1. Fiziologia neuronului La nivelul organismului animal, sistemele stimulo-integratoare sunt reprezentate de sistemul nervos şi sistemul endocrin, care datorită strânsei interrelaţii formează sistemul neuroendocrin. Sistemul endocrin este supus în permanenţă unui control reglator nervos. Sistemul nervos integrator este un ansamblu de structuri specializate în recepţionarea, transmiterea, depozitarea şi prelucrarea informaţiei. Din punct de vedere al naturii informaţiei procesate , cuprinde un sitem senzitiv, un sistem motor şi un sistem de integrare senzitivo-motorie. Sistemul nervos senzitiv, sau al analizatorilor, captează informaţia conţinută de diferiţi stimuli externi sau interni, o transportă la centrii nervoşi specializaţi, o depozitează şi o prelucrează. Sistemul nervos motor asigură capacitatea de răspuns adecvat a ţesuturilor şi organelor la stimulii recepţionaţi. Integrarea senzitivo-motorie se realizează la nivelul ganglionilor cerebroizi în cazul nevertebratelor, la vertebratele inferioare este preluată de talamus şi corpii striaţi, iar la vertebratele superioare revine cortexului cerebral, structură complexă, capabilă de activitate nervoasă superioară. Din punct de vedere al funcţiilor controlate , sistemul nervos cuprinde două componente: una care este răspunzătoare de integrarea organismului în mediul extern (sistemul nervos al vieţii de relaţie, sau somatic) şi alta care asigură reglarea funcţiilor interne (sistemul nervos al vieţii de nutriţie, sau vegetativ). Din punct de vedere structural şi topografic , distingem o componentă centrală, reprezentată de totalitatea centrilor nervoşi cuprinşi în nevrax şi o componentă periferică, formată din fibre nervoase grupate în nervi ce conectează sistemul nervos central cu senzorii şi cu efectorii. Activitatea sistemului nervos se desfăşoară pe bază de acte reflexe, care au ca bază structurală arcuri reflexe. 1.1. Structura funcţională a neuronului În alcătuirea sistemului nervos intră neuroni, celule gliale şi celule stem, capabile a se diferenţia în sensul formării de neuroni sau de celule gliale. Neuronii sunt celule alcătuite din corp celular (soma) şi prelungiri (dendrite şi axon). Cele trei componente ale neuronului prezintă particularităţi structurale diferite, perfect adaptate funcţiei. Corpul celular (soma) şi dendritele sunt învelite într-o membrană plasmatică subţire (neurilema), selectiv permeabilă pentru ioni şi în consecinţă polarizată electric. Canalele ionice sunt preponderent "de scurgere" şi ligand-dependente, dată fiind funcţia de componentă postsinaptică. Nucleul, situat de obicei central, este mare, în ciuda faptului că neuronul nu se divide. Nucleul este însă direct implicat în intensele sinteze proteice din somă, el deţinând informaţia genetică ce va fi transmisă prin transcriere neuriplasmei. Pe baza unor studii de hibridizare celulară, s-a constatat că în permanenţă se transcrie 1/3 din informaţia genomului, producându-se mai mult ARNm decât în orice alt tip de celulă. Din cauza înaltului nivel al activităţii de transcriere, cromatina nucleară este dispersată. În celulele gliale, prin contrast, cromatina se găseşte în aglomerări pe faţa internă a anvelopei nucleare. Majoritatea proteinelor formate de ribozomii liberi şi de poliribozomi rămân în corpul celular, în timp ce proteinele sintetizate pe reticulul endoplasmatic rugos (corpusculii Nissl sau corpii tigroizi) sunt exportate spre dendrite şi mai ales spre axon. Poliribozomii şi reticulul endoplasmatic rugos (RER) sunt caracteristici somei neuronului, axonul nu conţine RER şi nu poate sintetiza proteine. Reticulul endoplasmatic neted este implicat în depozitarea intracelulară a calciului şi menţinerea unei concentraţii constante a acestuia, de 10 -7 M. Creşterea prelungită a concentraţiei calciului intracelular peste această valoare duce la degenerarea şi moartea neuronilor. În neuroni, aparatul Golgi se găseşte de asemenea numai în soma. Ca şi în alte tipuri de celule, aceste structuri sunt angajate în glicozilarea terminală a proteinelor sintetizate de RER şi în formarea veziculelor de transport pentru aceste proteine, care sunt eliberate în citoplasmă şi trimise prin transport axonal spre porţiunea terminală a axonului. Neuronii cu proprietăţi secretorii (producători de neurohormoni) din nucleii hipotalamici posedă un aparat Golgi deosebit de dezvoltat. Transportul proteinelor de la aparatul Golgi depinde de modul de structurare a reţelei interne a citoscheletului. Citoscheletul neuronilor este format din microfilamente, neurofilamente şi microtubuli. Microfilamentele sunt formate din actină, o proteină contractilă caracteristică celulelor musculare, dar răspândită în multe alte tipuri de celule. Neurofilamentele se găsesc atât în dendrite, cât
Transcript
1. Fiziologia neuronului La nivelul organismului animal, sistemele stimulo-integratoare sunt reprezentate de sistemul
nervos şi sistemul endocrin, care datorită strânsei interrelaţii formează sistemul neuroendocrin. Sistemul endocrin este supus în permanenţă unui control reglator nervos.
Sistemul nervos integrator este un ansamblu de structuri specializate în recepţionarea, transmiterea, depozitarea şi prelucrarea informaţiei.
Din punct de vedere al naturii informaţiei procesate, cuprinde un sitem senzitiv, un sistem motor şi un sistem de integrare senzitivo-motorie. Sistemul nervos senzitiv, sau al analizatorilor, captează informaţia conţinută de diferiţi stimuli externi sau interni, o transportă la centrii nervoşi specializaţi, o depozitează şi o prelucrează. Sistemul nervos motor asigură capacitatea de răspuns adecvat a ţesuturilor şi organelor la stimulii recepţionaţi. Integrarea senzitivo-motorie se realizează la nivelul ganglionilor cerebroizi în cazul nevertebratelor, la vertebratele inferioare este preluată de talamus şi corpii striaţi, iar la vertebratele superioare revine cortexului cerebral, structură complexă, capabilă de activitate nervoasă superioară.
Din punct de vedere al funcţiilor controlate, sistemul nervos cuprinde două componente: una care este răspunzătoare de integrarea organismului în mediul extern (sistemul nervos al vieţii de relaţie, sau somatic) şi alta care asigură reglarea funcţiilor interne (sistemul nervos al vieţii de nutriţie, sau vegetativ).
Din punct de vedere structural şi topografic, distingem o componentă centrală, reprezentată de totalitatea centrilor nervoşi cuprinşi în nevrax şi o componentă periferică, formată din fibre nervoase grupate în nervi ce conectează sistemul nervos central cu senzorii şi cu efectorii.
Activitatea sistemului nervos se desfăşoară pe bază de acte reflexe, care au ca bază structurală arcuri reflexe.
1.1. Structura funcţională a neuronului În alcătuirea sistemului nervos intră neuroni, celule gliale şi celule stem, capabile a se
diferenţia în sensul formării de neuroni sau de celule gliale. Neuronii sunt celule alcătuite din corp celular (soma) şi prelungiri (dendrite şi axon). Cele trei
componente ale neuronului prezintă particularităţi structurale diferite, perfect adaptate funcţiei. Corpul celular (soma) şi dendritele sunt învelite într-o membrană plasmatică subţire
(neurilema), selectiv permeabilă pentru ioni şi în consecinţă polarizată electric. Canalele ionice sunt preponderent "de scurgere" şi ligand-dependente, dată fiind funcţia de componentă postsinaptică.
Nucleul, situat de obicei central, este mare, în ciuda faptului că neuronul nu se divide. Nucleul este însă direct implicat în intensele sinteze proteice din somă, el deţinând informaţia genetică ce va fi transmisă prin transcriere neuriplasmei. Pe baza unor studii de hibridizare celulară, s-a constatat că în permanenţă se transcrie 1/3 din informaţia genomului, producându-se mai mult ARNm decât în orice alt tip de celulă. Din cauza înaltului nivel al activităţii de transcriere, cromatina nucleară este dispersată. În celulele gliale, prin contrast, cromatina se găseşte în aglomerări pe faţa internă a anvelopei nucleare.
