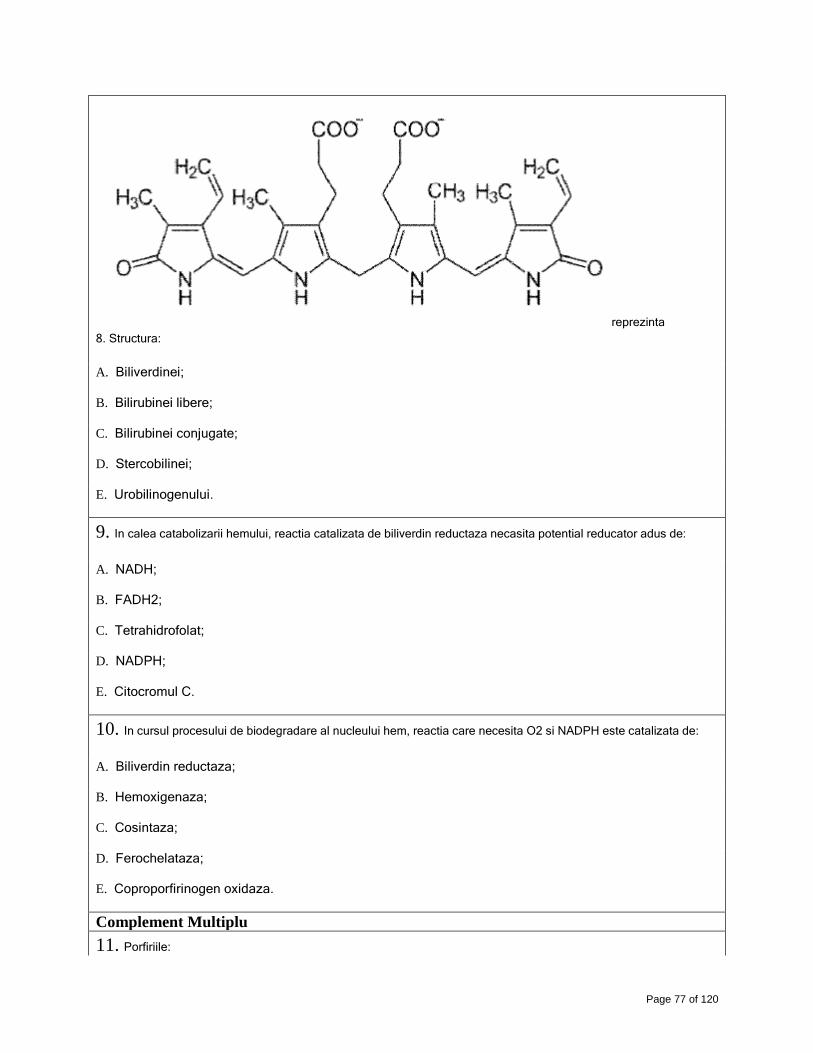
Page 1 of 120 Biochimie medicala, generala si aplicata, Anul III Farmacie, sem. II - (14) Capitolul 1. Catena respiratorie si fosforilarea oxidativa: componente, functionare, control, inhibitie, aspecte clinice Complement Simplu 1. Afirmatiile de mai jos sunt adevarate, cu exceptia: A. Catena respiratorie cuprinde trei complexe multienzimatice cu rol de pompe de protoni; B. Catena respiratorie cuprinde patru complexe multienzimatice; C. Catena respiratorie cuprinde patru complexe enzimatice cu rol de pompe de protoni; D. Una din pompele de protoni ale catenei respiratorii este NADHQ-reductaza; E. Una din pompele de protoni ale catenei respiratorii este citocrom oxidaza. 2. Ubiquinona (coenzima Q 10) este: A. Un transportor de electroni cu molecula lipofila; B. O proteina cu molecula mica, avand 10 nuclee hem; C. O molecula cu nucleu antrachinonic; D. O enzima care descompune peroxidul de hidrogen; E. Un complex multienzimatic al catenei respiratorii. 3. Afirmatia adevarata este: A. Decuplarea fosforilarii de oxidare nu mai permite trecerea fluxului de electroni prin catena respiratorie; B. Viteza transportului de electroni prin catena respiratorie creste atunci cand raportul ATP/ADP din mitocondrie creste; C. Transportorii mobili ai catenei respiratorii sunt ubiquinona si citocromul C; D. Complexele multienzimatice ale catenei respiratorii sunt codificate in totalitate la nivelul ADN nuclear; E. Gruparea prostetica a citocromilor este nucleul corinic cu ion Fe2+ complexat in centru. 4. Decuplarea lantului respirator de fosforilarea oxidativa presupune ca:
Transcript
Page 1 of 120
Biochimie medicala, generala si aplicata, Anul III
Farmacie, sem. II - (14)
Capitolul 1. Catena respiratorie si fosforilarea oxidativa: componente,
functionare, control, inhibitie, aspecte clinice
Complement Simplu
1. Afirmatiile de mai jos sunt adevarate, cu exceptia:
A. Catena respiratorie cuprinde trei complexe multienzimatice cu rol de pompe de protoni;
B. Catena respiratorie cuprinde patru complexe multienzimatice;
C. Catena respiratorie cuprinde patru complexe enzimatice cu rol de pompe de protoni;
D. Una din pompele de protoni ale catenei respiratorii este NADHQ-reductaza;
E. Una din pompele de protoni ale catenei respiratorii este citocrom oxidaza.
2. Ubiquinona (coenzima Q 10) este:
A. Un transportor de electroni cu molecula lipofila;
B. O proteina cu molecula mica, avand 10 nuclee hem;
C. O molecula cu nucleu antrachinonic;
D. O enzima care descompune peroxidul de hidrogen;
E. Un complex multienzimatic al catenei respiratorii.
3. Afirmatia adevarata este:
A. Decuplarea fosforilarii de oxidare nu mai permite trecerea fluxului de electroni prin catena respiratorie;
B. Viteza transportului de electroni prin catena respiratorie creste atunci cand raportul ATP/ADP din mitocondrie creste;
