenţe mari între soiuri. O ameliorare spectaculoasă a duratei de păstrare ce poate obţine utilizând mutante monogenice legate de maturizarea fructelor; genele “rin” (repening inhibitor) sau “nor” (non ripenin) (TIGCHELAAR şi col., 1978). Fructele plantelor rin/rin sau nor/nor nu sunt climaterice. Procesul de maturizare a fructelor climaterice a unui soi normal are loc în condiţiile blocării producţiei de etilenă, a respiraţiei, a sintezei carotenoizilor, a înmuierii fructelor şi a dezvoltării aromelor. Fructele rămân galbene şi se pot păstra mai multe luni la temperatura mediului ambiant, dar gustul lor este neplăcut.
Fructele plantelor de tip nor/+ se pot păstra câteva săptămâni, iar dezvoltarea coloraţiei în roşu este lentă şi incompletă. La heterozigoţii de tip rin/+, coloraţia fructului este normală şi durata păstrăririi nu este decât la câteva zile. Amelioratorii, în special în Israel, exploatează aceste gene mutante în cadrul hibrizilor, încercând să atenueze defectele pe planul coloraţiei şi a gustului.
Ameliorarea epocii de coacere. Epoca de coacere constituie un obiectiv de ameliorare de primă importanţă având în vedere faptul că este necesară acoperirea unei perioade de timp cât mai îndelungată cu tomate de calitate superioară pentru consumul în stare proaspătă. Din punct de vedere a epocii de coacere, soiurile de tomate din ţara noastră se clasifică în trei grupe principale: precoce (95-100 zile), semirprecoce (111-130 zile) şi tardive (peste 130 zile). Condiţiile climatice din ţara noastră permit cultivarea unor soiuri de tomate aparţinând oricăreia din clasele enumerate mai sus. Desigur, obţinerea unor soiuri de tomate de câmp timpurii şi extratimpurii constituie obiectivul major al cercetărilor legate de epoca de coace la această plantă, mai ales dacă se are în vedere că datorită crizei resurselor de energie, producţia de tomate în sere şi solarii încălzite nu va mai fi rentabilă.
În selecţie, pentru o anumită epocă de coace a fructelor de tomate, deosebit de importantă este corelaţie pozitivă dintre data înfloritului şi data maturităţii fructelor. Această corelaţie permite efectuarea selecţiei, în cadrul populaţiilor F2 şi următoarele încă înainte de plantarea lor în câmp. În acest scop, răsadul este ţinut în seră sau răsadniţe până în momentul deschiderii primelor flori. În funcţie de epoca de
coacere pentru care vrem să facem selecţie, încă din răsadniţă pot fi eliminate toate plantele care au o înflorire prea precoce sau prea tardivă.
Ameliorarea rezistenţa la boli şi dăunători. Tomatele fiind cultivate în condiţii diverse şi uneori în condiţii limitante pentru creştere şi dezvoltare, numărul agenţilor patogeni ce afectează această specie este foarte mare. Sunt aproape 200 de boli descrise la tomate, la nivel mondial.
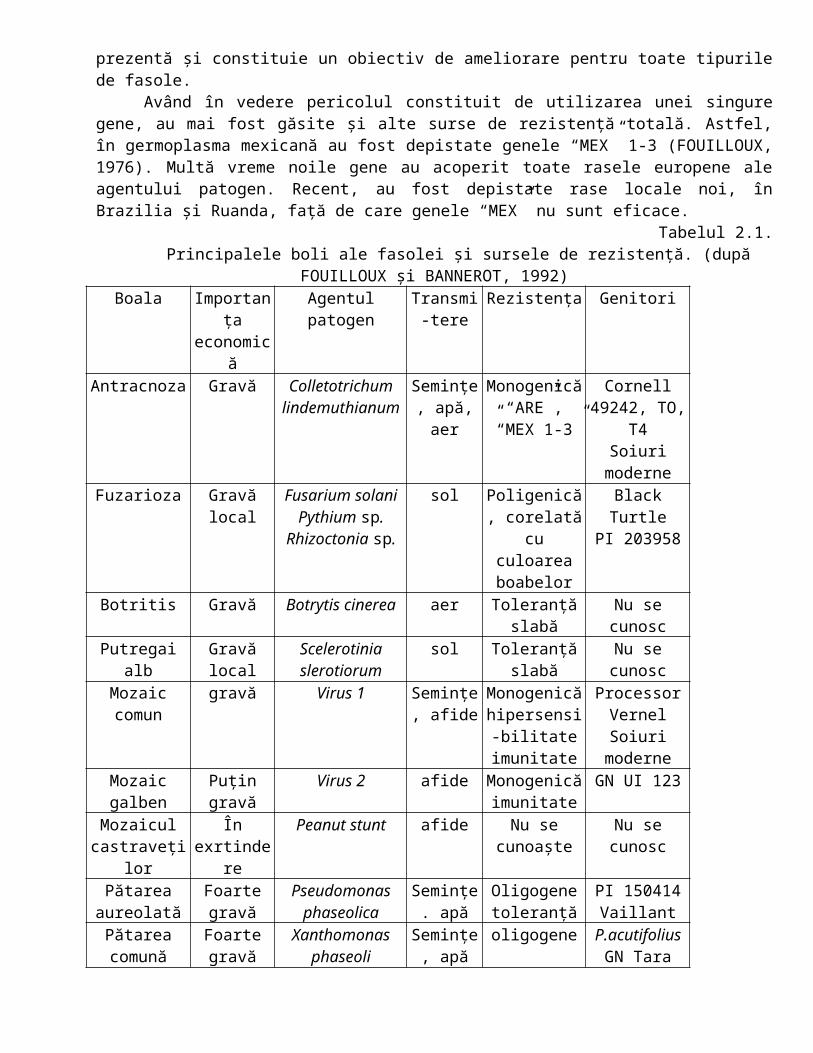
Începând cu anii 1940, cercetările pentru depistarea unor surse de rezistenţă s-au intensificat foarte mult, la început în SUA, apoi în Europa. Speciile sălbatice au fost foarte mult exploatate. După 1955 a fost elaborat primul bilanţ a resurselor disponibile (ALEZANDER şi HOOVER, 1955). Utilizarea acestora a fost mai mult cerută şi după 1960 au apărut primele soiuri reformante şi rezistente la mai multe boli. Peste 10 boli sunt controlate astăzi de surse de rezistenţă. Toate sunt monogenice, şi în cea mai mare parte dominante. Eficacitatea lor este valabilă în ceea ce priveşte nivelul expresiei şi stabilitatea (tabelul 3.2.)
Tabelul 3.2.Stadiul luptei genetice contra agenţilor patogeni de la tomate în ţările mediteraneene (după
PHILOUZE şi LATERROT, 1992)Agent patogen (boala) Gena de
rezistenţăNivelul expresiei
rezistenţeiStabilitatea rezistenţei
în timpCiuperci
Verticillium dalilie
Fusarium oxysporum f.sp. lycopersici
Rasa 0
Rasa 1 Fusarium oxysporum f.sp. radicis
lycopersiciPirenochaeta lycopersici
Cladosporium fulvum (Fulvia fulva)
Phytphtora infestansStemphylum sp.
Bacterii Pseudomonas tomato
ViruşiVirisul mozaicul tutunului (VMT)
Nematozi Meliodogine spp.
Ve
I
I-2
Fr 1pyl
Cf serie de genePh 2Sm
Pto
Tm-22
Mi
Foarte ridicat
Foarte ridicat
Foarte ridicat
Foarte ridicatMedie
Foarte ridicat
Medie Foarte ridicatră
Foarte ridicat
Foarte ridicată
Foarte ridicată
Bună, depăşită în mmulte zone
Medie, depăşită în unele zoneBună
Bună, recent introdusăBună
Medie, unele gene au fost depăşite
Bună Bună
Bună
Bună
Bună, depăşită în unele zone
Faţă de Clamidosporium fulvum situaţia este mai complexă. Această ciupercă evoluează foarte rapid în culturile protejate din Europa şi bazinul Mediteranean, fiind cunoscute peste 8 rase (LATERROT, 1990). Rezerva de gene de rezistenţă este importantă. Programele de ameliorare actuale tind să introducă cel puţin 2 gene, încă nedepăşite, în noii hibrizi F1, în fine opunând ciupercii o barieră mai eficace.
Dacă faţă de virusul mozaicului tutunului gena de hipersensibilitate Tm – 22 este la baza tuturor soiurilor rezistente actuale, trebuie precizat că rezistenţa poate să apară la 3 tipuri de genotipuri:
- Tm – 22 în stare homozigotă, - Tm – 22 în stare heterozigotă, - Tm – 22 în stare heterozigotă cu gena de toleranţă Tm – 1. (LATERROT, 1990)În unele ţări subrtopicale şi tropicale, bacteria Pseudomonas solanocearum provoacă pagube
considerabile. Pentru aceasta au fost puse în evidenţă mai multe surse de rezistenţă. Rezultate bune au fost obţinute cu rezistenţa indusă la soiul “Caracibe”, rezistenţă care provine de şa tomate primitive, această rezistenţă dde tip semidominant la hibrizii F1, s-a exprimat în toate ţările în care a fost studiată.
Cele mai păgubitoare boli ale tomatelor, în condiţiile din ţara noastră sunt: septorioza (Septoria licopersici) mana (Phitophtora infestans), pătarea brună a frunzelor (Cladosporium fulvum) şi virusurile. Combaterea pe cale chimică a acestor boli este destul de costisitoare, iar în cazul unor viroze, ea nu este nici măcar posibilă. Din aceste motive, crearea de soiuri şi hibrizi de tomate cu o rezistenţă genetică la aceste boli se impune ca un obiectiv de ameliorare de prim rang.
Principalele surse de rezistenţă la boli, folosite în ameliorare tomatelor, sunt speciile înrudite cu L.esculentum. Multe forme ale speciei L.peruvianum sunt rezistente la mană, pătarea brună şi mozaicul tutunului, iar subspecia pimplinellifolium este deosebit de rezistentă la fuzarioză.
Datele cuprinse în literatura de specialitate sunt destul de sărace în privinţa mecanismului ereditar al transmiterii rezistenţei la aceste boli. Au fost identificate trei gene majore, Cf1, Cf2 , Cf3 care conferă rezistenţă la pătarea brună a frunzelor (BARTON şi colab., 1995). De asemenea, PORTE (1941), obţine în experienţele sale dovezi evidente că şi rezistenţa la fuzarioză ar fi determinată de o singură genă majoră, dominantă.
Rezistenţa la mană pare a fi moştenită cantitativ (CLARKE, 1955) şi este, foarte adesea corelată negativ cu mărimea fructelor. Se pare că tot o corelaţie negativă există şi între rezistenţa la mozaicul tutunului şi mărimea fructelor. Pentru obţinerea unor descendenţi rezistenţi la aceste boli, în afara speciei L.peruvianum mai pot fi folosite ca surse de rezistenţă soiurile Kirov I, Smorovindnaia, care sunt practic imune la atacul de mană şi prezintă de asemenea o bună rezistenţă la viroze.
Amelioratorii mizează să acumuleze maximum de rezistenţă deja cunoscute şi folosite împreună cu caracterele agronomice şi de calitate a fructelor. În paralel au fost dezvoltate programe de ameliorare pentru a introduce în cultivare rezistenţe la boli încă necontrolate genetic sau la noi rase fiziologice adaptate la genele actuale exploatate. Pe plan mondial, pe tipuri de agenţi patogeni, pot fi menţionate mai multe programe.
Principalele programe de ameliorare a rezistenţei la boli criptogamice vizează următorii agenţi patogeni: Verticillium dablie (începutul exploatării rezistenţei parţiale la rase adaptate la gena Ve), Fusarium oxysporium f.sp. lycopersici (exploatarea rezistenţei de nivel înalt controlată de genele I3
preluate de la L. pennellii, eficace faţă de rasele adaptate la I şi I2), Leveillula taurica – Oidiopsis taurica (începutul exploatării unei rezisteneţ de înalt nivel controlată de gena Lv preluată de la L. chilense), Oidium licopersicum (începutul introducerii rezistenţei de înalt nivel preluată de la 25 specii sălbatice), Alternaria dauci f. sp. solani (exploatarea rezistenţei parţiale poligenice)
În privinţa atacului de baterioze sunt luaţi în studiu două specii: Corynebacterium michiganenese - Clavibacter michiganensis ssp. michiganensis (exploatarea rezistenţei parţiale oligo şi poligenice), Xanthomonas campestris pv. vesicatoria (exploatarea rezistenţei parţiale)
Principalele viroze urmăîrite a fi controlate genetic sunt: Virusul mozaicului castraveţilor (CMV), transmis de afide (începutul exploatării unei rezistenţe parţiale preluată de la L. hirsutum ), Virusul Y al cartofului (PVY) transmis de afide (exploatarea unei rezistenţe mono sau oligogenice preluată de la L. hirsutum), Virusul tomato spotted wiet (TSWV) transmis prin tripşi (începutul exploatării rezistenţei de la L. peruviannum), Virusul tomato Yellow Leaf Curl (TYLCV) transmis de Bemisia tabaci (exploatarea rezistenţei parţiale de la mai multe specii sălbatice).
Rezistenţa faţă de agenţii patogeni este pusă în evidenţă prin mijloace de încercare sistematică cu infecţii artificiale cu una sau mai multe suşe ale agenţilor patogeni de diferite origini sau în câmp în zone cu contaminare naturală foarte severă.
Pentru a sublinia importanţa observării materialului biologic în câmp, prezentăm 2 exemple. În Canada, existenţa unei epidemii de Pseudomonas tomato a evidenţiat rezistenţa la această bacterie a mai multor linii de tomate. Această rezistenţă s-a datorat unui părinte comun acelor linii, aparţinând speciei L. pimpinellifolium. În Bulgaria, o puternică epidemie de Laveillula taurica a permis găsirea unei rezistenţe într-un material obţinut prin încrucişarea cu L. chilense.
Tehnicile de alegere a materialului cu o anumită rezistenţă depind de tipul agentului patogen şi de expresia rezistenţei. Succesul şi aplicarea pe perioade lungi de timp aşa cum doresc amelioratorii, depinde, în cazurile mai dificile, de colaborarea între amelioratori şi fitopatologi.
În ceea ce priveşte cele aproximativ 10 rezistenţe integrate în cultivarele actuale şi populaţiile în curs de selecţia se poate considera că pentru două dintre ele există dificultăţi. Uneori este vorba de rezistenţele la Pyrenophora lycopersici şi Phztophtora infestans pune probleme de alegere, în cazul lor nivelul expresiei fiind foarte slab şi dependent de condiţiile mediului de testare în stadiu de plantulă. În alte cazuri, tehnicile sunt relativ mai simple.
Dintre insecte, singura specie pentru care sunt posibilităţi de ameliorare este Liriomyza. Pentru acest dăunător a început exploatarea rezistenţei parţiale de la speciile L. hirsutum şi L. cheesmanii.
Ameliorarea rezistenţa la frig. În calitatea lor de plante termofile tomatele sunt foarte sensibile la frig. Majoritatea soiurilor de tomate cunoscute în prezent îşi încetinesc sau îşi încetează creşterea şi dezvoltarea la temperaturi pozitive cuprinse între 5 şi 70C.
Crearea de soiuri şi hibrizi de tomate rezistente la frig se impune ca un obiectiv important de ameliorare. Astfel de soiuri ar aduce câştiguri din două puncte de vedere:
- plantarea mai timpuri e răsadului în câmp, aceasta atrăgând economii de energie şi fonduri,- extinderea culturii tomatelor în zone cu primăvari şi veri mai răcoroase.Surse valoroase de rezistenţă la frig sunt soiurile Stambovîi 2165, Karlic, VIR 2116 şi specia
L.hirsutum. Ameliorarea adaptabilităţii. Este vorba de crearea unor soiuri adaptate la diverse moduri de
cultură, la condiţii climatice şi pedologice extrem de variate, în maniera ameliorării productivităţii şi stabilităţii, a prelungirii perioadei productive şi la extinderea tomatelor în noi zone.
Lucrările efectuate în nordul Europei, în special în Olanda, au permis crearea unui material vegetal adaptat la condiţiile zi scurtă şi la lumină puţină, chiar dacă tomatele sunt plante pretenţioase faţă de lumină. Astfel au fost create cultivare capabile să producă întreg anul cu randamente extrem de ridicat.
Lucrările asupra rezistenţei la salinitate trebuie să permită creşterea productivităţii şi în zonele unde salinitatea solului sau a apei este o problemă. Terenul de salinitate cuprinde o foarte mare heterogenitate legată de natura şi concentraţia diferiţilor ioni minerali din sol sau apă. În plus, acestea prezintă fluctuaţii în spaţiu şi timp (JONES, 1987). Rezistenţa la salinitate a fost pusă în evidenţă, pe de o parte la L. esculentum în cadrul cultivarelor cu origine din zone aride (Egipt, Turcia) şi pe de altă parte la un număr mare de forme sălbatice, în particular L. chesmanii, sp. L. esculentum.
Între soiurile speciile L. esculentum există diferenţe mari în privinţa calităţii polenului şi a sensibilităţii la legarea fructelor la temperaturi scăzute. Un astfel de material a fost folosit mult la genitori, dar nu sunt speranţe prea mari din acest punct de vedere genitorii cei mai promiţători se pot găsi la speciile sălbatice.
Anumite ecotipuri din L. hirsutum, care cresc la altitudine, rezistă la frig, nu numai în privinţa calităţii polenului şi a legării fructelor, dar şi în privinţa germinării seminţelor şi creşterii.
O altă cale pentru ameliorarea legării la temperaturi scăzute este exploatarea partenocarpiei naturale. Există mai multe origini, găsite la L. esculentum sau la specii sălbatice din genul Lycopersicom, cel mai uşor de exploatat şi datorată genelor recesive pat-2 (PHILOUZE, 1983). Este vorba despre o partenocarpie facultativă: când condiţiile de mediu sunt favorabile microsporogenezei în urma polenizării şi fecundării se formează fructe cu conţinut normal de seminţe; dar când condiţiile devin nefavorabile se formează fructe partenocarpice, cu aspect şi mărime asemănătoare fructelor cu seminţe. Acest caracter a fost obiectul unui număr mare de lucrări şi programe de ameliorare, dar până în prezent nu au fost create soiuri partenocarpice foarte performante. Totuşi, această cale rămâne deschisă.
Bazele fiziologice care conferă o bună comportare la temperaturi ridicate sunt adesea diferite de la un soi la altul; stigmatul nu depăşeşte vârful staminelor la temperaturi ridicate, producţia de polen este suficientă, gameţii femeli şi masculi au o bună viabilitate, aptitudinea la partenocarpie, aptitudinea de a transfera produşii fotosintezei către organele de reproducere. Aceste elemente sunt vizate a fi acumulate într-un singur genotip prin programe de ameliorare complexe (STEVENS, 1979).
Adaptabilitatea pentru cultura forţată. De la început trebuie menţionat faptul că nu toate soiurile de câmp dau rezultate în culturile din spaţii acoperite. Soiurile pentru aceste spaţii prezintă unele însuşiri speciale:
- să aibă o bună rezistenţă la condiţiile de mediu specifice serelor şi solariilor unde este deficit de lumină, exces de umiditate şi variaţii diurne de temperatură
- să prezinte o foarte bună rezistenţă la atacul bolilor specifice spaţiilor acoperite;- să prezinte fructe intens colorate, uniforme ca mărime, cu pulpa tare, rezistente la transport şi
dacă este posibil să se maturizeze după recoltare.În ultima vreme asemenea soiuri şi hibrizi au fost creaţi şi introduşi în cultură. Soiurile pot servi
ca genitori în programele de ameliorare.Obţinerea de soiuri cu tulpină erectă şi creştere determinată constituie un obiectiv de ameliorare
tot mai prezent în ultima vreme datorită extinderii culturilor de tomate în câmp şi necesităţii mecanizării complete a lucrărilor de întreţinere şi chiar recoltare.
Soiurile şi hibrizii de tomate destinaţi a fi cultivaţi în câmp trebuie să prezinte anumite însuşiri, în principal: creştere determinată, tulpini erecte şi puternica care să nu necesite sisteme de susţinere, fructificare şi maturizare simultană a fructelor de pe două până la patru etaje, o desprindere uşoară a fructelor din codiţă.
Dintre aceste caractere, numai pentru câteva se cunoaşte mecanismul determinismului genetic. Pentru creşterea determinată se cunoaşte gena majoră recesivă “sp” diferită de gena “d”, care în stare homozigotă recesivă produce piticirea plantelor. Celelalte caractere de interes din acest punct de vedere au un determinism genetic poligenic cu stabilitate foarte slabă.
Soiurile şi hibrizii destinaţi culturilor de câmp în condiţiile mecanizării complete, îndeplinesc numai o parte din caracterele necesare. Principalul caracter care este deficitar este portul neerect, ceea ce face ca coacerea fructelor să fie neuniformă şi recoltarea să se facă cu greutate.
AMELIORAREA ARDEIULUI
Ardeiul a fost adus pe contionentul European după descoperirea Americii, la început în Spania şi Portugalia, apoi în Germania, Anglia şi Ungaria. Pe la începutul secolului XIX, ardeiul pătrunde din Asia Mică în sudul Rusiei şi în ţările balcanice. În aceiaşi perioadă această plantă pătrunde în ţara noastră, la începul în sudul ţării, apoi şi în celelalte zone.
În ţara noastră zonele foarte favorabile culturii ardeiului sunt c ele din Câmpia Română şi Câmpia de Vest, favorabile sunt cele din Câmpia şi Podişul Transilvaniei şi partea de est a Moldovei. În zonele mai răcăroase, cultura găseşte condiţii mai puţin prielnice (HORGOŞ, 2000).
Este foarte inetersant faptul că în fiecare ţară există variatăţi favorite, dar peste tot sunt cunoscute toate varietăţile, iar ameliorarea le cuprinde pe toate. În ţările cu nivel înalt de dezvoltare ardeiuol este foarte răspândit, fiind consumat crud, gătit sau conservat. Conservarea face ca ardeiul să fie o materie primă pentru industria alimentară. Conservarea devine tot mai importantă, formele sub care s epot conserva fructele fiind variate: fructe întregi sau feliate, fermentate sau în oţet. În unele ţări (Spania, Bulgaria, Ungaria) ca şi în ţara noastră ardeiul este un condiment şi colorant sun formă de pudră sau extras uleios.
Producţia mondială este crescândă, fiind estimată la peste 10 milioane tone obţinute pe o suprafaţă de peste 1 milion hectare. Apăroape jumătate este realizată în Asia. În ţările tropicale, ardeiul este un principal obiect al comerţului.
Creşterea producţiei mondiale se datorează fptului că prin introducerea în cultură a hibrizilor, producţiile la unitatea de suprafaţă au crescut foarte mult.
Obiective de ameliorare.
Obiectivele de ameliorare la ardei sunt complexe, fiind influenţate de sistemul de cultură.
Ameliorarea capacităţii de producţie. Capacitatea de producţie este un caracter complex, rezultat a mai multor componente morfologice: gradul de ramificare a plantei, numărul florilor, mărimea fructelor, grosimea pulpei, greutatea fructelor. Toate aceste caractere sunt specifice fiecărei varietăţi şi în legătură strânsă cu condiţiile de creştere şi dezvoltare.
Capacitatea de producţie, este în general ridicată. Ardeiul iute are o înflorire mult mai abundentă, dar fructele sunt mai mici. Producţia totală pentru un ciclu de producţie depinde şi de activitatea fiziologică a plantei, de rapiditatea cu care fructele ajung la maturitatea de recoltare. Acesta aspect este foarte important la ardeiul gras, la care, apariţia timpurie a fructelor şi o recoltare eşalonată este un obiectiv principal. La ardeiul gogoşar şi cel lung, care se utilizează cu frecvenţă mai mare pentru conservare, maturizarea tehnică poate fi mai târziu, dar este necesară o maturizare uniformă. Aceleaşi obiective sunt de bază şi în cazul ardeiului pentru boia.
Realizarea producţiei pe plantă depinde de numărul fructelor şi de mărimea acestora. Mărimea fructelor este specifică fiecărei varietăţi, dar poate fi variabilă în cadrul aceleiaşi varietăţi, aşa cum este cazul ardeiului iute.
La ardeiul gras, fructele pot avea greutatea de la 50 g până la peste 150 g. (IFRIM AURELIA, 2003). În funcţie de modul de utilizare se preferă fructe mai mari sau mai mici. Pentru conservare sunt preferate fructele mai mici, iar pentru consumul proaspăt fructele mai mari. Numărul fructelor pe plantă, în corelaţie cu mărimea lor, trebuie să depăşească valoarea de 10 fructe. (GLĂMAN şi colab., 2002)
La ardeiul gogoşar greutatea medie fructelor de pe o plantă trebuie să fie mai mare, să depăşească 100 g, să tindă spre 200 g. Cu cât fructele sunt mai mari cu atât sunt mai apreciate. Pentru a avea o bună rentabilitate la prelucrarea fructelor în procesul conservării, partea edibilă (pulpa fructului) trebuie să reprezinte peste 80% din greutatea fructului. (IFRIM AURELIA şi BĂLAŞA M., 2002)
La arediul lung foarte important este numărul fructelor pe plantă şi greutatea acestora. Numărul fructelor pe o plantă este mai redus decât la ardeiul gras sau iute pentru că recoltarea se face numai aproape de maturitate. Greutatea medie a unui fruct trebuie să depăşească 100 g, iar numărul lor să fie aproape de 10.
O categorie aparte de ardei lung este ardeiul pentru boia la care numărul fructelor pe plantă este elementul hotărâtor al capacităţii de producţie. Pe o plantă pot fi realizate peste 15 fructe cu greutatea medie a unui fruct în jur de 100 g. (MADOŞĂ şi colab., 2003)
La ardeiul iute numărul fructelor pe plnată este foarte mare, putând depăşi valoarea de 30 fructe pe plantă. Despre greutatea medie a unui fruct nu se pot face referiri precise deoarece variabilitatea este foarte pronunţată. Există forme cu fructe foarte mici (sub 10 g) sau forme cu fructe mari (peste 50 g). În privinţa ameliorării nu există direcţii foarte precise fiind necesară o gamă largă de soiuri.
Ameliorarea calităţii. Calitatea este un obiectiv la fel de important ca şi capacitatea de producţie. Calitatea cuprinde două aspecte principale: cel biochimic şi cel morfologic.
Din punct de vedere biochimic, fructele trebuie să fie bogate în vitamine, la ardei proporţia cea mai mare prezentând-o vitaminele A şi C. Vitaminele, substanţele dulci şi capsicina realizează gustul şi aroma fructelor. Cele mai preferate sunt fructele dulci, însă în anumite zone ale lumii, în special în Asia, sunt la fel de preferate şi cele iuţi. Varietăţile de ardei gras, gogoşar şi parţial cel lung, trebuie să aibă fructe dulci.
Fructe iuţi apar la varietăţile de ardei lung, şi bineînţeles la ardeiul iute. Există chiar şi forme de ardei gogoşar cu fructe iuţi. Acestea se pot recunoaşte uşor deoarece au fructele imature de culoare galbenă.
Ardeiul pentru boia poate avea fructe din ambele categorii. Pentru consumul curent se preferă boiaua dulce, dar pentru prepararea mezelurilor este preferată boiaua mai picantă. O componentă importantă a calităţii ardeiului pentru boia este conţinutul în substanţă uscată a fructului şi cel în pigmenţi. Formele care prezintă fructe cu un procent mai mare de substanţă uscată dau producţii mai mici, din acest motiv, la ardeiul pentru boia selecţia trebie făcută după cantitatea de boia obţinută la unitatea de suprafaţă. Pigmentaţia fructelor mature se determină mai dificil, dar în selecţie se poate lucra notându-se prin note produsul finit, după o scară de culori.
Aspectele morfologice implicate în realizarea calităţii sunt: forma, culoarea şi mărimea fructelor. Forma şi culoarea depind de preferinţele consumatorilor. O diversitate mai mare de forme şi culori
constituie un motiv de atracţie pentru consumatori. În privinţa mărimii sunt preferate fructele mai mari, cu pulpă groasă. Grosimea pulpei este foarte importantă la ardeiul gras şi la gogoşar. La ardeiul gogoşar, valorile acestui caracter trebuie să fie de peste 7 mm. Referitor la mărimea fructelor se poate spune că pentru a realiza producţii mari şi fructele trebuie să fie mari. Totuşi, sunt şi cazuri în care se preferă fructe mai mici la ardeiul iute sau chiar la ardeiul gras (când prepararea se face ca ardei umplut). Acestea au o frecvenţă destul de mare în recolta realizată prin cultivarea soiurilor. În cazul hibrizilor fructele sunt mai mari şi foarte uniforme.
Ameliorarea rezistenţei la boli. Arealul de cultură foarte mare al ardeiului, face ca la această cultură să existe un număr mare de boli. În climatul subtropical bolile sunt mai frecvente în sezonul ploios. În aceste zone, cele mai frecvente boli sunt Xanthomomas campestris pv.vesicatoria, Fusarium oxysporum, cercospora capsici, Colletotrichum capsici.