Majoritatea proteinelor formate de ribozomii liberi şi de poliribozomi rămân în corpul celular, în timp ce proteinele sintetizate pe reticulul endoplasmatic rugos (corpusculii Nissl sau corpii tigroizi) sunt exportate spre dendrite şi mai ales spre axon. Poliribozomii şi reticulul endoplasmatic rugos (RER) sunt caracteristici somei neuronului, axonul nu conţine RER şi nu poate sintetiza proteine.
Reticulul endoplasmatic neted este implicat în depozitarea intracelulară a calciului şi menţinerea unei concentraţii constante a acestuia, de 10-7 M. Creşterea prelungită a concentraţiei calciului intracelular peste această valoare duce la degenerarea şi moartea neuronilor.
În neuroni, aparatul Golgi se găseşte de asemenea numai în soma. Ca şi în alte tipuri de celule, aceste structuri sunt angajate în glicozilarea terminală a proteinelor sintetizate de RER şi în formarea veziculelor de transport pentru aceste proteine, care sunt eliberate în citoplasmă şi trimise prin transport axonal spre porţiunea terminală a axonului. Neuronii cu proprietăţi secretorii (producători de neurohormoni) din nucleii hipotalamici posedă un aparat Golgi deosebit de dezvoltat.
Transportul proteinelor de la aparatul Golgi depinde de modul de structurare a reţelei interne a citoscheletului. Citoscheletul neuronilor este format din microfilamente, neurofilamente şi microtubuli. Microfilamentele sunt formate din actină, o proteină contractilă caracteristică celulelor musculare, dar răspândită în multe alte tipuri de celule. Neurofilamentele se găsesc atât în dendrite, cât
şi în axon şi sunt formaţiunile care conferă rigiditatea şi menţin forma neuronului. Nu apar în vârfurile de creştere ale axonilor şi în spinele dendritelor, acestea fiind formaţiuni mai dinamice. Microtubulii sunt responsabili pentru transportul rapid al substanţelor prin dendrite şi mai ales prin axon. Au un diametru de 23 nm şi sunt formaţi dintr-o proteină numită tubulină. Proteinele neurofilamentelor au proprietăţi contractile, punând în mişcare sistemul de microtubuli. Împreună formează neurofibrilele, organite specifice neuronului, care pe lângă rolul structural îndeplinesc şi funcţia de a transporta proteine, vezicule cu mediator şi alte materiale necesare pentru a menţine integritatea structurală şi funcţională a celulei.
În axoni transportul anterograd (fig. 1.1) (ortodromic) se face cu viteză diferită, în funcţie de substanţele care sunt transportate. Viteza transportului axoplasmic lent este de 1-2 mm/zi. Astfel sunt deplasate substanţe cu rol hrănitor, proteine structurale de tipul actinei, neurofilamente şi microtubuli. Viteza transportului axoplasmic rapid este de 400 mm/zi; pe această cale ajung în zona sinaptică a axonului organite, vezicule cu mediatori şi glicoproteine membranare. Transportul rapid necesită Ca2+ şi ATP şi depinde de rata metabolizării oxidative a glucozei.
Fig. 1.1. Transportul axoplasmic anterograd (mediat de kinesină) şi retrograd (mediat de dineină). 1-nucleu; 2-RER; 3-aparat Golgi; 4-vezicule cu mediator aflate în drum spre butonii terminali; 5-neurofilamente; 6-microtubuli; 7- transport anterograd; 8-vezicule sinaptice; 9-eliberarea mediatorului prin exocitoză; 10-recuperarea prin endocitoză; 11-transport retrograd.
În dendrite, transportul anterograd se desfăşoară cu viteza de 0,4 mm/zi şi necesită de
asemenea ATP. Prin această modalitate sunt puşi în mişcare, pe lîngă alte substanţe, ribozomi şi ARN, sugerînd că şi dendritele sunt capabile de sinteze proteice. De altfel, la baza dendritelor şi chiar pătrunzând în acestea, pot fi observate concentrări de RER.
Prin transport axoplasmatic retrograd (antidromic) substanţele sunt deplasate centripet, dinspre terminaţiile prelungirilor spre corpul celular al neuronului. Astfel substanţele ce rezultă din inactivarea şi degradarea mediatorilor chimici la nivelul sinapselor sunt recuperate, prin endocitoză, în porţiunea terminală a axonului, conduse spre corpul celular şi reutilizate pentru noi sinteze. În cazul unor neuroni, menţinerea integrităţii funcţionale a sinapselor depinde de transportul transneural al unor substanţe trofice, ca factorul de creştere a nervilor, de-a curmezişul sinapsei. Odată ajuns la soma prin transport retrograd, factorul de creştere a nervilor activează mecanismele sintezei proteice.Tot antidromic sunt transportate spre corpul celular unele virusuri (herpetic, poliomielitic).
Neurofibrilele NU conduc impulsul nervos; acesta se propagă la nivelul neurilemei şi axolemei, prin mecanisme pe care le vom studia la subcapitolul despre conductibilitatea neuronului. S-a constatat că viteza de transport de-a lungul neurofibrilelor este constantă pentru una şi aceeaşi substanţă, indiferent de tipul de fibră nervoasă, în timp ce viteza de conducere a influxului nervos diferă în funcţie de tipul de fibră.
2
În corpul celular al neuronului se găsesc numeroase mitocondrii, ce furnizează ATP-ul necesar intenselor procese de sinteză a proteinelor (inclusiv a mediatorilor chimici). Dar cea mai mare concentrare a mitocondriilor se află la nivelul regiunii terminale a axonului, unde nu numai că produc ATP-ul utilizat ca sursă de energie în procesele asociate transmiterii sinaptice, ci furnizează şi substrate pentru sinteza unor substanţe cu rol de neurotransmiţători, ca de exemplu aminoacidul glutamat. Mai mult, mitocondriile conţin enzime implicate în degradarea moleculelor de mediatori chimici, ca monoaminoxidaza (MAO) ce degradează catecolaminele.
Axonul, prelungire unică, lungă, numită de aceea şi fibră nervoasă, conduce centrifug influxul nervos (PA) generat la nivelul conului axonic prin sumarea PL. Axonii neuronilor formează căile de conducere ascendente şi descendente din SNC, precum şi nervii periferici. Spre deosebire de dendrite, majoritatea axonilor sunt ramificaţi numai la capătul distal, unde formează arborizaţia terminală. Fiecare ramificaţie a arborizaţiei are un buton terminal, structură implicată în transmiterea sinaptică, mediată chimic, a influxului nervos de la un neuron la altul sau la organele efectoare. Axonii unor neuroni vegetativi eliberează mediatorii chimici din proeminenţe aflate pe traiectul lor, numite varicozităţi.
Axoplasma este străbătută pe toată lungimea sa de neurofibrile, care pătrund până în butonul terminal. Axolema este învelită de 3 teci: teaca de mielină, teaca lui Schwann şi teaca lui Henle.
Teaca lui Henle (endonervul) se află la exterior şi este de natură conjunctivă; asigură nutriţia şi protecţia neuronului. Sub ea se găseşte teaca lui Schwann, formată din celule gliale (celule Schwann), care aderă la axolemă şi se răsucesc în jurul axonului (fig. 1.2). Teaca de mielină, cu rol de izolator electric, situată în jurul axolemei, este produsă de celulele Schwann şi are structură lipoproteică, fiind de fapt o repliere a membranei plasmatice a celulei Schwann, care se răsuceşte în jurul axonului. Fibrele nervoase care posedă teacă de mielină bine dezvoltată sunt fibre mielinice, cu viteză mare de conducere a impulsului nervos. La fibrele amielinice, care au şi ele teacă Schwann, replierea membranei acestor celule în jurul axonului este redusă şi teaca de mielină este atât de subţire, încât nu-şi poate îndeplini funcţia izolatoare. În fibrele amielinice viteza de conducere a influxului nervos este considerabil mai mică decât în cele mielinice.