C. Transportorii mobili ai catenei respiratorii sunt ubiquinona si citocromul C;
D. Complexele multienzimatice ale catenei respiratorii sunt codificate in totalitate la nivelul ADN nuclear;
E. Gruparea prostetica a citocromilor este nucleul corinic cu ion Fe2+ complexat in centru.
4. Decuplarea lantului respirator de fosforilarea oxidativa presupune ca:
Page 2 of 120
A. Respiratia continua fara sinteza de ATP;
B. Mitocondria utilizeaza NADPH in loc de NADH;
C. Sinteza ATP continua dar respiratia inceteaza;
D. Nu are loc hidroxilarea unora din substrate;
E. Mitocondria utilizeaza ca donor de potential reducator citratul.
5. Afirmatia adevarata este:
A. Orice citocrom contine cupru;
B. Toti componentii catenei respiratorii care contin fier sunt hemoproteine;
C. Complexul enzimatic I al catenei respiratorii transfera electroni complexului II prin intermediul coenzimei Q;
D. Ubiquinona este transportorul de electroni mobil al Citocrom reductazei;
E. Ubiqiunona nu poate transporta electroni preluati de la FADH2.
6. Viteza de functionare a catenei respiratorii mitocondriale este reglata prin:
A. Scoaterea ATP din matricea mitocondriala;
B. Concentratii mari de glucoza in matricea mitocondriala;
C. Concentratii mari de colesterol in matricea mitocondriala;
D. Concentratii mari de glicerol in matricea mitocondriala:
E. Formarea de CO in ultima etapa a transportului de electroni prin catena.
7. Acumularea de mutatii somatice la nivelul ADN mitocondrial:
A. Constituie cauza declansatoare a neuropatiei optice ereditare Leber;
B. Constituie cauza declansatoare a encefalo-miopatiei insotite de acidoza lactica si convulsii;
C. Constituie cauza declansatoare sau favorizanta a unor maladii degenerative;
D. Constituie cauza decuplarii oxidarii de fosforilare;
E. Constituie cauza generarii gradientului de pH intre fata interna si cea externa a membranei mitocondriale.
8. Complexul III al catenei respiratorii:
A. Asigura transferul echivalentilor reducatori de la NADH,H+ la ubiquinona;
Page 3 of 120
B. Transfera echivalenti reducatori de la ubiquinona la citocromul c;
C. Transfera echivalenti reducatori de la succinat la ubiquinona;
D. Asigura sinteza ATP din ADP;
E. Nu este o pompa de protoni.
9. Complexul II al catenei respiratorii:
A. Este o pompa de protoni;
B. Preia potentialul reducator generat prin oxidarea succinatului in fumarat in ciclul acidului citric;
C. Contine nuclee hem C1 ca grupari prostetice;
D. Se mai numeste si citocrom reductaza;
E. Transfera potentialul reducator citocromului C.
10. Curgerea a 2 perechi proton-electron de la NADH,H+ la ubiquinona prin intermediul NADHQreductazei conduce
la:
A. Scoaterea a 2 protoni in afara matricei mitocondriale;
B. Reducerea completa a moleculei de oxigen;
C. Scoaterea a 4 protoni in afara matricei mitocondriale;
D. Oxidarea citocromului C;
E. Nici o varianta corecta.
Complement Multiplu
11. Componentele lantului mitocondrial transportor de electroni sunt:
A. NADH-Q reductaza;
B. Succinat –Q reductaza;
C. Fosfo fructo kinaza;
D. Citocrom-reductaza;
E. Citocrom-oxidaza.
12. Complexele enzimatice cu rol de pompe de protoni ale lantului mitocondrial transportor de electroni sunt:
A. NADH-Q reductaza;
B. Succinat-Q reductaza;
Page 4 of 120
C. Fosfofructokinaza;
D. Citocrom-reductaza;
E. Citocrom-oxidaza
13. ATP-sintaza mitocondriala este:
A. Un complex enzimatic cu rol de pompa de protoni din cadrul lantului mitocondrial transportor de electroni;
B. Un complex multimolecular care este fixat in membrana mitocondriala interna si care catalizeaza sinteza ATP;
C. Conditionata in functionarea sa de forta generata de gradientul de pH si de potentialul membranar;
D. O proteina avand ca grupari prostetice citocromi si clustere Fe-S;
E. Un compus cu energie libera standard ridicata, cu rol de stocator de energie.
A. Inhiba respiratia si sinteza mitocondriala de ATP;
B. Inhiba respiratia fara sa afecteze sinteza de ATP din ADP si Pa;
C. Permit consumul de oxigen fara sinteza de ATP;
D. Inhiba citocrom-oxidaza;
E. Anuleaza gradientul de pH dintre fata interna si cea externa a membranei mitocondriale.
15. Dintre dovezile experimentale care sustin teoria Mitchell fac parte:
A. Observatia conform careia pentru realizarea fosforilarii oxidative este esentiala pastrarea intacta a membranei mitocondriale interne;
B. Observatia conform careia substantele cu rol de transportori membranari de protoni determina decuplarea oxidarii de fosforilare;
C. Observatia conform careia ATP se poate sintetiza in lipsa gradientului de pH intre fetele interna si externa ale membranei mitocondriale interne;
D. Observatia conform careia ATP sintaza si catena respiratorie sunt sisteme biochimice care functioneaza impreuna in cadrul aceluiasi lant metabolic;
E. Observatiile conform carora NADHQ-reductaza, citocrom reductaza si citocrom oxidaza pompeaza protoni in exteriorul matricei mitocondriale.
16. Dintre dovezile experimentale care sustin teoria Mitchell fac parte:
Page 5 of 120
A. Observatia conform careia in veziculele cu membrane lipidice care contin inserate proteine bacteriene cu rol de pompa de protoni si ATP-sintaza se poate realiza sinteza de ATP;
B. Demonstrarea experimentala a proprietatii de pompa de protoni a succinat-Q reductazei;
C. Demonstrarea experimentala a proprietatii de pompa de protoni a citocrom reductazei;
D. Demonstrarea experimentala a proprietatii de pompa de protoni a citocrom oxidazei;
E. Evidentierea unui gradient de protoni intre interiorul si exteriorul matricei mitocondriale.
17. Dintre componentele ATP-sintazei mitocondriale fac parte:
A. Subunitatea F1 care catalizeaza sinteza ATP;
B. Citocromul c care transporta elecroni;
C. Doi ioni Cu cu valente variabile;
D. Inhibitorul F1 care regleaza curgerea protonilor si sinteza ATP;
E. Hemul b care, prin pozitionarea sa in medii diferite, are afinitati diferite pentru electroni.
18. Dintre inhibitorii catenei respiratorii care realizeaza blocarea transferului de electroni de la nivelul clusterelor Fe-
S catre ubiquinona fac parte:
A. Dimercaprolul;
B. Barbituratii;
C. Oligomicina;
D. Rotenona;
E. Piericidina A.
19. Agentii decuplanti ai oxidarii de fosforilare:
A. Disociaza oxidarea prin catena respiratorie de fosforilarea realizata de fosfofructokinaza mitocondriala;
B. Pot fi: 2,4-dinitrofenolul, ciano fenil hidrazona, pentaclorofenolul;
C. Pot fi: carboxamina, atractilozid, malonatul;
D. Sunt substante cu capacitatea de a complexa protonii, facilitand transportul lor prin membrana mitocondriala interna, anihiland gradientul de protoni necesar pentru sinteza ATP;
E. Se mai numesc si ionofori.
20. In cadrul mecanismelor de control al procesului de fosforilare oxidativa:
A. Fluxul de ADP orientat spre interiorul matricei este cuplat cu cel de ATP orientat spre exterior;
Page 6 of 120
B. Iesirea ATP din matrice este realizata prin intermediul unei proteine de transport;
C. Potentialul pozitiv de membrana mareste viteza de scoatere a ATP din matricea mitocondriala;
D. Potentialul pozitiv de membrana micsoreaza viteza de scoatere a ATP din matricea mitocondriala;
E. Un rol esential il are ionul cianura.
21. In cadrul mecanismelor de control al procesului de fosforilare oxidativa:
A. Prin transportul spre exeriorul matricei mitocondriale a ATP potentialul de membrana scade semnificativ;
B. Electronii sunt transportati prin catena respiratorie indiferent daca celula necesita sau nu ATP pentru activitatile sale metabolice;