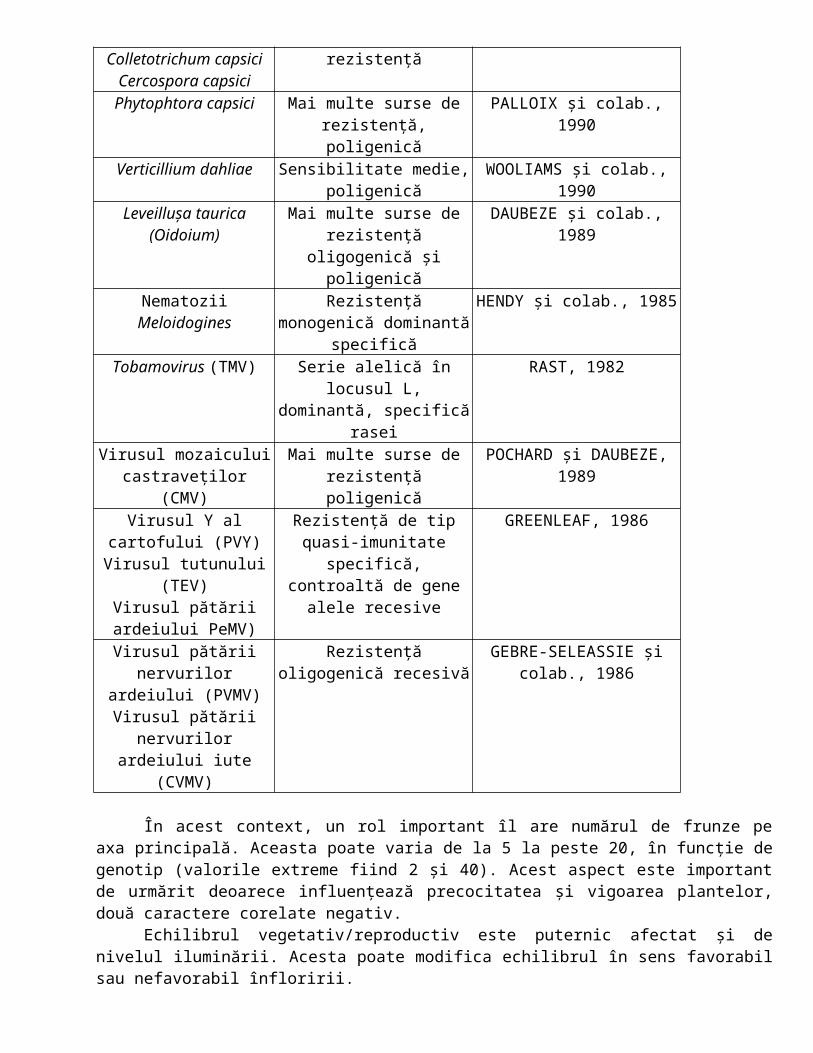
O altă gamă de agenţi patogeni au o arie de răspândire mult mai largă. Phytophtora capsici, Verticillum dahlie şi numeroşi viruşi. Principalii agenţi patogeni sunt prezentaţi în tabelul 3.4.
Ameliorarea adaptabilităţii. Până în prezent au fost puţine eforturi consacrate studiului proprietăţilor adaptative şi unei cunoaşteri mai bune a creşterii şi dezvoltării ardeiului. Variabilitatea morfologică şi fiziologică a formelor cultivate este foarte importantă. De altfel, elaborarea unor tehnologii de cultură în condiţii protejate pe parcursul diferitelor sezoane s-a dovedit a fi dificilă, fructificarea ardeiului fiind dependentă de condiţiile de mediu şi recunoscută ca foarte capricioasă. Astfel de studii de bază au fost efectuate în Olanda, Australia, Israel şi mai recent în Franţa, fiecare subliniind particularităţi aparte.
Este clar că echilibrul între creşterea vegetativă şi aptitudinile reproductive (calitatea florilor şi fructelor) joacă un rol major. Cultivatorul trebuie să menţină acest echilibru creat de ameliorator care poate fi uşor afectat în condiţiile unei culturi intensive. Dezechilibrul poate rezulta dintr-un deficit de fructificare debutat încă de la înflorit (fecundare defectuoasă) sau din contră, a unui exces care poate bloca dezvoltarea ulterioară a fructelor, care în final va duce la limitarea producţiei.
Tabelul 3.4.Principalele boli prezente la ardei şi sursele de rezistenţă utilizate (după POCHARD şi colab.,
1992)Agentul patogen Rezistenţa şi determinismul Referinţa biobliografică
În acest context, un rol important îl are numărul de frunze pe axa principală. Aceasta poate varia de la 5 la peste 20, în funcţie de genotip (valorile extreme fiind 2 şi 40). Acest aspect este important de urmărit deoarece influenţează precocitatea şi vigoarea plantelor, două caractere corelate negativ.
Echilibrul vegetativ/reproductiv este puternic afectat şi de nivelul iluminării. Acesta poate modifica echilibrul în sens favorabil sau nefavorabil înfloririi.
De exemplu, soiurile tropicale în zonele de origine realizează 5 frunze pe axa principală. Soiurile selecţionate pentru rezistenţă la temperaturi scăzute, când sunt cultivate în extrasezon vor avea o frunză mai puţin.
Condiţiile de cultură afectează şi cantitatea şi calitatea polenului, astfel că unele flori pot rămâne nefecundate. O sensibilitate mai redusă a polenului faţă de condiţiile de mediu poate fi indusă prin introgresia de gene favorabile de la Capsicum baccatum.
Prin urmărirea sistemului foliar şi a fertilităţii polenului se pot rezolva problemele legate de iluminarea insuficientă în spaţiile protejate, factor care poate fi limitativ pentru productivitate.
Ceilalţi factori (apă, substanţe nutritive, temperatură) trebuie asiguraţi la nivelul cerinţelor prin tehnologie. Valorile naturale a acestor factori, în condiţiile ţării noastre nu pot asigura realizarea unor producţii satisfăcătoare. Pentru toate zonele se impune irigarea şi asigurarea fertilizării optime. Nu se cunosc forme rezistente la secetă şi frig.
Ameliorarea poate contribui la crearea unor forme cu rezistenţă la sărurile din sol, germoplasma prezentând astfel de genotipuri, cu precădere specii sălbatice înrudite.
AMELIORAREA PĂTLĂGELELOR VINETE
Pătlăgelele vinete sunt cultivate pentru fructele lor cu calităţi gustative deosebite cât şi pentru pretabilitate la prelucrarea industrială. Gama de preparate culinare ce se poate obţine din pătlăgelele vinete este foarte largă, putând fi consumate singure sau asociate cu alte legume. Prepararea fructelor acestei legume se poate face prin prăjire şi fierbere sau pot fi conservate prin diferite metode (DAUNAY şi colab., 1995)
Deşi valoarea nutritivă a fructelor de pătlăgele vinete este mai scăzută comparativ cu alte specii de legume, ele sunt apreciate datorită valorii lor gustative şi a unor componente chimice, cum a r fi: unii compuşi antocianici ca de exemplu nasunina ( aflată mai ales în pericarpul fructelor), caroten, vitamina C, vitamina PP, acid aspartic, acid malic, substanţe pectice şi altele. Conţinutul în săruri minerale în fructe este destul de mare, cele mai importante elemente fiind: calciu, fosfor, fier, potasiu. (SUTEKI, 1989)
În stare proaspătă fructele au un gust amar dat de prezenţa unui glicoalcaloid, stereoidic-solanină, însă prin preparare aceasta substanţă este descompusă şi gustul amar dispare. Fructele de pătlăgele vine au un conţinut de substanţă uscată de 8-9 %, 3-4 % zahăr, 1,1 % pectine şi o valoare nutritivă de 18 kcal/kg.
Dintre binefacerile de ordin medical a fructelor de pătlăgele vinete trebuie menţionat faptul că prin consumarea acestor fructe scade procentul de colesterol din sânge la animale şi la om şi creşte diureza. (CHEN şi LI, 1995)
Obiective de ameliorare.
Ameliorarea capacităţii de producţie. Analizele au arătat că producţia este determinată în primul rând de greutatea fructului şi în al doilea rând de numărul de plantă (ANASO şi OKEREKE, 1990).
Prin utilizarea celor mai adecvate metode de ameliorare, inclusiv al folosirii speciilor înrudite, producţia de fructe pe plantă la pătlăgele vinete a atins 4,4 kg. Producţia pe plantă este dată de numărul
fructelor şi mărimea acestora. Numărul fructelor pe plantă poate fi cuprins în medie între 6 şi 80, cu o medie de 24. Majoritatea soiurilor prezintă sub 20 fructe pe plantă. Greutatea medie a fructelor este cuprinsă între 15 g şi 1,5 kg. (SWARUP, 1995). Lungimea fructelor variază de la 2,9 cm la 30,1 cm, iar diametrul între 2,5 cm şi 10,4 cm după CHEN şi LI (1995) sau 4-45 cm lumgime şi 2-356 cm diametru după SWARUP (1995)
Realizarea unor forme cu potenţial foarte ridicat de producţie impune modificarea substanţială a particularităţilor morfologice, biochimice, ecologice şi citologice ale plantelor. Desimea la unitatea de suprafaţă este dependentă de habitusul plantei. Înălţimea plantei poate fi cuprinsă între 70 şi 160 cm, media fiind, la soiurile cultivate de 100-120 cm. Numărul de ramificaţii pe plantă este de 5 până la 11, majoritatea soiurilor având 6-8 ramificaţii. Între numărul de ramificaţii şi producţia de fructe pe plantă există o corelaţie strânsă. (VADIVEL şi BAPU, 1990)
Viitoarele cultivare vor trebui să posede, printre altele, următoarele însuşiri: capacitatea mare de absorbţie a substanţelor minerale, în special a sărurilor de azot din sol, însuşire care necesită dezvoltarea substanţială a sistemului radicular, productivitatea ridicată a fotosintezei, dezvoltarea corespunzătoare a elementelor producţiei, rezistenţă la boli şi insecte.
Aceasta presupune că există în stocul de gene al speciilor cultivate, sau a speciilor înrudite, a genelor care să determine însuşirile respective şi de posibilitatea transferului genelor valoroase străine în noile forme ameliorate.
Majoritatea speciilor Solanum înrudite cu vânăta posedă un sistem radicular puternic dezvoltat şi au capacitatea de fructificare în ciorchine, realizând un număr foarte mare de fructe pe plantă. Speciile Solanum torvum, Solanum macrocarpon, Solanum integrifolium şi Solanum nodiflorum posedă însuşire de fructificare în ciorchine.
În general, este destul de dificil de reunit într-un soi capacitatea de producţia ridicată cu factorii ce conferă rezistenţă la condiţiile de mediu nefavorabil. Pentru sporirea rezistenţei la factorii adverşi de mediu de cele mai multe ori se apelează la speciile înrudite sălbatice, sau semisălbatice, care posedă asemenea însuşiri, dar acestea au un potenţial productiv redus
O cale sigură şi mult mai uşoară de sporirea producţiei este utilizarea efectului heterozis. Capacitatea de producţie a hibrizilor F1 de pătlăgele vinete este mai mare cu 24-43% faţă de părinţi, iar hibrizii F1de pătlăgele vinete au depăşit cu 51-76% producţia soiului martor.
CHEN N.C. şi LI H.M (1995) că hibrizii F1 s-au situat pe primele locuri la producţia totală comparativ cu formele homozigote.
Ameliorarea calităţii. Pentru stabilirea obiectivelor de ameliorare în domeniul calităţii recoltei trebuie pornit de la modul de prepare al fructelor şi de la preferinţele de consum. Fructele de pătlăgele vinete pot fi consumate ca atare, sau în urma prelucrării. Ele pot fi preparate prin coacere, fierbere, prăjire în ulei, sau ca murături, singure sau în amestec cu alte legume.
În funcţie de modul de preparare al fructelor, precum şi preferinţele de consum ce pot diferi de la o zonă geografică la alta, putem stabili obiective cu caracter general cum ar fi: pieliţa subţire a fructelor, pulpa de culoare alb-gălbuie cu textură fină, elastică, fără gust amar şi conţinut redus de seminţe. Cerinţele în privinţa dimensiunilor şi formei fructelor diferă foarte mult în funcţie de modul de preparare al lor.
Dacă la prepararea prin fierbere sub formă înăbuşită se preferă fructele mari, la conservarea sub formă de murături se preferă fructele de dimensiuni mici.
Un obiectiv important în procesul de ameliorare al pătlăgelelor vinete este obţinerea de soiuri, sau hibrizi cu conţinut ridicat în substanţa utile. Pentru păstrarea culorii albe a pulpei fructelor în timpul prelucrării se urmăreşte obţinerea de varietăţi cu conţinut redus în compuşi fenolici şi glicoalcaloizi, iar activitatea peroxidazelor să fie la un nivel scăzut. Aceste însuşiri sunt întrunite la un nivel optim la varietăţile cu fructe de culoare albă (PEARSON, 1983).
Pentru păstrarea sub formă congelată se preferă soiurile cu fructe alungite şi cu o bună capacitate de păstrare. Un rol important îl are şi conţinutul de substanţă uscată. Substanţa uscată în fructe este cuprinsă între 6,2 şi 21,4%. Varietăţile cu fructe mici au un conţinut de substanţă uscată în fruct mai mare comparativ cu cele care prezintă fructe mai mari.
La noi în ţară se preferă în general fructele de dimensiuni mijlocii, sau mari, de formă ovoid
alungită, sau piriform alungită, intens colorate, cu luciu persistent, lipsite de gust amar şi conţinut redus de seminţe.
Ameliorarea timpurietăţii. Timpurietatea reprezintă un obiectiv major în procesul de ameliorare la pătlăgele vinete. Înfloritul la această specie se desfăşoară paralel cu creşterea vegetativă atâta timp cât condiţiile de temperatură şi hrană sunt adecvate (SUTEKI, 1989).
Prin obţinerea de soiuri timpurii se măreşte perioada de fructificare şi de prelucrare a fructelor. În general soiurile timpurii având o perioadă mai mare, au o capacitate de producţie mai mare comparativ cu soiurile tardive (SWARUP, 1995).
La soiurile timpurii, la cultura în seră, prin scuturarea perioadei vegetative se reduc cheltuieli cu agentul termic, lucru deosebit de util pentru reducerea costurilor de producţie.
Pe lângă formarea mai devreme a florilor, soiurile timpurii trebuie să posede însuşirea de legare a fructelor în condiţiile termice mai scăzute din această perioadă. La multe soiuri primele flori avortează din cauza eşuării proceselor de polenizare, polenul fiind incapabil să germineze pe stigmat la temperaturi mai scăzute. Unele specii sălbatice de Solanum, manifestă exigenţe termice reduse, ele fiind capabile să reziste la îngheţurile uşoare din primăvară şi sunt apte să lege fructe la temperaturi joase. ele pot fi folosite în procesul de ameliorare a pătlăgelelor vinete pentru transferul acestor însuşiri.
Timpurietatea apreciată după numărul de zile de la transplantat până la prima recoltare poate fi cuprinsă între 70 şi 130 zile, media fiind de 84 zile. La soiurile timpurii prima floare se formează la nodul 5-6, iar la cele la nodul 8-9.Plantele la soiurile timpurii au o talie mai joasă, iar frunzele au dimensiuni mai reduse.
Soiurile de pătlăgele vinete din var. Orientale posedă însuşirea de fructificare timpurie şi pot fi folosite în lucrările de ameliorare pentru transferul acestei însuşiri.
Ameliorarera rezistenţei la boli şi dăunători. Pătlăgelele vinete sunt afectate de numeroase boli şi de atacul unor dăunători ce pot afecta din punct de vedere cantitativ şi calitativ recolta, sau pot compromite în totalitate cultura. Printre cele mai păgubitoare boli ce pot afecta cultura în zonele temperate enumerăm verticiloza, produsă de ciuperca Verticillium dahliae Kleb., ofilirea (fuzarioza) vinetelor produsă de ciuperca Fusarium oxysporum f.sp.melongena şi putregaiul fructelor produs de ciuperca Phytophthora parasitica (MARINESCU, 1986).
Dăunătorii care afectează cel mai frecvent cultura de pătlăgele vinete sunt gândacul din Colorado (Leptinotarsa decemlineata), păianjenul roşu (Tetranychus urticae) şi nematozii de rădăcină (Meloidogyne spp.).
Pentru combaterea acestor agenţi patogeni se efectuează un număr mare de tratamente chimice. Mijloacele de combatere chimică pot contribui la diminuarea atacului agenţilor patogeni, dar ele sunt foarte costisitoare şi destul de greu de aplicat, mai ales în perioada de fructificare când repausul după tratament depăşeşte intervalele dintre recoltării (HORGOŞ, 2003).
În plus, produsele chimice folosite la tratament pot afecta prădătorii şi paraziţii dăunătorilor, cu consecinţe negative de lungă durată.
Obţinerea de soiuri de pătlăgele vinete cu rezistenţă la atacul principalilor agenţi patogeni ar elimina aceste neajunsuri legate de utilizarea mijloacelor chimice de combatere. CLAUDE CH. şi CLAUDE F.; (1991) consideră ca un obiectiv prioritar introducerea rezistenţei la boli la pătlăgelele vinete şi apoi îmbunătăţirea însuşirilor morfologice, productive şi calitate a fructelor.
Studiile întreprinse până în prezent au arătat că nici un soi de pătlăgele vinete nu posedă un nivel corespunzător de rezistenţă la atacul agenţilor patogeni enumeraţi mai sus.
Speciile de Solanum mai apropiate, sau mai îndepărtate de pătlăgele vinete au fost considerate ca posibile surse de rezistenţă la boli. Încercările de a transforma genele de rezistenţă de la speciile sălbatice la formele cultivate au eşuat din cauza incompatibilităţi la hibridare a speciilor, sau a sterilităţii ridicate a hibrizilor obţinuţi. Din această cauză s-a recurs la folosirea speciilor sălbatice, sau a hibrizilor interspecifici ca portaltoi de rezistenţă pentru combaterea bolilor transmise prin sol. Ca portaltoi de rezistenţă în Japonia se folosesc speciile Solanum integrifolium şi Solanum torvum, precum şi hibridul F1
Solanum integrifolium x Solanum melongena. Înmulţirea speciilor sălbatice precum şi producerea seminţei hibride este dificilă şi foarte costisitoare, din această cauză metoda nu poate fi extinsă la culturile de câmp.
Prezenţă perişorilor gladulari pe tulpină şi frunzele plantelor pot diminua atacul produs de păianjenul roşu.
Ameliorarea altor caractere utile. Concentrarea producţiei la această specie cu fructificare îndelungată poate contrbui la reducerea cheltuielilor cu lucrările de întreţinere şi de recoltare. Inflorescenţa polifoloră şi fructificarea în ciorchine pot conduce la concentrarea producţiei la pătlăgele vinete şi la reducerea cheltuielilor.
Vânăta face parte din grupul speciilor cu spini ai genului Solanum. Există totuşi varietăţi lipsite de spini pe tulpină, sau frunzele plantei.
Lipsa spinilor pe plantă şi caliciul fructelor pot uşura lucrările de întreţinere şi mai ales de recoltare şi sortare a fructelor. Portul erect şi tufa strânsă a plantelor permit sporirea densităţii culturii cu efect benefic asupra producţiilor la unitatea de suprafaţă şi reducerea costurilor de producţie.
AMELIORAREA VERZEI
Varza face parte din familia Cruciferae care cuprinde un mare număr de specii cultivate. Scopurile pentru care sunt cultivate acestea sunt diverse. Unele sunt cultivate pentru alimentaţia umană cum sunt diferitele varietăţi de varză, ridiche sau cresonul, altele se cultivă ca plante furajere, varza şi rapiţa furajeră, gulia furajeră. Există şi specii utilizate pentru extragerea uleiurilor (rapiţa pentru ulei şi muştarul) sau utilizate ca şi condimente (muştarul şi hreanul). Aceste moduri variate de utilizare demonstrează larga variabilitate din cadrul familiei, dar mai cu seamă din genul Brassica, deoarece majoritatea speciilor aparţin acestui gen.
Cultivarea verzei este atestată din vremuri străvechi, încă din antichitate fiind menţionate mai multe varietăţi. Anticii egipteni şi eleni cunoşteau două forme de varză pentru căpăţână, cu frunza netedă şi cu frunza grofată.
În decusrul timpului interesul pentru varietăţile de varză a evoluat. Până nu de mult, atât la noi în ţară, dar şi pe plan mondial se cultiva cu precădere varza pentru căpăţână. Din a diua jumătate a secolului XX s-au mărit şi suprafeţele cultivate cu conopidă şi gulie. Celelalte varietăţi ocupă şi în prezent un loc mai puţin important în cultură. În vestul Europei se constată o scădere a interesului pentru varza pentru căpăţână în favoarea celorlalte varietăţi datorită unui curent de diversificare a hranei care este prezent în ţările mai dezvoltate. Pentru estul Europei şi Asia (în special China şi India) varza pentru căpăţână rămâne principala varietate cultivată.
Varza pentru căpăţână este urmată de conopidă. Extinderea acestei varietăţi a avut loc în vestul şi sudul Europei. La noi în ţară se constată un interes tot mai mare pentru aceasta, în special în vestul ţării. Tot în această parte de ţară sunt mai cunoscute şi gulia şi varza creaţă. În ultimii ani, în sudul Europei se cultivă pe suprafeţe foarte mari brocoli, varietate care a pătruns şi în Statele Unite ale Americii. În România această varietate, alături de varza chinezească sunt cunoscute numai după anul 1990. În trecut, au fost cultivate suprafeţe importante şi cu varză furajeră, dar interesul pentru aceasta a scăzut deoarece are o valoare furajeră redusă şi se conservă foarte dificil.
Răspândirea geografică a legumelor din grupa verzei s-a realizat datorită marii plasticităţi a acestei specii, din regiunile calde mediteraneene sau chiar subtropicale, până în regiunile nordice cu climă rece. În ţara npastră varza se cultivă din zona de câmpie până în zona montană.
Obiective de ameliorare.
În ameliorarea verzei există două gupe de obiective. Unele obiective sunt generale tuturor varietăţilor, altele însă sunt specifice fiecărei varietăţi în parte.
Ameliorarea capacităţii de producţie este obiectivul principal în orice program de ameliorare. La varza pentru căpăţână potenţialul productiv este foarte mare, însă diferă în funcţie de precocitate soiului. Producţia la unitatea de suprafaţă depinde de desimea plantelor şi de potenţialul fiecărei plante. La soiurile timpurii desimea poate fi mai mare pentru plantele ajung la maturitatea tehnică mai repede şi căpăţânile sunt mai mici. La varza de toamnă căpăţânile pot depăşi greutatea de 5 kg, ca urmare habitusul plantei va ocupa mai mult spaţiu. În această situaţie foarte importantă este mărimea rozetei de frunze şi
poziţionarea frunzelor. Rozeta cuprinde frunzele care nu participă la formarea căpăţânii. Ele ajută planta să crească deoarece sunt principalele frunze care efectuează fotosinteza de aceia numărul lor nu trebuie obligatoriu redus. Aceste frunze trebuie să prezinte un peţiol mai scurt şi să fie poziţionate cât mai spre verticală.
În funcţie de greutatea căpăţânii, care poate fi cuprinsă între limitele de 0,5 şi 5 kg. Foarte iumportantă este uniformitatea căpăţânilor, care la hibrizi este mult mai bună decât în cazul soiurilor. Căpăţânile mici nu trebuie să reprezinte mai mult de 5-7%. (GHICĂ, 2001) Producţiile realizate la unitatea de suprafaţă variază între 20 şi 60 t/ha.
Preferinţele consumatorilor faţă de mărimea şi greutatea căpăţânii sunt diferite. În general, căpăţânile foarte mari nu sunt preferate decât în bucătăriile unde se pregătesc cantităţi mari de hrană. Greutatea optimă a unei căpăţâni este în jur de 2 kg. Asemenea greutăţi se pot obţine uşor la varza de toamnă, dar la soiurile timpurii valorile sunt ceva mai reduse.
Producţia plantei este dependentă de dimensiunile căpăţânii şi de densitatea acesteia. Sunt preferate căpăţânile dense. Dimensiunile căpăţânii se determină la maturitate. Variabilitatea diametrului căpăţânii poate varia de la 10 cm la peste 40 cm, dar valorile cele mai frecvente sunt în jur de 25-30 cm Foarte important este stabilirea momentului optim al recoltării, care se face după realizarea unui studiu a dinamicii creşterii căpăţânii. Pe baza unor măsurători în câmp se determină volumul căpăţânii. Sunt luate în considerare înălţimea şi diametrul căpăţânii. La căpăţânile cu formă rotundă în secţiune transversală volumul se calculează cu formula:
V = I x D2 x 1/6 π,Pentru căpăţânile cu secţiune transversală neregulată, volumul se calculează cu formula:
V = I x Dmin. x Dmax. x 1/6 π I – înălţimea căpăţânii, D – diametrul căpăţânii, D.min. – diametrul minim al căpăţânii, D.max –
diametrul maxim al căpăţânii.Rezultatele obţinute din aceste calcule se corectează cu coeficienţi în funcţie de forma căpăţânii,
0,95 pentru căpăţânile rotunde, turtite sau puţin alungite şi 0,90 pentru căpăţânile ascuţite. Densitatea influenţează direct greutatea căpăţânii. Pentru determinarea acestui caracter există
aparate speciale cu care determinarea se poate efectua înainte de recoltare. După recoltare selecţia se poate face prin introducere în apă comparativ cu un soi martor a cărei densitate este bine determinată.
În procesul de ameliorare trebuie avute în vedere corelaţiile care există între caractere. Este cunoscut faptul că la varză, mărimea căpăţânii este corelată pozitiv cu tardivitatea. Este foarte dificil să se creeze soiuri precoce cu căpăţâni mari. La varza roşie mărimea căpăţânii este mult mai mică decât la varza albă, deci între culoarea antocianică a frunzelor şi mărimea redusă a căpăţânii este tot o corelaţie pozitivă.
La celelalte varietăţi productivitatea este determinată, aproximativ de aceleaşi caractere morfologice.
La varza creaţă elementele productivităţii sunt identice cu cele de la varza pentru căpăţână. Totuşi, căpăţânile sunt mai mici, iar densitatea este mult inferioară
La conopidă inflorescenţa trebuie să fie de peste 2 kg, iar frunzele din rozetă adunate. Poziţionarea frunzelor cât mai aproape de verticală are două avantaje. Pe de o parte permite desimi de plantare mai mari, iar pe de altă parte, asigură şi o protejare a inflorescenţa de razele solare şi ploi, ceşti factorii ducând la deprecierea ei (îngălbenire, atac de boli)
La gulie tuberofructul are mărimi şi greutăţi diferite, în funcţie de tipul cultivarului. Soiurile timpurii şi destinate cultivării în spaţii protejate prezintă teboerofructele mai mici, sub 500 g, pe când la soiurile de toamnă-iarnă, greutatea acestora poate depăşi 1 kg.
La varza de Bruxelles productivitatea este dată de numărul căpăţânilor pe plantă, de mărimea acestora şi de densitatea lor. Numărul căpăţânilor este dependent de lungimea tulpinii şi densitatea inserţiei frunzelor.
La brocoli, productivitatea are aceleaşi elemente componente ca şi la conopidă.Ameliorarea calităţii. Calitatea producţiei este dată de elemente morfologice şi biochimice. Din punct de vedere morfologic calitatea căpăţânii este dependentă de forma sa, fineţea frunzelor
interioare şi de mărimea cepului. Forma căpăţânii este variabilă, de la foarte turtită la foarte ascuţită. Pe
piaţa românească sunt preferate căpăţânile rotunde, uşor turtite sau alungite, dense, cu frunze subţiri şi puţin gofrate. La varza de toamnă căpăţânile turtite sunt preferate deoarece la conservarea prin murare ocupă mai puţin spaţiu. Formele plante sunt mai frecvente la varza de toamnă, pe când la varza timpurie căpăţânile sunt rotunde sau alungite, chiar ascuţite la vârf. Forma căpăţânii este un caracter destul de stabil cu toate că raportul între cele două dimensiuni (înălţime şi lăţime) poate varia de la un an la altul, în funcţie de condiţiile de climă. S-a constatat că formele sferice şi uşor turtite au şi o bună densitate, deci acestea ar fi formele care ar trebui reţinute în procesul de selecţie.
Cepul este porţiunea de tulpină pe care sunt inserate frunzele, fiind puternic lignificată este improprie consumului. Din acest motiv, mărimea cepului trebuie să fie cât mai redusă. Lungimea cepului poate reprezenta de la 25 până la 90% din înălţimea căpăţânii. Scurtarea cepului foarte mult nu este posibilă deoarece s-ar reduce forte mult numărul frunzelor din căpăţână. Prin ameliorare se consideră că este satisfăcătoare o lungime a cepului la nivelul a 50-60% din înălţimea căpăţânii. Pentru a se reduce lungimea cepului în condiţiile în care numărul frunzelor să rămână mare, grosimea acestora trebuie să fie redusă. Frunzele fine constituie un element de calitate în plus.
Densitatea căpăţânii este atât un element al calităţii, cât şi al productivităţii. Selecţionând forme cu cepul scurt se îmbunătăţeşte şi densitatea căpăţânii. Densitatea căpăţânii este un caracter dependent de tipul soiului, dar şi de condiţiile de climă. Soiurile timpurii prezintă căpăţânile mai puţin dense comparativ cu cele tardive. În condiţii de stres hidric, căpăţânile rămân mici şi cu densitate foarte scăzută. În ultimul timp, prin ameliorare au fost obţinuţi hibrizi timpurii la care densitatea căpăţânii este foarte bună, dar frunzele din căpăţână sunt groase şi puternic încreţite.
Gustul şi mirosul degajat la fierbere sunt alte componente ale calităţii. La realizarea acestor două componente contribuie conţinutul de grăsimi din frunză şi compuşii cu sulf. Determinismul acestora nu este cunoscut, iar selecţia se face numai prin observare directă.