Două celule Schwann învecinate nu se ating, între ele există un spaţiu numit strangulaţie (nod) Ranvier, la nivelul căruia mielina lipseşte. Strangulaţiile Ranvier permit conducerea saltatorie a influxului nervos şi formarea colateralelor axonului, cu rol important în stabilirea conexiunilor interneuronale multiple.
Evoluţia tecii de mielină. Nevertebratele au numai fibre nervoase amielinice. Teaca de mielină este specifică vertebratelor şi atinge dezvoltarea maximă la mamifere. S-a constatat experimental că embrionii mamiferelor sunt sensibili la excitanţi sonori. La embrionul uman, la 3 luni de la fecundare încep să se mielinizeze fibrele nervoase ale căilor somestezice (mai întâi cele auditive, apoi cele pentru tact şi durere). Observaţiile au arătat că acei copii cărora li se cântă, în viaţa intrauterină, vor fi mai veseli şi vor avea un sistem nervos mai echilibrat. Înţelegem de aici şi influenţele nefaste pe care le poate avea expunerea organismului în formare la stimuli sonori excesivi (faptul că mama, în timpul sarcinii, lucrează într-un mediu cu mult zgomot sau obişnuieşte să asculte o muzică agresivă, desele certuri familiale etc.).
Fig. 1.2. Formarea tecii de mielină. A-secţiuni transversale printr-un axon, în diferite stadii de mielinizare. Celula Schwann se poate răsuci în jurul axonului înfăşurându-l în straturi succesive de membrană plasmatică (săgeţile externe) sau poate creşte dinspre exterior spre interior (săgeţile interne). B-celulele Schwann sunt separate de un spaţiu îngust – nodul Ranvier. 1-celulă Schwann; 2-nucleul celulei Schwann; 3- axon; 4-teacă de mielină; 5-noduri Ranvier.
3
La mamifere, căile motorii se mielinizează mult mai târziu, în uter mişcările fătului fiind
reduse. La speciile la care puii încep să se deplaseze relativ independent la scurt timp după naştere, şi mielinizarea căilor motorii este mai timpurie. Acest lucru este valabil şi pentru păsările nidifuge, la care căile motorii se mielinizează mai repede decât la păsările nidicole. La om, fibrele motorii încep să-şi secrete teaca de mielină din a doua lună de viaţă extrauterină. Mai întâi începe mielinizarea căilor extrapiramidale, apoi a celor piramidale, procesul încheindu-se în jurul vârstei de 2 ani, când copilul are deja un mers sigur. Printre ultimii se mielinizează axonii nervilor ruşinoşi, care controlează sfincterele anal extern şi vezical extern.
Din punct de vedere al numărului de prelungiri, neuronii pot fi unipolari (cu o singură prelungire, axonică, lungă şi ramificată - neuronii de la nevertebrate), pseudounipolari (cu o singură prelungire, axonică, bifurcată - neuronii senzitivi din ganglionii spinali; unii autori îi consideră neuroni unipolari), bipolari (cu dendrita şi axonul ieşind din corpul celular la poli opuşi - celulele olfactive, neuronii bipolari din retină) sau multipolari (cu mai multe dendrite şi un singur axon - neuronii multipolari din retină, celulele mitrale din bulbii olfactivi, celulele piramidale Betz din cortexul motor, motoneuronii din coarnele anterioare medulare etc.). Unii neuroni sunt lipsiţi de dendrite tipice (neuronii senzitivi din ganglionii spinali), alţii nu au axon (majoritatea celulelor amacrine din retină).
Din punct de vedere funcţional, neuronii pot fi împărţiţi în 3 clase: neuroni aferenţi sau senzitivi, neuroni eferenţi sau motori şi neuroni intercalari. Neuronii aferenţi (lat. ad = la; ferre = a transporta) duc informaţia de la ţesuturile şi organele corpului la SNC, neuronii eferenţi (lat. ex = în afară) transmit semnale electrice de la SNC la celulele efectoare din muşchi şi glande, iar interneuronii fac legătura între primele două categorii.
Situaţi distal faţă de SNC, neuronii aferenţi sunt în legătură cu senzori (receptori) care, drept răspuns la variaţii mecanice, fizice sau chimice din mediul lor, determină ca în neuron să se producă semnale electrice. Senzorii pot conţine porţiuni specializate ale prelungirii neuronale sau pot fi alcătuiţi din celule receptoare de natură epitelială (epiteliu senzorial) aflate în relaţie funcţională de tip sinaptic cu prelungirea neuronului. Neuronii aferenţi conduc semnalele recepţionate de senzori de la periferia organismului spre nevrax. Aceşti neuroni sunt în majoritatea lor pseudounipolari, atipici prin faptul că au o singură prelungire, de obicei foarte lungă şi mielinizată, considerată a fi un axon. La mică distanţă după ieşirea din corpul celular, axonul se bifurcă. Porţiunea periferică vine în contact cu senzorul sau chiar îl formează; structural această porţiune este axon, deoarece este foarte lungă şi mielinizată, dar din punct de vedere funcţional este o dendrită, deoarece conduce impulsul nervos centripet (spre corpul celular) şi intră în componenţa senzorului. Porţiunea proximală a prelungirii este un axon tipic, ce intră în nevrax, unde stabileşte legături cu alţi neuroni. Corpul celular şi porţiunea periferică, lungă, a axonului se află în afara SNC şi numai o parte din capătul proximal pătrunde în măduvă sau în creier.
Corpul celular şi dendritele neuronilor eferenţi sunt găzduite în SNC, în timp ce axonii se prelungesc către periferie, unde stabilesc sinapse cu organele efectoare. Axonii neuronilor aferenţi şi eferenţi, exceptând o scurtă porţiune, formează nervii (sistemul nervos periferic).
Neuronii intercalari se găsesc în întregime în interiorul SNC. Ei reprezintă 99% din totalul neuronilor şi au o mare varietate de forme, proprietăţi fiziologice, mediatori chimici şi funcţii. Pentru fiecare neuron eferent (a cărui prelungire iese din SNC) există 10-20 neuroni aferenţi şi în jur de 200.000 de neuroni intercalari. Numărul de neuroni intercalari ce se interpun între anumiţi neuroni aferenţi şi eferenţi este diferit, în funcţie de complexitatea activităţii îndeplinite de respectiva reţea neuronală. De exemplu, arcul reflexului rotulian nu cuprinde interneuroni, axonul neuronului aferent (porţiunea centrală) se termină pe soma neuronului eferent. Spre deosebire de acesta, reflexele complexe implicate în vorbire sau în procesele de memorie pot cuprinde milioane de neuroni.
Caracteristicile celor 3 clase funcţionale de neuroni sunt rezumate în tabelul 1.1.
4
Tabelul 1.1. Clasele funcţionale de neuroni. Clasa funcţională de neuroni Proprietăţi
I. Neuroni aferenţi A. Transmit informaţii de la receptori la SNC. B. Majoritatea celulei (soma şi prelungirea lungă periferică) este în
afara SNC, doar prelungirea centrală, scurtă intră în nevrax. C. Din punct de vedere structural, nu au dendrite. Funcţia de
dendrită o îndeplineşte capătul ramificat al porţiunii distale a axonului.
II. Neuroni eferenţi A. Transmit informaţii de la SNC la organele efectoare (muşchi sau glande).
B. Soma, dendritele şi o mică porţiune din axon se află în SNC, iar cea mai mare parte a axonului este în afara SNC.
III. Neuroni intercalari A. Se află în întregime în interiorul SNC. B. Reprezintă 99% din totalul neuronilor.
1.2. Proprietăţile funcţionale ale neuronilor Funcţionarea sistemului nervos se bazează pe trei proprietăţi fundamentale ale neuronilor: a)
excitabilitatea; b) conductibilitatea; c) degenerarea şi regenerarea.