C. Electronii nu circula prin catena respiratorie daca nu se produce simultan si transformarea ADP in ATP;
D. Viteza de consum a oxigenului de catre mitocondrie scade substantial daca mediul se suplimenteaza cu ADP;
E. ATP-ADP translocaza realizeaza intrarea ADP in mitocondrie cuplata cu scoaterea ATP din mitocondrie.
22. Ionoforii:
A. Au capacitatea de a complexa cationi specifici;
B. Au structura hidrofila care permite strabaterea membranelor lipidice;
C. Pot anihila potentialul electric transmembranar;
D. Sunt substante care decupleaza oxidarea de fosforilare;
E. Cuprind ca reprezentanti valinomicina si nigericidina.
23. Acumularea de mutatii somatice la nivelul ADN mitocondrial constituie cauza favorizanta a:
A. Maladiilor degenerative (diabetul zaharat, maladia Parkinson);
B. Afectiunilor degenerative la nivelul tesuturilor a caror principala sursa energetica este glicoliza anaeroba;
C. Afectiunilor degenerative la nivelul tesuturilor in care metabolismul oxidativ este intens;
D. Afectiunilor degenerative ale miocardului si musculaturii striate;
E. Maladiei Tay-Sachs.
24. Dintre deficientele de functionare ale complexelor enzimatice ale catenei respiratorii fac parte:
Page 7 of 120
A. Neuropatia optica ereditara Leber;
B. Hipercolesterolemia familiala;
C. Hiperargininemie;
D. Encefalo miopatia-MELAS;
E. Miopatia mitocondriala infantila letala insotita de disfunctie renala.
25. Cei doi ioni Cu din componenta citocrom oxidazei:
A. Reprezinta cele doua nuclee hem din structura complexului;
B. Sunt identici ca si stare de oxidare si proprietati redox;
C. Sunt pozitionati in subunitati diferite ale complexului multimolecular;
D. Formeaza impreuna cu gruparile hem din vecinatate clustere care transfera electroni;
E. Au starea de oxidare 2+ invariabila.
26. Pentru sistemul biologic, molecula de oxigen reprezinta un acceptor terminal de electroni ideal deoarece:
A. Are cea mai mare afinitate pentru electroni din toate elementele sistemului periodic;
B. Reactioneaza foarte lent cu hidrogenul in absenta catalizatorilor sau altor factori activatori (temperatura);
C. Prin reducere se transforma intr-un compus netoxic;
D. Poate generea specii toxice prin reducerea sa partiala;
E. Poate fi partial redus, cu eliberarea produsilor rezultati in afara mitocondriei.
27. Citocrom reductaza:
A. Reprezinta un complex enzimatic cu rol de canalizare a potentialului reducator al FADH2 in catena respiratorie;
B. Canalizeaza transferul electronilor adusi de ubiquinol la citocromul C;
C. Este o “pompa de protoni”;
D. Nu este o “pompa de protoni”;
E. Are ca grupari prostetice mai multe tipuri de nuclee hem si clustere Fe-S.
28. Citocrom reductaza:
A. Este denumita si complexul III al catenei respiratorii;
Page 8 of 120
B. Este denumita si complexul II al catenei respiratorii;
C. Are o forta termodinamica mai mare decat complexul I al catenei respiratorii;
D. Are o forta termodinamica mai mica decat complexul I al catenei respiratorii;
E. Are o forta termodinamica mai mica decat complexul IV al catenei respiratorii.
29. NADH-Q reductaza:
A. Contine lanturi polipeptidice codificate exclusiv de genomul nuclear;
B. Contine lanturi polipeptidice codificate atat de genomul celular, cat si de genomul mitocondrial;
C. Realizeaza transferul potentialului reducator adus de NADH catre ubiqiunona;
D. Este denumita si complexul II al catenei respiratorii;
E. Poate suferi mutatii structurale de natura genetica, conducand la maladia denumita neuropatie optica ereditara Leber.
30. Dintre gruparile transportoare de electroni ale complexelor enzimatice ale catenei respiratorii fac parte:
A. Gruparea flavin;
B. Ionii Cu;
C. Ionii Zn;
D. Gruparea nicotin;
E. Clusterele Fe-S.
Capitolul 2. Ciclul acizilor tricarboxilici: reactii, rol biochimic, reglare, aspecte
clinice
Complement Simplu
1. In urma decarboxilarii oxidative a αceto-glutaratului in cadrul secventei de reactii din ciclul acizilor tricarboxilici
rezulta:
A. GTP si succinil-Coenzima A;
B. NADPH, H+, succinil-Coenzima A si apa;
C. NADH, H+, succinil-Coenzima A si dioxid de carbon;
D. Succinat, dioxid de carbon si GTP.
E. Piruvat, coenzima A si GTP.
Page 9 of 120
2. In urma condensarii oxaloacetatului cu acetil-Coenzima A in cadrul secventei de reactii din ciclul acizilor
tricarboxilici rezulta:
A. Coenzima A, izocitrat si apa;
B. Citrat, coenzima A si H+;
C. Cis-aconitat si coenzima A;
D. Fumarat , coenzima A si NADH, H+;
E. α cetoglutarat, succinil-coenzima A si GTP.
3. Care din urmatoarele transformari este reactie de alimentare a ciclului acizilor tricarboxilici cu intermediari?
A. Piruvat + ATP + CO2 →Oxaloacetat + ADP + Pa
B. Acetil Co A + Oxaloacetat →Citrat + CoA-SH
C. Oxaloacetat + GTP →Fosfoenol piruvat + CO2 + GDP
D. Succinil-Co A + GDP + Pa → Succinat + GTP + Co A-SH
E. Nici o reactie dintre cele de mai sus.
4. Ciclul acizilor tricarboxilici este:
A. O cale apartinand exclusiv laturii catabolice a metabolismului;
B. O cale apartinand exclusiv laturii anabolice a metabolismului;
C. O parte integranta a procesului prin care este obtinuta energia eliberata din descompunerea glucidelor, lipidelor si aminoacizilor;
D. O parte a catenei respiratorii;
E. O sursa de NADPH necesar biosintezelor reductive.
5. Enzimele ciclului acidului citric sunt localizate:
A. Numai in citoplasma;
B. Numai spre fata externa a membranei mitocondriale interne;
C. Numai spre fata interna a membranei mitocondriale interne;
D. Numai in matricea mitocondriala interna;
E. Atat in citoplasma cat si in mitocondrie.
6. Izomerizarea citratului in cadrul ciclului acizilor tricarboxilici:
Page 10 of 120
A. Este catalizata de citrat sintaza;
B. Este catalizata de izocitrat dehidrogenaza;
C. Este catalizata de aconitaza;
D. Este o reactie prin care se genereaza energie in mod direct;
E. Este o reactie de oxidare.
7. In cadrul ciclului acizilor tricarboxilici, regenerarea oxaloacetatului are loc in final prin:
A. Oxidarea succinatului;
B. Oxidarea malatului;
C. Hidratarea fumaratului;
D. Oxidarea izocitratului;
E. Decarboxilarea oxalo succinatului.
8. Singura reactie a ciclului acidului citric prin care se genereaza energie in mod direct este:
A. Izomerizarea citratului la izocitrat;
B. Conversia fumaratului in malat;
C. Conversia succinil-Co A in succinat;
D. Decarboxilarea oxidativa a αceto glutaratului;
E. Condensarea oxaloacetatului cu acetil Co A.
9. Succinat dehidrogenaza, enzima din ciclul acidului citric:
A. Este singura enzima a ciclului fixata in membrana interna mitocondriala;
B. Catalizeaza reducerea succinatului la fumarat;
C. Este o hem proteina;
D. Este o enzima avand drept coenzima NAD+;
E. Catalizeaza hidratarea succinatului la L-malat.
10. Dintre vitaminele hidrosolubile care servesc drept coenzime in ciclul acizilor tricarboxilici face parte:
A. Piridoxal fosfatul;
B. Tocoferolul;
Page 11 of 120
C. Biotina;
D. Acidul folic;
E. Tiamina.
Complement Multiplu
11. In cadrul secventei de reactii din ciclul acizilor tricarboxilici, izocitratul:
A. Este transformat in oxalosuccinat concomitent cu hidrogenarea unei molecule NAD+;
B. Este transformat in succinil-coenzima A concomitent cu hidrogenarea unei molecule NAD+;
C. Este oxidat;
D. Este redus;
E. Este scindat in doi compusi cu cate trei atomi de carbon.
12. In cadrul secventei de reactii din ciclul acizilor tricarboxilici succinatul:
A. Este decarboxilat;
B. Este convertit in oxaloacetat printr-o secventa de trei reactii concomitent cu hidrogenarea unei molecule de NAD+ si a unei molecule de FAD;
C. Este oxidat;
D. Este convertit in acetil-coenzima A printr-o secventa de trei reactii concomitent cu hidrogenarea unei molecule de NAD+si a unei molecule de FAD;
E. Este transformat in fumarat de catre enzima succinat dehidrogenaza.
13. Prin trecerea unei grupari acetil activate prin secventa de reactii a ciclului acizilor tricarboxilici:
A. Sunt reduse in total 3 molecule FAD;
B. Sunt reduse in total 3 molecule NAD+;
C. Se formeaza o legatura macroergica (GTP);
D. Este redusa o singura molecula NAD+;
E. Este redusa o singura molecula FAD.
14. Ciclul acizilor tricarboxilici este:
A. O cale metabolica care se desfasoara in citosolul celulelor;
B. Un proces amfibolic;
Page 12 of 120
C. O cale metabolica care constituie parte comuna finala a oxidarii principalelor categorii de substante-combustibil;
D. Alimentat cu oxaloacetat provenit in reactia de carboxilare a piruvatului, catalizata de piruvat carboxilaza;
E. O cale metabolica care se desfasoara in matricea mitocondriala.
15. Aconitaza:
A. Catalizeaza numai reactia de deshidratare a citratului;
B. Catalizeaza numai reactia de hidratare a cis-aconitatului;
C. Contine clustere Fe-S;
D. Catalizeaza procesul de izomerizare a citratului la izocitrat;
E. Contine Fe neheminic.
16. In ciclul Krebs nu exista:
A. Patru reactii NAD+-dependente;
B. O reactie FAD-dependenta;
C. O fosforilare la nivel de substrat;
D. O dehidrogenaza NADP-dependenta;
E. O enzima cu biotina drept coenzima.
17. Nu apartin ciclului Krebs transformarile:
A. α-Cetoglutarat →succinil-Co A;
B. Fumarat →Malat;
C. Malat →Fosfoenol piruvat;
D. Citrat →Izocitrat;
E. Citrat → Acetil-Co A + Oxaloacetat
18. Nicotin amid adenin dinucleotid constituie coenzima pentru urmatoarele enzime din ciclul Krebs:
A. Succinat dehidrogenaza;
B. Izocitrat dehidrogenaza;
C. α-Ceto glutarat dehidrogenaza;
Page 13 of 120
D. Aconitaza;
E. Malat dehidrogenaza.
19. Reactiile care furnizeaza schelete hidrocarbonate pentru sinteza de aminoacizi neesentiali pornind de la
intermediari ai ciclului Krebs sunt:
A. Conversia lactatului in piruvat;
B. Reactia de interconverise a piruvatului si alaninei;
C. Reactia de interconversie a oxaloacetatului si aspartatului;
D. Decarboxilarea oxaloacetatului la fosfoenol piruvat;
E. Reactia de izomerizare a citratului la izocitrat.
20. Reactia de carboxilare a piruvatului cu formare de oxaloacetat catalizata de piruvat carboxilaza:
A. Constituie o reactie de transfer a intermadiarilor ciclului Krebs catre gluconeogeneza;
B. Constituie o reactie de alimentare a ciclului Krebs cu oxaloacetat;
C. Este o reactie importanta pentru mentinerea concentratiei adecvate de oxaloacetat necesare pentru functionarea ciclului Krebs;