Componentele morfologice implicate în realizarea calităţii sunt diferite în funcţie de varietate.La conopidă inflorescenţa trebuie să prezinte o formă regulată, de culoare albă, fără frunze
interioare cu primordiile florale la suprafaţa inflorescenţei şi cu dezvoltarea ramificaţiilor florifere cât mai tardivă.
La varza de Bruxelles căpăţânile trebuie să fie dense şi uniforme, cu aptitudini pentru recoltare mecanizată.
La gulie tuberofructul trebuie să fie de formă regulată, cu număr redus de vase lemnoase şi o structură fină a acestora.
La brocoli inflorecenţa trebuie să fie densă, colorată uniform şi fără frunze interioare, cu o bună rezistenţă la boli criptogamice şi bacterioze
Din punct de vedere biochimic, calitatea este influenţată de conţinutul de substanţă uscată, proteine, grăsimi, compuşi liberi de azot, celuloză şi unele vitamine.
Căpăţâna de varză este foarte bogată în apă, atât la varza albă, cea roşie cât şi la cea creaţă. Apa, poate reprezenta între 92 şi 94 %, restul fiind substanţă uscată. Varza de Bruxelles prezintă un conţinut mai ridicat de substanţă uscată, putând fi cuprins între 13 şi 16%. La varza pentru căpăţână limitele de variabilitate sunt mult mai largi, de la 5 la 13,5%.
Cantităţile cele mai mari de componente nutritive se întâlnesc la varza de Bruxelles: compuşi liberi de azot 6,22%, grăsimi 4,46%, proteină brută 4,46%, celuloză 1,57%. Tot această varietate prezintă şi cantităţile mai mari de provitamina A, vitamina C şi vitaminele B1 şi B2. (BECKER, 1950) Varza de Bruxelles prezintă şi cele mai mari procente de proteină digestibilă (aproape 5%) şi acizi graşi (peste 8%) (ALEXE CONSTANŢA şi MĂRIUŢEI ALEXANDRINA, 2001)
Conopida şi brocoli se aseamănă cu varza de Bruxelles din punct de vedere biochimic, iar gulia este mai bogată în substanţă uscată.
Pe lângă analizele precise de determinare a principiilor nutritive, se urmăreşte şi prezenţa unor substanţe toxice remanente din tehnologia aplicată ( pesticide, nitraţi) care se pot acumula în organele vegetative. În frunze acestea se pot acumula în cantităţi mai mari. Având în vedere că varza pentru căpăţână se consumă şi nefiartă, toxicitatea acestor substanţe este mult mai mare.
Ameliorarea rezistenţei la crăpare a căpăţânii. Rezistenţa la crăpare este obiectiv specific verzei pentru căpăţână. Acest caracter este influenţat de condiţiile de climă şi este foarte frecvent când după
perioade foarte secetoase survin ploi abundente. Totuşi, există şi o influenţă a genotipului, deoarece apar diferenţe de la un genotip la altul. Aceste diferenţe se datorează ritmului de creştere a frunzelor şi rezistenţei la secetă. Productivitatea este mai puţin influenţată de crăpare, dar calitatea este mult depreciată. Determinarea acestei însuşiri se face după ce căpăţânile au ajuns la maturitate, prin numărarea zilelor de la maturizarea şi până la crăparea a 50% din căpăţâni. Acest mod de determinare se aplică foarte rar, fenomenul observându-se după perioadele secetoase urmate de ploi. În condiţiile aplicării unei tehnologii corespunzătoare, cu asigurarea necesarului de apă prin irigare, fenomenul este puţin prezent.
S-a constatat că la soiurile timpurii există o corelaţie negativă între precocitate şi rezistenţa la crăpare, dar fenomenul este semnalat şi la cele de toamnă.
Ameliorarea rezistenţei la păstrare în timpul iernii este un obiectiv specific soiurilor târzii. La varza pentru căpăţână se pune problema păstrării frăgezimii, dar şi a compoziţiei biochimice. Frăgezimea scade datorită pierderii apei. Cel mai bine se păstrează varza roşie, care până în primăvară pierde din greutate 8-10%, urmând varza albă cu pierderi de 10-15% şi varza creaţă, la care pierderile sunt mai mari, de 10-20%. Biochimic, se semnalează pierderi mari de glucide şi mai reduse de celuloză şi substanţe minerale. Se înregistrează pierderi ş de vitamina C, dar aceasta se păstrează mai bine decât în fructe. (NEAGU, 1967)
Conopida şi brocoli nu se pot păstra o perioadă lungă de timp deoarece pierderile de apă sunt mult mai rapide. Gulia însă, este varietatea care se poate păstra pe o perioada destul de îndelungată, fără pierderi în greutate şi calitative foarte importante.
Ameliorarea rezistenţei la boli şi dăunători este un obiectiv comun tuturor varietăţilor. Toate varietăţile de varză sunt sensibile la atacul aceloraşi agenţi patogeni. Principalii agenţi patogeni care produc boli criptogamice sunt Plasmodiophora sp., Alternaria sp., Mycosphaerella sp., Botrytis sp., Pythium sp., Phoma sp.. Pe lângă aceştia se mai semnalează atacuri de viruşi (mozaicul) sau bacterioze (Xanthomonas sp.). Comuni sunt şi dăunătorii: albiliţa, purecii şi musca cruciferelor.
Ereditatea rezistenţei faţă de boli şi dăunători este destul de puţin cunoscută şi în cele mai multe cazuri este poligenică, greu de transferat de la o varietate la alta. Pentru unii agenţi patogeni se cunosc mai multe rase fiziologice. Aşa este cazul herniei (Plasmodiophora brassicae). Gene specifice acestor rase sunt cunoscute la varza furajeră şi la unele varietăţi sălbatice de la care au fost transferate la varza cultivată.
Pentru nervaţiunea neagră a frunzelor (Xanthomonas campestrtis) se cunosc gene de rezistenţă dominante preluate din surse asiatice şi transferate în soiuri mai vechi cultivate.
Pentru atacul dăunătorilor nu se cunosc gene specifice de rezistenţă.Ameliorarea rezistenţei la boli este o activitate destul de dificilă deoarece trebuie depistate în
permanenţă gene noi datorită evoluţiei agenţilor patogeni. Transferul acestora nu pune probleme deosebite, doar în cazul apelării la varietăţi şi specii sălbatice, deoarece de la acestea se înrăutăţesc aptitudinile productive şi calitative.
Ameliorarea rezistenţei la emiterea tijelor florale este un obiectiv important pentru toate varietăţile. La speciile bienale fructificarea este condiţionată de realizarea unor temperaturi scăzute. Acestea trebuie să acţioneze asupra platelor semincere plantate din toamnă. Ele pot acţiona şi asupra răsadului, dacă este plantat prea devreme şi în acest caz emiterea tijelor florale are loc în primul an de vegetaţie. Această situaţie poate apărea la soiurile timpurii, care se plantează foarte devreme. Selecţia se poate realiza prin înfiinţarea culturilor de seminceri din toamnă prin plantarea în septembrie pentru a începe formarea căpăţânii la sfârşitul toamnei. La intrarea în iarnă căpăţânile se secţionează şi se verifică pornirea în creştere a lăstarilor floriferi. Plantele la care aceşti lăstari au început să se dezvolte, se vor elimina.
Ameliorarea adaptării la mecanizare tehnologiei. În perioada de vegetaţie majoritatea lucrărilor se pot efectua mecanizat, dar lucrarea de recoltat trebuie efectuată încă manual la multe varietăţi. Recoltarea mecanizată este condiţionată de mai multe elemente, cum sunt: dimensiunile plantelor şi dimensiunile şi fragilitatea organelor recoltate.
În perioada actuală recoltarea mecanizată se practică la varza pentru căpăţână sau la gulie. A fost încercată şi la celelalte varietăţi, dar deoarece organele recoltate sunt foarte fragile, pentru a recolta fără
pierderi şi deprecieri cultura trebuie să fie foarte uniformă. Uniformitatea se impune în privinţa maturizării dar şi a dezvoltării organelor recoltate.
AMELIORAREA CEPEI
Ceapa ocupă un loc important în cadrul culturilor de legume, la nivel mondial cultura cepei situându-se pe locul patru. Interesul pentru această specie reiese şi din caracteristicile sale biologice care face mai dificil procesul de ameliorare: specie bulboasă, fotoperiodică, cu repaus marcat, bianuală şi alogamă.
Cultura cepei este extinsă aproape în întreaga lume. Cele mai mari suprafeţe se cultivă în Asia, aici fiind cultivate mai multe specii. În emisfera nordică, cultura cepei se extinde de la 100 la 650
latitudine.Schimburile comerciale cu producţia de ceapă sunt destul de reduse, din totalul producţiei, numai
10% face obiectul acestor sc himburi. Această situaţie se datorează condiţiilor de climă şi preţului de cost care sunt diferite de la o ţară la alta.. în ţările din sud costurile de producţie sunt mai reduse, potenţialul de producţie fiind mai mare şi precocitatea superioară. Cu toate acestea se simte un surplus de producţie în marile ţări cultivatoarepentru că majoritatea ţărilor îşi asigură necesarul de consum.
Sortimentul de soiuri este destul de vast, fiind dictat de posibilităţile diferite de nutilizare. La ceapă se pot distinge trei moduri de cultură şi mai multe variante thnologice. Fiecare dintre acestea impun anumite însuşiri, deci în procesul de ameliorare apar obiective diferite şi precise. Cele trei moduri de cultură sunt: cultură prin răsad, cultură prin arpagic şi cultira directă.
Cultura prin răsad este mai costisitoare doarece se obţine răsadul care se repică la 45-60 zile. Avantajul acestui tip de cultură este că se obţin culturi foarte uniforme.
Cultura prin arpagi este cea mai răspândită la noi în ţară şi se realizează în două etape. În primul an se seamănă la densităţi foarte mari pentru a se obţine bulbii (arpagicul) care se vor planta în câmp în primăvara anului viitor. Acest mod de cultură are unele dezavantaje. În primul an se pot produce pierderi la recoltare şi în păstrare, iar în al doilea an apare riscul înfloririi care atrage deprecierea calităţii şi a producţiei.
Cultura directă se înfiinţează prin semănatul al locul definitiv la sfârşitul iernii, în zonele mai calde se poate face chiar din toamnă. Semănatul din toamnă are unele avantaje, dar şi riscul influneţei gerurilor din timpul iernii şi înflorire prematură. Alt inconvenient este răsărirea deficitară şi neuniformă. (DUMITRESCU, 2001)
3.5.3. Obiective de ameliorare.
Condiţiile actuale de cultură impun o exigenţă foarte mare faţă de omogenitatea tuturor caracterelor urmărite în procesul de ameliorare. Ansamblul exigenţelor pieţei de bulbi proaspeţi sunt uniformizate la nivel mondial. Cele mai valoroase se dovedesc a fi soiurile olandeze din punct de vedere morfologic, dar acestea nu sunt adaptate la condiţiile de climă specifice altor zone, în special a celor din sud.
În plus, se constată o puternică concurenţă între amelioratori în privinţa satisfacerii cerinţelor cultivarelor. Extinderea ariei de cultură a unui soi şi satisfacera pretenţiilor cultivatorilor a făcu ca ameliorarea să se îndrepte spre crearea de hibrizi comerciali. În prezent, în vestul Europei cultura cepei este direcţionată spre cultivarea hibrizilor, acest lucru ducând la monopolizarea comerţului mondial cubulbi de către aceste ţării, în special de către Olanda. Pe lângă productivitatea foarte bună, hibrizii au şi avantajul că pot fi semănaţi direct.
Ameliorarea capacităţii de producţie. La ceapa pentru bulbi, care se seamănă primăvara, productivitatea este principalul obiectiv. Structura plantei şi fiziologia dezvoltării sale, impun amelioratorilor o analiză profundă a componentelor productivităţii, a acumulării substanţei uscate în bulbi, dar şi a factorilor implicaţi în conservare. (BREWSTER, 1977). Structura plantei în perioada de vegetaţie, în special tulpina redusă şi producţia utilă bazată pe îngroşarea frunzelor, limitează acţiunea ameliorării. De fapt coeficientul de eficacitate a lucrărilor de selecţie variază foarte puţin din cauza
acestei structuri. Odată cu creşterea productivităţii în materie proaspătă se poate reduce conţinutul de substanţă uscată, deoarece între capacitatea de producţie şi conţinutul în substanţă uscată există o strânsă corelaţie negativă. Din acest motiv, potenţialul productiv nu poate fi crescut exagerat pentru că reducând prea mult conţinutul bulbilor în substanţă uscată, scade capacitatea de păstrare.
O altă cale de sporire a potenţialului productiv este mărirea duratei perioadei de vegetaţie, între potenţialul productiv şi tardivitate fiind o corelaţie pozitivă. Nici tardivitatea nu poate fi exagerată, în funcţie de condiţiile specifice ale regiunii, există o anumită dată limită a recoltatului.
Cele două posibilităţi de creştere a capacităţii de producţie impun un examen foarte atent a acetor corelaţii şi o verofocare a posibilităţilor gnetice în concordanţă cu condiţiile de mediu. Dintre factorii de cultură, cel ami important este asigurarea deficitului hidric, care este foarte greu de depăşit în condiţiile semănatului în primăvară.
Nivelul producţiilor trebuie să fie cât mai mare posibil, să tindă spre valoarea de 80 t/ha. Pentru a obţine astfel de producţii bulbii trebuie să fie foarte mari. Forma bulbilor are importanţe ei, fiind preferată forma alungită celei aplatizate deoarece permite o desime mai mare de plante la unitatea de suprafaţă. Atingerea unor producţii mari depinde şi de precocitatea bulbificării. Genotipurile care bulbifică devreme au o stabilitate a producţiei de la un an la altul mai bună. Totuşi în această direcţie nu trebuie exagerat, deoarece formele foarte precoce au un potenţial productiv mai scăzut.
Depăşirea pragului de 80 t/ha este foarte greu de realizat şi ar puta fi posibilă numaI odată cu creşterea indicelui de durată foliară, adicăprelungirea duratei de viaţă a frunzelor. Semănatul din toamnă ar putea contribui la aceasta. Ca urmare, principala caracteritică ce trebuie îmbunătăţită ete rezistenţa la iernare combinată cu o maturizare cât mai târzie. De fapt, totul se rezumă la prelungirea duratei perioadei de vegetaţie (BREWSTER, 1977)
Un rol important în creşterea potenţialului productiv este mărimea bulbilor. Variabilitatea acestui caracter este foarte largă la ceapa pentru bulbi. La soiurile mai vechi, bulbii sunt mai mici (50-80 g.), dar sunt şi soiuri cu bulbi foarte mari, cu greutatea de până la 300 g. Cele mai extinse soiuri în cultură prezintă bulbi cu greutăţi de peste 100 g în cazul bulbilor sferic-turtiţi şi greutăţi de epste 200 g la soiurile cu bulbi alungiţi (IFRIM AURELIA şi BĂLAŞA, 2001). Ereditatea acestui caracter este complexă, efectele de bază fiind cele de aditivitate, prezentă fiind şi dominanţa.
Mărimea bulbilor depinde de numărul frunzelor pe plantă. De numărul frunzelor depinde procesul fotosintezei, dar şi realizarea bulbului. Bulbul fiind o metamorfozare a frunzelor, mărimea bulbului depinde de numărul frunzelor şi de grosimea acestora. Acest caracter este important şi în cazul cepei pentru frunze.
Ameliorarea calităţii. Pretenţiile comercializării impun anumite caractere morfologice care asigură o bună calitate. Diametrul bulbilor trebuie să fie de peste 8 am. Foile exterioare galbene arămii sau galbene aurii. Foarte importantă este rezistenţa foilor exterioare care asigură protecţie la manipulări şi în timpul păstrării. La mărirea duratei de păstrare în depozite contribuie şi conţinutul de substanţă uscată a foilor interioare care trebuie să fie cât mai ridicat.
Dintre caracterele cele mai importante care contribuie la realizarea calităţii sunt forma şi culoarea bulbilor, care sunt date de preferinţele pieţei, şi compoziţia biochimică.
Forma bulbilor este variabilă, de la plată până la foarte alungită. Cei mai preferaţi sunt bulbii globuloşi, aceştia având şi avantajul că se pot recolta mecanizat mult mai uşor. Determinismul genetic al acestui caracter este poligenic cu puternice efecte de dominanţă.
Culoarea bulbilor este diversă. Cele mai răspândite sunt soiurile cu bulbi de culoare galbenă, dar există şi zone unde se preferă soiurile cu bulbi de culoare roşie. Culoarea albă a bulbilor este mai rar întâlnită, pe piaţă fiind doar o curiozitate.
Din punct de vedere biochimic, substanţele care participă la realizarea calităţii sunt zaharurile şi uleiurile eterice. Aceste două componente contribuie la realizarea gustului. Conţinutul de zahăr este în jur de 5-6% din substanţa uscată. Determinarea uleiurilor eterice este mai dificil de realizat. Rolul lor este hotărâtor în realizarea gustului, în special a gustului iute. Uleiurile bogate în sulf imprimă un gust iute mai accentuat. Cei mai importanţi componenţi bogaţi în sulf sunt sulfura şi bisulfura de alil. (KUCKUCK ŞI KOBABE, 1962). S-a constatat că între gustul iute şi culoarea foilor exterioare există unele corelaţii.
Bulbii de culoare galbenă sunt mai iuţi decât cei de culoare roşie sau albă. De asemenea, bulbii mari sunt mai puţin iuţi decât cei mai mici.
Ameliorarea rezistenţei la boli şi dăunători. Ceapa este atacată de un număr mare de agenţi patogeni şi dăunători a căror importanţă este dată de pagubele pe care le produc, dar şi de posibilitatea de se face o combatere pe cale genetică. În cercetările de ameliorare au fost depistate surse de gene, însă spectrul raselor fiziologice şi agresivitatea acestora sporindu-se, găsirea unor surse noi este un obiectiv permanent.
Cele mai frecvente boli care apar în condiţiile din România sunt mana cepei (Peronospora destructor), putregaiul umed (Erwinia carotovora) şi putregaiul cenuşiu (Botrytis allii), iar dintre dăunători musca cepei (Hylemya antiqua).
Pentru rezistenţa la mană există variabilitate foarte mare, forme rezistente găsindu-se printre populaţiile locale sau la soiurile vechi. De asemenea, gene de rezistenţă există în cadrul speciei Allium altaicum sau A.chinensis, numai că transferarea lor este destul de dificilă. Genele implicare în realizarea acestui tip de rezistenţă sunt diverse. Există at’t gene minore cât şi gene majore. Cele mai cunoscute gene majore sunt recesive (s1, s2)
Determinismul rezistenţei la putregaiuri este mai puţin cunoscut. Ca surse de rezistenţă sunt semnalate speciile Allium ascalonicum şi A.fistulosum.
La nivel european se lucrează cu genitori cunoscuţi, cum este soiul Italia Red pentru rezistenţa la Peronospora schlieideni, Dorato di Parma pentru rezistenţa la Fusarium sp.; Dulce de Spania pentru rezistenţa la Phoma sp., mai multe soiuri cu bulbi galbeni sau roşii pentru rezistenţa la Colletotrichm sp.. Soiurile româneşti sunt tolerante la majoritatea agenţilor patogeni în condiţiile unor atacuri de intensitate medie.
Rezistenţa la boli poate fi sporită prin acordarea unei atenţii sporite hibridării îndepărtate. Această metodă are perspective noi de aplicare odată cu dezvoltarea unor tehnici ale iotehnologiilor, în spcial a culturii de embrioni imaturi.
Atacul de musca cepei nu poate fi combătut pe cale genetică, până în prezent nu sunt cunoscute forme care să nu fie atacate de acest dăunător. Se poate face numai o departajare a unor forme foarte sensibile sau mai puţin atacate.
Ameliorarea capacităţii la păstrare. La ceapa pentru bulbi, capacitatea de păstrare a bulbilor în timpul iernii este una dintre cele mai importante însuşiri. Deprecierea bulbilor în timpul păstrării are loc din cauza putrezirii şi a încolţirii prea devreme. Ambele aspecte sunt influenţate de condiţiile de păstrare, de condiţiile în care s-a realizat recolta, dar şi de valoarea genotipului. Putrezirea bulbilor pe parcursul păstrării se datorează atacului de boli în timpul vegetaţiei, ca urmare soiurile care se păstrează mai bine sunt cele mai rezistente la boli. Conţinutul de apă sin bulb poate favoriza dezvoltarea agenţilor patogeni, astfel că soiurile cu un conţinut mai mare de substanţă uscată au o capacitate de păstrare mai bună. Conţinutul bulbului în apă influenţează şi încolţirea, dar acest aspect este legat şi de starea de repaus specifică soiului, dar şi de umiditatea şi temperatura din depozit.
Selecţia pentru această însuşire trebuie realizată după conţinutul în substanţă uscată a bulbului, duritatea foilor interioare şi rezistenţa foilor exterioare.
Ameliorarea aptitudinilor pentru recoltare mecanizată. Din acest punct de vedere sunt necesare anumite cerinţe din punct de vedere morfologic. Cei mai favorabili sunt bulbii globuloşi, cu coletul fin, cu formă regulată, simetrici care se pot rostogoli uşor. Importantă este şi maturizarea rapidă şi uniformă. Pentru a nu se deprecia în timpul recoltatului, foile interioare ale bulbului trebuie să fie cărnoase şi dure, iar cele exterioare aderente de bulb şi rezistente pentru al proteja de loviturile care apar la recoltat şi în timpul transportului.
Ameliorarea soiurilor de ceapă destinate deshidratării. În ultimul timp ceapa se utilizează tot mai mult în industria alimentară, în special în prepararea concentratelor pentru supe. Pentru a avea un randament bun la prelucrare elementul cel mai important este conţinutul de substanţă uscată. Formele cu un procent mare de substanţă uscată dau un randament superior la deshidratare în condiţiile un or cheltuieli de producţie reduse. Între conţinutul de substanţă uscată şi productivitate există o corelaţie negativă greu de modificat. Din acest motiv, pentru industrializarea prin deshidratare este necesar să se creeze soiuri speciale care să prezinte procentajul de substanţă uscată corespunzător la un nivel al
productivităţii apropiat de cele maxime întâlnite la soiurile obişnuite. Pe lângă aceste însuşiri aparte, aceste soiuri trebuie să prezinte caracterele impuse de recoltarea mecanizată.
Ameliorarea soiurilor de ceapă de toamnă. Ceapa de toamnă se cultivă în Europa pe suprafeţe restrânse, dar este o cultură foarte extinsă în Asia, în special în Japonia. În Europa, tehnologii specifice cepei de toamnă apar în producerea de sămânţă la ceapa pentru bulbi.
La ceapa de toamnă cea mai importantă caracteristică este precocitatea bulbificării şi maturizarea rapidă care permit recoltarea începând cu luna mai, perioadă în care bulbii din recolta anului trecut nu mai pot fi păstraţi. În condiţiile României, ceapa de toamnă poate fi cultivată pentru producţii timpurii în zonele cu climă mai blândă. Pentru acest mod de cultură, prin ameliorare trebuie create soiuri care să prezinte o bună rezistenţă la iernare, semănatul făcându-se toamna. În primele faze de vegetaţie, după răsărire, ritmul de creştere trebuie să fie rapid, apoi trebuie să încetinească pentru a se realiza acumulările necesare rezistenţei la condiţiile de iernare. Germoplasma existentă permite realizarea unor asemenea caracteristici fiziologice. Problemele care se rezolvă foarte greu la acest tip de ceapă sunt fermitatea slabă a bulbilor şi nedezvoltarea suficientă şi corespunzătoare a foilor exterioare cu rol de protecţie. Astfel, producţie acestor soiuri trebuie consumată într-o perioadă scurtă deoarece nu prezintă capacitate de păstrare. Pentru asigurarea continuităţii consumului în perioada de vară, la acest tip de ceapă este necesară diversificarea sortimentului de soiuri după perioada de vegetaţie. Pentru a mări rezistenţa la păstrare în perioada de vară se pot utiliza genitori din zonele tropicale care sunt adaptaţi depozitării bulbilor recoltaţi în condiţii de temperaturi şi umiditate mai ridicate. Ameliorarea pe baza acestei germoplasme este dificilă deoarece de la formele tropicale pot fi preluate şi unele caractere nedorite, cum este o slabă rezistenţă la temperatri scăzute.
Ameliorarea soiurilor pentru ceapă verde. Cultura pentru ceapă vede este realizată în scopul valorificării foliajului. Cultura de ceapă verde se înfiinţează toamna, chiar la sfârşitul verii, iar valorificarea se face primăvara devreme. Caracterele importante pentru asemenea soiuri sunt o foarte bună rezistenţă la iernare, bulbificare foarte precoce şi un foliaj cât mai bogat. Frunzele trebuie să fie în număr mare, foarte lungi şi subţiri, drepte şi fragede.
AMELIORAREA MORCOVULUI
Morcovul este o specie introdusă în cultură mult mai târziu decât alte legume cu toate că a fosr cunoscută ca plantă medicinală. O mare perioadă de timp, morcovul a fost consumat din natură. După luarea în cultură, preogresele obţnute prin ameliorare au apărut la scurt timp, ca urmare astăzi există un număr mare de soiuri cu caractere productive şi calitative deosebite. Comparativ cu formele sălbatice, soiurile actuale au suferit modificări morfologice majore.
Ca urmare a cultivării şi selecţiei, rădăcina subţire a morcovului sălbatic a devenit mult mai mare, cărnoasă şi fragedă bogată în zaharuri şi vitamine. Morcovul prezintă o plasticitate ecologică foarte pronunţată astfel că ameliorarea poate duce la crearea unei game de cultivare care să răspândească această cultură pe areale tot mai extinse utilizarea tot mi largă în consum în stare proaspătă sau preparat în diferite moduri, impune diversifciarea lucrărilor de ameliorare în diferite diferite direcţii.
În urma ameliorăii, sortimentul de soiuri s- divrsificat foarte mult, atât în privinţa formei rădăcinii cât şi a calităţii. Pretenţiile faţă de calitatea rădăcinilor au crescut tot mai mult, urmare a acestora apărând soiuri cu rădăcini foarte bogate în zahăr (peste 10%) şi în vitaminele A, B şi C. Rezultate deoasebite au fost obţinute şi în privinţa potenţialului productiv, potenţialul biologic putând depăşi 50 t/ha. La asemenea valori s-a ajuns prin introducerea în cultură a hibrizilor comerciali.
Obiective de ameliorare.
Amelioarrea morcovului a progresat mult, în multe ţări, soiurile fiind înlocuite cu hibrizi comerciali. Progrese mari au fost realizate şi în privinţa calităţii, procentul de zahăr şi vitamine au fost mult îmbunătăţite, la fel ca frăgezimea rădăcinii şi capacitatea de păstrare.
Ameliorarea capacităţii de producţie. Productivitatea la morcov este condiţionată de o serie de caractere şi însişiri. Producţia la unitatea de suprafaţă este dată de desimea plantelor şi productivitatea unei plante.
Desimea plnatelor la unitatea de suprafaţă este dependentă de mărimea rădăcinilor dar şi de mărimea rozetei de frunze. Pentru a se permite o creştere a desităţilor la unitatea de suprafaţă, frunzele din rozetă trebuie să fie poziţionate cât mai aproape de verticală, astfel obţinându-se şi o mai bună valorificare a luminii. Desimile prea mari nu sunt benefice deoarece rădăcinile pot creşte deformat (contorsionat şi neregulat).
Caracterele morfologice care influenţează capacitatea de producţie sunt: forma rădăcinii, lungimra rădăcinii, diametrul rădăcinii şi greutatea acesteia. Nivelul producţiilor care se pot atinge prin cultivarea unor soiuri cu parametrii morfologici corespunzători, este de peste 40-45 t/ha. (POTLOG şi colab., 1982) Prin cultivarea hibrizilor, la care uniformitatea recoltei ste foarte ridicată, producţiile care se pot obţine pot tinde spre limita de 50 t/ha.
Forma rădăcinii poate fi diversă, de la forma aproape sferică până la forma cilindrică sau conic alungită. Sunt preferate formele alungite pentru a obţine producţii mai mari la unitatea de suprafaţă. Pe solurile uşoare, unde dislocarea rădăcinilor la recoltat se face cu uşurinţă, se pot cultiva şi soiurile cu rădăcini cilindrice, acestea având un volum şi o greutate mai mare. Pe solurile unde dislocarea se face mai dificil sunt preferate rădăcinile uşor conice cu vârful rotunjit, pentru că nu se rup la recoltat.
Lungiumea nu poate trece de animite limite pentru că la recoltat apar pierderi, mai ales în cazul rădăcinilor foarte adânci şi subţiri. Lungimea optimă a rădăcinii de morcov este de 20-25 cm. La asemeneas dimensiuni în timpul recoltatului nu se produc pierderi, iar pe parcursul trasportului şi depozităirii rădăcinile se pot păstra întregi. La varietăţile de morcov furajer rădăcinile pot ajunge până la 30 cm, sau chiar mai lungi. La acestea partea lemnoasă fiind mai dezvoltată asigură o bună rezistenţă la rupere. Pentru consumul alimentar, rădăcinile cu cilindrul central foarte dezvoltat nu sunt preferate pentru că sunt dure.
Diametrul rădăcinii este o altă componentă care contribuie la realizarea volumului rădăcinii. Valoarea diametrului trebuie să fie de 5-6 cm. La morcovul furajer, grosimea rădăcinii poate depăşi valoarea de 10-12 cm. Grosimea prea mare face ca cilindrul central să fie mai dezvoltat în detrimentul calităţii.