1.2.1. Excitabilitatea Citosolul are o compoziţie foarte diferită de a lichidului extracelular. De exemplu, concentraţia potasiului şi a fosfatului este mai mare în interiorul celulei decât în afara ei, în timp ce pentru sodiu, calciu şi clor situaţia este inversă. Aceste diferenţe sunt necesare funcţionării normale a multor enzime intracelulare; bunăoară, sintezele proteice de la nivelul ribozomilor necesită o concentraţie relativ ridicată de potasiu. Diferenţele de concentraţie sunt create şi menţinute de membrana plasmatică, prin stabilirea unei bariere de permeabilitate în jurul citosolului. Plasmalema are permeabilitate selectivă, care asigură păstrarea în celulă a substanţelor necesare, dintre care unele - în special proteinele - cu cost metabolic prea ridicat pentru a fi irosite, şi previne pătrunderea în interiorul celulei a substanţelor ce nu-i sunt utile. De asemenea, posedă mecanisme (canale ionice, pompe ionice, proteine transportoare) care ajută unele molecule să străbată bariera membranară în vederea îndeplinirii de către celulă a funcţiilor sale specifice. Excitabilitatea este proprietatea fundamentală a oricărei celule vii de a reacţiona la acţiunea unui stimul prin modificarea permeabilităţii ionice a membranei sale plasmatice şi generarea unui potenţial electric. La nivel de organ efector, reacţia de răspuns se traduce prin îndeplinirea unor funcţii specifice, ca secreţia glandulară, contracţia musculară etc. Toate celulele vii sunt excitabile, dar gradul lor de excitabilitate este adaptat structurii lor funcţionale. Celula cea mai excitabilă este neuronul, de rapiditatea sa de reacţie chiar la stimuli cu intensitate mică depinzând capacitatea de integrare şi coordonare a organismului. Urmează, în ordine, fibra musculară striată, muşchii scheletici trebuind să fie capabili de răspunsuri contractile rapide şi precis gradate la comenzile emise de sistemul nervos central, apoi celulele musculare de tip cardiac şi fibrele musculare netede. Structuri înalt excitabile sunt şi senzorii analizatorilor, precum şi alte celule cu rol de senzor grupate în zone strategice din interiorul organismului. Celule mai puţin excitabile, în care modificările de permeabilitate membranară sunt de mai mică amploare şi răspunsul specific al celulelor nu se bazează pe generarea unor potenţiale de acţiune, sunt hepatocitul şi celulele ţesutului conjunctiv, inclusiv cele osoase. Excitant sau stimul poate fi orice agent din mediu capabil să determine o reacţie din partea unei structuri vii (în fapt, orice variaţie a unei forme de energie din mediu, care la nivel celular capătă valoare informaţională). Pentru a fi investită cu calitatea de stimul, variaţia de energie trebuie să îndeplinească o serie de condiţii, adică să se supună legilor generale ale excitabilităţii:
l. Pentru a produce o reacţie din partea celulei, excitantul trebuie să aibă o anumită valoare minimă: intensitate-prag sau liminară.
2. Stimulii subliminari produc doar reacţii locale la nivel membranar, ce nu se pot transmite mai departe ca potenţiale de acţiune. Dar, dacă stimulii subliminari acţionează repetat şi cu o frecvenţă care să permită însumarea lor, ei pot genera o reacţie din partea celulei sub forma unui potenţial de acţiune.
5
3. La nivelul unei singure celule sau al unei unităţi funcţionale (neuron, unitate motorie, sinciţiu), un stimul liminar produce o reacţie maximală, iar un stimul superior ca intensitate (supraliminar) produce aceeaşi reacţie ca şi excitantul liminar - legea "tot sau nimic". Această lege nu mai este valabilă în cazul agregatelor supracelulare (de ex. muşchiul scheletic în ansamblul său).
4. Pentru a evita adaptarea structurilor vii faţă de variaţia unei forme de energie, stimulul trebuie să acţioneze cu o anumită bruscheţe pentru a produce o reacţie.
5. Excitarea repetată cu un stimul de aceeaşi natură atrage după sine modificarea temporară a pragului de excitabilitate al celulei faţă de acel stimul (adaptare).
6. Reacţia de răspuns la un stimul nu apare instantaneu, ci după un anumit interval de timp (în general foarte scurt – perioadă de latenţă) necesar desfăşurării proceselor fizico-chimice declanşate de stimul. Cu excepţia apei, componentele majore ale lichidului extracelular sunt ionii Na+ şi Cl-, în timp ce fluidul intracelular conţine concentraţii mari de K+, anioni proteici şi fosfaţi. Fenomenele ce rezultă din distribuţia acestor particule încărcate electric joacă un rol important în funcţionarea celulelor. Când particule cu încărcătură electrică opusă se află în acelaşi mediu (loc), forţa care le atrage una spre alta poate fi utilizată pentru a efectua un travaliu. Dimpotrivă, pentru a separa aceste particule, trebuie acţionat cu o forţă care să învingă atracţia dintre ele. Deci, sarcinile electrice de sens contrar au potenţialul de a efectua un travaliu, dacă se pot apropia una de alta. Acest potenţial poartă numele de potenţial electric sau, deoarece este determinat de diferenţa de sarcină dintre două puncte, diferenţă de potenţial - pe scurt, potenţial. Diferenţa de potenţial se măsoară în volţi (V), dar în sistemele biologice ea este foarte mică, de aceea se foloseşte ca unitate de măsură milivoltul (mV).
Potenţialul membranar de repaus În condiţii de repaus, toate celulele au o diferenţă de potenţial între faţa externă şi cea internă a membranei, cu faţa externă încărcată pozitiv şi faţa internă încărcată negativ. Acesta este potenţialul membranar de repaus (PMR), a cărui mărime variază între -5 şi -100 mV, în funcţie de tipul de celulă (în neuron, este de ordinul a -40 pînă la -75 mV). PMR al celulelor excitabile se poate modifica rapid ca răspuns la o excitaţie, această proprietate fiind esenţială pentru funcţionarea lor. PMR se caracterizează printr-un exces de ioni negativi în interiorul celulei şi un exces de ioni pozitivi în exterior. Sarcinile negative în exces din celulă, nefiind suficiente sarcini pozitive care să le neutralizeze, sunt atrase electric spre excesul de sarcini pozitive de la exterior, şi invers. Astfel, ionii în exces se adună într- un strat subţire la suprafaţa internă şi la cea externă a membranei plasmatice, în timp ce majoritatea mediului extracelular, precum şi cea mai mare parte a citosolului sunt practic neutre din punct de vedere electric (fig. 1.3). De precizat că numărul de sarcini pozitive şi negative care se aliniază de o parte şi de alta a membranei şi generează diferenţa de potenţial reprezintă o fracţiune infinitezimală din numărul total de sarcini electrice dintr-o celulă.
Fig. 1.3. Excesul de sarcini pozitive de la exteriorul celulei şi excesul de sarcini negative din interior se aglomerează în apropierea membranei plasmatice. În realitate, aceste sarcini în exces reprezintă doar o mică parte din numărul total de ioni de la exteriorul şi din interiorul celulei.