D. Este inhibata de acumularea acetil-Co A;
E. Este o reactie care se produce cu consum de energie furnizata prin hidroliza unei molecule de ATP.
21. Pentru tesuturile in care functia principala a ciclului acizilor tricarboxilici este cea de furnizor de energie:
A. Controlul catenei respiratorii constituie si o modalitate de control a ciclului Krebs;
B. Viteza de desfasurare a ciclului Krebs depinde de rata de utilizare a ATP;
C. Reactia care regleaza viteza ciclului Krebs este cea de conversie a fructozei in fructozo-1,6bis fosfat;
D. Viteza de desfasurare a ciclului Krebs depinde exclusiv de activitatea aconitazei;
E. Viteza de desfasurare a ciclului Krebs depinde exclusiv de activitatea citrat dehidrogenazei.
22. Controlul enzimatic al ratei de desfasurare a ciclului acizilor tricarboxilici se poate desfasura:
A. La nivelul reactiilor de decarboxilare;
B. La nivelul reactiilor la echilibru;
C. La nivelul reactiei catalizate de citrat sintaza;
D. La nivelul reactiei catalizate de α-cetoglutarat dehidrogenaza;
Page 14 of 120
E. La nivelul reactiilor catalizate de transaminaze.
23. O reactie importanta pentru mentinerea concentratiei adecvate de oxaloacetat in mitocondrie, necesara pentru
“pornirea” ciclului Krebs:
A. Este cea de sinteza a fosfoenol piruvatului din creatin fosfat;
B. Este cea catalizata de piruvat carboxilaza;
C. Este cea de transaminare dintre glutamat si piruvat;
D. Este activata allosteric de excesul de acetil-Co A;
E. Este cea de convertire a piruvatului in lactat, catalizata de lactat dehidrogenaza.
24. Viteza de desfasurare a ciclului acidului citric depinde de:
A. Aportul de cofactori ai dehidrogenazelor care actioneaza in respectiva cale metabolica;
B. Statusul energetic general al celulei;
C. Raportul [ATP]/[ADP] din celula;
D. Reactia catalizata de tiamin pirofosfat transcetolaza;
E. Activitatea fosfofructo kinazei mitocondriale.
25. Dintre enzimele ciclului Krebs a caror activitate este modulata de rapoartele [ATP]/[ADP] si [NADH]/[NAD+] fac
parte:
A. Transcetolaza;
B. Gluco kinaza;
C. Citrat sintaza;
D. Izocitrat dehidrogenaza;
E. Lactat dehidrogenaza.
26. Generarea unei legaturi macroergice in cadrul reactiilor ciclului Krebs:
A. Are loc prin izomerizarea citratului;
B. Este o reactie catalizata de succinil CoA sintetaza;
C. Conduce la formarea unei molecule de ATP;
D. Are loc prin desfacerea unei legaturi tioesterice macroergice;
E. Are loc prin hidrogenarea FAD.
Page 15 of 120
27. In cadrul secventei de reactii ale ciclului acizilor tricarboxilici:
A. Hidratarea fumaratului la malat are loc stereospecific;
B. Prin hidratarea fumaratului se obtine exclusiv D-malat;
C. Prin hidratarea fumaratului se obtine exclusiv L-malat;
D. Conversia fumaratului la malat este catalizata de malat dehidrogenaza;
E. Conversia fumaratului la malat este catalizata de fumaraza.
28. Transferul intermediarilor ciclului Krebs catre gluconeogeneza este realizat:
A. Prin transaminarea piruvatului la alanina;
B. Prin decarboxilarea oxaloacetatului la fosfoenol piruvat;
C. De catre enzima piruvat carboxilaza;
D. De catre enzima fosfo enol piruvat carboxikinaza;
E. Cu aport energetc adus de ATP.
29. In cadrul sirului de transformari incluse in ciclul acizilor tricarboxilici:
A. Se genereaza energie prin oxidarea gruparilor acetil activate;
B. Se genereaza intermediari pentru procesele de biosinteza;
C. Se furnizeaza potential reducator pentru catena respiratorie;
D. Se indeparteaza compusi cu potential toxic ridicat;
E. Se sintetizeaza nuclee tetrapirolice.
30. In cadrul secventei de reactii ale ciclului acizilor tricarboxilici:
A. Succinatul este oxidat la fumarat cu transferarea potentialului reducator pe o molecula NAD+;
B. Succinatul este oxidat la fumarat cu transferarea potentialului reducator pe o molecula FAD;
C. Succinatul este oxidat la fumarat cu transferarea potentialului reducator pe o molecula NADH;
D. Succinatul este oxidat la fumarat cu transferarea potentialului reducator pe o molecula FADH2;
E. Enzima care catalizeaza oxidarea succinatului este inhibata de oxaloacetat
Page 16 of 120
Capitolul 3. Metabolismul glucidelor: glicoliza si oxidarea acidului piruvic,
aspecte clinice
Complement Simplu
1. In cursul reactiilor de degradare a glucozei prin calea glicolizei:
A. 1,3 bis-fosfogliceratul este convertit la 3-fosfoglicerat cu obtinerea unei legaturi macroergice (ATP);
B. Fosfoenol piruvatul este convertit in piruvat cu obtinerea unei legaturi macroergice (ATP);
C. Citratul este convertit la izocitrat printr-o deshidratare urmata de o rehidratare;
D. Corecte A si B;
E. Corecte A, B si C.
2. Procesul de decarboxilare oxidativa a piruvatului rezultat din glicoliza se desfasoara:
A. In acelasi compartiment cellular in care are loc glicoliza;
B. La nivelul ribozomilor;
C. In interiorul mitocondriei;
D. In cisternele lizozomale;
E. In citosolul eritrocitelor.
3. Enzima-cheie a glicolizei este:
A. Hexokinaza ;
B. Enolaza ;
C. Lactat-dehidrogenaza;
D. Fosfofructokinaza ;
E. Fosfoglicerat mutaza.
4. In urma procesului de degradare anaeroba prin calea glicolitica a unei molecule de glucoza rezulta:
A. doua molecule ATP si o molecula NAD, H+;
B. o molecula GTP, o molecula FADH2 si 3 molecule NADH, H+;
C. doua molecule ATP si 3 molecule NADH,H+;
D. doua molecule ATP;
Page 17 of 120
E. nici o varianta corecta.
5. In calea glicolitica glucoza este fosforilata la glucozo-6-fosfat de catre:
A. Hexofosfataza in parenchimul hepatic si in pancreasul endocrin;
B. Glucofosfataza in parenchimul hepatic si in pancreasul endocrin;
C. Glucokinaza in parenchimul hepatic si in pancreasul endocrin;
D. Hexokinaza in parenchimul hepatic si in pancreasul endocrin;
E. Corecte C. si D.
6. Fosforilarea glucozei la glucozo-6-fosfat in calea glicolitica se desfasoara avand ca agent de fosforilare:
A. ATP;
B. UTP;
C. GTP;
D. ATP sau UTP;
E. NADPH
7. Fata de glucokinaza, hexokinaza are:
A. KM mai mare;
B. KM mai mic;
C. KM identic in valoare absoluta;
D. Afinitate mai mica pentru substrat;
E. Corecte A. si D.
8. Functia glucokinazei este:
A. De a inlatura glucoza din sange, imediat dupa aportul alimentar;
B. De a creste concentratia glucozei in sange atunci cand organismul necesita aceasta;
C. De a cataliza fosforilarea fructozo-6-fosfatului;
D. De a cataliza hidroliza glucozo-6-fosfatului;
E. De a finaliza transformarile glucozei pe calea glicolitica.
9. In secventa de reactii a glicolizei:
Page 18 of 120
A. Gliceraldehid 3-fosfatul este transformat in 1,3 bis fosfoglicerat prin reducerea gruparii aldehidice;
B. Gliceraldehid 3-fosfatul este transformat in 1,3 bis fosfoglicerat prin oxidarea gruparii aldehidice;
C. Gliceraldehid 3-fosfatul este transformat in 1,3 bis fosfoglicerat prin reducerea gruparii aldehidice si fixarea unui rest fosfat;
D. Gliceraldehid 3-fosfatul este transformat in 1,3 bis fosfoglicerat prin oxidarea gruparii aldehidice intr-o reactie care decurge in ambele sensuri cu aceiasi viteza;
E. Gliceraldehid 3-fosfatul este transformat in 1,3 bis fosfoglicerat prin aditionarea gruparii aldehidice la o molecula fosfat anorganic.