Diametrul rădăcinii este un cracter mai puţin stabil compartiv cu lungimea rădăcinii. Valorile diametrului sunt influenţate de condiţiile de cultură, în special de num ărul plantelor la unitatea de suprafaţă. Din acest motiv selecţia hotărâtoare pentru capcitatea de producţie se realizează după lungimea şi greutatea rădăcinilor.
Greutatea rădăcinilor este este principala componentă a productivităţii. Acest caracter depinde de altle, adică de volumul rădăcinii şi de conţinutul în substanţă uscată. Rădăcinile de morcov pot avrea greutăţi de 35 g, până la peste 250 g, iar la morcovul furajer, chiar aproape de 500 g. rădăcinile fragede, bogate în apă, sunt mai grele. Prin pierderea apei scade şi greutatea rădăcinii. Procentul de substanţă uscată din rădăcină poate depăşi valoarea de 14%, dar la majoritatea soiurilor pentru consum curent este în jur de 10-12%. La soiurile care trebuie păstrate în timpul iernii, procentul mai mare de substanţă uscată îmbunătăţeşte capacitatea de păstrare. (POTLOG şi colab., 1982) Greutăţile rădăcinilor, la soiurile actuale sunt apropiate de 150-200 g, în condiţii tehnologice obişnuite. (IFRIM AURELIA şi BĂLAŞA M., 2001)
Ameliorarea calităţii. Calitatea rădăcinilor de morcov are mai multe componente de ordin morfologic şi biochimic. Componentele morfologice sunt culoarea rădăcinii şi dezvoltarea cilicndului central (sau inima rădăcinii)
Culoarea rădăcinii este relizată pe baza prezenţei carotenului. Culorile întâlnite pot de alb, galben, portocaliu-cărămiziu, sau diferite nuanţe de violet la unele subsepcii asiatice. În zona cilindrului central culoarea este mai deschisă. Din acest motiv culoarea se apreciază după intensitatea ei, dar şi după uniformitatea acesteia în secţiune transversală. Intensitatea culorii este corelată şi cu conţinutul de substanţă uscată. Morcovii intens coloraţi au şi un conţinut de substanţă uscată mai mare şi nu sunte preferaţi de către consumatori. Variaiblitatea conţinutului de carotină, ca urmare şi a intensităţii culorii, este destul de mare. La soiurile extinse în cultură, carotina reprezintă 6-12 mg la 10 g substanţă uscată,
dar sunt menţionate şi valori de 25 mg la 10 g substanţă uscată. (BANGA, 1962). Repartizarea carotinei de-a lungul rădăcinii este diferită. În treimea inferioară a rădăcinii valorile sunt mai reduse, de aceea determinările se efectuează la mijlocul rădăcinilor. În funcţie de conţinutul de cortină sunt delimitate cinci clase de culoare, după care se poate efctua selecţia. Stabilirea culorii se face prin efectuarea de secţiuni transversale în rădăcină, sau combinat, prin secţiunin transversale şi lomgitudinale. În rădăcina de morcov există două tipuri de carotina.: carotina α şi carotina β. Cea mai activă este carotina β. Pe lângă carotină, în rădăcina de morcov mai există şi alţi compuşi carotinoizici, dar procentajul lor este foarte redus, ca urmare şi importanţa lor este nesemnificativă. Stabilirea cantităţii şi a tipului de carotină presupune efectuarea unor analize chimice laborioase, şi de cele mai multe ori selecţiam se face numai prin observaţii vizuale asupra culorii. Conţinutul în carotină prezintă un determinism genetic complex neelucidat încă. Se pare că un rol important în au efectele de aditivitate. (ARDELEAN, 1986)
Mărimea cilindrului central este o altă componentă a calităţii rădăcinii de morcov. Această zonă a rădăcinii este colorată mai puţin intens, este mai puţin fragedă şi mai săracă în zahăr şi vitamine. Singura importanţă a acestei porţiuni este aceea că imprimă rezistenţă la ruperea rădăcinilor în timpul recoltatului şi a transportului. În selecţie se vor reţine formele la coloraţia rădăcinii este uniformă şi cilindrul cenral este mai redus. Reducerea cilindrului central nu trebuie exagerată nu numai pentru că se va micşora rezistenţa la rupere, ci şi pentru că semincerii vor avea un număr redus de tulpini florifere. Selecţia în funcţie de mărimea cilindrului central se poate realiza şi cu ajutorul unui parametru sintetic, denumit indice de calitate. Acesta se determină ca raport între diamaetul rădăcinii şi diametrul cilindrului central. Valorile optime sunt cuprinse între 2 şi 3. (ARDELEAN, 1982). Variabilitatea mărimii şi coloraţiei acesteia este mare, cunoscându-se şi genotipuri favorabile din acest punct de vedere.
Compoziţia biochimică a morcovului este complexă, motiv pentru care produsele din morcov sunt indicate în alimentaţia copiilor. Prin ameliorare, trebuie depăşiţi parametrii calitativi ai actualelor soiuri. Glucidele sunt un compuşi organici foarte importanţi, care în rădăcina de morcov trebuie să fie peste 10 %. Pe lângă glucide, există şi cantităţi mici de proteine ( apropape 2%) şi lipide ( sub 1%). Foarte importantă este vitamina C, care trebuie să depăşească valoarea de 7 mg/100 g produs, β carotenul care trebuie să fie peste 10 mg la 100 g produs. Pe lângă acestea foarte importante şi cantităţile de potasiu, calciu şi sodiu. (DINICĂ MONICA şi colab., 2002) Pe lângă compuşii de bază amintiţi, în cantităţi mai rerduse se găsesc şi vitaminele E, K, B1, B2, PP, unele metale (fier, cupru, zinc) şi un spectru larg de aminoacizi. (DINCĂ MONICA, 2002) Toate aceste elemnte trebuie luate în considerare în procesulo de ameliorare deoarece consumul morcovilor în hrana copiilor este tot mai extins.
Ameliorarea rezistenţei la boli şi dăunători. Cele mai frecvente boli ale morcovului sunt putregaiurile: putregaiul umed produs de Erwinia carotovora şi putregaiul uscat produs de Sclerotinia slerotiorum. Studiile în privinţa rezistenţei la aceste două boli sunt destul de puţine, astfel că nu sunt semnalate gene specifice de rezistenţă. Atacul are aceleaşi intensităţi la toate subspeciile celor două specii. Pentru găsirea unor forme mai tolerante ar trebuie intensificate căutările în germoplasma locală. Dificultăţile legate de studiul acestotr boli se datorează şi faptului că atacul este influenţat şi de condiţiile tehnologice, reducerea frecvenţei atacului putându-se obţine şi prin măsuri preventive. Pe lângă aceste boli, în cultrile de morcov mai sunt semnalate făinarea (Esysiphe umbeliuferarum), cercosporioza morcovului (Cercosporiosa carotae), fuzarioza morcovului ( Fusarium oxysporum) şi mozaicul morcovului. (GLĂMAN şi colab., 2003)
Dintre dăunători, cel mai frecvent este musca morcovului (Psila rosae). La fel ca în cazul bolilor, nu se cunosc forme care să nu fie atacate de acest dăunător.
Ameliorarea precocităţii. În privinţa precocităţii, gama soiurilor este destul de restrânsă deoarece deoarece necesităţile pieţei nu impun producţii extratimpurii. Păstrarea mrcovului în depozite se poate face până la începutul verii, perioadă în care se poate începe recoltarea noii culturi. Producţiile timpurii sunt destul de mici, dar prin preţul de valorificare, cultura poate fi rentabilă. Pentru consumul din vară până toamna, trebuie create soiuri care să aibe o dezvoltare foarte rapidă în prima etapă de forma a rădăcinii. La soiurile tardive, prima etapă de creşere este mai lentă, dar rădăcinile care se formează sunt mult mai mari. La soiurile timpurii rădcinile sunt foarte fragede şi capacitatea de păstrare este mai redusă.
Ameliorarea rezistenţei la emiterea tulpinilor florifere în primul an. Emiterea tulpinilor florifere în primul an este o deficienţă care duce la scăderea producţiei şi a calităţţii. Rădăcinile plantelor care au
emis tulpini florifere aunt mult mai mici şi cu cilindrulc entral foarte bine dezvoltat. Determinsimul genetic al acestei însuşiri este foarte complex şi necunoscut. O influenţă foarte mare o au condiţiile de mediu, dintre care temperatura.
AMELIORAREA FASOLEI Fasolea comună este principala sursă de proteine în alimentaţia umană în numeroase ţări în curs
de dezvoltare. Proteina de fasole este foarte valoroasă din punct de vedere calitativ, conţinând majoritatea aminoacizilor esenţiali, la un preţ mult inferior proteinelor de origine animală. Statisticile, destul de imprecise, indică producţii destul de scăzute la unitatea de suprafaţă. Aceasta se datorează faptului că este o plantă extensivă. Producţiile la hectar diferă foarte mult de la o ţară la alta. Nivelurile sunt foarte ridicate în vestul Europei şi nordul Americii şi extrem de mici în celelalte zone.
Fasolea a fost luată în cultură de foarte mult timp în America de Sud, aici găsindu-se şi cele mai mari suprafeţe cultivate. După aducerea în Europa, suprafeţele cultivate s-au extins foarte repede. Încă de la introducerea în Europa, au fost consumate atât boabele cât şi fructele. Selecţia empirică a cultivatorilor a fost foarte insistentă în direcţia eliminării progresive a structurilor fibroase din păstaie pentru a ajunge la tipul de păstaie fină, apoi la tipul de plantă de la care se consumă atât boabele cât şi păstăile. În carul variabilităţii naturale au fost descoperite forme la care boabele rămân verzi mai multă vreme. Astfel a fost creat un nou tip de fasole, cea cultivată pentru boabe verzi. Suprafeţele cultivate cu asemenea soiuri sunt destul de reduse, la noi în ţară, soiuri de acest tip nu se cunosc.
Pentru cultivatori, fasolea aduce unele avantaje, fiind o bună premergătoare pentru majoritatea culturilor. Suprafeţele reduse se datorează unor impedimente legate de mecanizarea lucrărilor de întreţinere şi recoltat, cât şi datorită cerinţelor ridicate faţă de condiţiile de mediu şi tehnologie
Obiective de ameliorare.
Diversele tipuri de fasole şi modurile de utilizare implică un număr mare de obiective de ameliorare. Eforturile ameliorării trebuie să fie concentrate spre fiecare tip în parte. Elementele definitorii ale fiecărui tip sunt, ele însele obiective de ameliorare. Cu toate acestea sunt şi obiective comune, cum sunt. Rezistenţa la boli şi dăunători, creşterea productivităţii şi stabilitatea producţiei, calitatea producţiei.
Ameliorarea capacităţii de producţie. Productivitatea la fasole este realizată pe baza unui complex de factori care influenţează numărul păstăilor pe plantă şi numărul boabelor în păstaie. Nu trebuie uitată nici densitatea la unitatea de suprafaţă. Ameliorarea a adus un aport mare la îmbunătăţirea multor componente ale capacităţii de producţie. În condiţii de câmp, la fasolea pentru păstăi, numărul plantelor la unitatea de suprafaţă poate trece de 25 plante/m2, în condiţii de seră, cultivând forme speciale acest element tehnologic poate avea valori de 5-15 plante/m2, forme la care producţia pe plantă este mult sporită. La fasolea pentru boabe desimile sunt mai mari. În crearea soiurilor destinate cultivării pentru boabe se urmăreşte obţinerea unor forme cu tufă compactă şi creştere limitată care să permită realizarea unor desimi de 40 plante/m2. Realizarea unui anumit număr de plante la unitatea de suprafaţă depinde de portul plantei. La fasolea cu creştere determinată se urmăreşte menţinerea unei talii relativ înalte pentru a se putea obţine un număr mare de păstăi pe plantă, dar cu o bună rezistenţă la cădere. Creşterea taliei trebuie să se realizeze foarte rapid, iar înflorirea să se desfăşoare pe o perioadă de timp cât mai redusă pentru a uniformiza maturizarea. Desimea mai depinde şi de unele însuşiri fiziologice, cum sunt: rezistenţa la secetă şi rezistenţa la boli. Formele sensibile din acest punct de vedere nu suportă desimi mari. O altă însuşire importantă care influenţează densitatea este rezistenţa la brumele târzii din primăvară. Genotipurile sensibile pierd plante din cauza temperaturilor scăzute care pot să apară în primele faze de vegetaţie.
Productivitatea unei plante are mai multe componente de ordin morfologic: numărul păstăilor fertile, numărul boabelor din păstaie, mărimea şi greutatea boabelor. (MUREŞAN şi CRĂCIUN, 1971)
Numărul păstăilor pe plantă variază în funcţie de tipul de creştere şi perioada de vegetaţie. La formele cu creştere limitată, numărul păstăilor pe plantă variază între 15 (la formele precoce) şi 25 (la cele tardive), acestea fiind şi limitele minime acceptate în procesul de selecţie. Aceste limite pot fi atinse
la fasolea de grădină, ultimele soiuri româneşti prezentând peste 12 păstăi pe plantă, în funcţie de condiţiile climatice. (LASCU, 2003; IFRIM AURELIA şi BĂLAŞA, 2002)
Variabilitate foarte pronunţată există şi pentru numărul boabelor din păstaie. Elitele trebuie să prezinte păstăi cu lungimea de cel puţin 6 cm, conţinând minim 6-7 boabe în păstaie. Având în vedere aceşti parametrii, pe o plantă trebuie să se realizeze, în medie, cel puţin 60 de boabe la soiurile precoce şi 90 de boabe la cele tardive.
Prin ameliorare a crescut foarte mult producţia de boabe pe plantă, ajungându-se până la 50 g/plantă în condiţii de câmp sau 100 g/plantă în condiţii de seră. La creşterea producţiei pe plantă poate contribui în mod hotărâtor şi heterozisul, care prezintă limite largi de variaţie, de la 20% până la peste 100%. În plus, prin crearea noilor soiuri s-a căutat reducerea perioadei de vegetaţie, la mai puţin de 4 luni la soiurile destinate a fie cultivate în câmp, şi la numai 3 luni în cazul soiurilor şi hibrizilor pentru sere. Astfel, în sere se pot obţine chiar 4 generaţii pe an.
La fasolea pentru boabe, producţia pe plantă este dată de numărul păstăilor, numărul boabelor în păstaie şi mărimea boabelor. Dintre acestea, caracterul cel mai stabil este mărimea boabelor. Mărimea boabelor se exprimă cel mai frecvent prin intermediul masei a 1000 de boabe. Pentru acest caracter, variabilitatea în cadrul genului este foarte pronunţată, fiind cuprinsă între 150 g şi 1250 g. La fasole există mai multe categorii de soiuri după acest aspect. Masa a 1000 de boabe, la soiurile cultivate poate fi cuprinsă între 150 şi 650 g. Din acest punct de vedere, la nivel mondial există cinci categorii de soiuri de fasole pentru boabe. Patru dintre acestea prezintă boabele de culoare albă şi o categorie, boabele de culoare verde. Soiurile cu boabe de culoare albă pot fi cu boabe foarte mari (MMB = 550-650 g), cu boabe mari (MMB = 450-550 g), cu boabe mijlocii ( MMB = 350-450 g) şi cu boabe mici ( MMB = 150-250 g). Se consideră că cele cu boabe mari au şi o calitate mai bună. Soiurile cu boabe de culoare verde, au boabele mici ( MMB = 180-250 g), au vigoarea plantelor redusă, iar boabele pot fi colorate în diferite nuanţe de verde şi galben. Producţiile de boabe la nivel mondial sunt destul de mici. Randamentul de sub 2000 kg/ha nu satisface o agricultură intensivă. Motivarea culturii de fasole pentru boabe începe de la producţia de 2500 kg/ha. Această producţie poate fi realizată fără dificultate dacă se cultivă soiuri performante. Conservarea formelor actuale are o mare importanţă în ameliorarea productivităţii la fasole. Pentru a proteja variabilitatea genetică sunt necesare soiuri din toate tipurile.
La fasolea pentru păstăi proaspete notarea producţiei totale are o importanţă discutabilă, deoarece preţul de vânzare a păstăilor este variabil. Recoltarea se face eşalonat şi producţia de păstăi creşte treptat pe parcursul perioadei de vegetaţie, dar calitatea scade la fel ca preţul de vânzare. Ca urmare este foarte important ca planta să producă un număr mare de păstăi la primele recoltări când calitatea şi preţul sunt foarte bune.
Ameliorarea calităţii. La fasolea pentru păstăi, păstaia poate avea două structuri fibroase. O structură longitudinală care formează “aţele” şi una oblică, mai puţin cunoscută. Aceste structuri fibroase se dezvoltă în cursul creşterii păstăii şi face ca după aproximativ 10 zile, păstăile să nu mai poată fi consumate. Păstăile cu structură longitudinală, aţoasă, trebuie recoltate foarte tinere, pe când cele cu structură oblică, care nu prezintă aţe pot fi recoltate şi mai târziu. Alte caractere de calitate sunt culoarea păstăilor, cilindricitatea şi rectilinitatea. Se consideră că optimul pentru calitate este când substanţa uscată în păstaie ajunge la 10-12%, indiferent de mărimea păstăilor. Acest procentaj se atinge la o vârstă a păstăilor de 10-15 zile (OLIVIER, 1971; FOUILLOUX, 1976). Pentru a nu pierde din calitate recoltările, trebuie efectuate succesiv, în perioada optimă. În practică, recoltarea nu se face după conţinutul în substanţă. În practică, criteriul de bază, este calibrul păstăilor şi fineţea lor. Ameliorarea a lucrat în direcţia creării unor soiuri cu păstăi foarte fine, astfel că acest caracter este foarte bine fixat genetic. Cerinţele actuale direcţionează selecţia spre păstăi extrafine în scopul, înlocuirii soiurilor cu păstăi aţoase. Din punct de vedere biochimic, păstăile de fasole conţin vitamina C, vitamina B1, vitamina B2, provitamina A.
Pentru fasolea pentru boabe, prezenţa fibrelor este apreciată deoarece uşurează treieratul. Calitatea la fasolea pentru boabe este destul de arbitrar definită. Pentru industrializarea boabelor uscate, acestea trebuie să absoarbă o cantitate cât mai mare de apă pentru ca produsul, obţinut să aibă frăgezimea necesară. Frăgezimea este dependentă de genotip dar şi de apa cu care se lucrează. Calitatea culinară a boabelor de fasole depinde şi de gustul acestora. Se pare că există o savoare care diferă de la un soi la
altul. Ca şi criterii de selecţie pentru calitate culinară a boabelor sunt grosimea cojilor şi prezenţa substanţelor antinutriţionale cum sunt alfa-galactocidele care sunt răspunzătoare de balonare.Din punct de vedere biochimic se urmăreşte creşterea conţinutului de proteină. Pentru această însuşire, variabilitatea este cuprinsă între 12 şi 35%. Soiurile cultivate se situează la mijlocul intervalului de variabilitate (23-25%). În procesul de selecţie, pe lângă conţinutul de proteină, se urmăreşte şi îmbunătăţirea calităţii acesteia. Proteina din boabele de fasole este printre puţinele proteine de origine vegetală, bogate în lizină şi triptofan. Din spectrul aminoacizilor esenţiali, lizina reprezintă peste 4%, iar triptofanul peste 1,4%. Din punct de vedere a calităţilor culinare interesează o bună capacitate de fierbere foarte bună şi un procent redus de coji. Fierberea trebuie să se realizeze în maxim o oră, realizându-se şi o uniformitate corespunzătoare. În laborator această însuşire se testează prin determinarea capacităţii de fierbere a unui bob. Coeficientul de fierbere obţinut trebuie să fie sub valoarea 7.Pentru îmbunătăţirea calităţii se urmăreşte şi scăderea procentului de coji. Valoarea maximă admisă trebuie să se situeze sub 6%. Îmbunătăţirea acestui caracter se realizează destul de greu, mai cu seamă atunci când se doreşte obţinerea unor genotipuri cu boabe mari. Mărimea boabelor este corelată direct cu procentul ridicate de coji, ca urmare formele cu procent redus de coji prezintă boabe mici.
Ameliorarea rezistenţei la boli . Primele lucrări de selecţie la fasole au avut ca scop îmbunătăţirea rezistenţei la boli. Aceasta a fost justificată de pagubele importante datorate atacului diferiţilor agenţi patogeni. Selecţia pentru rezistenţă la boli permite ameliorarea concomitentă şi a productivităţii şi calităţii. Principalele boli şi sursele de rezistenţă sunt prezentate în tabelul 3.7.
Cele mai importante boli for fi prezentate în continuare. Mozaicul comun (Virus 1) deformează frunzele şi diminuează considerabil producţia. Pentru
această boală există două sisteme de rezistenţă. Primul sistem este de tip imunitate şi este legat de prezenţa genelor recesive. Acest sistem se
bazează pe teoria genă pentru genă şi a fost aplicat utilizând variabilitatea parazitului şi a gazdei. Datorită vulnerabilităţii, acest sistem nu se mai utilizează singur.
Al doilea sistem este de tip hipersensibilitate dată de gena “I” prezentă la soiurile Processor şi Cabanais. Acest sistem este folosit la fasolea pentru păstăi. Multă vreme nu a fost utilizat la fasolea pentru boabe, dar în ultima vreme a fost încercat şi în această direcţie. Dezavantajul acestui sistem este că la temperaturi mari, de peste 300C, se poate declanşa o reacţie mortală a plantelor. Cu toate acestea, aproape toate soiurile moderne prezintă gena de hipersensibilitate, deoarece în cultură apariţia acestui sindrom nu întruneşte condiţii decât în anumiţi ani.
Gene pentru rezistenţă la viroze se pot prelua de la speciile Phaseolus coccineus, Phaseolus aureus,, Phaseolus mungo
Antracnoza (Colletotrichuim lindemuthianum) este provocată de o ciupercă ce distruge plantele tinere şi produce pete mari pe păstăi şi boabe. În unele regiuni, această boală este un element limitant al culturii. Genele de rezistenţă la această boală sunt cunoscute de mult timp. Progresul decisiv în ameliorarea rezistenţei la această boală a fost realizat odată cu descoperirea genei “ARE”, genă care conferă rezistenţă pentru patotipurile cunoscute în Europa. (MASTENBROEK, 1960) Graţie punerii la punct a unui test de infecţie artificială şi datorită eredităţii simple au fost create soiuri cu o foarte bună rezistenţă. În vestul Europei, majoritatea soiurilor prezintă gena “ARE” şi rezerva de agent patogen s-a redus foarte mult. În condiţiile României, cultivându-se multe populaţii locale, boala este încă prezentă şi constituie un obiectiv de ameliorare pentru toate tipurile de fasole.
Având în vedere pericolul constituit de utilizarea unei singure gene, au mai fost găsite şi alte surse de rezistenţă totală. Astfel, în germoplasma mexicană au fost depistate genele “MEX” 1-3 (FOUILLOUX, 1976). Multă vreme noile gene au acoperit toate rasele europene ale agentului patogen. Recent, au fost depistate rase locale noi, în Brazilia şi Ruanda, faţă de care genele “MEX” nu sunt eficace.
Tabelul 2.1.Principalele boli ale fasolei şi sursele de rezistenţă. (după FOUILLOUX şi BANNEROT, 1992)
Pătarea aureolată (Pseudomonas phaseolica) este o boală bacteriană care provoacă pagube mari în anii cu umiditate ridicată. Dacă atacul se produce asupra plantelor tinere, acestea pot fi distruse foarte rapid. Atacul se manifestă prin pete create de coloniile de bacterii în jurul cărora se difuzează toxine. Atacul se poate manifesta şi pe păstăi sub formă de pete unsuroase.
Împotriva acestei boli se cunoaşte toleranţa provenită de la linia PI 150414. Ereditatea acestei toleranţe este complicată, fiind implicate trei gene majore, mai mult sau mai puţin recesive. (HILL şi colab., 1972; FOUILLOUX, 1975). Testarea infecţiilor artificiale este greoaie din acest motiv selecţia este de durată. Pentru aceasta se cunosc soiuri rezistente extinse în cultură (Gratiot, Sanilac ) sau se pot prelua gene de la specia Phaseolus aborigeus.
La fasole mai există şi alte boli (rugina, mozaicul galben), dar fiind specifice zonelor tropicale, în arealul european nu se întâlnesc decât foarte rar. În condiţiile nerespectării unei rotaţii corespunzătoare, poate fi important şi atacul de fuzarioză, boală provocată de un complex de trei agenţi patogeni. Crearea formelor rezistente pentru această boală este dificilă, numărul genelor implicate fiind prea mare. Rezistenţa la rugină poate fi îmbunătăţită prin transferul de gene de la Phaseolus coccineus, Phaseolus aureus, Phaseolus acutifolius, Phaseolus mungo.
În ultimul timp a fost semnalat tot mai des pe plan european atacul de mozaicul castraveţilor faţă de care nu se cunosc surse de rezistenţă în cadrul speciei Phaseolus vulgaris.
Ameliorarea rezistenţei la dăunători. Cel mai frecvent şi periculos dăunător al fasolei este gărgăriţa. Atacul de gărgăriţă se poate îmbunătăţi prin modificarea unor însuşiri legate de înflorit, perioada de realizare a păstăilor sau de constituţia boabelor. Gene de rezistenţa la atacul de gărgăriţă pot fi preluate din soiurile Michigan sau Robust, ori de la specia Phaseolus lunatus.
Ameliorarea nutriţiei cu azot. Fiind o leguminoasă, fasolea poate fixa azotul atmosferic, dar se impune o îmbunătăţire a acestei activităţi. Eficacitatea fixării azotului se măsoară prin conţinutul de ureide. Metodele de determinare a acestora sunt destul de imprecise. În plus, ameliorarea acestui caracter este îngreunată de faptul că o bună capacitate de fixare a azotului este direct corelată cu tardivitatea.
Ameliorarea precocităţii. Precocitatea este foarte importantă în cultura fasolei. Datorită sensibilităţii fasolei faţă de temperaturi ridicate şi faţă de secetă se recomandă cultivarea soiurilor precoce, care în perioadele critice în privinţa acestor factori, sunt mai bine dezvoltate şi sunt mai puţin afectate.
La fasolea pentru păstăi precocitatea este importantă şi în direcţia obţinerii, mai degrabă a unui sortiment de soiuri care să furnizeze cu regularitate producţie proaspătă pe piaţă sau pentru conservare.
La fasolea pentru boabe, precocitatea este legată de adaptabilitate. Cu toate că se ştie că formele mai tardive sunt mai productive, în cazul lungirii perioadei de vegetaţie producţia poate fi compromisă de factorii de climă.
Ameliorarea pretabilităţii pentru recoltarea mecanizată. Recoltarea mecanizată reduce din costurile cu lucrările manuale. Principalele cerinţe din acest punct de vedere sunt crearea unor soiuri cu talie uniformă, rezistente la cădere şi cu maturizare omogenă a păstăilor. Acestea sunt caractere necesare atât la fasolea pentru boabe cât şi la cea pentru păstăi, mai ales la soiurile destinate conservării.
La fasolea cultivată pentru păstăi destinate consumului imediat recoltarea nu se poate face decât manual Pentru a scădea costul recoltatului manual, trebuie selecţionate forme la care recoltarea să se efectueze printr-o singură trecere, la care maturizarea să se realizeze grupat. Pentru a creşte productivitatea culegătorilor, portul, plantei trebuie să fie mai înalt, iar păstăile mai lungi şi mai grele. Realizarea unor păstăi lungi, drepte şi fără aţe este destul de dificilă deoarece, în general, păstăile lungi nu sunt drepte. Alungirea păstăii se face pe baza îmbogăţirii în parenchim. Obţinerea unor păstăi lungi şi cu aspect plăcut nu este posibilă. De aceea lungimea păstăii trebuie păstrată în limite rezonabile, la soiurile pitice în jur de 17-19 cm, iar la cele volubile, pot tinde spre 30 cm.
La fasolea destinată conservării recoltarea se face cu maşini speciale, care realizează o “pieptănare” a plantelor într-o singură trecere. Condiţia principală din acest punct de vedere este maturizarea foarte grupată. Pentru a evita pierderile excesive la recoltarea mecanizată a păstăilor, acestea trebuie să fie bine repartizate de-a lungul plantei şi să fie inserate la cel puţin 10 cm de la nivelul solului. Aceasta presupune plante relativ înalte. Pentru a avea şi o calitate bună a recoltatului, păstăile nu trebuie rupte în ciorchini, ca urmare fiecare păstaie trebuie să se detaşeze uşor, pentru a nu fi smulse plante la recoltat, înrădăcinarea trebuie să fie adâncă, tulpinile rezistente la cădere şi frângere, iar ramificaţiile inflorescenţelor puternice. Prinderea păstăilor pe ramificaţii trebuie să fie cât mai fragilă. În cazul în care conservarea se va face sub formă de păstăi întregi, acestea trebuie să fie mai scurte şi foarte uniforme. Lungimea optimă este de 11-12 cm. Pentru ca sucul din conserve să nu fie colorat, boabele trebuie să fie albe. Chiar dacă bobul nu este ajuns la maturitate, culoarea lui se transmite în suc în urma fierberii.
Ameliorarea rezistenţei la temperaturi excesive. Temperaturile excesive, atât cele ridicate cât şi cele scăzute sunt foarte dăunătoare plantelor de fasole.
Temperaturile scăzute, apropiate de 00C sunt foarte dăunătoare în primele faze de vegetaţie. Pentru îmbunătăţirea rezistenţei la temperaturi scăzute se pot efectua hibridări cu speciile înrudite sau poate fi utilizată cu succes mutageneza.