Mărimea PMR este determinată de următorii factori: (1) diferenţa în concentraţiile ionilor de o parte şi de alta a barierei membranare; (2) permeabilitatea diferită a membranei pentru diferite specii de ioni; (3) transportul pasiv (difuziune) al Na+ (influx) şi K+ (eflux) prin membrană, în sensul gradientului de concentraţie; (4) echilibrul Gibbs-Donnan; (5) osmoza; (6) transportul activ al ionilor prin pompa de Na+ / K+. Diferitele mecanisme de transport pasiv şi activ sunt astfel coordonate încât să menţină ionii şi alte substanţe intracelulare la concentraţii compatibile cu viaţa. În consecinţă concentraţiile ionilor din interiorul celulei nu se echilibrează cu cele din lichidul extracelular, ci există într-o stare staţionară diferită faţă de acesta. De exemplu, concentraţia Na+ intracelular (10 mM într-o celulă musculară) este mult mai scăzută decît a Na+ extracelular (140 mM), deci sodiul poate intra în
6
celulă prin transport pasiv, difuziune în sensul gradientului de concentraţie. Rata intrării sodiului este egalată de rata transportului activ al sodiului spre exteriorul celulei via pompa de Na+/K+. Astfel, concentraţia intracelulară a sodiului este menţinută la un nivel scăzut şi constant, deşi sodiul intră şi iese în permanenţă din celulă. Procesul este invers pentru ionii de potasiu, care sunt menţinuţi la o concentraţie ridicată în celulă, comparativ cu exteriorul. Ieşirea pasivă a potasiului este contrabalansată de intrarea activă prin pompa de Na+/K+ (fig. 1.4).
Fig. 1.4. Conceptul de stare staţionară. Rata intrării Na+ în celulă, în sensul gradientului electrochimic, este egalată de rata transportării Na+ spre exterior, prin activitatea pompei de Na+/K+. În acest fel, concentraţia intracelulară a Na+ nu se modifică. În mod similar, rata efluxului pasiv de K+ este egalată de rata introducerii active a K+ în celulă, prin aceeaşi pompă. Energia furnizată prin hidroliza unei molecule de ATP serveşte pentru deplasarea spre exterior a 3 ioni de sodiu şi introducerea în celulă a 2 ioni de potasiu.
Atât în interiorul celulei cât şi în lichidul extracelular există şi alţi ioni: Mg2+, Ca2+, H+, HC03-,
HP042-, S04
2-, grupări amino şi carboxil ale aminoacizilor, anioni proteici mari; dar sodiul, potasiul şi clorul sunt prezenţi în cele mai mari concentraţii şi deci joacă rolul hotărâtor în generarea PMR. Concentraţiile ionilor de Na+, K+ şi Cl- în mediul extracelular şi în citosolul unei celule nervoase sunt prezentate în tabelul 1.2. Tabelul 1.2. Distribuţia celor mai importanţi ioni de o parte şi de alta a membranei plasmatice
a unei celule nervoase
Ionul Concentraţia (mM) Extracelular Intracelular
K+ 5 150 Na+ 150 15 Cl- 110 10
Aşa cum am mai arătat, membrana plasmatică este selectiv permeabilă, lăsând să treacă numai anumiţi ioni sau molecule şi numai în anumite proporţii. Această proprietate poate fi caracterizată prin coeficientul de permeabilitate membranară, particular fiecărei specii chimice (tabelul 1.3.).
Tabelul 1.3. Coeficienţii de permeabilitate pentru principalii ioni implicaţi în realizarea
potenţialului membranar de repaus; Pr- = anioni proteici. Ionul Coeficientul de permeabilitate membranară
Na+ 0,1 K+ 1,0 Cl - 2,0 Pr - 0
Se observă că, pentru fiecare ion de sodiu care intră în celulă, 10 ioni de potasiu părăsesc
celula şi 20 de ioni de clor intră, în timp ce anionii mari proteici nu pot părăsi celula. Concentraţia mare a sodiului în lichidul extracelular, precum şi deplasarea ionilor de potasiu în sensul gradientului lor de concentraţie conferă electropozitivitate feţei externe a membranei; anionii proteici, împreună cu influxul de clor conferă electronegativitate feţei interne a membranei. 7
Datorită încărcăturii electrice diferite a celor două feţe ale membranei, se stabileşte şi un gradient electric care, alături de gradientul de concentraţie, influenţează deplasarea ionilor. În cazul sodiului, ce se deplasează de la exteriorul membranei (electropozitiv) spre interior (electronegativ), gradientul electric facilitează trecerea ionilor prin membrană. Potasiul în schimb, se deplasează de la interior spre exterior, în sensul gradientului de concentraţie, dar în sens invers gradientului electric.
Potenţialul membranar la care forţa gradientului electric este egală şi de sens contrar cu forţa gradientului de concentraţie este numită potenţial de echilibru pentru acel ion. La potenţialul de echilibru nu există nici o deplasare a ionului prin membrană, cele două forţe egale anulându-se reciproc.
Difuziunea ionilor prin membrană, guvernată de gradientul electrochimic şi de permeabilitatea selectivă, este influenţată şi de prezenţa, în interiorul celulei, a anionilor mari proteici, care nu pot străbate membrana, dar conferă electronegativitate feţei sale interne. Datorită anionilor proteici, între interiorul celulei şi micromediul extern se stabileşte un echilibru cunoscut sub numele de echilibrul Gibbs-Donnan, caracterizat printr-o distribuţie inegală a ionilor difuzibili între două soluţii separate de o membrană, impermeabilă pentru cel puţin o specie ionică (formată din ioni prea mari pentru a străbate membrana).
Pentru a înţelege acest fenomen, să considerăm două compartimente lichidiene, separate printr-o membrană permeabilă pentru apă şi electroliţi, dar total impermeabilă pentru o specie ionică (anionii proteici) (fig. 1.5.). Presupunem că, iniţial, în ambele compartimente există doar apă; dacă în compartimentul I se adaugă KCl, K+ şi Cl- vor difuza în compartimentul II, până când concentraţiile lor vor fi egale în cele două compartimente (fig. 1.5a). Dacă în compartimentul I este adăugată sarea de potasiu a unui anion proteic (KA), prezenţa acestuia va modifica echilibrul ionic de o parte şi de alta a membranei (fig. 1.5b): va atrage în compartimentul I mai mulţi ioni pozitivi (K+) şi mai puţini ioni negativi (Cl-). Astfel, se va menţine electroneutralitatea în fiecare compartiment, dar concentraţiile ionilor difuzibili nu vor mai fi egale în cele două compartimente. Aceste concentraţii pot fi exprimate algebric, după cum se observă în fig. 1.5.c.
(c)
Fig. 1.5. Echilibrul Gibbs-Donnan. (a) – KCl adăugat în compartimentul I difuzează în compartimentul II, până la egalizarea concentraţiilor celor doi ioni; (b) - prezenţa unui anion nedifuzibil în compartimentul I va modifica raportul de concentraţii al ionilor difuzibili; (c) – noile concentraţii ionice stabilite pot fi exprimate algebric.
La un PMR de -70 mV, o valoare caracteristică neuronului, nici sodiul şi nici potasiul nu se află la potenţialul lor de echilibru, deci există un continuu eflux de K+ şi influx de Na+. Cu toate acestea, concentraţia sodiului în interiorul celulei şi a potasiului în lichidul extracelular nu creşte, deoarece sodiul pătruns în celulă şi potasiul ieşit la exterior sunt permanent trimişi înapoi în compartimentele lor printr-un mecanism activ de transport membranar - pompa de Na+/K+ (fig. 1.6.).
8
Fig. 1.6. Pompa de Na+/K+. pentru fiecare moleculă de ATP consumată, 3 ioni de sodiu sunt expulzaţi şi 2 ioni de potasiu sunt introduşi în celulă.