10. Care enzima catalizeaza o reactie reversibila in calea glicolitica?
A. Piruvat dehidrogenaza;
B. Aldolaza;
C. Fosfoenol-piruvat-carboxikinaza;
D. Glucokinaza;
E. Glucozo-6-fosfataza.
Complement Multiplu
11. Situsuri de formare de legaturi macroergice in cadrul glicolizei sunt:
A. Reactia de conversie a 1,3 bis fosfogliceratului in 3, fosfoglicerat;
B. Reactia de conversie a fosfoenol piruvatului in piruvat;
C. Reactia de conversie a glucozo 6 fosfatului in fructozo 6 fosfat;
D. Reactia de conversie a fructozo 6 fosfatului in fructozo 1,6 bis fosfat
E. Reactia de conversie a fructozo 1,6 bis fosfatului in dihidroxi aceton fosfat si gliceraldehid 3 fosfat.
A. O proteina cu masa moleculara mica, care serveste ca molecula-primer pentru grefarea primei molecule de glucoza in cadrul sintezei glicogenului;
B. Enzima de ramificare, care transfera o parte a catenei poliglucidice la un lant alaturat;
C. Enzima-cheie a glucogenogenezei;
D. O molecula-efector care creste gradul de ramificare al moleculelor de glicogen in pancreas si tesutul adipos;
E. O maladie genetica in care organismul stocheaza glicogen fara a-l putea utiliza.
2. Enzimele principale care controleaza metabolismul glicogenului sunt:
Page 31 of 120
A. Fosfoglucomutaza si pirofosfataza;
B. Hexokinaza si glucokinaza;
C. Glicogen fosforilaza si glicogen sintaza;
D. Piruvat carboxikinaza si glucozo 6 fosfataza;
E. Fosfoglucomutaza si glucokinaza.
3. Suntul pentozo-fosfatilor este:
A. O cale majora de metabolizare a glucozei prin care se genereaza NADPH, H+ pentru biosinteze reductive si riboza pentru biosinteza nucleotidelor si acizilor nucleici;
B. O cale majora de metabolizare a glucozei cu generare de legaturi macroergice (ATP) si potential reducator (NADPH,H+);
C. O cale alternativa de metabolizare a glucozei la acid glucuronic cu generare de potential reducator (NADPH,H+);
D. O prescurtare a caii de metabolizare a glucozei prin glicoliza, care are ca enzime caracteristice fosfofructokinaza si gluconolacton hidrolaza;
E. O cale metabolica activa numai in tesutul nervos, derm si cortexul glandelor suprarenale.
4. Calea acidului glucuronic:
A. Se desfasoara in tesutul pulmonar si are ca rezultat sinteza de pentoze si de acid ascorbic la primate;
B. Se desfasoara in ficat si are ca scop major excretia compusilor straini organismului si produsilor de catabolizare proprii;
C. Este o cale majora de metabolizare a glucozei, prezenta in toate tesuturile (ubiquitara);
D. Este mult amplificata in cazul unei maladii genetice denumita pentozurie esentiala ;
E. Corecte B si D.
5. In procesul biosintezei glicogenului:
A. Glucoza este fosforilata la glucozo-1-fosfat de catre hexokinaza;
B. Glucoza este fosforilata la glucozo-6-fosfat de catre hexokinaza;
C. Glucozo-1-fosfat este transformata in glucozo-6-fosfat de catre fosfoglucomutaza;
D. Glucoza este activata cu ajutorul energiei furnizate de creatin fosfat;
E. Adaugarea de noi molecule de glucoza in catena glicogenului se face la capatul reducator.
Page 32 of 120
6. In procesul biosintezei glicogenului:
A. Enzima de ramificare transfera o catena lineara cu minimum 6 unitati de glucoza pe un lant alaturat;
B. Molecula de glucoza este incarcata energetic prin sinteza ATP-glucozei;
C. Molecula de glucoza este incarcata energetic prin sinteza GTP-glucozei;
D. Molecula de glucoza este incarcata energetic prin sinteza CTP-glucozei;
E. Se produce energie metabolica inmagazinata in UTP.
7. Glicogenoliza:
A. Reprezinta calea metabolica de sinteza a glicogenului la nivelul ficatului si muschilor;
B. Este o cale metabolica identica cu glicogenogeneza;
C. Reprezinta calea metabolica de hidroliza a glicogenului prin hidroliza legaturilor glicozidice α1-4 si α1-6;
D. Are ca molecula furmizoare de energie UTP;
E. Se desfasoara exclusiv in tesutul hepatic.
8. Enzimele principale care controleaza metabolismul glicogenului sunt:
A. Fosfoglucomutaza si enzima de ramificare;
B. Glicogen fosforilaza si glicogen sintaza;
C. Hexokinaza si pirofosfataza;
D. Aldolaza si cetolaza;
E. Fosfofructo kinaza si piruvatcarboxi kinaza.
9. In muschi glicogen fosforilaza este activata de:
A. Insulina;
B. Glucagon;
C. Adrenalina;
D. Iodotironina;
E. Estradiol.
10. Maladia von Gierke:
Page 33 of 120
A. Este cauzata de deficienta enzimei glicogen sintaza;
B. Are ca patologie caracteristica incaracrea hepatocitelor si a tubulilor renali cu glicogen;
C. Se manifesta clinic prin hiperglicemie si hipolipemie;
D. Este o boala infectioasa;
E. Este cauzata de absenta enzimei de deramificare.
Complement Multiplu
11. Glicogenul este stocat in:
A. Eritrocitele tinere;
B. Eritrocitele mature;
C. Ficat;
D. Tesut muscular;
E. Pancreas.
12. Care din etapele descrise fac parte din procesul de biosinteza al glicogenului?
A. Fosforilarea glucozei la glucozo 6 fosfat de catre hexokinaza;
B. Carboxilarea piruvatului la oxaloacetat de catre piruvat carboxilaza;
C. Transformarea glucozo 6 fosfatului in glucozo 1 fosfat de catre fosfoglucomutaza;
D. Izomerizarea glucozo 6 fosfat in fructozo 6 fosfat de catre fosfoglucoizmeraza;
E. Conversia lactatului la piruvat de catre lactat dehidrogenaza.
13. Glicogenoliza reprezinta:
A. O cale distincta de glicogenogeneza;
B. Procesul de hidroliza al moleculelor de glicogen in molecule de glucoza;
C. Procesul reversibil al glicogenogenezei;
D. Un proces ce se desfasoara preponderant in eritrocitele tinere;
E. Un proces inhibat de insulina.
14. Bolile de tezaurizare a glicogenului:
A. Sunt un grup de maladii genetice caracterizate prin depozitarea in organism a unui tip anormal de glicogen;
Page 34 of 120
B. Se pot manifesta prin incarcarea hepatocitelor si a tubulilor renali cu glicogen;
C. Se pot trata prin transplant de ficat;
D. Se manifesta prin liza hematiilor imbatranite;
E. Pot cauza lipsa rezistentei la efort fizic.
15. Etape ale suntului pentozofosfatilor sunt:
A. Fosforilarea glucozei cu obtinerea de glucozo 1 fosfat de catre hexokinaza;
B. Dehidrogenarea glucozo 6 fosfatului cu obtinerea 6 fosfo glucono lactonei de catre glucozo 6 fosfat dehidrogenaza NADP+ dependent;