Temperaturile ridicate sunt foarte dăunătoare în perioada de înflorire. Asociate cu seceta, aceste condiţii de mediu afectează înflorirea şi fecundarea. Pentru a evita perioadele cu condiţii de stres, se recomandă obţinerea soiurilor precoce, cu o perioadă de vegetaţie de 70-90 zile sau includerea în genotip a unor gene de rezistenţă preluate de la speciile înrudite mai rezistente.
Ameliorarea rezistenţei la secetă. Seceta este periculoasă aproape în toate fazele de vegetaţie. Cele mai mari pagube le produce când este asociată cu temperaturile ridicate. Pe lângă cultivarea soiurilor precoce, lucrările de ameliorare pot utiliza genele de rezistenţă prezente la unele forme cultivate foarte bine adaptate cum sunt populaţiile locale din Moldova sau de la speciile înrudite, cum ar fi Phaseolus acutifolius şi Phaseolus aureus
Ameliorarea fasolei pentru boabe semiuscate Acesta este un obiectiv special conturat în unele centre de ameliorare din vestul Europei. Pentru conservarea boabelor în condiţii calitative mai bune, se pare că recoltarea trebuie realizată la umiditatea de 40%. Din acest motiv, pentru ca maşinile de recoltat să lucreze fără pierderi, boabele trebuie să fie foarte omogene ajunse toate la umiditatea de recoltare. Culoarea lor trebuie să fie verde şi să nu fie îmbătrânite. Un nou criteriu de selecţie impus de consumatori şi de către industrializatori este fineţea boabelor. . Până acum, legătura dintre vârstă şi calitate pe de o parte şi tinereţea şi mărimea boabelor pe de altă parte, nu au existat. Se impune urmărirea cu atenţie a omogenităţii mărimii boabelor şi a frăgezimii acestora, asemănător criteriilor de la mazăre. Deocamdată, fasolea pentru boabe verzi este mai puţin cunoscută la noi în ţară, dar cererea pe plan mondial este în creştere. Ca principale obiective în selecţie sunt: boabe mari şi sferice care să se desfacă uşor din păstaie.
Boabele verzi se impun faţă de cele uscate pentru că pot fi congelate şi obţinerea semipreparatelor este mai uşoară.
AMELIORAREA MAZĂREI
Mazărea, ca celelalte leguminoase, aduce în alimentaţie şi furajare un important aport proteic, vitamine şi săruri minerale. Mazărea de grădină este un produs care se poate consuma în stare proaspătă sau conservată. Boabele de mazăre constituie un aliment bogat în proteine şi glucide, aducând un aport energetic important. Din acest motiv sunt foarte indicate în alimentaţia copiilor şi a persoanelor care depun eforturi deosebite.
Utilizarea în furajare este posibilă fie sub formă de boabe, fie sub formă de masă verde, în combinaţie cu cereale păioase, rezultând borceaguri de toamnă sau de primăvară. În alimentaţia umană, mazărea se foloseşte mai mult sub formă de boabe verzi, şi foarte rar sub formă de boabe uscate sau păstăi verzi. Pentru că producţia proaspătă se obţine într-o perioadă de timp foarte restrânsă din an, cantităţile cele mai mari de boabe verzi sunt destinate conservării.
Deşi utilizările mazărei pot fi multiple, extinderea în cultură este restricţionată de unele neajunsuri ale soiurilor existente, neajunsuri care trebuie eliminate prin procesul de ameliorare. În primul rând se impune creşterea nivelului productiv al soiurilor, apoi, obţinerea unor modificări de nivel morfologic prin care să se uşureze recoltarea mecanizată şi fără pierderi.
Obiective de ameliorare.
Ameliorarea capacităţii de producţie. Capacitatea de producţie la mazăre este dependentă de mai multe elemente de ordin morfologic şi fiziologic. Elementul morfologic hotărâtor este numărul păstăilor fertile pe plantă. Acesta depinde de numărul nodurilor florifere şi de numărul florilor la un nod. Optimizarea acestor elemente, alături de optimizarea unor însuşiri fiziologice, pot soluţiona obţinerea unor producţii mai mari la această cultură.
În mod obişnuit, majoritatea nodurilor florifere sunt amplasate în jumătatea inferioară a tulpinii, ceea ce face ca unele dintre păstăi să se piardă sau să se deprecieze. Din acest punct de vedere, un obiectiv important este repartizarea nodurilor florifere pe toată lungimea tulpinii, sau gruparea acestora spre vârf.
Cu toate că numărul nodurilor florifere este destul de mare, nu toate fructifică la nivel maxim, aşa cum nici în păstaie nu se obţine numărul maxim posibil de boabe. Aceasta se datorează capacităţii asimilatorii deficitare care duce la imposibilitatea hrănirii tuturor păstăilor, multe rămânând sterile.
Din cele prezentate reiese că o altă componentă importantă a producţiei este numărul boabelor din păstaie. Nici acest element nu se poate realiza la nivelul potenţialului posibil. Deşi numărul punctelor de fructificare din păstaie este mare, unele rămân sterile din aceleaşi motive, imposibilitatea asigurării asimilatelor. În privinţa acestui caracter se tinde la valori de 8-10 boabe în păstaie, chiar şi peste 12.
O altă componentă a producţiei pe plantă este mărimea boabelor, însă aceasta nu este direct corelată cu capacitatea de producţie deoarece mărimea boabelor este corelată negativ cu numărul lor. Formele cu boabe mari realizează un număr restrâns de boabe în păstaie. Astfel, formele cu boabe puţine pot prezenta peste 300 g masa a 1000 boabe, iar cele cu boabe mai multe în păstaie nu depăşesc o masă a 1000 boabe de 200 g.
Având în vederea cele prezentate, principala însuşire ce trebuie îmbunătăţite este hrănirea normală a punctelor de fructificare prin sporirea capacităţii de asimilaţie şi modificarea suprafeţei foliare. O mare parte din asimilate se pierd pentru întreţinerea sistemului foliar.
Reducerea suprafeţei foliare poate duce la creşterea capacităţii de producţie şi chiar la îmbunătăţirea rezistenţei la cădere datorită transformării foliolelor în cârcei care duc la menţinerea tulpinilor în poziţie verticală. Pentru aceasta poate fi utilizată gena af care transformă foliolele în cârcei sau gena st, care reduc mărimea stipelelor. La tipul de plante “afila”, suprafaţa foliară se reduce cu până la 40%, iar productivitatea creşte cu până la 20%, comparativ cu formele cu foliaj normal. (COUSIN,
1992)Pe lângă aceste gene mai sunt folosite genele rogue care reduc lăţimea foliolelor, sau combinaţii
între genele “af” şi “ti” care produc cârcei transformaţi în foliole foarte reduse ca mărime. Genitorii pentru creşterea capacităţii de producţie trebuie să fie caracterizaţi prin cel puţin 10-12
păstăi pe plantă, cu minim 7-8 boabe în păstaie şi o masă a 1000 boabe în jur de 300 g. În ameliorarea capacităţii de producţie trebuie avută în vedere şi influenţa activităţii simbiozei cu
bacteriile fixatoare de azot. Formele cu foliaj normal mai productive prezintă şi o activitate superioară în fixarea azotului atmosferic, deci există o corelaţie directă. Se poate crede că energia folosită în întreţinerea sistemului foliar la plantele normale, la cele de tip “afila” poate fi dirijată spre simbioza fixatoare de azot. Studiile arată că la formele cu foliaj redus, activitatea fixatoare de azot este mai redusă, deşi productivitatea lor este ridicată. La aceste forme corelaţie respectivă nu mai este valabilă. (COUSIN şi colab., 1987)
Ameliorarea calităţii. Calitatea la mazăre trebuie privită diferit, în funcţie de destinaţia producţiei: pentru furajare sau în alimentaţie.
Importanţa boabelor de mazăre se datorează conţinutului în proteină care trebuie să depăşească 28-33% (valoarea amximă fiind de peste 35%). Un procentaj mai mare apare la formele cu bob zbârcit, comparativ cu cele cu bob neted. Variabilitatea existentă dă posibilitatea obţinerii unor forme superioare în această privinţă, dar trebuie avută în vedere corelaţia negativă dintre conţinutul în proteină şi capacitatea de producţie. Din acest motiv, este foarte greu a se obţine forme productive şi bogate în proteină.
Între mărimea boabelor şi conţinutul în proteină nu există o corelaţie directă. Principalii factori care influenţează bogăţia în proteină sunt condiţiile anului şi poziţia păstăilor pe plantă
Realizarea procentului maxim de proteină este dependentă şi de atacul bolilor, în special al făinării. În cazul unui atac puternic, procentul de proteină în bob scade foarte mult.
Raportul aminoacizilor esenţiali din proteina de mazăre este favorabil. Cu toate acestea creşterea conţinutul în metionină şi triptofan este binevenită, având în vedere faptul că aceşti aminoacizi sunt deficitari în majoritatea proteinelor de origine vegetală.
Opus procentului de proteină este conţinutul în amidon. Acesta este mai ridicat la formele cu boabe netede şi depinde de combinaţiile genelor “Ra” şi “Rb”, dominante sau recesive. Aflate în stare dominantă (Ra Ra Rb Rb), determină boabe netede, iar combinaţiile dintre stările dominante şi cele recesive duc la apariţia diferitelor grade de zbârcire.
Din punct de vedere alimentar, calitatea presupune îmbunătăţirea coeficientului de fierbere pentru a se obţine o fierbere rapidă şi uniformă. Această însuşire este corelată cu forma boabelor. Boabele cu formă rotundă au o capacitate de fierbere mai bună.
Tot din punct de vedere culinar, procentul de coji cât mai scăzut este un obiectiv important de realizat. Valoarea maximă a acestuia trebuie să fie sub 5%.
La soiurile destinate consumului sub formă de boabe verzi, foarte important este conţinutul în zahăr. Procentul de zaharuri simple trebuie să depăşească 7%, dar se cunosc şi forme cu peste 10% zahăr în boabe. Boabele trebuie să fie dulci şi fragede, să aibă formă rotundă regulată, culoare uniformă, iar coaja să fie moale dar să nu crape uşor. Gustul bun şi aroma plăcută trebuie să se păstreze chiar dacă recoltarea mai întârzie, Culoarea boabelor depinde de modul de prelucrare. Pentru mazărea refrigerată, boabele trebuie să aibă o culoare verde oliv intensă, iar pentru conserve în cutii, verde deschis. Procentul boabelor faţă de păstăi este de 30 - 54%. Pentru soiurile de mazăre pentru conserve, procentul boabelor mici trebuie să fie cât mai mare, iar culoarea verde să nu se altereze în timpul prelucrării. La aceste soiuri, coacerea boabelor trebuie să fie cât mai uniformă pentru a se recolta toate odată. O altă cerinţă este ca boabele destinate conservării să nu crape în timpul manipulării, iar în conserve să nu apară tulbureala datorată amilozei. Dacă mazărea se prelucrează imediat după recoltare, tulbureala nu se formează. Din acest motiv trebuie create soiuri speciale pentru conserve.
Soiurile de la care se consumă şi păstăile verzi nu trebuie să aibă aţe. Soiuri cu păstăi complet fără aţe nu există, dar există soiuri la care aţele sunt absente la păstăile foarte tinere.
Ameliorarea precocităţii. Sensibilitatea mazărei faţă de temperaturile ridicate impune crearea de soiuri precoce, în special pentru zona de sud-est a României. Soiurile timpurii prezintă inconvenientul
unor producţii scăzute, precocitatea fiind corelată direct cu o capacitate mică de producţie şi un procent de proteină scăzut. Această corelaţie nu poate fi desfăcută decât prin utilizarea transgresiunilor care pot apărea în cadrul populaţiilor hibride, bazate pe acumularea de gene favorabile dominante.
În privinţa precocităţii, soiurile cele mai convenabile sunt cele semitimpurii, cu o perioadă de vegetaţie de 60-90 zile, soiuri la care capacitatea de producţie este satisfăcătoare. Crearea lor trebuie să ţină cont de faptul că precocitatea este un caracter poligenic bazat pe aditivitatea genelor implicate.
Ameliorarea rezistenţei la temperaturi scăzute. Realizarea unor producţii mari depinde de factorii de mediu ai anului, dintre aceştia, un rol foarte important avându-l temperaturile scăzute. Mazărea fiind o plantă care se seamănă primăvara foarte timpuriu, suportă temperaturi destul de scăzute în primele faze de vegetaţie (în jurul a –50C).
Crearea unor forme cu rezistenţă mult mai mare este posibilă folosind gene preluate de la speciile înrudite, cea mai folosită fiind Pisum arvense. (SCURTU, 2001) Pe baza acestei specii se pot crea şi forme de toamnă care pornesc în vegetaţie, primăvara foarte devreme, acestea maturizându-se cu 10 zile mai devreme decât formele de primăvară. Asemenea soiuri pot realiza producţii foarte mari. (COUSIN, 1976)
Variabilitatea acestui caracter este pronunţată, în Asia Mică fiind cunoscute varietăţi ce pot rezista până la –220C. Soiurile străine, forme de toamnă, nu pot fi cultivate decât în zonele cu ierni blânde din Banat sau sud-vestul Olteniei.
Ameliorarea rezistenţei la secetă. Seceta este foarte păgubitoare în faza de formare a fructelor. În condiţii de secetă şi arşiţă, producţia este diminuată datorită umplerii deficitare a boabelor, sau chiar de avortarea unor puncte de fructificare. (MADOŞĂ, 2004)
Selecţia pentru acest caracter se poate face prin observarea sistemului radicular. Formele cu sistem radicular profund sunt mai rezistente, dar acestea prezintă boabe foarte mici, reflectate într-o producţie scăzută.
Îmbunătăţirea rezistenţei la secetă se poate îmbunătăţi prin încrucişare cu unele specii înrudite, cele mai bune fiind Pisum arvense şi Pisum fulvum.
Ameliorarea rezistenţei la boli şi dăunători. Bolile pot diminua producţia şi calitatea acesteia foarte mult. Pentru majoritatea bolilor nu se cunosc surse de rezistenţă ci numai unele soiuri sau specii tolerante.
Rezistenţa la antracnoză este controlată poligenic, majoritatea surselor fiind tolerante. În unele soiuri cultivate de origine olandeză (soiul Rondo fiind cel mai valoros) sunt introduse gene de la Pisum elatius. Acestea conferă rezistenţă la Ascochyta pisi. Dacă este vorba de Ascochyta pinodes, sursele de rezistenţă sunt necunoscute.
Fuzarioza este o altă boală foarte răspândită. Cele mai tolerante sunt soiurile americane, de la care genele de rezistenţă pot fi transferate în procesul de ameliorare. Aceste transfer se realizează destul de dificil datorită numărului mare de rase fiziologice ale agentului patogen, dar şi datorită faptului că sunt implicate un număr mare de gene, pentru unele rase, gene dominante, pentru altele gene recesive.
Rezistenţa faţă de atacul de Oidium sp. Este mai puţin cunoscută. Au fost identificate gene recesive în unele populaţii din Mexic şi Peru.
Atacul de Peronospora pisi poate provoca pagube importante în anii umezi şi reci. În unele soiuri străine au fost stabilite surse recesive de rezistenţă.
Virozele sunt foarte frecvente la mazăre, fiind provocate de viruşi diferiţi. Pentru acestea au fost stabilite surse importante de rezistenţă în germoplasma provenită din India, bazate pe gene recesive.
Bacteriozele (Pseudomonas sp.) sunt prezente în majoritatea anilor. Pentru diferitele rase ale acestui agent patogen se cunosc gene de rezistenţă, gene provenite din surse engleze sau franceze. Pe baza acestora se poate crea o toleranţă la atac.
Dintre dăunători, cel mai frecvent şi mai periculos este gărgăriţa mazărei. Sursele precise de rezistenţă nu sunt precizate. Se ştie că un atac mai redus apare când nu există concordanţă între înflorire şi perioada de zbor maxim a insectei. Ca urmare, formele mai precoce sunt mai rezistente, aceasta fiind în fond, o rezistenţă aparentă.
Ameliorarea aptitudinilor pentru recoltarea mecanizată. Recoltarea mazărei se realizează, de obicei, în două etape. Morfologia plantelor face ca în timpul recoltatului să apară un procent mare de
pierderi. Acestea pot fi eliminate prin crearea unor forme rezistente la cădere şi scuturare şi cu maturizare uniformă a păstăilor.
Rezistenţa slabă la cădere este cauzată de tulpinile înalte şi de structura anatomică deficitară a tulpinii. Deşi este necesară obţinerea unor forme cu talie mai redusă, o reducere a taliei prea mult face ca inserarea păstăilor să fie aproape de nivelul terenului, păstăile respective pierzându-se la recoltat. Ca urmare, formele cu talie pitică nu sunt de dorit. (MADOŞĂ, 2000)
Cele mai favorabile sunt formele cu talie mijlocie, tulpină elastică şi groasă şi o grupare a inflorescenţelor spre vârful tulpinii. Inserarea primelor păstăi spre baza tulpinii face ca la recoltat, acestea să nu fie tăiate de aparatul de tăiere al combinei. De asemenea, păstăile inserate spre vârful tulpinii nu ajung pe sol şi îmbolnăvirea lor se realizează mai greu.
Crearea unor forme cu tulpină elastică şi groasă, şi cu înflorire grupată la vârful tulpinii se poate obţine prin încrucişări cu forme cu tulpină fasciată. Folosirea acestora trebuie făcută cu atenţie deoarece, pe lângă aceste caractere favorabile, mai transmit si unele însuşiri negative: tardivitate şi producţie mică din cauza păstăilor şi boabelor mici.
Rezistenţa la cădere se poate îmbunătăţi şi prin crearea formelor cu frunze transformate în cârcei foarte ramificaţi, care duc la legarea plantelor unele de altele, formându-se o masă de plante care se susţin, astfel căderea fiind împiedicată. Aceste forme prezintă şi tulpini mai scurte şi o fructificare rapidă şi uniformă.
Pierderi la recoltat apar şi din cauza slabei rezistenţe la scuturare. Selecţia pentru îmbunătăţirea rezistenţei la scuturare se poate face prin observarea stratului pergamentos din păstaie cu care rezistenţe este direct corelată. Gene de rezistenţă pot fi preluate de la mazărea de grădină, formele bogate în zahăr fiind mai rezistente la scuturate decât formele obişnuite.
Cel de-al treilea aspect ce intervine în uşurarea recoltării mecanizate este uniformizarea maturizării. O coacere mai uniformă o prezintă tipul de plante cu flori grupate. Acest obiectiv poate fi îmbunătăţit folosind la hibridări tipul fasciat, uniformitatea maturizării fiind legată de înflorirea într-un interval de timp scurt.
Ameliorarea aptitudinilor pentru fixarea azotului. Aceste constituie un obiectiv special. Variabilitatea acestui caracter este foarte pronunţată. Se cunosc forme care nu prezintă nodozităţi, forme provenite din Asia (Pisum transcaucazicum) care nu prezintă nodozităţi în contact cu Rhisobium legunimosarum. Deşi se cunoaşte variabilitatea acestui caracter, nu se cunoaşte suficient determinismul genetic implicat în realizarea lui. Intensitatea activităţii de fixare a azotului nu depinde de precocitate, de sistemul foliar, sau de alte însuşiri. În unele cazuri se constată corelaţie între această activitate şi unele caractere, iar în alte situaţii corelaţiile nu există. Din acest motiv ameliorarea acestui caracter este foarte dificilă.
AMELIORAREA VIŢEI DE VIE
Plantaţiile cu viţă de vie au o mare extindere pe glob, în special în Europa. Cultura extinsă arată importanţa acordată acestei plante de către foarte multe popoare. Viţa de vie este cultivată pentru consumul fructelor proaspete, dar cel mai mult pentru obţinerea vinului.
Practica cultivării viţei de vie este o activitate foarte veche, la fel de vechi fiind şi preocupările faţă de selecţionarea unor forme mai productive şi superioare din punct de vedere calitativ. Această activitate desfăşurată de-a lungul secolelor a făcut ca la viţa de vie să fie cunoscute peste 10.000 de soiuri, din toate categoriile: pentru vin, struguri de masă sau pentru stafide. Multe dintre soiurile vechi se mai cultivă şi astăzi pe, suprafeţe însemnate. La aceasta contribuie atât calitatea acestor soiuri, cât şi durata de viaţă foarte lungă a plantaţiilor viticole.
Cele mai vechi soiuri au fost create prin selecţie în masă, dar mai târziu au fost aplicate şi selecţia clonală sau selecţia variaţiilor mugurale. Forte rar s-a practicat selecţia în populaţii obţinute din seminţe. În aceste cazuri variabilitatea este sporită de hibridările care se pot realiza. Din perioada străveche, în care ameliorarea s-a făcut la întâmplare, se mai cunosc încă multe soiuri şi populaţii locale. Acestea se găsesc,
nu numai în colecţii ci şi în cultură. În ultimul timp li se acordă atenţie tot mai mare, existând preocupări de depistare şi salvare de la pierdere. Vechile cultivare pot fi utilizate ca material iniţial în procesul de ameliorare. Pe lângă buna adaptare la condiţii specifice de mediu, nu poate fi omisă nici capacitatea de producţie şi calitatea acesteia.
La sfârşitul secolului al XVIII-lea au fost încercate primele hibridări dirijate. Aceste hibridări au fost încercate din cauza pericolului produs de apariţia filoxerei în Europa. Hibrizii între viţa americană şi cea europeană au salvat podgoriile europene, cu toate că din punct de vedere calitativ şi chiar productiv, nu egalează vechile soiuri. Aceşti hibrizi au stat la baza creării portaltoilor rezistenţi la filoxeră. Ulterior, selecţiile aplicate au dus la obţinerea unor hibrizi direct producători, rezistenţi la filoxeră, productivi şi cu o calitate apropiată de cea a soiurilor nobile.
În secolul al XX-lea hibridarea a rămas o metodă de bază, dar programele de ameliorare s-au desfăşurat în două direcţii speciale: de obţinere a unor soiuri roditoare folosite ca altoi şi de obţinere a unor soiuri portaltoi. Hibridarea a fost ajutată şi de alte metode, care s-au diversificat tot mai mult. Mutageneza şi poliploidia sunt deja metode binecunoscute. Dezvoltarea biotehnologiilor şi a diferitelor metode ale ingineriei genetice, deschid noi perspective în ameliorarea viţei de vie.
Strugurii sunt fructe foarte apreciate pentru calităţile lor. Conţinutul ridicat în zahăr şi aroma strugurilor de masă, sărurile minerale şi vitaminele pun strugurii pe acelaşi loc cu alte fructe. Importanţa soiurilor pentru vin nu mai trebuie menţionată. Vinurile sunt nelipsite zilnic pe masa multor popoare sau măcar cu ocazii speciale. Vinul constituie obiectul unor importante schimburi comerciale la nivel mondial.
Obiective de ameliorare.
Ameliorarea viţei de vie este o activitate foarte complexă. În condiţiile, tehnologiei şi cerinţelor actuale această activitate se desfăşoară în două direcţii: crearea de soiuri roditoare şi crearea de portaltoi corespunzători.
Ameliorarea productivităţii. Productivitatea este o însuşire complexă, condiţionată de baza ereditară a genotipului şi într-o anumită măsură şi de condiţiile pedo-climatice sau de agrotehnica aplicată. Zestrea genetică este hotărâtoare în realizarea caracterelor fructificării, iar agrotehnica şi condiţiile pedo-climatice reglează încărcătura de rodire şi parţial mărimea şi numărul fructificaţiilor.
Producţia la hectar depinde de densitatea plantelor şi de capacitatea de producţie a fiecărei plante.Densitatea plantelor depinde de vigoarea plantelor, dar cum la viţa de vie vigoarea este controlată
prin tăieri, densitatea depinde de sistemul de cultură adoptat. Sigur că în alegerea sistemului de susţinere şi de tăieri trebuie să se aibă în vedere particularităţile genotipului, deoarece sunt diferenţe de la un genotip la altul.
Viţa de vie creşte nedeterminat având şi o mare capacitate de lăstărire. Menţinerea unui anumit habitus al plantei se face efectuând numeroase lucrări de tăiere şi conducere, atât în perioada de repaus, cât şi în vegetaţie. În cadrul genului Vitis şi a genurilor înrudite, nu sunt cunoscute forme cu creştere determinată, dar sunt semnalate forme a căror vigoare este mai redusă. De exemplu, unele specii americane (V.aestivalis, V.candicans) au o vigoare redusă comparativ cu V.vinifera.
În funcţie de vigoarea butucului, pe o plantă poate fi prezent un număr mai mare sau mai mic de lăstari, deci o încărcătură de rod mai mare sau mai mică.
Vigoarea plantei şi capacitatea de lăstărire sunt caractere determinate poligenic. În general, vigoarea mare şi capacitatea puternică de lăstărire se transmit dominant.
Producţia pe butuc are ca şi componente morfologice, numărul de ciorchini, mărimea ciorchinilor şi mărimea boabelor. În funcţie de destinaţia producţiei, aceste elemente au comportare diferită. La soiurile pentru masă, ciorchinii trebuie să fie mari şi să prezinte boabe mari şi uniforme. La strugurii pentru vin este importantă mărimea ciorchinilor, dar mult mai important este conţinutul boabelor în must, mărimea boabelor nefiind relevantă.
Numărul ciorchinilor pe butuc se reglează prin tăieri, dependent de vigoarea butucilor şi de agrotehnica aplicată. La soiurile de struguri pentru masă se lasă o încărcătură de rod mai mică, pentru a avea siguranţa obţinerii unor ciorchini mari şi cu boabe mari. La soiurile de struguri pentru vin, numărul
ciorchinilor pe butuc va fi mult mai mare. Potenţialul este extrem de mare. La viţele conduse în gospodăriile populaţiei , pe un butuc pot fi realizaţi peste 100 de ciorchini. În plantaţiile de la care se aşteaptă producţii mari la hectar şi o bună calitate, numărul ciorchinilor care se realizează pe un butuc, este mult mai mic. Deoarece aceste caracter este controlat prin tăieri, nu poate fi tratat ca un caracter stabil, cu o anumită bază ereditară.
Mărimea ciorchinilor este un caracter de soi, determinat poligenic, având la baza realizării fenotipice, efecte de aditivitate. Cu toate acestea, condiţiile de mediu şi agrotehnica aplicată îl pot influenţa. În condiţii de secetă şi a unui număr mare de ciorchini pe butuc, mărimea ciorchinilor se va reduce. Odată cu reducerea mărimii ciorchinelui se reduce şi mărimea boabelor.
La soiurile se struguri pentru masă, ciorchinii mari sunt foarte apreciaţi. Ei trebuie să fie de tip răsfirat pentru ca boabele să ni se deformeze şi să aibă dezvoltare uniformă. La strugurii pentru vin sunt de dorit ciorchini mai mici şi de tip compact, chiar dacă şi boabele sunt mai mici. De fapt, între aceste caractere există corelaţii directe destul de strânse. Boabele mari se obţin numai în ciorchini mari şi răsfiraţi, iar în ciorchini de tip compact se realizează numai boabe mici. Uneori, din cauza unor condiţii nefavorabile (secetă, temperaturi ridicate) apare un anumit procent de boabe mici şi în ciorchinii strugurilor de masă.
Mărimea boabelor este un caracter cantitativ bazat pe efecte de aditivitate. Caracterul ciorchine dens, cu boabe mici se transmite mai fidel decât caracterul ciorchine mari şi boabe mari. Mărimea boabelor este foarte importantă la strugurii de masă şi s-a constatat că este corelată uneori cu tardivitatea coacerii boabelor.
Aceste componente morfologice ale productivităţii pot fi cuprinse într-o singură observaţie, şi anume producţie pe butuc. Evaluarea productivităţii butucului nu se face în primul an de rodire, ci după 2-3 ani. La soiurile de struguri pentru vin producţia unui butuc trebuie să fie de cel puţin 3 kg, iar soiurile de struguri pentru masă limita minimă a producţiei este de 5 kg.
La strugurii pentru vin , productivitatea plantei se poate evalua şi prin determinarea cantităţii de must realizată pe butuc. De regulă, se determină randamentul în must al ciorchinilor. Această însuşire este corelată cu epoca de coace e a boabelor. Cu cât coacerea este mai târzie, cu atât randamentul este mai bun. În selecţia soiurilor de struguri pentru vin se vor reţine ca valoroase formele la care din 100 kg struguri se obţin cel puţin 75 l must.
Ameliorarea calităţii. Calitatea recoltei are mai multe componente, în funcţie de utilizarea strugurilor.
La soiurile de struguri pentru vin cel mai important este conţinutul în zahăr şi acizi, însă la cei pentru masă importante sunt şi alte caractere de ordin morfologic. Realizarea unor vinuri de calitate, competitive pe plan mondial nu este posibilă fără cultivarea unor soiuri nobile, cu calitate superioară şi excluderea hibrizilor producători direcţi. (TĂNASE, 2002)
Procentul de zahăr şi aciditatea sunt două caracteristici care contribuie împreună la realizarea calităţii, cu toate că transmiterea lor ereditară este separată. Foarte important este raportul între cele două grupe de substanţe şi nu valorile acestora. Valorile procentuale sunt mult influenţate de condiţiile de mediu (umiditate, temperaturi, lumină). Raportul între ele este un caracter mai stabil. La strugurii pentru vin, zahărul contribuie la realizarea unei anumite tării, iar aciditatea (alături de alţi compuşi) realizează gustul şi buchetul.
La strugurii pentru masă, cele două grupe de substanţe dau gustul şi aroma. Principalele fracţii glucidice care se găsesc sunt glucoza şi fructoza. La strugurii pentru vin este de dorit o proporţie majoritară a fructozei, pe când la cei pentru masă, a glucozei.