Acest mecanism are o dublă contribuţie la realizarea PMR: 1) indirectă, prin menţinerea concentraţiilor Na+ şi K+ de o parte şi de alta a membranei; 2) directă, prin faptul că numărul ionilor de sodiu şi de potasiu transportaţi prin membrană nu este egal. Pompa de Na+/K+ funcţionează ca o pompă electrogenă, deoarece scoate din celulă 3 Na+ pentru 2 K+ pe care-i introduce. Acest transport inegal face ca valoarea PMR să fie mai mare decît cea rezultată exclusiv în urma difuziunii ionilor în sensul gradientului de concentraţie. În majoritatea celulelor contribuţia directă, electrogenă a pompei de Na+/K+ la realizarea PMR este mică. Pe lângă sodiu şi potasiu, ionii de clor sunt prezenţi în cantitate mare, mai ales în lichidul extracelular. Membrana plasmatică a majorităţii celulelor este permeabilă pentru Cl-, dar nu prezintă pompe ionice pentru acesta. Potenţialul membranar stabilit de interacţiunile ionice descrise până acum va determina şi deplasarea Cl-. Electronegativitatea din interiorul celulei este forţa care va determina deplasarea Cl- spre exterior, pînă la apariţia unui gradient de concentraţie (concentraţie mai mică de Cl- în interiorul celulei decât la exterior). Forţa gradientului de concentraţie va produce difuzarea Cl- înapoi în celulă, opunându-se astfel forţei de gradient electric. Rezultatul este că potenţialul de echilibru pentru Cl- este egal cu PMR şi Cl- nu contribuie la valoarea potenţialului de membrană. În neuroni, majoritatea sarcinilor negative din interiorul celulei nu se datorează Cl-, ci moleculelor organice, mai ales anionilor mari proteici, care nu pot străbate membrana plasmatică. O altă modalitate de a ne referi la o membrană încărcată electric este să spunem că membrana este polarizată electric, adică are un potenţial diferit de zero. Dacă PMR este punctul de referinţă faţă de care măsurăm modificările de potenţial membranar, trecerea la o stare mai polarizată (cu interiorul membranei mai electronegativ şi exteriorul mai electropozitiv) o numim hiperpolarizare, iar trecerea la o stare mai puţin polarizată (mai aproape de 0 mV) o numim depolarizare (fig. 1.6). Modificări ale potenţialului membranar pot fi produse prin alterarea temporară a permeabilităţii membranei pentru anumiţi ioni, sub influenţa unui stimul.
Canalele ionice Implicate în producerea PA, ele poartă numele de canale voltaj-dependente, deoarece devin operante când se produc modificări ale valorii potenţialului membranar (modificări de voltaj). Deşi există deosebiri între proprietăţile canalelor de Na+ şi de K+ din membrana celulelor nervoase (spre exemplu, canalele de Na+ se deschid mai rapid decât cele de K+ şi se închid instantaneu după un anumit interval de timp, independent de starea de polarizare a membranei, în timp ce canalele de K+ se deschid mai încet şi nu se inactivează spontan, ci continuă să rămână deschise câtă vreme membrana este depolarizată), ambele tipuri de canale sunt considerate canale voltaj-dependente rapide. Vom vedea, la studierea excitabilităţii miocardului, că există în unele celule şi canale voltaj-dependente
9
lente, a căror acţiune, combinată cu cea a canalelor rapide, duce la prelungirea perioadei de depolarizare, ce ia forma unui platou, imediat după atingerea vârfului PA. În stare de repaus menţinerea PMR, fără apariţia unor variaţii de voltaj, se datorează difuziunii pasive a ionilor prin alt tip de canale, numite canale ionice de scurgere ("leaking" channels). Trecerea ionilor prin aceste canale este determinată de forţele descrise la subcapitolul privind PMR. Unele membrane plasmatice posedă şi un alt tip de canale, ce se deschid în urma legării unei molecule proteice (mediator chimic, hormon) numită ligand la o moleculă receptor ataşată canalului, cu formarea unui complex ligand-receptor. Este cazul membranelor neuronale postsinaptice şi al membranelor celulelor-ţintă pentru diferite specii de hormoni. Asemenea canale poartă numele de canale ligand-dependente (ligand-gated); un exemplu îl reprezintă canalele membranelor postsinaptice ale sinapselor avînd ca mediator chimic acetilcolina (Ach). Legarea Ach la un receptor membranar determină deschiderea canalelor ligand-dependente, sub forma unui por cu diametrul de 0,65 nm, care permite ionilor pozitivi cu diametru mai mic decît al porului să pătrundă în celulă. Canalele mecano-dependente sunt deschise de o modificare mecanică (tracţiune) la nivelul membranei plasmatice. Astfel de canale sunt prezente în membranele cililor celulelor senzoriale auditive şi vestibulare, excitate prin îndoirea cililor.
Fig. 1.6. Hiperpolarizarea şi depolarizarea. Potenţialele de echilibru pentru Na+ şi pentru K+ (ENa= +60 mV; EK= -90 mV) sunt prezentate în relaţie cu PMR (-70 mV) şi cu starea nepolarizată a membranei (punct izoelectric = 0 mV).
Potenţialul de acţiune (PA) Potenţialul de acţiune este o modificare locală (apare la locul de acţiune al unui stimul asupra membranei celulare) şi rapidă a potenţialului membranar, cu durată redusă (1 ms pentru neuron), timp în care potenţialul se poate modifica cu 100 mV şi membrana se poate repolariza, refăcând PMR. Numai celulele nervoase, musculare şi unele celule glandulare au membrane plasmatice capabile de a produce PA. Aceste membrane se numesc excitabile, iar proprietatea lor de a produce PA se numeşte excitabilitate. În cadrul sistemului nervos, informaţia poate circula în organism la distanţe mari, sub forma PA (impuls nervos).
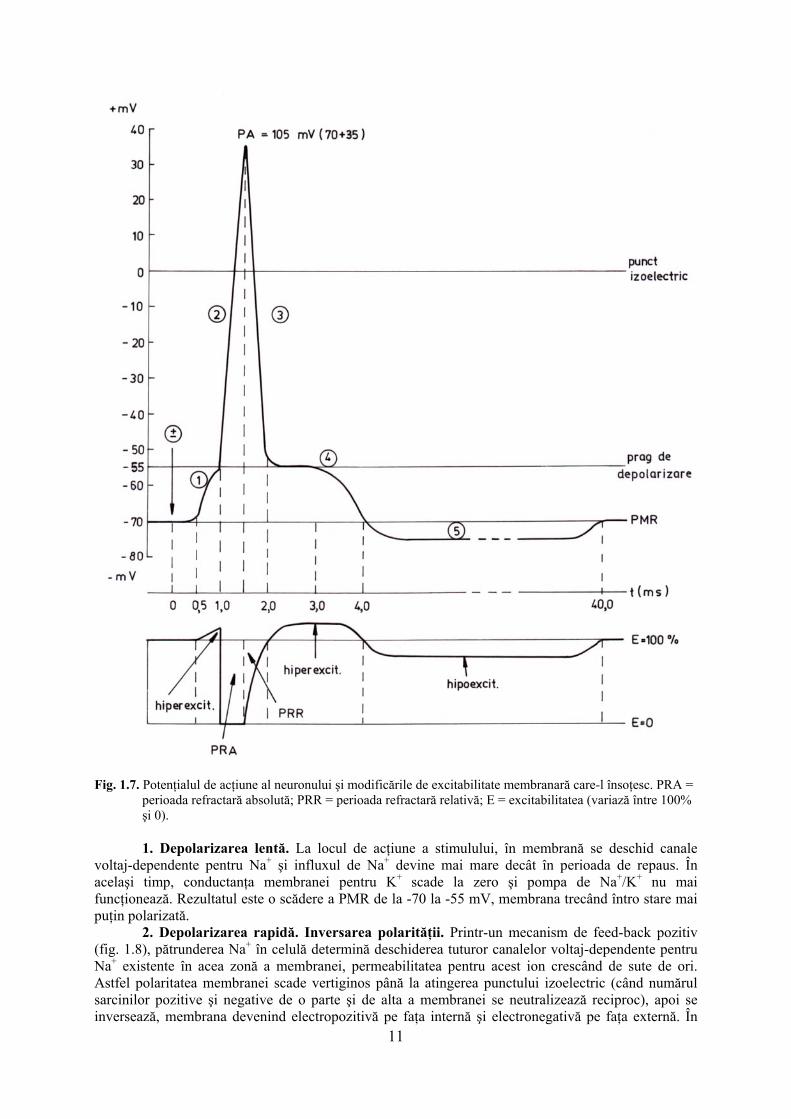
Bazele ionice ale generării PA. Mecanismele modificării de permeabilitate membranară. În cazul formării PA, apare o modificare temporară a permeabilităţii membranare, în timp ce gradientul de concentraţie rămîne neschimbat şi dirijează sensul de deplasare al ionilor. În stare de repaus, în membrana celulei sunt deschise mai ales canale de K+ şi de Cl- şi doar foarte puţine canale de Na+. Din această cauză PMR este mai apropiat de potenţialul de echilibru al potasiului decât de cel al sodiului. În timpul generării unui PA şi apoi a revenirii la polaritatea de repaus a membranei, distingem două evenimente majore - depolarizarea şi repolarizarea - care se petrec în mai multe etape (fig. 1.7).