C. Hidroliza 6 fosfo gluconolactonei cu obtinerea de 6 fosfogluconat de catre transcetolaza;
D. Transformarea 6 fosfogluconatului in 3 ceto 6 fosfogluconat (instabil) de catre 6 fosfogluconat dehidrogenaza;
E. Transformarea glucozo 6 fosfatului in glucozo 1 fosfat de catre fosfoglucomutaza.
16. Etape ale suntului pentozofosfatilor sunt:
A. Decarboxilarea spontana a 3 ceto 6 fosfogluconatului cu formarea ribulozo 5 fosfatului;
B. Epimerizarea ribulozo 5 fosfatului cu obtinerea xilulozo 5 fosfatului;
C. Izomerizarea gliceraldehid 3 fosfatului cu obtinerea dihidroxi aceton fosfatului;
D. Izomerizarea citratului in cis-aconitat;
E. Crearea unei legaturi 1-6 glicozidice intre doua molecule de glucozo 6 fosfat.
17. Etape ale caii acidului glucuronic sunt:
A. Oxidarea UDP glucozei cu generarea de UDP-glucuronat de catre UDP glucozo dehidrogenaza;
B. Reactia glucozo-1 fosfatului cu UTP, cu generarea de UDP glucoza;
C. Izomerizarea glucozo 6 fosfatului la glucozo 1 fosfat de catre fosfoglucomutaza;
D. Izomerizarea gliceraldehid 3 fosfatului cu obtinerea dihidroxi aceton fosfatului;
E. Hidroliza 6 fosfo gluconolactonei cu obtinerea de 6 fosfogluconat de catre transcetolaza.
18. Amplificarea sintezei de acid glucuronic pornind de la glucoza poate fi indusa de:
A. Uree;
B. Barbital;
Page 35 of 120
C. Clorobutanol;
D. Glucoza in exces;
E. Acidul piruvic.
19. Dintre bolile de tezaurizare a glicogenului fac parte:
A. Sindromul McArdle;
B. Neuropatia optica ereditara Leber;
C. Maladia Cori;
D. MELAS;
E. Alcaptonuria.
20. In urma contractiei musculare:
A. Glicogenogeneza se amplifica in muschi de cateva sute de ori;
B. Glicogenoliza se amplifica in muschi de cateva sute de ori;
C. Are loc activarea rapida a glicogen fosforilazei prin fosforilarea sa;
D. Are loc activarea rapida a glicogen sintazei prin defosforilarea sa;
E. Are loc sinteza amplificata de glicogenina.
21. Suntul pentozo fosfatilor:
A. Este o cale majora de obtinere a energiei din moleculele de glucoza;
B. Nu genereaza ATP;
C. Genereaza potential reducator pentru catena respiratorie;
D. Se desfasoara in matricea mitocondriala;
E. Cuprinde doua etape, dintre care una oxidativa.
22. Reactiile suntului pentozo fosfatilor:
A. Se petrec in citosol;
B. Se petrec in matricea mitocondriala;
C. Se petrec in reticulul endoplasmatic;
D. Cuprind o etapa oxidativa si una neoxidativa;
Page 36 of 120
E. Cuprind doua etape oxidative care se desfasoara in doua compartimente intracelulare diferite.
23. In prima etapa a suntului pentozo fosfatilor:
A. Are loc transformarea ribulozo-5-fosfatului in glucozo-6-fosfat;
B. Participa la procesul catalitic transcetolaza;
C. Are loc dehidrogenarea glucozo-6-fosfatului cu formarea de 6 fosfogluconolactona;
D. Sunt generate 2 molecule NADPH;
E. Are loc transferul unei unitati cu 2 atomi de carbon de la o cetoza catre o aldoza.
24. O mutatie genetica care induce deficit de functionare al enzimei glucozo-6-fosfat dehidrogenaza:
A. Are drept consecinta blocarea caii glicolitice;
B. Are drept consecinta blocarea caii acidului glucuronic;
C. Are drept consecinta blocarea producerii de NADPH;
D. Are drept consecinta liza eritrocitara la administrarea de antimalarice;
E. Are drept consecinta anemia falciforma.
25. Dintre diferentele de catabolizare ale glucozei prin cele doua cai majore fac parte:
A. Generarea de pentoze fosforilate in cadrul suntului pentozo fosfatilor;
B. Locul diferit de desfasurare al reactiilor;
C. Generarea de potential reducator sub forma de NADH in cadrul suntului pentozo fosfatilor;
D. Generarea de legaturi fosfat macroergice in cadrul glicolizei;
E. Existenta unor intermediari comuni.
26. Suntul pentozo fosfatilor:
A. Se desfasoara cu mare intensitate in glanda mamara in lactatie;
B. Se desfasoara cu mare intensitate in muschii scheletici;
C. Se desfasoara cu mare intensitate in tesuturile in care au loc biosinteze reductive;
D. Este activat de insulina;
E. Produce NADH pentru mentinerea glutationului in stare oxidata in eritrocite. 147 Calea acidului glucuronic:
27. UDP-glucuronatul:
Page 37 of 120
A. Este forma activa a glicogenului;
B. Este forma activa a glucuronatului;
C. Reactioneaza cu bilirubina pentru a genera bilirubina conjugata;
D. Reactioneaza cu acidul uric pentru a genera acidul uric conjugat;
E. Se excreta exclusiv pe cale digestiva.
28. Calea acidului glucuronic:
A. Genereaza ATP;
B. Nu genereaza ATP;
C. Genereaza NADPH;
D. Nu genereaza NADPH;
E. Porneste de la glucoza-6-fosfat.
29. Spre deosebire de glicogenul hepatic, glicogenul muscular:
A. Are rolul de a pune la dispozitie hexoze pentru propriile celule;
B. Se gaseste in procent mult mai mic in celule;
C. Se gaseste in celule in procent de peste 16%;
D. Are rolul de a pune la dispozitie hexoze pentru mentinerea constanta a glicemiei;
E. Are structura moleculara neramificata.
30. Calea acidului glucuronic:
A. Are ca semnificatie biologica generarea de potential reducator pentru biosinteze
B. Are ca semnificatie biologica excretarea compusilor straini organismului (xenobiotice);
C. Are ca semnificatie biologica excretarea produsilor de catabolizare ai organismului
D. Se desfasoara in muschii scheletici;
E. Se desfasoara in ficat.
Page 38 of 120
Capitolul 6. Metabolismul lipidelor. Catabolismul acizilor grasi: digestia si
1. Digestia lipidelor din aport alimentar incepe din:
A. Cavitatea bucala;
B. Traiectul esofagian;
C. Stomac;
D. Citosolul celulelor enterale;
E. Colonul transvers.
2. Deficientele in structura acil gras translocazelor au ca manifestari clinice:
A. Pierderi de memorie si neuropatie dupa exercitii fizice exesive;
B. Voma, diaree si crampe musculare in urma dietelor sarace in lipide;
C. Hiperglicemie si poliurie;
D. Crampe muscular care se accentueaza dupa mese, diete bogate in lipide sau exercitii fizice;
E. Inflamarea tesutului adipos.
3. Locul majoritar de acumulare al triacilglicerolilor la mamifere este:
A. Citoplasma hepatocitelor;
B. Citoplasma neuronilor;
C. Citoplasma adipocitelor;
D. Citoplasma miocitelor;
E. Eritrocitul matur.
4. Chilomicronii:
A. Sunt sintetizaţi în intestin în timpul absorbţiei lipidelor si secretaţi în sistemul limfatic şi de acolo, în sânge;
B. Nu contin colesterol din aport exogen;
C. Sunt sintetizati in adipocite si secretati in sistemul limfatic şi de acolo, în sânge;
D. Sunt particule monomoleculare cu forme neregulate;
Page 39 of 120
E. Sunt agregate lipidice.
5. Evenimentul iniţial în beta oxidarea acizilor grasi este:
A. Activarea acizilor grasi prin formarea unui tioester cu Coenzima A;
B. Formarea unui acil adenilat cu ATP;
C. Hidroliza triacilglicerolilor de catre lipaze;
D. Secretia epinefrinei, nor-epinefrinei si insulinei;
E. Intrarea acizilor grasi in mitocondrie.
6. Activitatea lipazelor din adipocite este reglata de catre:
A. Estradiol;
B. ACTH;
C. Testosteron;
D. Ocitocina;
E. Cortisol.
7. Inhibitia activitatii lipazice este realizata de:
A. Glucagon;
B. Adrenalina;
C. ACTH;
D. Insulina;
E. Nor-adrenalina.
8. Degradarea oxidativa a acizilor grasi are loc in:
A. Citosol;
B. Reticul endoplasmatic;
C. Citoplasma adipocitelor;
D. Membrana mitocondriala interna;
E. Matricea mitocondriala.
9. Transportul acizilor grasi in compartimentul celular unde are loc β-oxidarea lor se face prin intermediul:
Page 40 of 120
A. Coenzimei A;
B. Carnitinei;
C. ATP;
D. Proteinei transportoare de grupari acil;
E. Chilomicronilor.