Culoarea fructelor este un caracter cu ereditate puternică şi foarte stabil. În realizare culorii boabelor sunt implicate două perechi de gene, în funcţie de contribuţia cărora apar cele trei nuanţe de culoare: negru, roşu sau alb. Gena dominantă “B” determină culoarea neagră, iar gena dominantă “R” culoarea roşie. Formele homozigot recesive vor avea boabe de culoare albă, iar cele în care sunt prezente ambele gene dominante, vor avea nuanţe foarte închise.
Forma boabelor poate prezenta importanţă la strugurii pentru masă, influenţând aspectul comercial. Caracterul este determinat poligenic şi s-a constatat că formele rotunjite domină asupra celor alungite. Unele segmente de piaţă apreciază formele curioase, cum sunt cele turtite sau foarte alungite,
sub formă de ţurţure. Între forma bobului, gust şi aromă nu există corelaţie. Aroma este o componentă foarte importantă atât la soiurile pentru masă cât şi la cele pentru vin.
Realizarea aromei este o rezultantă a participării multor substanţe: raportul glucide-aciditate, proporţia glucozei şi a fructozei, prezenţa esterilor şi a aldehidelor, natura acestora, prezenţa metanolului şi a uleiurilor volatile sau a unor substanţe astringente. Foarte dăunători sunt unii compuşi preluaţi de la speciile americane care se transmit dominant şi care conferă boabelor gust ştiros.
La strugurii de masă cea mai cunoscută aromă este cea de muscat (sau tămâioasă). Aceasta se transmite dominant şi poate fi uşor realizat prin hibridări.
Pe lângă analizele biochimice de determinarea a procentului de zahăr şi a acidităţii, strugurii mai pot fi bonitaţi prin observaţii în privinţa calităţii. La strugurii pentru vin se pot face degustări şi observaţii stabilind gustul şi aroma, culoarea şi limpezimea mustului. La strugurii pentru masă se bonitează gustul, aroma, consistenţa pulpei, fineţea pieliţei şi aderenţa acesteia de pulpă, mărimea seminţelor. În plus, la strugurii pentru masă pot fi efectuate determinări referitoare şi la conţinutul în vitamine, săruri minerale, aspecte puţin avute în vedere în aplicarea selecţiei.
Ameliorarea epocii de coacere. Coacerea strugurilor se poate realiza în diferite epoci, putând fi eşalonată pe o perioadă de 3 luni. Între epoca de coacere a strugurilor şi calitate este o strânsă corelaţie. Soiurile tardive au calitate superioară. Cu toate acestea , în special la soiurile de struguri pentru masă, sunt necesare şi soiuri timpurii. La strugurii pentru masă este necesară existenţa unei game largi de soiuri pentru a se asigura piaţa cu fructe proaspete, mai ales că păstrarea strugurilor din soiurile mai timpurii nu se poate face pe o perioadă prea lungă de timp. În zonele în care toamnele sunt mai scurte şi răcoroase, cultivarea soiurilor tardive este riscantă pentru că nu se acumulează zahăr suficient şi calitatea recoltei va fi mediocră.
La soiurile pentru vin, primordială este calitatea şi nivelul producţiei, acestea fiind net superioare la formele tardive. Pentru regiunile în care iarna se instalează mai devreme sunt necesare şi soiuri semiprecoce sau semitardive, pentru a avea siguranţa coacerii până la recoltat.
Precocitatea sau tardivitatea sunt însuşiri poligenice destul de stabile. Totuşi, influenţa condiţiilor de climă se face simţită. Temperaturile din perioada de vegetaţie pot influenţa maturizarea boabelor. În anii foarte calzi şi secetoşi, coacerea este mai timpurie decât în anii reci. Din acest motiv, pe parcursul perioadei de vegetaţie trebuie observate toate fazele de creştere şi dezvoltare, şi făcută o corelare a acestor observaţii cu condiţiile de climă.
Selecţionarea acestei însuşiri nu se poate realiza decât după intrarea pe rod. La soiurile speciei V.vinifera au semnalate unele corelaţii care pot uşura selecţia în privinţa epocii de coacere. Astfel, între coacerea strugurilor şi durata perioadei de cădere a frunzelor există o corelaţie directă. Cu cât frunzele cad într-un interval de timp mai scurt, cu atât coacerea fructelor va fi mai precoce. Alt caracter care poate fi observat este coacerea lemnului. Epoca de coacere a lemnului coincide cu intrarea în pârgă a strugurilor. Ca urmare, observând aceste caractere la plantele din primii ani de viaţă (înainte de rodire) se poate aprecia epoca de maturizare a viitoarei clone. (POPESCU, 1966)
La puieţii obţinuţi din seminţe precocitatea se păstrează neschimbată, ca şi la genotipul din care provin. În hibridări, precocitatea este transmisă cel mai fidel de către soiurile precoce cu origine nordică.
Ameliorarea rezistenţei la ger şi iernare. Rezistenţa la iernare este un caracter foarte important la viţa de vie, de el depinzând stabilitatea recoltelor. Nivelul rezistenţei depinde de foarte mulţi factori, începând cu coacerea lemnului, continuând cu condiţiile iernii şi cu nivelul temperaturilor scăzute din primăvară.
Viţa de vie nu este o specie foarte rezistentă la ger. Unele forme sunt chiar destul de sensibile. În general, soiurile europene cele mai rezistente suportă geruri de scurtă durată de până la –200C. Speciile sălbatice suportă şi geruri mai aspre.
Rezistenţa la gerurile din timpul iernii a devenit o însuşire foarte importantă după elaborarea tehnologiilor moderne cu conducere înaltă, în care îngroparea butucilor nu se mai practică. În aceste condiţii, se impune o bună rezistenţă a altoiului cât şi a portaltoiului.
Din studiile realizate s-a dedus că rezistenţa la ger este un caracter determinat de un număr redus de gene minore. În creşterea rezistenţei la ger un rol important l-a avut mutaţiile naturale, deoarece încrucişările naturale ale viţelor europene cu specii mai rezistente nu au fost posibile, deoarece speciile
americane sau asiatice au pătruns în Europa destul de târziu.Principalii genitori pentru rezistenţa la ger sunt speciile americane (V.riparia şi V.rupestris), sau
cele asiatice (V.amuriensis şi V.davidii). cele mai rezistente sunt speciile asiatice. Utilizarea acestor specii în ameliorarea soiurilor nobile trebuie făcută cu atenţie deoarece de la acestea se transmit şi multe caractere nevaloroase. Principalul defect preluat de la speciile americane este gustul astringent al strugurilor. Foarte valoroase sunt aceste specii în ameliorarea portaltoilor. Dacă portaltoiul este foarte rezistent asigură protecţia butucului împotriva acţiunii gerului la nivelul rădăcinilor, dar poate induce un anumit nivel al rezistenţei şi la soiul folosit ca altoi.
În condiţiile României, crearea unor soiuri foarte rezistente este necesară în aproape toate zonele ţării, chiar dacă frecvenţa iernilor mai aspre este mai mare numai în anumite regiuni.
Ameliorarea coacerii lemnului. Coacerea lemnului are influenţă asupra rezistenţei la ger a plantei, asupra prinderii la altoire, asupra intrării pe rod şi chiar a maturizării fructelor.
Prin termenul de coacere a lemnului se înţelege acumularea unei anumite cantităţi de hidraţi de carbon la nivelul coardelor. Hidraţii de carbon produc o creştere a concentraţiei sucului celular din coarde, astfel creându-se o mai bună rezistenţă la îngheţ. Dacă nu se poate determina concentraţia de hidraţi de carbon din coarde, coacerea lemnului poate fi apreciată în funcţie de culoarea acestuia. Se consideră că lemnul este copt când coardele îşi schimbă culoarea, din verde în gălbui sau brun roşcat.
Gradul de coacere a lemnului diferă de la un soi la altul şi de la o specie la alta, asemănător variaţiilor de maturizare a strugurilor.
Coacerea lemnului este la fel de importantă şi în cazul portaltoilor. Cea mai bună coacere a lemnului o au portaltoii Riparia Gloire
Ereditatea coacerii lemnului este destul de puţin cunoscută. S-a observat că ereditatea acestei însuşiri este foarte scăzută. Determinismul genetic nu este elucidat, dar se presupune că este poligenic, aflat foarte mult sub influenţa condiţiilor de mediu. De la un an la altul apar fluctuaţii mari în privinţa hidraţilor de carbon acumulaţi în coarde.
Ameliorarea rezistenţei la secetă. Viţa de vie nu este foarte pretenţioasă faţă de umiditate, dar pentru zonele mai secetoase sunt necesare şi soiuri rezistente. În unele zone din ţară există soiuri locale cu un nivel bun al rezistenţei la secetă. Acestea s-au format pe soluri nisipoase (în sudul Olteniei) sau pe soluri cu roca mamă la suprafaţă care nu reţin apa. Astfel de soiuri vechi pot fi utilizate ca genitori.
În condiţiile în care se cultivă viţe altoite, rezistenţa la secetă trebuie să fie foarte bună pentru portaltoii folosiţi.
Rezistenţa la secetă trebuie îmbunătăţită concomitent cu rezistenţa la temperaturi ridicate. De regulă, aceşti doi factori de stres acţionează împreună. Soiurile folosite ca altoi trebuie să-şi păstreze aparatul foliar funcţional în condiţii de deficit hidric şi arşiţă.
Cele mai valoroase surse de rezistenţă la secetă sunt unele specii americane: V.rotundifolia, V.aestivalis, V.lincecunii.
Dintre soiurile cultivate, multe pot fi utilizate ca genitori alături de unii hibrizi producători direcţi. Atenţie mare trebuie acordată alegrii genitorilor, deoarece multe soiuri sunt foarte sensibile.
Ameliorarea rezistenţei la boli şi dăunători. Viţa de vie este atacată de mai mulţi agenţi patogeni, dar cel mai păgubitor este Plasmopara viticola, care produce mana viţei de vie. Atacul de mană este bine corelat cu condiţiile de climă, în special cu umiditatea şi temperatura. În condiţii de umiditate ridicată, la temperaturi de cel puţin 110C, atacul se poate realiza. Cu cât temperatura va creşte, atacul va fi mai rapid, temperatura optimă fiind în jur de 200C.
În cadrul soiurilor cultivate există o variabilitate a rezistenţei la mană, unele soiuri prezentând o rezistenţă acceptabilă (Băbească, Cabernet, Negru vârtos, Traminer). Sursele de rezistenţă cele mai valoroase se găsesc printre speciile americane: V.riparia, V.berlandierii, V.rotundifolia şi V.cinerea.
În funcţie de sursă, determinismul genetic al rezistenţei la mană poate fi monogenic sau poligenic. Determinismul monogenic duce la o necrozare rapidă a ţesuturilor în locul unde s-a realizat atacul. Sistemul poligenic al rezistenţei are acţiune asupra micelului ciupercii, inhibându-i dezvoltarea (VICTORIA LEPĂDATU, 1970 citat de ARDELEAN; 1986)
Testarea rezistenţei la mană trebuie efectuată de timpuriu, chiar plantele din primul an de viaţă pot fi supuse infecţiilor artificiale, testările făcându-se în seră.
Alte boli faţă de care sursele de rezistenţă sunt mai puţin cunoscute, sunt micoplasmozele. Cele mai frecvente în ţara noastră sunt îngălbenirea aurie şi înnegrirea lemnului. Nivelul sensibilităţii diferă de la un soi la altul, astfel că un studiu riguros al posibililor genitori poate duce la depistarea unor surse de toleranţă. (TOMOIOAGĂ LILIANA, 2002)
Cel mai periculos dăunător este filoxera. Strategiile de luptă împotriva filoxerei s-au schimbat de-a lungul timpului. Principala şi prima cale de a combate atacul de filoxeră a fost altoirea pe portaltoi rezistenţi, deoarece atacul la rădăcină este cel mai periculos, neputând fi combătut pe cale chimică. Cea de a doua cale încercată a fost cultivarea unor hibrizi producători direcţi.
Pentru a cultiva soiurile nobile care dau vinuri de calitate superioară, singura posibilitate este altoirea acestora pe portaltoi valoroşi. Hibrizii cultivaţi pe rădăcini proprii dau vinuri de calitate inferioară. Aceştia au fost obţinuţi prin încrucişări cu specii americane de la care au moştenit şi o serie de caractere nevaloroase.
Portaltoii de viţă de vie rezistenţi la atacul de filoxeră se creează tot cu ajutorul speciilor americane, acestea fiind singurele care prezintă gene de rezistenţă. Aceste specii nu se folosesc direct ca portaltoi. Cei mai valoroşi portaltoi se obţin prin încrucişări ale acestor specii cu V.vinifera sau încrucişându-le între ele. Pentru rezistenţa la filoxeră, dintre speciile americane, cele mai valoroase sunt V.riparia, V.rupestris, V.berlandieri, V.cinerea. la noi în ţară, unele dintre aceste specii au fost deja folosite în obţinerea binecunoscuţilor portaltoi din grupa Crăciunel.
Crearea unor soiuri nobile rezistente la filoxeră, care să fie cultivate pe rădăcini proprii rămâne încă o problemă de viitor. Transferul genelor de la speciile americane este posibil mai cu seamă că diferitele metode ale ingineriei genetice permit acest lucru, fiind excluse dezavantajele metodelor clasice (a hibridării).
Orice cale este urmată, ameliorarea rezistenţei la filoxeră rămâne un obiectiv permanent datorită faptului că parazitul evoluează continuu, noile rase nefiind contracarate de fondul de gene utilizat. Speciile americane din genul Vitis, chiar şi alte genuri, trebuie studiate în continuare pentru a găsi noi surse de rezistenţă.
Din numeroase studii efectuate s-a constatat că rezistenţa la atacul de filoxeră prezintă un determinism poligenic. Chiar speciile americane, cunoscute ca rezistente, prezintă grade diferite de rezistenţă. Complexitatea însuşirii de rezistenţă şi diversitatea parazitului, face ca în ameliorare alegerea materialului cu care se lucrează este hotărâtoare. Când se efectuează încrucişări, trebuie să se lucreze cu cei mai valoroşi genitori. La hibridarea speciei V.vinifera cu cele americane, se va lucra cu soiurile care prezintă toleranţă faţă de filoxeră: Cabernet Sauvignon, Băbească neagră, Zghihară.
Selecţia formelor rezistente se face numai în condiţii de infecţie artificială, existând metode de testare bine puse la punct.
Obiective speciale de ameliorare. În ameliorarea viţei de vie apar şi o serie de obiective speciale: adaptarea la condiţii speciale de climă şi sol, obţinerea unor soiuri cu struguri fără seminţe, obţinerea unor soiuri cu flori hermafrodite, aptitudini pentru recoltarea mecanizată.
Cu toate că viţa de vie este o specie cu adaptabilitate foarte bună, există situaţii când sunt necesare forme şi mai rezistente, în cazul înfiinţării podgoriilor pe terenuri aparte (nisipuri, soluri pietroase, soluri bogate în calcar) sau în zone cu climă deosebită (secetoasă, foarte umedă). Astfel de soiuri rezistente, pot valorifica terenuri improprii pentru alte culturi. Îmbunătăţirea însuşirilor de rezistenţă a factorii de stres este posibilă în cadrul genului Vitis fiind cunoscute specii valoroase pentru asemenea caractere, atât printre cele de origine americană, cât şi asiatică. Nu trebuie uitate nici vechile forme din specia V.vinifera, în special cele din zona mediteraneană (europeană, asiatică şi chiar africană).
Strugurii fără seminţe au utilizări speciale. La strugurii de masă sunt apreciate soiurile cu seminţe puţine şi mici sau chiar cei fără seminţe. La soiurile pentru stafide, lipsa seminţelor este obligatorie.
Lipsa seminţelor este un caracter monogenic recesiv, însă expresivitatea genei poate fi modificată de alţi factori ereditari. Soiurile de struguri pentru masă fără seminţe se obţin prin hibridări între genitori tetraploizi şi diploizi, deci vor fi forme triploide. Principalii genitori care se utilizează sunt soiurile cu seminţe puţine sau chiar fără seminţe. Alături de vechile soiuri Kiş-Miş şi Corinth, pot fi folosite şi soiurile americane, care sunt mai recente. (GRECU, 2001)
La soiurile pentru stafide, pe lângă lipsa seminţelor, boabele trebuie să mai prezinte şi alte
însuşiri: procent ridicat de substanţă uscată, pulpa moale şi aroma puternică.Crearea soiurilor cu flori hermafrodite este importantă pentru înfiinţarea plantaţiilor pure şi pentru
stabilitatea producţiei de la un an la altul. Din studiile efectuate s-a constatat că florile hermafrodite prezintă determinism monogenic dominant, iar florile unisexuate sunt determinate monogenic recesiv. Totuşi, aceste gene se pot găsi sub acţiunea unor gene modificatoare. La această concluzie s-a ajuns de la constatarea că în urma hibridării unor soiuri cu flori hermafrodite cu soiuri unisexuate, se obţin populaţii în care raporturile de segregare depind de genitor. Unele soiuri (cum sunt Muscatul de Hamburg, Reisling Italian, Chasselas d’ore) transmit foarte bine urmaşilor caracterul floare hermafrodită.
Recoltarea manuală a strugurilor este o lucrare costisitoare. Pentru ca această lucrare să se poată realiza mecanizat, soiurile trebuie să prezinte anumite însuşiri: coacerea să fie foarte uniformă, sistemul radicular să fie foarte bine dezvoltat pentru ca butucul să fie puternic ancorat în sol, frunzele să cadă odată cu ajungerea la maturitate a strugurilor şi într-un interval de timp foarte scurt, iar boabele să fie prinse pe ciorchine mai slab (dar să depăşească limita sensibilităţii la scuturare). În sortimentul mondial există soiuri care îndeplinesc astfel de cerinţe, ele putând fi folosite ca genitori.
AMELIORAREA POMILOR FRUCTIFERIMărul este o specie rustică, care are o mare capacitate de adaptare, din această cauză fiind extins în cultură în diverse zone ecologice, până în zone montane cu ierni destul de aspre. În lume, cultura este extinsă între 250 şi 600 latitudine nordică, iar în sud pe toate continentele, până în cele mai sudice regiuni.
Merele au diverse utilizări, putând fi consumate în stare proaspătă sau prelucrate sub formă de sucuri, compot, gemuri şi chiar alcool. După citrice, producţia de mere se situează pe locul doi în lume în cadrul producţiei de fructe. Această situaţie se datorează faptului că producţiile la unitatea de suprafaţă au crescut continuu, sunt stabile, dar şi pentru că merele sunt fructe valoroase prin compoziţia biochimică pe care o au. Pe lângă glucide, merele sunt bogate în vitamine ( vitaminele C, PP, B 1, B2, B6) şi săruri minerale.
Multiplele întrebuinţări ale mărului au făcut ca această specie să fie luată în studiu încă de la dezvoltarea ştiinţelor biologice şi horticole. De la cultivarea mărului, care s-a produs în Asia şi Europa, această specie s-a extins şi pe celelalte continente. Vechimea cultivării mărului este dovedită de descoperirile arheologice, în locuinţele oamenilor primitivi fiind găsite mere carbonizate. Fructele găsite au prezentă o variabilitate mare, pe lângă mere mici au fost găsite şi fructe mari ceea ce denotă existenţa unei variabilităţi încă din cele mai vechi timpuri, variabilitate în crearea căreia a fost implicată şi activitatea umană.
O contribuţie foarte importantă la îmbogăţirea şi perfecţionarea sortimentului de soiuri la măr au avut-o lucrările de ameliorare dirijată efectuate în vestul Europei începând cu sfârşitul secolului al XVIII-lea. În a doua jumătate a secolului al XIX-lea asemenea lucrări au luat amploare şi în estul Europei şi pe continentul American. Urmare a eforturilor de ameliorare, unii autori consideră că în lume, în perioada actuală există câteva mii de soiuri şi populaţii locale.
În ultimele decenii cercetările au fost concentrate spre perfecţionarea tehnologiei şi mai puţin spre diversificarea materialului biologic cultivat. Livezile moderne cuprind un număr mare de pomi la hectar, până la 2500 pomi. Pentru aceasta altoirea se face pe portaltoi speciali care conferă o reducere a vigorii concomitent cu utilizarea unor altoi cu talie specifică şi particularităţi aparte în ceea ce priveşte creşterea şi fructificarea. Prin crearea unor astfel de modificări de ordin morfologic se urmăreşte şi reducerea costurilor provocate de lucrările manuale, dar datorate combaterii chimice a bolilor şi dăunătorilor.
În România cultivarea mărului este o activitate foarte veche, dar lucrări de ameliorare fundamentate ştiinţific nu s-au desfăşurat decât după al doilea război mondial. Din perioada ameliorării empirice se mai păstrează încă unele populaţii locale sau soiuri foarte vechi, atât în colecţii cât şi în cultură în zonele premontane.
Ultimele direcţii ale cercetării în domeniul culturii mărului sunt legate de studiul poluării şi implicaţiile acesteia în activitatea fiziologică a plantelor, dar şi măsura în care substanţele chimice folosite în aplicarea tratamentelor se regăsesc în fruct. Prin ameliorarea însuşirilor de rezistenţă problema
poluării chimice poate fi rezolvată. (ROŞCA şi colab., 2003)
Obiective de ameliorare.
Obiectivele ameliorării la soiurile folosite ca altoi sunt direcţionate spre rezistenţa la boli criptogamice, la bacterioze şi la dăunători, reducerea lucrărilor manuale în vegetaţie şi a însuşirilor organoleptice ale fructelor. Sigur ca noile soiuri care prezintă astfel de însuşiri trebuie să posede şi caractere agronomice, cum ar fi productivitatea mare a pomului, aspect atractiv şi formă regulată a fructului.
Ameliorarea capacităţii de producţie. Productivitatea la unitatea de suprafaţă este dată de numărul pomilor la hectar şi de producţia medie a unui pom. Numărul pomilor la hectar depinde de vigoarea acestora. Pentru a mări numărul pomilor, vigoarea trebuie redusă prin crearea unor forme speciale sau prin sistemul de tăieri aplicat.
Studiul variabilităţii în cadrul speciei cultivate arată o variaţie continuă a caracterelor implicate în definirea portului pomului şi a tipului de fructificare. Se consideră patru ideotipuri arhitecturale.
Ideotipul coloană sau fus este constituit dintr-o axă garnisită cu ramificaţii scurte pe toată lungimea ei. Această arhitectură este corelată destul de strâns cu o alternanţă la rodire. Acest tip de creştere este datorat unei alele dominante depistată dintr-o populaţie mutantă a soiului McIntosh (LAPINS şi WATKINS, 1973). Se fac eforturi de asociere a rezistenţei la boli şi a stabilităţii producţiei, în special în ţările anglo-saxone, unde acest ideotip este mai răspândit.
Ideotipul “spur” este caracterizat prin formarea de ramificaţii scurte repartizate pe toate şarpantele. Modul de creştere este de tip va, portul înălţat, putând fi condus după dorinţă. Pomii au tendinţă spre alternanţă la rodire.
Ideotipul piramidal este foarte frecvent. Ramificaţiile sunt descrescânde spre vârf şi sunt dispuse de-a lungul unei tije tronconice. Fructificarea are loc pe lemn de la 2 la 4 ani.
Ideotipul natural prezintă o uşoară dominanţă a tijei principale care favorizează dezvoltarea unor ramificaţii laterale. Fructificarea are loc pe ramuri coronariene şi nu se manifestă alternanţa de rodire. Pomii pot fi conduşi de la sol fără susţinere.
Alegerea ideotipurilor depinde de tehnologia care se aplică. Sunt preferate formele “spur” şi cele care fructifică pe ramificaţii scurte pentru crearea livezilor intensive în care să se creeze densităţi mari la hectar. În direcţia modificării arhitecturii pomilor trebuie făcute eforturi pentru a se renunţa la lucrările manuale, prin simplificarea conducerii pomilor.
Producţia pe pom este dependentă de ideotipul cu care se lucrează. În medie, pe un pom se obţin 60-80 kg fructe, dar în condiţiile speciale, producţia unui pom poate depăşi 700 gk. (NEAGU, 1967)
Producţia pe pom este dependentă de numărul fructelor pe pom şi de greutatea lor. Aceste caractere sunt legate de ideotipul cu care se lucrează. În perioada actuală, când tipul de înfiinţare al livezilor este cel intensiv, ideotipurile cu care se lucrează sunt cele cu vigoare redusă pentru a putea creşte numărul pomilor la unitatea de suprafaţă. În aceste condiţii limitele maxime ale valorilor pentru cele două caractere implicate în realizarea producţiei pe pom nu vor fi cele maxime.
Numărul fructelor pe pom este un caracter complex, dependent de vigoarea pomului, vârstă şi condiţii de cultură. De asemenea, acest caracter este corelat direct cu mărimea fructelor. Având în vedere această situaţie nu se precizează un număr precis de fructe pe pom. Astfel numărul fructelor pe pomii selecţionaţi trebuie să fie între 30 şi 100 fructe de mărime corespunzătoare în primul an de rodire normală. Elitele care prezintă o asemenea încărcătură de fructe vor fi foarte productive după 5-6 ani de rodire, când începe rodirea la potenţial maxim. Având în vedere faptul că în primul an de rodire potenţialul plantei este foarte redus, se recomandă ca selecţia să fie efectuată după observaţiile din anul doi de rodire. În acest an, atât numărul fructelor este mai mare cât şi mărimea lor.
Mărimea fructului poate fi exprimată prin dimensiunile acestuia şi prin greutatea lui. În primii ani de fructificare, fructele sunt mai mici, pomul având o vigoare redusă, mărimea fructelor este foarte mult influenţată de numărul lor. Mărimea fructelor are o serie de implicaţii practice. Din acest motiv se recomandă ca fructele să nu fie exagerat de mari pentru că ramurile se pot rupe, cad uşor şi se deteriorează mai repede în timpul transportului. Fructele sub 200 g ar fi cele mai dorite conform acestor
principii. Totuşi, în ultimul timp sunt comercializate şi apreciate şi fructele mai mari, greutatea lor putând fi de până la 500 g.
Mărimea fructelor este un caracter cu determinism genetic forte complex şi destul de instabil. Chiar dacă dimensiunile şi greutatea fructului sunt dependente de genotip, în cadrul aceluiaşi genotip, cu cât fructele pe pom sunt mai multe, cu atât vor fi mai mici. Pe măsură ce vârsta pomilor creşte, această corelaţie se diminuează.
Transmiterea la urmaşi a mărimii fructelor este diferită de la un genitor la altul. În cele mai multe cazuri, mărime fructelor la hibrizi este intermediară formelor prentale. Uneori se întâlnesc şi descendenţe hibride cu puternic heterozis, la care fructele depăşesc în mărime ambii părinţi. Datorită variabilităţii materialului hibrid, în procesul de ameliorare a mărului o parte din hibrizi trebuie eliminaţi devreme pe criterii productive. Aceste eliminări se efectuează pe baza unor corelaţii între unele caractere ale pomilor tineri şi productivitatea pomilor maturi. Cel mai sigur indiciu este recolta din anul doi de rodire. Hibrizii care produc fructe multe la a doua rodire sunt consideraţi valoroşi, aceasta indicând faptul că viitorul pom va intra pe rod mai timpuriu şi va fi cu o bună productivitate.
Pentru a se obţine fructe mari, important este şi numărul fructelor în inflorescenţă. Este de dorit ca într-o inflorescenţă să rămână un singur fruct. Pentru a elimina rărirea chimică a fructelor, acest caracter poate fi selecţionat, autorărirea fiind observată cu o frecvenţă mai mare la unele soiuri. Prin aceasta, pe pom se obţin fructe uniform dezvoltate. (BUDAN şi BRANIŞTE, 2001)
Pentru ca producţiile să nu fie influenţate de polenizarea deficitară în livezile de producţie, în ultimul timp s-a constatat ca obiectiv de ameliorare aparte, obţinerea soiurilor autocompatibile. Prin aceasta se pot înfiinţa livezi pure. Rezultatele se lasă aşteptate, dar la nivel mondial există deja selecţii parţial autcompatibile şi cu o bună rezistenţă la rapăn.
Eficacitatea selecţiei pentru capacitatea de producţie, dar şi efectul economic la înfiinţării unei plantaţii este influenţată de durata perioadei juvenile. Intrarea puieţilor pe rod are implicaţii asupra duratei procesului de ameliorare. Imprimarea unei precocităţi din acest punct de vedere se poate face apelând la speciile înrudite, în special la Malus baccata sai M.prunifolia. Intrarea pe rod a puieţilor obţinuţi din seminţe este corelată direct cu intrarea pe rod a pomilor altoiţi. Astfel, selecţia hibrizilor trebuie să aibă în vedere, pe lângă productivitatea hibrizilor şi durat perioadei juvenile. Durata perioadei juvenile depinde de soi. Sunt soiuri foarte precoce, care intră pe rod în anul 3-4, şi foarte tardive, la care intrarea pe rod are loc după 10-12 ani. Intrarea pe rod a puieţilor este dependentă de vigoarea pomilor. Genotipurile care au o capacitate de creştere foarte rapidă vor atinge dimensiunile optime pentru rodire mult mai devreme decât cela cu creştere lentă. Aceasta se poate observa încă din primii ani de viaţă ai puieţilor. Ca urmare, puieţii foarte slab dezvoltaţi trebuie eliminaţi încă din anii 2-3 de viaţă.