10
Fig. 1.7. Potenţialul de acţiune al neuronului şi modificările de excitabilitate membranară care-l însoţesc. PRA = perioada refractară absolută; PRR = perioada refractară relativă; E = excitabilitatea (variază între 100% şi 0).
1. Depolarizarea lentă. La locul de acţiune a stimulului, în membrană se deschid canale
voltaj-dependente pentru Na+ şi influxul de Na+ devine mai mare decât în perioada de repaus. În acelaşi timp, conductanţa membranei pentru K+ scade la zero şi pompa de Na+/K+ nu mai funcţionează. Rezultatul este o scădere a PMR de la -70 la -55 mV, membrana trecând întro stare mai puţin polarizată.
11
2. Depolarizarea rapidă. Inversarea polarităţii. Printr-un mecanism de feed-back pozitiv (fig. 1.8), pătrunderea Na+ în celulă determină deschiderea tuturor canalelor voltaj-dependente pentru Na+ existente în acea zonă a membranei, permeabilitatea pentru acest ion crescând de sute de ori. Astfel polaritatea membranei scade vertiginos până la atingerea punctului izoelectric (când numărul sarcinilor pozitive şi negative de o parte şi de alta a membranei se neutralizează reciproc), apoi se inversează, membrana devenind electropozitivă pe faţa internă şi electronegativă pe faţa externă. În
această fază se dezvoltă vârful (spike-ul) PA, care ajunge, în celula nervoasă, la aproximativ +30 mV, aproape de valoarea potenţialului de echilibru al sodiului. Declanşarea depolarizării rapide este dependentă de atingerea pragului de depolarizare, care pentru neuron este minim în membrana conului axonic, la -55 mV. În celula nervoasă, PA durează aproximativ 1 ms, dar poate fi mult mai lung în anumite tipuri de celule musculare.
Fig. 1.8. Relaţia de feed-back pozitiv dintre depolarizarea membranei şi creşterea permeabilităţii pentru sodiu.
3. Repolarizarea rapidă. Potenţialul membranar revine rapid aproape de nivelul său de
repaus, datorită: (a) închiderii canalelor pentru Na+ care au fost deschise pe durata depolarizării rapide; astfel sodiul pătruns în etapa precedentă rămâne prizonier în celulă; (b) deschiderii canalelor voltaj-dependente pentru K+. Astfel efluxul masiv de K+ reface rapid electropozitivitatea feţei externe a membranei. 4. Repolarizarea lentă (postrepolarizarea sau postpotenţialul negativ, după denumiri mai vechi) apare ca o întârziere a repolarizării şi se datorează excesului de sarcini pozitive ale ionilor de sodiu sechestraţi în celulă prin închiderea canalelor pentru sodiu. Acesta nu poate fi compensat în totalitate prin ieşirea K+, a cărui conductanţă a atins deja valoarea maximă şi începe să scadă. Această etapă durează 2-3 ms. 5. Posthiperpolarizarea (postpotenţialul pozitiv) este o creştere a diferenţei de potenţial transmembranar peste valoarea PMR, putând ajunge pînă la -80 mV. În condiţiile în care canalele pentru Na+ continuă să fie închise, iar o parte a canalelor pentru potasiu deschise, reintră în funcţie pompa de Na+/K+, care scoate activ Na+ din celulă. K+ difuzează prin canale la exterior şi este reintrodus în celulă de pompă, în timp ce Na+ nu poate pătrunde în celulă, este doar scos. Astfel în interiorul celulei se creează un deficit temporar de sarcini pozitive, ceea ce determină hiperpolarizarea membranei pentru o durată de câteva zeci de ms. Ca urmare a implicării, în această etapă, a mecanismului de transport activ prin pompa ionică, se înregistrează o creştere a consumului energetic al celulei excitabile, însoţită de eliberare de căldură. Pe măsură ce pompa ionică scoate Na+ şi reintroduce K+ în celulă, permeabilitatea membranei revine la normal şi la sfârşitul posthiperpolarizării potenţialul transmembranar se restaurează la valoarea PMR (-70 mV, pentru celula nervoasă). Modificările excitabilităţii membranare în timpul generării PA
În perioada de repaus, când nici un stimul nu acţionează asupra membranei, aceasta prezintă o diferenţă de potenţial transmembranar de -70 mV şi o excitabilitate de 100 % (normală). Orice stimul liminar sau supraliminar va modifica permeabilitatea membranei pentru ioni, va deschide canalele voltaj-dependente pentru Na+ şi va depolariza membrana, generând un PA. La un nou stimul, venit în timpul producerii acestui PA, membrana se va comporta în mod diferit, în funcţie de etapa în care se află PA şi de intensitatea noului stimul (fig. 1.7). În faza (1) de depolarizare lentă, excitabilitatea membranară creşte uşor peste normal (hiperexcitabilitate), fenomen datorat, în unele cazuri, sumaţiei mai multor stimuli subliminari iar în alte cazuri unui singur stimul, de valoare prag sau peste prag. În etapa (2) de depolarizare rapidă, excitabilitatea membranei scade brusc la zero; acum sunt deschise toate canalele voltaj-dependente pentru sodiu din zona membranară excitată, există un influx masiv de Na+, astfel încât membrana nu mai are resurse pentru a răspunde la un nou stimul care ar
12
acţiona în această perioadă. Aceasta este perioada refractară absolută (PRA). Importanţa PRA constă în aceea că ea determină numărul de PA care pot fi generate în unitatea de timp într-o membrană excitabilă (labilitatea). Cu cât PRA este mai scurtă, labilitatea neuronului creşte. Majoritatea celulelor nervoase pot produce până la 100 PA/s, iar unele pot lucra, pentru perioade scurte de timp, chiar la frecvenţe mai mari. În faza (3) de repolarizare rapidă, excitabilitatea membranei creşte treptat, pe măsură ce repolarizarea avansează. Canalele pentru Na+ s-au închis în totalitate în momentul de vârf al PA, au stat închise un foarte scurt interval şi acum sunt disponibile pentru a genera un nou PA. Este perioada refractară relativă (PRR), în care un stimul prag nu produce PA, dar un stimul supraliminar are această posibilitate. PRR corespunde cu permeabilitatea maximă a membranei pentru K+ (eflux masiv al acestui ion); un stimul supraliminar poate deschide mai multe canale pentru Na+, astfel încât influxul de sodiu să întreacă efluxul de potasiu şi să permită intrarea în funcţie a mecanismului de feed-back pozitiv descris anterior. Faza (4) de repolarizare lentă (postrepolarizarea) este contemporană cu o uşoară creştere a excitabilităţii peste valoarea normală (hiperexcitabilitate). Acum în celulă există un exces de sarcini pozitive (Na+) rămase aici în urma închiderii canalelor pentru sodiu, astfel încât chiar un stimul aflat cu puţin sub pragul de excitabilitate poate produce un PA.
În timpul posthiperpolarizării (5) care urmează, excitabilitatea membranei scade uşor sub valoarea normală (hipoexcitabilitate) din cauza deficitului de sarcini pozitive de pe faţa internă a membranei, produs de reintrarea în funcţie a pompei de Na+/K+. Odată cu refacerea PMR, şi excitabilitatea membranei revine la normal.
Potenţiale locale Curenţi catodici şi anodici. Electrotonusul.