10. Degradarea catenelor saturate de acizi grasi activati are loc prin:
A. Clivarea secvenţialăde unităţi cu câte doi atomi de carbon din catena acidului gras;
B. Clivarea secvenţialăde unităţi cu câte trei atomi de carbon din catena acidului gras;
C. Clivarea secvenţialăde unităţi cu câte un atom de carbon din catena acidului gras;
D. Oxidarea secventiala a fiecarui atom de carbon din catena pana la CO2;
E. Nici un raspuns corect.
Complement Multiplu
11. Etape ale βoxidarii acizilor grasi sunt:
A. Oxidarea acil gras-Coenzimei A de catre acil CoA dehidrogenaza cu formare de enoil CoA;
B. Transportul gruparilor acil gras activate in matricea mitocondriala dupa conjugarea lor cu carnitina;
C. Sinteza malonil-CoA pornind de la acetil CoA si HCO3-;
D. Elongarea catenei acidului palmitic in reticulul endoplasmatic al hepatocitelor;
E. Carboxilarea biotinei pe baza energiei furnizate de ATP la nivelul acetil Co A carboxilazei.
12. Degradarea oxidativa a acizilor grasi nesaturati:
A. Necesita fata de procesul de oxidare al acizilor grasi saturati doua enzime suplimentare:
B. o izomeraza si o reductaza;
C. Se desfasoara in matricea mitocondriala dupa activarea gruparii acil gras si transportul ei din citosol;
D. Se desfasoara in cisternele lizozomale dupa activarea gruparii acil gras si transportul ei din citosol;
E. Necesita fata de procesul de oxidare al acizilor grasi saturati un set de enzime complet diferit;
13. Cetogeneza:
A. Implica sinteza si descompunerea 3-hidroxi-3 metil glutaril CoA de catre doua enzime;
Page 41 of 120
B. Genereaza molecule-combustibil destinate tesuturilor extrahepatice;
C. Generaza molecule-combustibil destinate tesutului hepatic;
D. Este reglata prin controlul mobilizarii acizilor grasi liberi din tesutul adipos prin mecanisme hormonale;
E. Are loc numai in conditii patologice.
14. Cetogeneza este reglata prin:
A. Controlul mobilizarii acizilor grasi liberi din tesutul adipos, efectuat preponderant hormonal;
B. Activarea carnitin palmitoil transferazei I hepatice;
C. Raportul de repartizare al acetil CoA generate din βoxidarea acizilor grasi intre procesul de cetogeneza si ciclul aciilor tricarboxilici;
D. Pompele de protoni ale catenei respiratorii din membrana mitocondriala interna;
E. Procesul de homeotermie realizat la nivelul hipotalamusului.
15. Acil CoA sintetaza:
A. Catalizeaza oxidarea Acil Co A la enoil Co A;
B. Catalizeaza reducerea catenei acil activate;
C. Induce formarea unei duble legaturi in configuratie trans;
D. Induce formarea unei duble legaturi in configuratie cis;
E. Utilizeaza pentru cataliza energia furnizata de doua legaturi macroergice.
16. In reactia de formare a trans ∆enoil Co A, in cadrul caii metabolice a βoxidarii acizilor grasi:
A. Are loc hidrogenarea unei molecule FAD;
B. Are loc hidrogenarea unei molecule NAD;
C. Catalizatorul este enoil Co hidrataza;
D. Catalizatorul este acil Co A dehidrogenaza;
E. Are loc hidroliza unei molecule ATP.
17. Enoil Co A hidrataza:
A. Este o enzima stereospecifica;
B. Nu hidrateaza legaturile duble cu structura cis;
Page 42 of 120
C. Hidrateaza si dublele legaturi cu structura cis;
D. Are ca produs de reactie L-3 hidroxiacil CoA;
E. Este o enzima citosolica.
18. Acil Co a dehidrogenazele:
A. Catalizeaza reactia de transformare a 3 ceto acil Co A in acil Co A;
B. Catalizeaza reactia de transformare a acil Co A in trans ∆enoli Co A;
C. Utilizeaza ca acceptor de potential reducator molecula NAD;
D. Utilizeaza ca acceptor de potential reducator molecula FAD;
E. Utilizeaza pentru reactia pe care o catalizeaza energie furnizata de ATP.
19. Acil Co A dehidrogenazele:
A. Cuprind enzime specifice de grup pentru catene hidrocarbonate lungi (12-18 atomi de carbon);
B. Cuprind enzime specifice de grup pentru catene hidrocarbonate medii (9-4 atomi de carbon);
C. Genereaza exclusiv legaturi duble in conformatie cis;
D. Genereaza exclusiv legaturi duble in conformatie trans;
E. Genereaza atat duble legaturi cis cat si trans.
20. In cazul oxidarii acizilor grasi nesaturati:
A. Calea βoxidarii este identica cu a acizilor grasi saturati;
B. Sunt necesare doua enzime suplimentare fata de calea oxidarii acizilor grasi saturati: o dehidrogenaza si o oxidaza;
C. Sunt necesare doua enzime suplimentare fata de calea oxidarii acizilor grasi saturati: o izomeraza si o reductaza;
D. Mecanismul de transport a acizilor in matricea mitocondriala este identic;
E. Este necesara modificarea configuratiei trans a dublei legaturi din catena acidului gras nesaturat in configuratie cis cu ajutorul unei izomeraze.
21. In cazul oxidarii acizilor grasi polinesaturati:
A. Sunt necesare trei enzime suplimentare fata de calea oxidarii acizilor grasi saturati: 2,4 dienoil CoA reductaza,o izomeraza si o reductaza;
B. Sunt necesare doua enzime suplimentare fata de calea oxidarii acizilor grasi saturati: o izomeraza si 2,4 dienoil CoA reductaza;
Page 43 of 120
C. 2,4 dienoil CoA reductaza transforma compusul 2,4 dienoil CoA in cis ∆3 enoil CoA;
D. 2,4 dienoil CoA reductaza transforma compusul 2,4 dienoil CoA in trans ∆3 enoil CoA;
E. Calea metabolica se desfasoara in citosol, nemainecesitand transportul acizilor grasi in matricea mitocondriala.
22. βoxidarea acizilor grasi in peroxizomi:
A. Conduce la malonil CoA;
B. Difera de cea mitocondriala prin catalizatorul si acceptorul de hidrogen al reactiei de oxidare a L-3-hidroxiacli Co A;
C. Difera de cea mitocondriala prin catalizatorul si acceptorul de hidrogen al reactiei de oxidare a acil Co A;
D. Conduce la acetil CoA;
E. Genereaza peroxid de hidrogen.
23. Cetogeneza are loc:
A. In cazul unei rate foarte crescute a oxidarii acizilor grasi;
B. Exclusiv in tesutul hepatic;
C. In tesutul hepatic si tesutul nervos;
D. Numai in situatii patologice;
E. In tesuturile care utilizeaza corpii cetonici drept molecule-combustibil.
24. Corpii cetonici sunt:
A. D (-) 3-hidroxipropionatul;
B. Acetoacetatul;
C. Acetona;
D. Acetobutiratul;
E. D (-) 3-hidroxibutiratul.
25. Cetogeneza implica:
A. Sinteza 3-hidroxi-3 metil glutaril CoA;
B. Descompunerea 3-hidroxi-3 metil glutaril CoA;
Page 44 of 120
C. Sinteza si descompunerea 3-hidroxi-3 metil glutaril CoA;
D. Actiunea HMG-CoA sintazei;
E. Actiunea HMG-CoA liazei.
26. Corpii cetonici sunt oxidati:
A. In tesutul hepatic;
B. In tesuturile extrahepatice;
C. Pana la saturarea caii de conversie (12 mM/L corpi cetonici in sange);
D. Producand o intensificare semnificativa a consumului de oxigen de catre organism;
E. Cu generare de cetonurie.
27. Cetogeneza:
A. Poate fi privita ca un mecanism alternativ de oxidare a acizilor grasi in conditiile saturarii ciclului Krebs cu acetil CoA;
B. Este o cale metabolica patologica;
C. Este o cale metabolica care se poate exacerba in anumite conditii fiziologice sau patologice;
D. Este exacerbata numai in situatii patologice;
E. Se produce in matricea mitocondriala a celulelor hepatice.
28. Deficienta in carnitina:
A. Se poate intalni la nou-nascutii prematuri;
B. Se poate trata prin administrare orala de carnitina;
C. Induce acidoza si cetonurie;
D. Induce hipercetonemie;
E. Impiedica un transport optim al acizilor grasi in mitocondrie.
29. Reglarea cetogenezei prin activitatea carnitin palmitoil transferazei I hepatice presupune:
A. Stabilirea exacta a proportiei dintre cantitatea de acizi grasi metabolizati prin oxidare si cantitatea de acizi grasi care sunt esterificati cu glicerol;