Pe parcursul procesului de selecţie trebuie efectuate o multitudine de observaţii. Cele mai importante observaţii în vegetaţie se efectuează în primii ani de fructificare. Pomul este notat în privinţa vigorii şi a siluetei în funcţie de circumferinţa trunchiului, a numărului de dezvoltării ramificaţiilor. Sunt cuantificate observaţiile privind atacul bolilor, luându-se în considerare şi atacul pe fructe. Tot în această perioadă are loc bonitarea fructelor în funcţie de mărime, culoare, importanţa şi repartizarea culorii dominante, forma şi deschiderea tubului calicial, textura şi suculenţa pulpei, savoare, aspectul epidermei şi defectele acre pot apărea la fructe. Pe baza acestor observaţii se pot stabili unele corelaţii între caractere pentru a simplifica lucrările ulterioare de selecţie. De asemenea, pot fi calculate eritabilitatea caracterelor şi unii parametrii genetici.
Ameliorarea calităţii. În privinţa calităţii fructelor, la nivel mondial apare o tendinţă de specializare a soiurilor, pentru fructe destinate consumului proaspăt sau pentru fabricarea sucurilor şi concentratelor.
Calitatea fructelor este un caracter complex fiind compusă dintr-o serie de caractere organoleptice şi biochimice. Din punct de vedere organoleptic, principalele caractere sunt forma fructului, culoarea fructului şi a pulpei, gustul şi aroma. Biochimic, principalele componente ale calităţii sunt conţinutul în glucide, acizi şi vitamine.
Forma fructului poate fi foarte variabilă. Ereditatea acestui caracter este destul de puţin cunoscută. Se consideră că forma fructului este un caracter poligenic în care principalul rol îl au efectele aditive. Forma fructului este determinată pe baza raportului între diametrul fructului şi înălţimea acestuia.
Cunoscând valorile acestora la formele parentale, se poate estima valoarea care va apărea la hibrizi. Faţă de forma fructelor nu există preferinţe. Formele deosebite sunt chiar atrăgătoare pentru consumatori. Important este ca fructele să fie simetrice, fără asperităţi şi proeminenţe. Cele mai răspândite forme sunt cele rotunde, puţin aplatizate sau tronconice.
Culoarea fructelor este o componentă a calităţii pentru care nu există linii clar urmărite în ameliorare. Există variaţii de culoare: roşu, galben sau verde, unicolore sau cu combinaţii de culori. Culoarea fructului este dată de combinarea unei culori de fond cu o culoare de acoperire, care nu este obligatorie.
Culoarea de fond poate avea diferite nuanţe de la galben la verde. Determinismul culorii de fond este poligenic, considerându-se că pentru fiecare culoare există câte o grupă de gene separată. (BROWN, 1975) Aceleaşi gene sunt implicare şi în realizarea culorii pulpei.
Culoarea de acoperire este datorată prezenţei antocianilor care determină apariţia culorii roşii. La fructele de culoare galbenă sau verde, culoarea de acoperire lipseşte. Distribuirea culorii de acoperire pe suprafaţa fructului este diferită, putând fi totală, cu aceiaşi intensitate sau cu intensitate variabilă sau parţială, sub formă de pete sau dungi. La rândul lui, modul de distribuire a culorii de acoperire prezintă determinism genetic aparte. Pentru distribuirea culorii de acoperire sub formă de dungi sunt răspunzătoare gene majore, deoarece din încrucişarea unor soiuri cu fructe roşii cu soiuri cu fructe galbene sau verzi, nu pot fi obţinute fructe dungate, dar pot apărea fructe cu parţial colorate, pătate cu roşu. Raporturile de segregare diferă în funcţie de genotipurile cu care se lucrează.
Prezenţa culorii de acoperire este de cele mai multe ori dominantă asupra lipsei acestei. Din încrucişarea a două soiuri cu culoare de acoperire pot apărea şi fructe galbene sau verzi, ceea de demonstrează că genele răspunzătoare pentru culoarea de acoperire sunt în stare heterozigotă.
Culoare pulpei, la majoritatea soiurilor este de alb sau diferite nuanţe de gălbui, mai rar verde. Transmiterea acestor culori la descendenţi este controlată de mai multe gene şi nu prezintă reguli precise. Încrucişând între ele forme parentale cu aceiaşi culoare a pulpei în descendenţă pot apărea şi fructe de altă culoare. Totuşi, din încrucişarea unor genitori cu fructe cu pulpa albă nu pot apărea fructe cu pulpa galbenă, la fel cum din părinţi cu fructe cu pulpa galbenă nu pot apărea descendenţi cu fructe cu pulpa albă. În cazul hibridărilor între forme cu fructe cu pulpa de culoare verde, la urmaşi nu poate apărea culoarea galbenă şi albă a pulpei. Din încrucişarea a doi genitori cu pulpa de culoare crem, la descendenţi pot apărea toate tipurile de culori. Un caz aparte îl prezintă culoarea roşie a pulpei, care este determinată independent de celelalte culori, la baza acesteia fiind o genă dominantă preluată de la Malus niedzwtzkiana.
Gustul este o însuşire complexă care depinde de cantitatea şi proporţia de glucide, acizi şi substanţe aromatice din fruct. Calităţile gustative ale merelor sau format de-a lungul timpului odată cu trecerea de la starea sălbatică la cea cultivată sub directa influenţă a omului.
Combinarea acidităţii cu conţinutul în zaharuri este foarte importantă în realizarea gustului, fiecare dintre cele două grupe de substanţe având importanţa lor. Fructele cu aciditate slabă sunt fade şi nesavuroase. Cele foarte bogate în acizi sunt acre şi astringente, nepreferate de către consumatori. Cu toate că cele două grupe de substanţe sunt legate între ele în realizarea gustului, transmiterea lor este independentă.
Concentraţia în glucide se stabileşte prin determinarea procentului de zahăr din sucul obţinut din fructe. După modul de comportare a acestui parametru în descendenţele hibride se pare că este un caracter poligenic, descendenţii fiind situaţi apropiat mediei părinţilor. Aciditatea se transmite după alt model. Determinarea acidităţii se face prin intermediul stabilirii concentraţiei de acid malic. Concentraţiile mari de acid malic se transmit după modelul unui caracter dominant, pe când cele mici după modelul unor caractere cantitative. În cazul încrucişării a doi părinţi cu fructe foarte acide, toţi descendenţii for prezenta aciditate mare. În caz contrar, segregarea este diferită, de la genitor la genitor. Soiurile şi populaţiile locale vechi sunt deficitare în privinţa raportului glucide – aciditate. Aciditatea este mult mai pronunţată decât la soiurile ameliorate străine. (ARDELEAN, 1986)
Aroma fructelor este dată de peste 30 compuşi aromatici. Contribuţia fiecăruia la realizarea aromei nu a putut fi separată, motiv pentru care nu se pot efectua analize chimice precise, aroma
stabilindu-se prin degustări. Aroma trebuie să fie plăcută, fină, fructele să aibă miros plăcut, iar gustul să nu fie astringent şi acru.
Savoarea fructelor este determinată şi de suculenţa şi textura pulpei. Determinările la fructele proaspete nu sunt hotărâtoare deoarece, suculenţa se poate pierde în timpul păstrării. Textura pulpei trebuie fă fie mediu afânată. Nu sunt apreciate de către consumatori nici fructele cu pulpa tare, dar nici cele făinoase. Menţinerea calităţilor din acest punct de vedere depinde de prezenţa stratului de ceară pe fructe. Stratul de pruină împiedică pierderea apei menţinând fructele suculente. determinismul acestui caracter nu este pe deplin elucidat, dar se presupune că în realizarea lui intervin mai multe gene. Pe lângă rolul protector, stratul de ceară conferă şi un aspect plăcut şi atrăgător fructelor, în special la fructele de culoare roşie.
Fructele destinate industrializării, în linii mari, trebuie să îndeplinească aceleaşi condiţii de calitate ca şi fructele destinate consumului proaspăt. În cazul în care se doreşte obţinerea de gemuri şi concentrate, suculenţa este mai puţin importantă, foarte important fiind conţinutul în glucide. La acestea importantă este şi lipsa seminţelor din fruct. Soiuri fără seminţe se pot realiza, spre exemplu soiurile triploide. Dezvoltarea unor fructe fără seminţe prin partenocarpie la măr nu este cunoscută. S-a constatat că la unele soiuri, numărul seminţelor în fruct este foarte mic. Acesta a dus la ideea că lipsa seminţelor din fruct are o bază ereditară însă aceasta nu a putu fi pusă în evidenţă. Pentru industrializare, conţinutul în pectoze.
Din punct de vedere alimentar, conţinutul în vitamine este foarte important, fructele fiind consumate în stare proaspătă aduc aportul cel mai mare de vitamine. Dintre vitamine, pentru starea de sănătate a consumatorilor importanţa cea mai mare o prezintă vitamina C. Conţinutul în vitamina C este variabil de la un soi la altul. Determinarea conţinutului de vitamina C trebuie realizată prin analize speciale, acesta putând depăşi 250 mg la 1 kg fructe. În tipul păstrării o parte din vitamina C se pierde dacă păstrarea nu se realizează în condiţii optime.
Ameliorarea rezistenaţei la boli şi dăunători. Mărul este specia care necesită, poate, cel mai mare număr de intervenţii fitosanitare. În medie, într-un ciclu de vegetaţie sunt efectuate aproape 20 de tratamente împotriva bolilor, insectelor şi acarienilor. În unele situaţii, cum este atacul de foc bacterian, eficacitatea tratamentului este foarte scăzută. Oricare ar fi parazitul pentru care se urmăreşte obţinerea rezistenţei, selecţia are trei etape : găsirea sursei de rezistenţă, studiul extinderii bolii şi elaborarea strategiei de ameliorare. Principalele lucrări de selecţie pentru rezistenţa la boli se efectuează în perioada juvenilă (primii 4-5 ani de vegetaţie)
Găsirea sursei de rezistenţă începe cu studiul soiurilor ce aparţin speciei cultivate. Din păcate, în interiorul speciei se pot găsi doar genitori cu rezistenţă parţială., de la aceştia, transmiterea în urma hibridării fiind dificilă. Din acest motiv cele mai importante surse de rezistenţă rămân speciile sălbatice care, prezintă, în general rezistenţă totală bazată pe determinism simplu (una până la trei gene), însă de la aceste specii se preiau şi caractere nevaloroase, cum ar fi fructele mici şi de slabă calitate.
A doua etapă trebuie să cuprindă un studiu a agentului patogen şi o testare a durabilităţii sursei de rezistenţă. Aceasta este o fază esenţială în reuşita ameliorării. În această etapă trebuie să existe colaborare cu fitopatologii şi entomologii, care să cunoască foarte bine agenţii patogeni şi dăunătorii. În final, trebuie reţinute acele surse de rezistenţă care au efect faţă de o gamă cât mai largă de rase şi biotipuri agenţilor patogeni sau dăunătorilor. Poate apărea însă un risc foarte mare în această strategie, apariţia unor rase şi biotipuri noi faţă de care sursele găsite să nu prezinte eficacitate. Importanţa riscului depinde de gradul de cunoaştere a agenţilor patogeni. La măr, sunt specificate pentru fiecare din agenţii patogeni, un anumit număr de rase, dar rase noi pot apărea oricând ca rezultat a diversificării agenţilor patogeni. (LESPINASSE, 1989)
Punerea la punct a strategiei de ameliorare (ultima etapă) constă, de fapt în realizarea unui program de hibridare pentru obţinerea celor mai bune combinaţii de gene de rezistenţă la boli. În planificarea hibridărilor nu trebuie uitate nici caracterele agronomice. Dacă unul din genitori prezintă valoare redusă din punct de vedere agronomic, metoda care trebuie aplicată este back-cross-ul.
Rezistenţa la boli obţinută prin ameliorare poate fi de două tipuri. Primul tip este rezistenţă controlată genetic relativ simplu, putând fi situaţia unor blocuri de gene înlănţuite (cazul rezistenţei la rapăn datorată genelor “Vf”). Al doilea tip are la bază 2-3 gene complementare (cum este cazul
rezistenţei la făinare prezentă la M.zumi). Al doilea tip de rezistenţă există pentru rapăn, făinare, foc bacterian. Mai există şi rezistenţa bazată pe o singură genă specifică unei singure rase a agentului patogen, însă acest tip nu se utilizează deoarece, faţă de o altă rasă, soiul este total sensibil.
Ameliorarea rezistenţei la rapăn (Venturia inaegualis) este un obiectiv principal deoarece soiurile cultivare, practic, toate sunt sensibile. Există însă şi unele soiuri foarte vechi care sunt mai puţin atacate. La acestea, rezistenţa prezintă un determinism poligenic foarte dificil de transferat în programe de ameliorare. O ameliorare mai uşoară se poate realiza pe baze rezistenţei existentă la speciile înrudite cu specia cultivată. Durabilitatea rezistenţei a făcut ca să fie catalogate cinci surse de rezistenţă de tipul blocurilor de gene îmbunătăţite sau a genelor cu efect compelementar (LESPINASSE, 1989). Cele mai utilizate gene de rezistenţă sunt genele "Vf" preluate de la Malus floribunda. Transferul şi selecţia se desfăşoară mai rapid dacă se lucrează în sere şi pepiniere. Prima etapă este hibridarea, urmată de plantarea puieţilor hibrizi în primul an în seră. Frunzele foarte tinere ale puieţilor din primul an se infectează artificial. Infecţia se face prin stropire cu suspensie de conidii de la cele mai frecvente rase ale agentului patogen. Urmează o etapă de incubare la o temperatură de 180C într-o atmosferă cu umiditate ridicată. După incubare boala se manifestă pe frunze şi se pot face notările şi selecţia deoarece rezistenţa plantei adulte este foarte bine corelată cu cea a puietului din primul an.
Rezistenţa poligenică nu este totală şi nu există o legătură foarte strânsă între rezistenţa la nivelul foliajului şi cea la nivelul fructelor. Din acest motiv testarea continuă în pepinieră unde se observă şi rezistenţa la făinare. Selecţia definitivă are loc după primii ani de fructificare pentru a fi observat şi atacul la fructe şi calitatea acestora.
De obicei, se adoptă strategii de selecţie mai complexe în care se asociază pentru aceiaşi boală două mecanisme independente, alături de mecanisme similare pentru alţi agenţi patogeni sau dăunători.
Pe lângă rezistenţa la rapăn, atenţie sporită se acordă rezistenţei la atacul de făinare (Pedosphaera leucotricha). Făinarea este o boală foarte frecventă la unele soiuri, altele fiind mai puţin atacate. Agentul patogen atacă lăstarii şi foliajul putând duce la diminuarea producţiei şi a calităţii acesteia. În condiţiile României, făinarea are o mare extindere în toate zonele tării. Dacă boaba se instalează timpuriu, producţia pomului poate fi serios afectată. (BOLBOSE CECILIA, 2003) Determinsimul genetic al acestei boli este de tip cantitativ, pentru a imprima rezistenţă bună hibrizilor, ambi genitori trebuie să fie rezistenţi. Pe lângă rezistenţa de tip poligenic au fost identificate şi surse monogenice de rezistenţă în cadrul speciei Malus robusta, gena fiind dominantă (“Pl1”), dar şi în cadrul speciei M.zumi (“Pl2”).
Alte boli faţă de care sunt preocupări mai recente în ameliorarea mărului sunt: focul bacterian, cancerul produs de Nectria galligena, monilioza produsă de Monilina fructigena sau diferite boli care apar la fructe în timpul conservării.
Pentru integrarea tuturor obiectivelor de ameliorare sunt elaborate scheme generale de ameliorare. Cea mai folosită schemă are la bază încrucişări urmate de etape succesive de selecţie. Principalul scop urmărit este îmbunătăţirea rezistenţei la boli care se realizează prin asocierea a două mecanisme diferite pentru ca rezistenţa să fie de durată. Hibridările se fac într-o anumită ordine, deoarece în cele mai multe cazuri, la obţinerea hibridului dorit participă mai mulţi genitori. Pentru că rapănul este boala cea mai frecventă, toţi genitorii trebuie să prezinte rezistenţă la rapăn, fie de tip poligenic, fie de tip mai simplu (genele “Vf”, “Vr”). Fiecare din genitori trebuie să prezinte separat gene de rezistenţă la făinare (genele “z”), la focul bacterian şi la păduchele cenuşiu (genele “Y”).
Pentru continentul American se acordă atenţie şi ruginii (Gymnosporangium juniperi – virgianae). Boli mai puţin cunoscute, dar cu extindere tot mai mare sunt şi micoplasmozele care provoacă proliferarea mărului, elasticitatea lemnului la măr, atrofia fructelor de măr (CONSTANTIN GABRIELA, 2002)
Cu toate că în urma ameliorării rezistenţa la boli a soiurilor noi s-a îmbunătăţit mult, aceste soiuri s-au extins în producţie foarte lent datorită livezilor existente dar şi datorită pieţei, existând cerinţe faţă de fructele soiurilor cunoscute. Ameliorarea trebuie să continue îmbunătăţirea rezistenţei la boli deoarece combaterea chimică nu este exclusă în totalitate nici la cele mai valoroase soiuri.
Ameliorarea rezistenţei la dăunători. Cel mai răspândit dintre dăunători este păduchele lânos (Erisoma lanigerum). Acest dăunător este foarte periculos pentru că atacă toate organele plantei, atât cele aeriene cât şi rădăcinile. Atacul părţilor aeriene poate fi combătut prin tratamente chimice, dar atacul la
rădăcină depinde de rezistenţa genetică. Din observaţiile cultivatorilor s-a constatat că există diferenţe între soiuri în privinţa rezistenţei. Acesta a făcut ca să fie căutate surse de rezistenţă. Determinismul rezistenţei a fost studiat de-a lungul timpului şi au fost depistate, atât surse monogenice cât şi poligenice de rezistenţă.
Primele surse de rezistenţă au fost descoperite la soiul Northern Spy, ulterior fiind găsite surse de rezistenţă şi în cadrul unor forme ale speciilor Malus sargentii şi M.coronaria.
Rezistenţa soiului Northern Spy este foarte cuprinzătoare, neexistând diferenţe la nivelul rezistenţei faţă de diferitele rase fiziologice ale dăunătorului. Aceasta a dus l concluzia că rezistenţa acestui soi este de tip poligenic, cu toate că a fost susţinută şi ideea rezistenţei monogenice, la baza acesteia stând o genă dominantă (“Er”). Introducerea surselor de rezistenţă la păduchele lânos este un obiectiv important atât pentru ameliorarea soiurilor de măr cât şi în ameliorarea portaltoilor.
Un alt dăunător foarte frecvent este Cydia pommonella însă faţă de acesta nu sunt cunoscute gene de rezistenţă. Cu toate eforturile de ameliorare a rezistenţei la acest dăunător, combaterea atacului se rezumă la tratamentele chimice şi, mai recent la cele biologice. Alături de acest dăunător, depreciere fructelor este făcută şi datorită atacului de acarian auriu (Aculus achlechtendali), dăunător mai puţin cunoscut, ca urmare şi rezistenţa genetică a soiurilor este la fel de puţin studiată. (TOMOIOAGĂ LILIANA, 2002)
Tot mai frecvente sunt atacurile de larve miniere. Numărul speciilor care sunt cuprinse în această categorie este mare, făcând parte din diferite genuri. Între soiuri nu se constată diferenţe în privinţa rezistenţei şi nici nu se cunosc surse de rezistenţă pentru că studiile în acest sens sunt foarte puţine. (COPĂCESCU VALERIA, 2003)
Ameliorarea rezistenţei la ger. Extinderea arealului de cultură a mărului este limitată de nivelul temperaturilor. Cu toate că mărul este o specie rezistentă la temperaturi scăzute, valori foarte coborâte ale gerurilor din timpul iernii pot fi dăunătoare pomilor. Asemenea situaţii apar în zonele montane şi chiar de deal, unde cultura mărului este foarte extinsă.
Rezistenţa la ger este un caracter complex în privinţa determinismului genetic. S-a constatat că unii genitori transmit mai bine urmaşilor acest caracter decât alţii. Rezistenţa la ger depinde de o serie de factori interni şi externi. Factorii interni hotărâtori sunt reacţia chimico-fizică a celulelor la acţiunea temperaturilor scăzute, durata repausului plantei în tipul iernii şi capacitatea de regenerare a ţesuturilor.
Ereditatea rezistenţei la ger nu este bine elucidată. Se consideră că la baza rezistenţei sunt implicate mai multe gene, descendenţii situându-se la nivelul mediei părinţilor sau mai apropiat de părintele mai slab rezistent. Din aceste constatări s-a dedus că efectele predominante în realizarea rezistenţei la ger sunt cele aditive. (WATKINS şi SPANGELO, 1970)
Îmbunătăţirea rezistenţei la ger se poate realiza pe baza germoplasmei genului Malus. Speciile M.baccata, M.prunifolia şi M.ioensis sunt specii cu o bună rezistenţă la temperaturi scăzute. Transferul rezistenţei de la M.baccata aduce pe lângă rezistenţa foarte bună şi o serie de caractere nevaloroase, cum ar fi mărimea redusă a fructelor şi slaba calitate a acestora. În lucrările de selecţie pentru rezistenţa la ger trebuie vizate şi alte caractere care pot fi preluate de la speciile înrudite, cum este înflorirea tardivă. În ceea ce priveşte rezistenţa la ger, trebuie evaluat gradul rezistenţei atât la nivelul lemnului cât şi la nivelul mugurilor.
Pentru a se transmite o rezistenţă cât mai apropiată de cea a părintelui cel mai valoros, este bine ca acesta să fie utilizat ca formă mamă. S-a observat o corelare a rezistenţei descendenţilor cu rezistenţa formei mamă.
Ameliorarea epocii de înflorire. Epoca de înflorire este foarte importantă pentru cultura mărului. De ea depinde realizarea polenizării având în vedere faptul că majoritatea soiurilor sunt autosterile. Simultaneitatea la înflorit a soiurilor dintr-o plantaţie este obligatorie pentru polenizare şi realizarea producţiei de fructe. Pe de altă parte, epoca de înflorire are implicaţii în evitarea influenţei gerurilor târzii din primăvară.
Din punct de vedere practic se doreşte ca înflorirea să se realizeze cât mai târziu în primăvară deoarece riscul datora gerurilor târzii este eliminat. Trebuie avute în vedere însă, corelaţiile acestei însuşiri cu alte caractere. Formele care înfloresc târziu realizează maturizarea tardivă a fructelor, iar calitatea acestora este slabă. Ca urmare, pentru obţinerea unor soiuri de vară şi toamnă nu pot fi utilizaţi
ca genitori forme cu înflorire foarte întârziată. Data înfloritului este un caracter destul de stabil, fiind considerat un caracter poligenic. În urma
hibridărilor, distribuirea descendenţilor se produce în jurul mediei părinţilor. În cazul încrucişărilor între părinţi extremi (înflorire foarte tardivă cu înflorire foarte precoce) adesea apar transgresiuni, mult mai precoce decât părintele cel mai precoce şi mult mai tardive decât părintele cel mai tardiv. Transgresiunile nu apar la încrucişările dintre genitori asemănători în privinţa epocii de înflorire.
Pentru a uşura munca de selecţie a acestui caracter se poate lua în considerare corelaţia pozitivă dintre precocitatea înfloritului şi precocitatea apariţiei frunzelor la puieţii din primii ani de viaţă. Astfel se poate face o selecţie a formelor dorite încă înainte de intrarea pe rod. (MURAWSKI, 1967)
Ameliorarea precocităţii. Programele de ameliorare urmăresc să acopere o perioadă cât mai lungă de timp cu epoca de maturizare a fructelor, adică o coacere eşalonată de la extratimpuriu la foarte tardiv. Eforturile cele mai mari se depun însă pentru obţinerea soiurilor tardive, cu mare capacitate de conservare.
După epoca de maturizare a fructelor, soiurile de măr se împart în soiuri de vază, soiuri de toamnă, soiuri de iarnă şi soiuri de iarnă târzie.
În privinţa epocii de coacere, la măr variabilitatea este foarte mare. În condiţiile ţării noastre, primele soiuri pot fi recoltate la începutul lunii iulie, iar cele mai tardive, la sfârşitul toamnei. Maturizarea fructelor este o însuşire ereditară cu determinism complex. În urma încrucişărilor, hibrizii se comportă intermediar formelor parentale, cu tendinţă de apropiere de unul dintre părinţi. Nu au fost semnalate cazuri clare de dominanţă. Foarte frecvente transgresiunile, motiv pentru care se consideră că acest caracter este determinat poligenic.
Încrucişând între ele soiuri de vară se obţin cu precădere hibrizi la nivelul părinţilor, dar şi variaţii mai tardive. Din încrucişarea unor soiuri de vară cu soiuri de iarnă rezultă populaţii majoritar de toamnă, dar şi un procent destul de însemnat de forme de vară. Foarte puţine vor fi formele de iarnă. De aici se constată o tendinţă de asemănare a hibrizilor cu părintele mai timpuriu.
Atingerea unui anumit nivel al precocităţii se poate obţine printr-o alegere corespunzătoare a formelor parentale, în special a formei mamă, având în vedere originea genetică şi geografică a acesteia.
Ameliorarea rezistenţei la secetă. Rezistenţa la secetă este un obiectiv mai puţin luat în considerare deoarece în zona principală de cultură a mărului precipitaţiile sunt suficiente. În condiţiile României, sunt necesare soiuri rezistente pentru sudul şi sud-estul ţării. Îmbunătăţirea rezistenţei la secetă se poate realiza numai pe baza genelor provenite de la speciile înrudite, deoarece nu se cunosc soiuri cultivate valoroase din acest punct de vedere.
Ameliorarea capacităţii de păstrare. Păstrarea merelor în depozit este condiţionată de condiţiile de păstrare, dar şi de însuşirile genetice ale soiului. Rezistenţa la păstrare este observată de la începutul maturării de consum până la apariţia procesului de autoliză. O păstrare mai îndelungată o prezintă soiurile tardive şi cele cu maturizare după recoltare. Acestea au o activitate fiziologică a fructelor încetinită şi un start de ceară pe fructe bine dezvoltat astfel că păstrarea se poate face până în primăvară. Rezistenţă bună la păstrare trebuie să aibă şi soiurile de vară, deoarece de la recoltat până la consum, de cele mai multe ori, trece o anumită perioadă de timp. În menţinerea calităţii pe parcursul păstrării foarte importantă este capacitatea de menţinere a apei în fruct. Păstrarea depinde de zestrea ereditară a soiului, de condiţiile de depozitare, dar şi de tehnologia aplicată, în special nivelul de fertilizare. (CHIRA şi POPESCU, 2001; ŞERBĂNOIU ALBERTINA, 2003)
Pentru realizarea unei rezistenţe la păstrare bune, la toate tipurile de soiuri, este necesar ca în hibridări, ambii genitori să fie valoroşi.
ÎNMULŢIREA LA O SPECIE DE POMI – LEGUME - VIE
AMELIORAREA TOMATELORMetode de ameliorare.Tomatele au fost introduse recent în ameliorare. Variabilitatea speciei provine din introducerea
repetată în Europa a unor provenienţe din Mexic, iar mai târziu apariţia unor forme diverse din diferite ţări europene sau SUA; create de către grădinari şi comercianţi provenind de la unele amestecuri, hibridări accidentale sau mutaţii. Tomatele nu au făcut obiectul ameliorării prin hibridări controlate urmate de alegere în descedentare hibride, de cât după anii 1920 în SUA, iar mai târziu, după anii 1960 au fost realizate selecţii adevărate în alte ţări.
Până la această dată, obiectivul a fost obţinerea unor soiuri stabilizate (tip linie pură) care se deosebea între ele prin precocitate, mărimea şi forma fructelor. După 1983, ALABUVEETTE şi TITARD subliniază importanţa heterozisului la tomate, care se manifestă printr-o creştere a potenţialului productiv al hibridului comparativ cu formele parentale, aceasta imaginând premisele cultivării în producţia hibrizilor.
toate cultivarele actuale disponibile au fost obţinute prin metode clasice de ameliorarea plantelor autogame la care, tot mai mult se impun hibrizii F1 performanţi. Selecţia genealogică, de departe, este metoda cea mai folosită, ea este eficace pentru fixarea unor caractere cu determinism simplu, numeroase la tomate, permiţând în acelaşi timp eliminarea rapidă a genotipurilor cu defecte, în special cele legate de aspectul fructelor, care fac incultivabile astfel de soiuri. Sele3cţia individuală este folosită în ameliorare pentru caracterele cu eritabilitate slabă, cum este cazul conţinutului de substanţă uscată solubilă în fruct.
Selecţia este metoda cea mai veche folosită în crearea de noi soiuri. În prezent metoda este foarte puţin folosită deoarece procesul de ameliorare de începe cu efectuarea de hibridări pentru producerea de variabilitate. Astfel, selecţia va fi o metodă care însoţeşte una din metodele folosite pentru provocarea variabilităţii.
Selecţia aplicartă la tomate, în întreaga lume, în cursul, ultimilor 40 de ani s-a dovedit eficace şi a dus la obţinerea unei palete de soiuiri extrem de diversificate. Rezultatele aplicării metodelor clasice de ameliorare continuă să apară şi în viitor.
Amelioratorii de tomate, de scurt timp practică selecţia recurentă, alternând generaţiile de interîncrucişări cu autopolenizarea pentru acumularea genelor favorabile pentru anumite caractere cu determinism poligenic spre exemplu rezistenţa la viroze (Yellow Leafe Curl), rezistenţa la musca minieră (Liriomyza ssp.) şi într-o manieră mai largă, adaptabilitatea la condiţiile de mediu şi calitatea fructelor. Aceste metode se completează una pe alta şi se pot folosi alternativ în programele de ameliorare. Pe de altă parte testarea combinaţiilor hibride trebuie făcută cu regularitate în timpul ameliorării.