În experimente efectuate cu curent galvanic (continuu), Pflüger a demonstrat că într-o celulă nervoasă traversată de un curent subliminar se produc modificări de excitabilitate. Să considerăm doi electrozi, unul negativ (catod) şi altul pozitiv (anod), alimentaţi de la o sursă de curent continuu şi plasaţi pe suprafaţa unei celule excitabile, de exemplu pe axolema unui neuron (fig. 1.9a). Trebuie reamintit că simpla trecere a curentului electric prin electrozi nu produce nici o reacţie din partea membranei celulare, ea fiind sensibilă doar la variaţii ale energiei electrice; stimularea se poate produce numai la închiderea sau la deschiderea circuitului. La trecerea prin circuit a unui curent electric slab, cu valoare de stimul subliminar, nu este iniţiat un PA, dar excitabilitatea membranei se modifică în dreptul celor doi electrozi. La catod are loc o acumulare de sarcini negative la suprafaţa membranei, ceea ce duce la scăderea valorii PMR şi deci a pragului de depolarizare. Fenomenul poartă numele de catelectrotonus şi determină o hiperexcitabilitate locală. La anod are loc o acumulare de sarcini pozitive la suprafaţa membranei, ceea ce accentuează polarizarea ei şi duce la creşterea pragului de depolarizare. Acest anelectrotonus determină o hipoexcitabilitate locală.
Potenţialele electrotonice, fiind produse de stimuli electrici subliminari, au toate caracteristicile unor potenţiale locale; ele nu iniţiază PA dar "pregătesc" membrana celulară pentru acţiunea unui stimul liminar modificându-i, într-un sens sau altul, capacitatea de a reacţiona. Astfel se explică de ce, la acţiunea unor stimuli cu intensitate crescândă, PA apare mai repede la catod, în zona devenită hiperexcitabilă şi mai târziu (doar la stimuli mai puternici) la anod, în zona de hipoexcitabilitate (fig. 1.9b).
13
Fig. 1.9. Punerea în evidenţă a potenţialelor electrotonice (a) şi relaţia dintre amplitudinea potenţialului şi intensitatea stimulului (IS), la catod (C) şi la anod (A), în cazul aplicării de stimuli subliminari (b). SE – înregistrarea semnalului electric (închiderea, respectiv deschiderea circuitului). PL – potenţialele locale rezultate în urma stimulării.
Potenţiale locale Potenţialele locale (PL) sunt modificări ale potenţialului membranar care privesc doar o mică
regiune a membranei şi „mor” la 1-2 mm distanţă de locul de origine. Ele pot apărea fie sub forma unei depolarizări, fie sub forma unei hiperpolarizări şi sunt produse de stimuli subliminari. Se mai numesc şi "potenţiale gradate" pentru că au amplitudine variabilă, proporţională cu intensitatea stimulului. Curentul local "curge" aşa cum curge apa de-a lungul unui furtun fisurat. Se pierd sarcini de-a lungul membranei pentru că membrana este în continuare permeabilă pentru ioni, iar rezultatul este descreşterea curentului cu distanţa. Altfel spus, propagarea PL se face decremental. La apariţia unui PL, va exista un schimb de sarcini electrice între locul său de origine şi regiunile adiacente ale membranei, aflate în stare de repaus. Astfel, PL creează curent electric în lichidul extracelular şi în fluidul intracelular, curenţi care sunt proporţionali cu amplitudinea potenţialului. Prin convenţie, direcţia de deplasare a sarcinilor pozitive este considerată direcţia de deplasare a curentului. Pe faţa internă a membranei, sarcinile pozitive se vor depărta de locul excitării, spre zone mai negative, caracteristice PMR. Pe faţa externă a membranei, sarcinile pozitive se vor îndrepta dinspre regiunile pozitive adiacente spre locul de excitare, mai electronegativ. Acest curent local scade gradul de separare a sarcinilor (depolarizare) în regiunile din jurul zonei uşor depolarizate prin excitarea cu stimulul subliminar. Curentul local este purtat de ioni ca K+, Na+, Cl- şi HC03
-. Întrucât PL nu prezintă prag de depolarizare (depolarizările locale pot fi produse de stimuli mult mai mici decât un stimul liminar) şi nici perioadă refractară (frecvenţa de acţiune a stimulilor poate fi foarte mare, deoarece modificările în încărcătura membranară sunt mici şi nu necesită mecanisme speciale pentru restaurarea stării de repaus), ele se pot însuma şi pot atinge astfel pragul de depolarizare necesar pentru generarea unui PA (fig. 1.10). Sumaţia poate fi temporală, dacă în aceeaşi regiune a membranei se aplică succesiv, cu frecvenţă mare, stimuli subliminari, sau spaţială, dacă se aplică simultan stimuli subliminali în puncte foarte apropiate ale membranei.
Proprietăţile PL şi PA sunt prezentate comparativ în tabelul 1.2. După localizare sau după funcţia pe care o îndeplinesc, distingem mai multe tipuri de PL: - potenţiale de receptor, care se nasc, sub influenţa stimulilor specifici, în celulele receptoare
ale analizatorilor şi pot fi potenţiale generatoare (PG) sau receptoare (PR), după cum celula receptoare este un neuron sau o celulă epitelială senzorială;
- potenţiale electrotonice (catelectrotonus şi anelectrotonus); - potenţiale postsinaptice, care apar în membrana postsinaptică şi pot fi excitatoare (PPSE) sau
inhibitoare (PPSI), după cum se produc prin depolarizarea, respectiv prin hiperpolarizarea membranei; - un tip special de potenţial postsinaptic este potenţialul local terminal de placă (PLTP),
generat la membrana postsinaptică a plăcii motorii;
14
Fig. 1.10. La catod se pot genera PA prin însumarea unor potenţialelor locale produse de stimuli subliminali ce nu depăşesc 80% din intensitatea stimulului prag. La anod, însumarea acestor PL produce o stare de hiper-polarizare a membranei. IS – intensitatea stimulilor subliminali, ca procente din intensitatea stimulului prag.
- potenţialele produse în mod spontan, în absenţa unui excitant extern, de celulele pacemaker generatoare de automatism din ţesutul nodal excitoconductor al inimii, de unele celule musculare ale intestinului subţire sau de neuroni din centrii inspiratori bulbari.
Tabelul 1.4. Proprietăţile PL şi PA Potenţiale locale Potenţiale de acţiune
1. Amplitudinea este proporţională cu intensitatea stimulului. Nu se supun legii "tot sau nimic".
l. Amplitudinea este maximă (independentă de intensitatea stimulului) dacă a fost atins pragul de depolarizare. Se supun legii "tot sau nimic".
2. Nu au prag de depolarizare. 2. Au un prag de depolarizare de 10-15 mV. 3. Nu au perioadă refractară. 3. Au perioadă refractară, absolută şi relativă. 4. Se pot însuma. 4. Nu se pot însuma. 5. Sunt conduse decremental, amplitudinea scade cu distanţa.
5. Sunt conduse nedecremental, amplitudinea rămâne constantă.
6. Durata variază în funcţie de condiţiile iniţiale. 6. Durata este constantă, specifică tipului celular. 7. Pot fi depolarizări sau hiperpolarizări. 7. Sunt depolarizări. 8. Pot fi iniţiate de stimuli din mediu (în celule receptoare), de mediatori chimici (la nivelul sinapselor) sau spontan.
8. Sunt iniţiate de depolarizarea membranei până la pragul de depolarizare.
9. Codifică intensitatea stimulului în amplitudine. 9. Codifică intensitatea stimulului în frecvenţă. Importanţa fiziologică a PL este deosebită, deoarece reprezintă, la nivelul sinapselor, unica modalitate de comunicare între celulele excitabile şi creează, prin sumaţie, condiţiile necesare iniţierii
























![P] ]v } µo }v µ ]]o} ]vuµvX Z] ]vµ ] SECTORULUI ... · 2 8. 1 ö 1 al Monumentelor Istorice 9. £ 1 ò & 1 1 1 1 1 1 ©1 1](https://static.documente.net/doc/80x56/5fb7a327e1e27656a034c5c9/p-v-o-v-o-vuvx-z-v-sectorului-2-8-1-1-al-monumentelor.jpg)