Hibridarea este metoda de bază aplicată în ameliorarea tomatelor. Se aplică toate tipurile de hibridare de la cea apropiată până la cea îndepărtată. Folosirea hibridării în ameliorarea tomatelor s-a practicat încă din secolul al XIX-lea. Odată cu dezlegarea eredităţii caracterelor, hibridarea a început a fi folosită programat pentru obţinerea combinaţiilor de caractere dorite.
Colecţiile din centrele de ameliorare a tomatelor sunt foarte ample şi oferă material iniţial bogat pentru realizarea de combinaţii hibride din ce în ce mai complexe.
Hibridarea interspecifică este tot mai mult folosită în ameliorarea tomatelor, odată ce s-a descoperit că în cadrul speciilor înrudite există gene de rezistenţă la temperaturi scăzute, la atacul bolilor sau gene pentru sporirea conţinutului fructelor în vitamina C. Astfel încrucişările cu L.peruvianum şi L.hirsutun sunt foarte utilizate. Hibridările cu aceste specii nu sunt dificil de realizat, problema principală care se pune este sterilitatea hibrizilor F1, în special la combinaţiile cu L.hirsutum.
Back-cross-ul este o variantă a hibridării foarte utilizată la tomate. Soiurile actuale prezintă performanţe productive importante şi o calitate a fructelor destul de bună. Multe dintre ele sunt însă deficitare în privinţa rezistenţei la boli, în special la septorioză, mană, fuzarioză, pătarea brună a frunzelor şi la viroze. Gene în privinţa rezistenţei la aceste boli pot transferate prin back-cross de la speciile înrudite sau de la soiurile care le posedă. După hibridarea cu forma donatoare urmează retroîncrucişări cu soiul valoros până la refacerea genotipului acestuia în cadrul lui existând şi genele preluate de la donator.
Heterozisul valorificat prin cultivarea hibrizilor este metoda cea mai recentă cu toate că la plantele autogame ameliorarea este bazată pe crearea de soiuri.
WELINGTON (1912) a observat prezenţa heterozisului la tomate şi a încercat utilizarea lui în practică. De atunci crearea de hibrizi comerciali F1 a fost încercată şi au fost evidenţiate sporuri faţă de media părinţilor, dar şi sporuri faţă de cel mai valoros dintre părinţi în privinţa productivităţii şi calităţii fructelor. Un mare avantaj al hibrizilor este acela că unele corelaţii nu pot fi uşor rupte în cadrul soiurilor, dar la hibrizi ele sunt mai uşor de realizat. Astfel, corelaţiile dintre mărimea fructelor şi tardivitate, dintre mărimea fructelor şi conţinutul redus în vitamina C nu pot fi desfăcute prin aplicarea selecţiei în populaţii hibride. Ele se manifestă însă foarte slab la hibrizii F1. Foarte valoroşi sunt hibrizii timpurii care dau şi producţii foarte mari.
Heterozisul este la fel de pronunţat chiar dacă condiţiile mediu sunt mai puţin favorabile. Analiza factorilor ce influenţează productivitatea arată că ameliorarea acesteia vine dintr-o mai bună legare a fructelor; un procent mai mare de flori care produc fructe, fructe mai mari care se datorează unui număr mai mare de seminţe (masa fructului unui soi este corelată direct cu numărul de seminţe care le conţin). Dacă se descompune mai departe fenomenul legării, se ajunge la concluzia că în condiţii nefavorabile, de exemplu temperaturi scăzute, trebuie să existe heterozis sau dominanţă pentru cantitatea de polen formată şi pentru calitatea polenului (estimată in vitro prin colorarea cu carmin acetic), şi heterozis pentru valoarea de fecundare a polenului (estimată in vivo prin numărul de seminţe de către flori castrate ale unei forme mamă cultivată în condiţii optime).
Un alt avantaj al hibrizilor F1, este acela că permit acumularea unor gene dominante de rezistenţă la mai multe boli. Ca şi la alte specii cultivate, comercializarea hibrizilor F1 permite protejarea materialului vegetal şi rentabilizarea procesului de ameliorare.
Hibrizii F1 câştigă progresiv teren la toate tipuri de culturi începând cu cele mai profitabile: culturile din seră (unde nu se mai întâlnesc hibrizi), apoi culturile palisate din câmp, apoi culturile nepalisate din câmp pentru fructe proaspete şi sfârşind cu cele pentru industrializare (la care hibrizii F1
care au fost introduşi recent). În acelaşi timp, pentru ţările mai puţin avansate, pentru care preţul seminţei hibride este foarte costisitor, este prioritară crearea soiurilor (de tip linie pură), introducând în acestea, în special gene de rezistenţă la boli.
În crearea hibrizilor comerciali la tomate, ca forme parentale se folosesc soiuri sau chiar linii obţinute prin metode clasice de ameliorare. S-a constat că cele mai performante soiuri sunt şi cele mai bune forme parentale ale hibrizilor comerciali. Pentru depistarea acestora, pe lângă performanţele productive este testată şi capacitate combinativă. Foarte mult timp ca forme parentale au fost folosite linia L 10 şi soiurile Bison pentru culturi timpurii şi soiul Rutgers pentru culturile tardive.
La noi în ţară au fost create linii proprii care stau la baza primilor hibrizi comerciali româneşti. Producerea seminţei hibride comerciale nu este perfectată îndeajuns. Castrarea formelor mamă se
face manual la fel ca şi polenizările artificiale. Astfel preţul de cost al seminţei hibride creşte. Prin utilizarea androsterilităţii acest neajuns ar putea fi eliminat. La tomate se cunoaşte androsterilitatea genică datorată genelor “ps”. În cazul acestui tip de androsterilitate în urma înmulţirii liniei androsterile se obţine un amestec de plante androsterile şi androfertile. Acestea se produc ca răsad şi se ţin în sere până la înflorire când se face separarea lor prin observarea florilor. În câmp se vor planta numai plantele androsterile.
Androsterilitatea citoplasmatică nu se cunoaşte la tomatele cultivate. Cu toate că există sistemul operaţional prin care se pot obţine gene mutante recesive de androsterilitate, unele prezentând markerii necesari (morfologici sau izoenzime) legaţi de genele de androsterilitate, producerea seminţei hibride are la bază forme mamă normale, însă această situaţie trebuie schimbată. Prin utilizarea androsterilităţii se
reduce mult preţul de cost al seminţei hibride fiind eliminat un volum mare de lucrări manuale (BĂLAŞA, 2001)
Mutageneza este o metodă destul de eficientă la tomate. Inducerea mutaţiilor se încearcă pentru obţinerea unor gene care trebuie preluate de la speciile înrudite. Folosirea unor astfel de gene mutante are avantajul că nu mai este necesară îndepărtarea genelor nedorite care se preiau de la speciile sălbatice.
Agenţii mutageni ei mai folosiţi sunt radiaţiile ionizante. Se recomandă două iradieri, una la polen şi alta la seminţele rezultate din polenizarea cu polen iradiat. Principalele caractere afectate de mutaţii sunt cele cu determinism simplu, cum ar fi: culoarea şi forma fructelor, prezenţa sau absenţa pubescenţei, tipul de creştere şi chiar obţinerea unor forme precoce.
Metode neconvenţionale de ameliorare. Variabilitatea care există la speciile sălbatice din genul Lycopersicon oferă avantaje care trebuie exploatate. Pentru aceasta diferitele tehnici ale biotehnologiilor oferă perspective total noi.
Markerii moleculari vor fi un mare ajutor pentru ameliorarea tomatelor. Există deja rezultate în acest domeniu. Astfel sunt menţionate legăturile între markerii izoenzimici şi genele de androsterilitate sau rezistenţă la boli, legăturile între markerii RFLP şi conţinutul în substanţă uscată solubilă din fruct. Existenţa unei hărţi comozomiale cu o densitate destul de mare de markeri RFLP şi dezvoltarea de noi markeri moleculari vor permite impoprtante progrese în ameliorare.
Inginerie geneteică deschide şi alte perspective. Proiecte mai mult sau mai puzţin elaborate, mai mult sau mai puţin avansate sunt aplicate în ameliorarea tomatelor. Din această catregorie pot fi amintite obţinerea de plante rezistente la erbicide, la insecte şi diverşi agenţi patogeni, obţinerea plantelor androsterile. Obţinerea tomatelor transgenice permite deja revizuirea completă de către tehnologi a ipotezelor emise cu referire la maturizarea fructelor.
Metodele neconvenţionale deschid perspective obţinerii unor soiuri cu arhitectură deosebită, a unei producţii superioare din punct de vedere calitativ şi liberă de substanţele nocive rămase de la numărul mare al tratamentelor chimice pentru combaterea bolilor care se mai practică în tehnologiile clasice.
AMELIORAREA MARULUIMetode de ameliorare.
Hibridarea şi mutageneza sunt metodele cele mai folosite în ameliorarea mărului. Mutageneza se utilizează la crearea a unor caractere inexistente la specia cultivată, , pe când hibridarea se foloseşte pentru a corecta unele soiuri deja extinse în cultură, pe baza speciilor înrudite.
Selecţia. Selecţia aplicată în valorificarea variabilităţii naturale este metoda care a stat la baza obţinerii primelor soiuri ameliorate. Variaţii naturale mai pot apărea în livezile de producţie, dar depistarea lor este dificilă, trebuind explorat un volum mare de material biologic. Aceste variaţii sunt frecvente datorită mutaţiilor naturale şi a hibridărilor datorate alogamiei. Cel mai uşor sunt depistate variaţiile mugurale datorate mutaţiilor naturale. Acestea pot fi utilizate direct ca noi soiuri, prin altoire. Hibrizii naturali se evaluează greu fiind neceară urmărirea populaţiilor hibride pentru valorificarea segregării, aceasta îngreunând procesul de ameliorare.
Tipul de selecţie care se aplică este selecţia vegetativă bazată pe principiile unei selecţii individuale. Prin selecţie pot fi create soiuri noi care prezintă formă deosebită a fructelor, culori şi combinaţii noi de culori, forme cu rezistenţă la boli şi dăunători îmbunătăţită şi chiar forme valoroase din punct de vedere productiv şi calitativ.
În perioada actuală, când procesul de ameliorare impune îmbunătăţirea unei game largi de caractere, selecţia este o metodă care însoţeşte metodele prin care se ajunge la variabilitatea dorită.
Hibridarea între soiurile cele mai valoroase se foloseşte în ameliorarea mărului de aproape un secol .Programe de hibridare se realizează continuu . Foarte frecvent se efectuează hibridări interspecifice, în special pentru îmbunătăţirea rezistenţei la boli. Aceste hibridări sunt urmate de back-cross, luându-se însă măsuri de prevenire a consangvinizării.
Hibridarea între soiuri se realizează cu scopul combinării caracterelor valoroase prezente la genitorii utilizaţi. În procesul hibridării se recomandă castrarea florilor la forma mamă pentru siguranţă, cu toate că multe soiuri sunt caracterizate de autosterilitate. Momentul optim pentru castrat este cu 2-3 zile înainte de deschiderea florii. Într-un corimb se castrează numai o singură floare, celelalte îndepărtându-se. Polenizarea se efectuează cu polen recoltat anterior. Pentru aceasta, de la forma tată se recoltează ramuri cu inflorescenţe neînflorite, care se aduc în camere încălzite şi se ţin la soare în vase cu apă. La o temperatură de 18-200C, în 2-3 zile anterele crapă şi eliberează polenul care poate fi folosit. Eliberarea polenului poate fi grăbită prin introducerea ramurilor în apă caldă la 30-320C, astfel putându-se obţine polen în 24 ore. Polenul poate să îşi păstreze viabilitatea până la 10-15 zile, dar se recomandă utilizarea polenului mai proaspăt, de până la trei zile. Momentul optim pentru polenizare este atunci când pe stigmate se observă secreţii lipicioase. Aceste secreţii nu se formează pe vreme secetoasă. Deoarece polenizarea se realizează cu dificultate, cantităţile de polen care se distribuie pe stigmate trebuie să fie cât mai mari. Controlul prinderii se poate efectua la o săptămână după polenizare.
Hibridarea intraspecifică oferă posibilitatea reunirii în cadrul aceluiaşi genotip a caracterelor valoroase provenite de la mai mulţi genitori. Alegerea genitorilor este etapa cea mai importantă deoarece impune o foarte bună cunoaştere a genotipului acestora. Mărul fiind o specie alogamă fără posibilităţi de consangvinizare, cunoaşterea genotipului este dificil de realizat. Multe gene recesive pot constitui surprize în prima generaţie hibridă. Dacă acestea sunt valoroase pot fi selecţionate încă din F1. Prelucrarea generaţiilor segregante la măr nu se practică în mod curent deoarece se prelungeşte foarte mult procesul de ameliorare. Pentru ca hibrizii să fie valoroşi se recomandă ca cei doi genitori să prezinte un număr cât mai mic de caractere nevaloroase.
S-a constata că şi la măr genotipurile sunt caracterizate de o anumită capacitate combinativă, astfel că unele soiuri sunt foarte valoroase ca genitori, transmiţând urmaşilor caracterele valoroase pe care le prezintă. Din acest motiv, foarte multe soiuri au la bază genitori comuni. La aceştia, caracterele valorose se comportă ca şi caractere dominante cu toate că determinismul lor genetic nu este foarte bine studiat. Caracterele fructelor, vigoarea plantelor, precocitatea înfloritului se comportă ca şi caractere dominante. Cu toate acestea, raporturile de segregare mendeliene nu se respectă. Aceasta se explică prin heterozigoţia accentuată a genitorilor şi prin diferenţele foarte mari dintre ei din punct de vedere genetic. O segregare foarte puternică se observă în descendenţele seminale, dar prelucrarea ei nu poate fi realizată datorită intrării pe rod foarte tardivă a puieţilor din seminţe. În segregare, marea majoritate a puieţilor se situează în jurul mediei părinţilor pentru majoritatea caracterelor. Puţine sunt cazurile extreme. Multe caractere se manifestă apropiat de forma mamă, dar sunt şi caractere cu manifestare apropiată de cea a formei tată (rezistenţă la ger, înflorire târzie). Alte caractere au o comportare cu totul întâmplătoare (forma şi dimensiunile fructelor, greutatea fructelor şi capacitatea de păstrare).
Hibridarea între soiuri se utilizează pe scară largă pentru obţinerea unor combinaţii dorite de caractere morfologice. Mai puţin eficientă este pentru combinarea însuşirilor gustative. Rezultatul final al hibridării este nu numai o corectare a unor soiuri mai vechi ci şi apariţia unor combinaţii de caractere inedite, sau chiar a unor caractere noi.
Hibridarea îndepărtată este larg utilizată în ameliorarea mărului cu toate că este utilizată mult mai recent decât hibridarea intraspecifică. Dat fiind numărul identic al cromozomilor la majoritatea speciilor din genul Malus, hibridarea interspecifică nu întâmpină greutăţi. Sunt menţionate hibridări reuşite cu speciile M.zumi. M.prunifolia, M.baccata, M.ringo, M.micromalus, M.niedzwetzkiana, M.halliana, M.ioensis şi chiar cu specii din genul Pyrus.
Utilizarea hibridării intraspecifice se face cu scopul de a îmbunătăţi însuşirile de rezistenţă la factorii de mediu nefavorabili (în special rezistenţă la ger şi secetă), dar şi a rezistenţei la atacul bolilor şi dăunătorilor. Speciile care se pot utiliza în acest scop au fost menţionate la categoriile de germoplasmă. Acestea au fost încercate astfel că multe soiuri conţin gene de rezistenţă preluate de la diferite specii ale genului Malus. Totuşi, acestea trebuie explorate în continuare pentru a găsi noi surse de gene impuse de diversificarea agenţilor patogeni.
Mutageneza. Inducerea mutaţiilor este o metodă foarte eficace şi se poate realiza folosind diverşi factori mutageni. Această metodă este aplicată de foarte multă vreme în ameliorarea mărului cu rezultate foarte bune.
Dintre factorii mutageni, cei fizici se aplică pe scară largă. Cel mai folosite sunt radiaţiile gama ale cobaltului 60 asupra mugurilor dorminzi, radiaţiile X, neutronii termici. După tratament urmează selecţia mutantelor în funcţie de caracterele agronomice. (DE COURTYE, 1971) Principalele dificultăţi a acestui mod de lucru sunt himerele, deoarece mutaţia nu afectează toate celulele mugurelui. Himerele sunt de mai multe tipuri, în funcţie de structura apexului. (PRATT, 1983)
Pe lângă agenţii mutageni fizici au fost încercaţi şi cei chimici, dar cu rezultate mai puţin satisfăcătoare.
DE COURTYE recomandă înmulţirea în pepinieră (generaţia a doua de multiplicare) a 5 muguri succesivi formaţi în apex după aplicarea tratamentului mutagen, acest număr fiind impus de filotaxia mărului care este 2/5.
Încă din pepinieră, aproximativ 60% din mutante pot fi depistate cu siguranţă, ele fiind cel mai adesea în stadiu de himeră. Pentru restul, adică mutante care sunt în stadii de sciziune izolate este nevoie să se realizeze o multiplicare vegetativă suplimentară. Mutantele ce se pot obţine la măr pe această cale pot influenţa portul pomului, tipul de ramificare şi fructificare, culoarea fructelor, productivitatea.
Poliploidia. Poliplodia cu toate că este o mutaţie de genom constituie o metodă de ameliorare aparte datorită utilizării în anumite scopuri. Forme triploide şi tetraploide sunt cunoscute şi ca variaţii naturale. În procesul de ameliorare, acestea se obţin cu ajutorul colchicinei aplicată pe muguri sau pe vârfuri de creştere ale lăstarilor.
Tetraploizii se obţin direct din formaţiunile colchicinizate şi pot fi uşor recunoscuţi fiind foarte viguroşi, cu tendinţă de îndesire, cu frunze groase. Fructele sunt mult mai mari şi turtite. Identificarea tetraploizilor se poate face mai devreme şi mai sigur după mărimea şi densitatea stomatelor.
Triploizii se obţin prin încrucişarea unor forme diploide cu forme tetraploide. Din astfel de populaţii pot fi selecţionate soiuri valoroase, caracterele negative ale tetraploizilor fiind mai puţin evidente. Avantajul acestor forme este lipsa seminţelor ceea ce le recomandă pentru industrializare.
Metode neconvenţionle de ameliorare. Biologia celulară deschide căi noi, graţie culturilor “in vitro”. La atingerea obiectrivelor de ameliorare care combină un număr mare de caractere foarte diferite, biotehnologiile, pot avea un rol hotărâtor în ameliorare şi înmulţire la măr. După definitivarea hărţii genetice a mărului vor putea fi elaborate programe de transfer de gene prin diferite metode ale ingineriei genteice.
Tehnicile de regenerare şi obţinerea protoplaştilor sunt puse la punct la mar (PATAT- OCMATT şi calab, 1988). Mutageneza “in vitro” este o metodă care se aplică asupra organelor sau celulelor cultivate “in vitro” fiind urmată de regenerare. Acestea permit selecţia unor mutante omogene în toţi locuşi izogenici (CERVEAN , 1990). Noi perspective sunt deschise de obţinerea de plante haploide prin androgeneză ginogeneză. Haploizi pot ajuta la o mai bună cunoaştere a speciei, manipulării prin fuziune de protoplaşti, inducerea de mutaţii, urmate toate de dublarea numărului de cromozomi şi creare a dihaploizilor ( LESPINASSE şi GODICHEAU, 1980, ZHANG şi colab. 1990)
AMELIORAEA VIŢEI DE VIE
Metode de ameliorare.
Selecţia. Selecţia este metoda prin care au fost create un număr foarte mare de soiuri şi au fost obţinute progrese în privinţa tuturor caracterelor importante. Aplicarea selecţiei se bazează pe variabilitatea pronunţată a unor soiuri vechi sau pe variaţiile mugurale care apar ca efect a mutaţiilor naturale.
Selecţia variabilităţii naturale se poate aplica eficient în plantaţiile vechi. Pentru a găsi elite valoroase se explorează un volum mare de material biologic. Butucii se marchează şi până la selecţia finală se urmăresc mai mulţi ani. Această urmărire constă în efectuarea de măsurători biometrice asupra fructificaţiilor, dar şi analize de calitate. Butucii care se constată că sunt valoroşi vor servi ca plante mamă pentru înmulţire în scopul creării unor clone care se vor testa în culturi comparative.
Variaţiile mugurale pot fi mai valoroase însă sunt mai greu de găsit deoarece variabilitatea nouă
se manifestă numai la nivelul unei corzi sau a unui lăstar, şi nu la nivelul întregului butuc.Ambele situaţii de valorificare a variaţiilor naturale sunt de fapt selecţii individuale clonale.
Selecţia clonală se aplică pe scară largă deoarece la înmulţirea prin seminţe se recurge doar în cazul hibridărilor. Populaţiile rezultate din seminţe sunt greu de urmărit, fiind foarte neuniforme, iar până la fructificare trebuie să treacă un număr mare de ani.
Plantele urmărite prin selecţie clonală se urmăresc până în anii 3-5 de fructificare şi numai după aceea sunt supuse înmulţirii spre a fi testate în culturi comparative.
Prin selecţie clonală se pot îmbunătăţi vechile soiuri creând selecţii superiore din punct de vedere productiv, calitativ sau a însuşirilor de rezistenţă. Noile selecţii din soiurile vechi se pot extinde repede în producţie deoarece nu mai trebuie prezentate cultivatorilor, aceştia cunoscâdu-le. Ele nu prezintă caractere noi marcante ci numai o îmbunătăţire a performanţelor.
Prin selecţie clonală pot fi create şi soiuri noi atunci când se porneşte de la variaţiile mugurale. Bonitatea acestora trebuie făcută foarte atent, atât în privinţa caracterelor morfologice ale strugurilor, cât şi a celor de calitate sau de rezistenţă.
Selecţia clonală se foloseşte atât în crearea soiurilor folosite ca altoi, cât şi a portaltoilor. În cazul portaltoilor, unde foarte importantă este rezistenţa la boli şi dăunători, este necesară aplicarea unor testări în condiţii de infecţie artificială înainte de înmulţirea spre testarea în culturi comparative. După înmulţire, când se dispune de material suficient este necesară şi o testare a afinităţii la altoire.
Hibridarea. Hibridarea este cea mai utilizată metodă de creare a variabilităţii. Se aplică în toate variantele, atât intraspecifică, cât şi interspecifică.
La speciile cu flori hermafrodite trebuie efectuată castrarea formei mamă. Lucrarea se execută înainte de înflorit, cu ajutorul unor pensete, cu care se smulge corola care formează un capişon deasupra stigmatului. Odată cu corola se smulg şi staminele. Pentru castrat, pe un ciorchine se lasă doar 20-40 de flori de la mijlocul acestuia. După smulgerea corolei se verifică dacă nu au rămas resturi de stamine sau chiar stamine întregi. Până la polenizare, care are loc după 2-3 zile, florile castrate se izolează. Polenizarea trebuie făcută când stigmatul devine receptiv, de preferat dimineaţa, între orele 6 şi 10. Foarte importantă este alegerea genitorilor. Se recomandă ca genitorul care prezintă cele mai multe caractere valoroase să fie folosit ca mamă.
Hibridarea intraspecifică este cea mai folosită. Hibridarea se realizează uşor, obţinându-se hibrizi fertili. Cele mai valoroase combinaţii hibride se obţin când se încrucişează soiuri din grupe ecologice diferite şi când ambii genitori sunt valoroşi. Caracterele nevaloroase se elimină foarte greu din populaţia hibridă.
Hibridarea inetrspecifică este folosită în ameliorarea însuşirilor de rezistenţă (ger, boli şi dănători) atât în ameliorarea soiurilor nobile cât şi a portaltoilor. Hibridările între speciile subgenului Euvitis reuşesc destul de bine, cele cu specii ale sungenului Muscadinia fiind mai dificile. Şi în cazul acestor hibridări alegerea genitorilor are un rol important în obţinerea unor hibrizi cu puţine caractere nedorite. În general, de la speciile înrudite se transmit şi multe caractere nefolositoare. Pentru a mări variabilitatea se recomandă a fi studiată şi generaţie F2 în urma hibridărilor îndepărtate. Această situaţie este impusă de faptul că de la speciile înrudite rămânând un număr mare de caractere nefolositoare, indivizii care acumulează numai caractere favorabile sunt puţini.
Pentru eliminarea mai rapidă a însuşirilor nevaloroase de la hibrizii îndepărtaţi, cât şi pentru a reface fertilizatea (în cazul în care aceasta este redusă) se recomandă utilizarea back-cross-ului. Ca părinte recurent se va utiliza soiul ce aparţine speciei V.vinifera sau a celei mai valoroase specii, în cazul portaltoilor. Numărul generaţiilor de back-cross nu poate fi prea mare. Pentru a nu prelungi foarte mult durata ameliorării , nu se vor practica decât 2-3 back-cros-uri (maxim).
Mutageneza. Mutageneza este o metodă care a fost încercată cu succes în ameliorarea viţei de vie. Se pot aplica atât factori mutageni fizici cât şi chimici. Rata mutaţiilor este destul de redusă, însă pot fi afectate unele caractere morfologice implicate în realizarea productivităţii şi a calităţii (boabe mari, fără seminţe, coacere precoce, aromă deosebită). Unele soiuri răspund mai bine la inducerea de mutaţii decât altele. Acestea trebuie depistate şi utilizate.
Aplicarea factorilor mutageni se face asupra mugurilor, a vârfurilor de creştere, a grăunciorilor de polen sau a seminţelor. Pentru a avea şansa depistării unor mutaţii valoroase, materialul supus mutaţiilor
trebuie să fie numeros.Selecţia mutantelor valoroase se face după metoda selecţiei clonale.Poliploidia. Poliploidia se foloseşte ca metodă de ameliorare în atingerea unor obiective speciale.
Poliploidizarea se realizează prin tratarea vârfurilor de creştere cu colchicină. Creşterea efectului colchicinei şi o lăstărire mai puternică se obţine dacă în soluţie se adaugă şi acid giberelic.
Formele tetraploide sunt mai viguroase, cu boabe mai mari şi sunt lipsite de seminţe au seminţe puţine. Verificvarea tetraploizilor se poate face observând caracterele morfologice specifice lor, dar pentru siguranţă se recomandă studiul citologic. Odată selecţionate, clonele tetraploide se înmulţesc vegetativ şi pot duce la apariţia unui soi noi.
Prin încrucişarea tetraploizilor cu diploizii pot fi create forme triploide. Acestea sunt forme total sterile, nu produc struguri cu seminţe şi prezintă toate caracterele specifice tetraploizilor. Faţă de aceştia au avantajul că prezintă o coacere a lemnului mai bună.
Consangvinizarea. Consangvinizarea este o metodă care s-a încercat în scopul valorificării unor gene recesive ascunse de starea heterozigotă a plantelor. Rezultatele nu au fost cele aşteptate deoarece ajungerea la un grad ridicat de homozigoţie prin consangvinizare, necesită foarte mult timp. Consangvinizarea poate fi folosită şi în scopul valorificării heterozisului rezultat din încrucişarea unor linii consangvinizate. Valoarea heterozisului nu justifică cheltuielile cu consangvinizarea. În plus, heterozisul este foarte frecvent şi la încrucişările dintre soiuri. Hibrizii F1 dintre soiuri, care prezintă heterozis pot fi înmulţiţi vegetativ, astfel heterozisul stabilizându-se.
Consangvinizarea şi heterozisul sunt metode care au fost încercate, dar s-a renunţat la ele în favoarea unor metode mai rapide şi cu eficienţă mai bună.
Metode neconvenţionale. Culturile “in vitro” s-au dezvoltat foarte mult la viţa de vie. La început au fost folosite pentru înmulţirea liberă de boli, dar în prezent de încearcă folosirea lor în provocare de variabilitate (mutageneză “in vitro”, hibridare somatică, etc.). avantajul acestor metode este foarte mare: poate fi urmărit un număr mare de plante, acestea pot fi supuse testărilor de rezistenţă în spaţii protejate, iar dacă este vorba de transferarea unor gene de la speciile înrudite, acest lucru devine posibil fără a fi transmise şi caractere nedorite.
Ameliorarea portaltoilor.
Ameliorarea portaltoilor este un obiectiv permanent în ameliorarea viţei de vie. Portaltoii trebuie să prezinte o sumă de însuşiri aparte. Portaltoii trebuie să prezinte un grad mare de adaptare la diferite condiţii de climă şi sol. Situaţiile cele mai deosebite apar pe terenurile nisipoase, cele bogate în calcar sau pietroase. GRECU, 2001)
Foarte bună trebuie să fie afinitatea la altoire cu soiurile nobile. La folosirea speciilor americane, multe soiuri nu prezintă prindere bună la altoire cu soiurile nobile. Acest defect a fost îndepărtat prin hibridările efectuate între speciile americane şi V.vinifera.
Portaltoii trebuie să fie şi rezistenţi la secetă. Pentru aceasta sistemul lor radicular trebuie să fie profund şi să imprime butucului un foliaj mai redus şi un coeficient de transpiraţie mic.
Rezistenţa la ger este o altă însuşire cerută portaltoilor. Aceasta este o caracteristică ereditară, dar depinde şi de gradul de coacere a lemnului.
Pentru a se înmulţi rapid, portaltoii trebuie să aibă o mare capacitate de lăstărire şi o creştere foarte rapidă a lăstarilor pentru a se obţine un număr cât mai mare de butaşi la unitatea de suprafaţă.
Nu se poate vorbi despre portaltoi valoroşi dacă nu prezintă o foarte bună rezistenţă la atacul de filoxeră (chiar imunitate) şi la atacul principalelor boli.
Aceste caractere şi însuşiri trebuie testate în procesul de ameliorare, portaltoii fiind studiaţi în culturi comparative.