B. Controlul eliminarii acetonei pe cale pulmonara;
C. Inhibitia acestei enzime atunci cand perioada de repaus alimentar se prelungeste excesiv;
Page 45 of 120
D. Activarea lipazelor din tesutul adipos;
E. Activarea acestei enzime in perioadele de repaus alimentar.
30. Dintre corpii cetonici:
A. Acetoacetatul este cu usurinta oxidat in tesuturile extrahepatice;
B. Acetona este cel mai usor de utilizat drept molecula combustibil de catre tesuturile extrahepatice;
C. D(-) 3 hidroxibutiratul este dificil de oxidat in vivo si se evapora in mari cantitati prin plamani;
D. D(-) 3 hidroxibutiratul este cu usurinta oxidat in tesuturile extrahepatice;
E. Nici unul din corpii cetonici nu este utilizat drept molecula-combustibil.
10. Etapa initiala in sinteza acizilor grasi este:
A. Sinteza acetil-Co A
B. Sinteza aceto acetil-Co A;
C. Sinteza geranil-Co A;
D. Sinteza mevalonatului;
E. Sinteza malonil-Co A.
Complement Multiplu
11. Etapa initiala a biosintezei de novo a acizilor grasi:
A. Are loc in reticulul endoplasmatic hepatic;
B. Este catalizata de o enzima allosterica avand drept coenzima piridoxal fosfatul;
C. Consta in sinteza malonil-CoA pornind de la acetil CoA si HCO3-;
D. Este catalizata de o enzima avand drept coenzima biotina;
Page 48 of 120
E. Se desfasoara in citosol.
12. Procesul de elongare a catenei acidului palmitic provenit prin sinteza de novo a acizilor grasi:
A. Are loc in reticulul endoplasmatic hepatic;
B. Se produce prin adaugare la catena acil gras-Co A de unitati de 2 atomi de carbon prin intermediul malonil Co A;
C. Se produce prin adaugare la catena acil gras-Co A de unitati de 2 atomi de carbon prin intermediul acetil Co A;
D. Utilizeaza ca agent reducator NADPH, H+;
E. Utilizeaza ca agent reducator NADH, H+.
13. Ateroscleroza este caracterizata ca:
A. depunere a colesterolului sau a esterilor colesterolului din lipoproteinele care contin apo B-100 in tesutul conjunctiv al peretilor arteriali;
B. depunere a colesterolului sau a triacil glicerolilor din chilomicroni in tesutul conjunctiv al peretilor arteriali;
C. depunere a lipidelor polinesaturate din lipoproteinele care contin apo B-48 in tesutul conjunctiv al peretilor arteriali;
D. depunere a colesterolului sau a esterilor colesterolului din lipoproteinele de tip LDL, VLDL sau IDL in tesutul conjunctiv al peretilor arteriali;
E. depunere a colesterolului si glucidelor din lipoproteinele circulante in tesutul conjunctiv al peretilor arteriali.
14. Enzima-cheie a procesului de biosinteza de novo a colesterolului:
A. Catalizeaza sinteza mevalonatului;
B. Este inhibata de colesterolul in exces adus prin aport alimentar;
C. Are ca substrat geranil pirofosfatul;
D. Utilizeaza ca donor de potential reducator NADH, H+;
E. Utilizeaza ca donor de potential reducator NADPH, H+.
15. Acizii biliari primari:
A. Sunt sintetizati la nivelul ficatului;
B. Sunt sintetizati la nivelul veziculei biliare;
C. Sunt acidul colic si acidul chendeoxicolic;
Page 49 of 120
D. Sunt acidul colic si acidul deoxicolic;
E. Sunt sintetizati pe baza de potential reducator adus de NADH, H+.
16. Procesul de sinteza al eicosanoizilor:
A. Are ca substrat comun arahidonatul;
B. Presupune aport alimentar de linoleat;
C. Presupune aport alimentar de stearat;
D. Are ca substrat comun 5, 8, 11, 14 eicosantetraenoatul;
E. Nu are loc in celulele animalelor superioare.
17. Deficitul de vitaminăC:
A. Interferă în sinteza acizilor biliari în reacţia de hidroxilare iniţială;
B. Conduce in final la acumularea de colesterol şi la ateroscleroză;
C. Nu afecteaza in mod direct metabolismul colesterolului;
D. Are consecinte asupra procesului excretiei colesterolului;
E. Nu are consecinte in procesul excretiei colesterolului atunci cand exista NADPH suficient in celule.
18. Despre procesul de excretie al colesterolului sunt adevarate afirmatiile:
A. Colesterolul se elimina din organism numai sub forma esterificata;
B. Jumatate din colesterolul excretat este eliminat prin fecale, dupa transformarea sa in acizi biliari;
C. Jumătate din colesterolul excretat este eliminat ca atare;
D. Principala forma de eliminare a colesterolului sunt agregatele HDL;
E. Colesterolul se elimina din organism conjugat cu acid glucuronic.
19. Hipercolesterolemia familialăeste:
A. O maladie infectioasa;
B. O maladie în care colesterolul este depozitat în diverse ţesuturi datorităconcentraţiei mari în colesterol a HDL plasmatice;
C. O maladie ereditara;
D. O maladie in care apar noduli de colesterol subcutani numiti xantomas;
Page 50 of 120
E. O maladie care are drept consecinte ateroscleroza, cauzata de depozitarea unui strat de colesterol pe peretii vaselor sanguine şi ingustarea lor.
20. Ateroscleroza este:
A. Depunere a colesterolului sau a esterilor colesterolului din lipoproteinele care conţin apo B100 proteine în ţesutul conjunctiv al pereţilor arteriali;
B. Corelata cu nivelul colesterolului seric;
C. O consecinta a defictului genetic care provoaca hipercolesterolemia familiala;
D. Corelata cu concentratia acizilor biliari primari;
E. Corelata cu nivelul celular al acetil CoA.
21. Controlul enzimatic al procesului de biosinteza de novo a colesterolului este realizat prin urmatoarele
mecanisme:
A. Inhibitia biosintezei mARN specific al HMG-CoA reductazei in prezenta excesului de steroli;
B. Inhibitia biosintezei ADN specific al HMG-CoA reductazei in prezenta excesului de colesterol esterificat;
C. Inhibitia biosintezei mARN specific al enzimei-cheie a procesului in prezenta excesului de steroli;
D. Inhibitia vitezei procesului de translatie al informatiei mARN specific enzimei-cheie a procesului in prezenta metabolitilor nesterolici derivati de la mevalonat;
E. Numarul receptorilor membranari pentru LDL.
22. Controlul enzimatic al procesului de biosinteza de novo a colesterolului este realizat prin urmatoarele
mecanisme:
A. Cresterea vitezei de degradare a HMG-CoA reductazei in prezenta unui continut scazut de derivati ai colesterolului si de mevalonat;
B. Cresterea vitezei de degradare a HMG-CoA reductazei in prezenta unui continut ridicat de derivati ai colesterolului si de mevalonat;
C. Modularea capacitatii catalitice a HMG-CoA reductazei prin fosforilare-defosforilare;
D. Amplificarea biosintezei colesterolului o data cu scaderea concentratiei de ATP;
E. Inhibarea HMG-CoA reductazei de catre excesul de colesterol adus prin aport alimentar la nivelul ficatului.
23. Controlul procesului de biosinteza de novo a colesterolului este realizat prin urmatoarele mecanisme:
A. Modularea vitezei de sinteza a proteinelor-receptor membranar pentru LDL prin elementul de reglare (SRE) situat pe gena responsabila de sinteza acestei proteine;
B. Absenta patologica a receptorilor LDL;
Page 51 of 120
C. Inhibitia HMG-CoA reductazei prin fosforilare sub actiunea unei protein kinaze-AMP dependente;
D. Marirea aportului de oxigen necesar sintezei prin amplificarea ritmului respirator;
E. Aparitia de noduli de colesterol subcutani si la nivelul articulatiilor.
24. Controlul procesului de biosinteza de novo a colesterolului:
A. Este foarte strict in organism, deoarece excesul provoaca stari patologice, contribuind la formarea placii aterosclerotice;
B. Este reglat numai la nivelul ficatului, organul in care se realizeaza biosinteza acestui compus;
C. Poate fi dereglat prin mutatii genetice;
D. Se realizeaza exclusiv la nivelul enzimei-cheie a procesului;
E. Implica formarea de xantomas la nivelul tendoanelor.
25. Transportul colesterolului in tesuturi:
A. Se realizeaza prin intermediul lipoproteinelor;
B. Se realizeaza de la nivelul ficatului prin intermediul VLDL, care contin exclusiv colesterol liber;
C. Se realizeaza de la nivelul ficatului prin intermediul VLDL, care contin colesterol liber si colesterol esterificat;
D. Se realizeaza de la nivelul ileonului prin intermediul chilomicronilor;
E. Se realizeaza de la nivelul ileonului prin intermediul HDL.
26. Boli in care se inregistreaza nivele sanguine ridicate de: VLDL, IDL si reziduuri de chilomicroni sau LDL sunt:
A. Hipertiroidismul;
B. Hepatita virala cu virus A;
C. Diabetul zaharat;
D. Nefroza lipidica;
E. Anemia pernicioasa.
27. Acizii biliari primari:
A. Sunt deversati in vezica biliara sub forma libera;
B. Sunt deversati in vezica biliara sub forma conjugata cu glicina sau taurina;
C. Se gasesc in secretia biliara sub forma de saruri de calciu si magneziu;
Page 52 of 120
D. Se gasesc in secretia biliara sub forma de saruri de sodiu si potasiu;
E. Necesita pentru sinteza O2 si NADPH.
28. Colesterolul este:
A. Un compus intalnit in toate organismele vii de pe Pamant;
B. Precursorul de biosinteza al hormonilor corticosteroizi;
C. Precursorul de biosinteza al hormonilor tiroidieni;
D. Precursorul de biosinteza al hormonilor sexuali;
E. Un compus ubicuitar la organismele eucariote.
29. In legatura cu sinteza mevalonatului, in cadrul caii metabolice de biosinteza a colesterolului, sunt adevarate:
A. Reactia este catalizata de HMG CoA reductaza;
B. Reactia este ireversibila;
C. Reactia consuma potential reducator adus de FADH2:
D. Precursorii de sinteza sunt acetil-CoA si metil malonil-CoA;
E. Reactia are loc in citosol.
30. In cadrul caii metabolice de biosinteza a colesterolului, mevalonatul:
A. Este transformat in epoxid de squalen prin trei reactii consecutive, care consuma 3 molecule ATP;
B. Trece in izopentenil pirofosfat pe baza energiei furnizate de hidroliza a 3 legaturi macroergice din ATP;
C. Este sintetizat pornind de la acetil CoA si aceto acetil CoA;
D. Este izomerizat in dimetil alil pirofosfat;
E. Este un modulator al vitezei de desfasurare a caii metabolice.
Capitolul 8. Metabolismul lipidelor. Notiuni privind metabolismul acil glicerolilor
si al sfingolipidelor, aspecte clinice. Schema generala a repartizarii si utilizarii
lipoproteinelor in organism, aspecte clinice privind dislipidemiile
Complement Simplu
1. Utilizarea glicerolului rezultat in urma hidrolizei acilglicerolilor depinde de:
Page 53 of 120
A. Lipazele hormon sensibile din tesutul adipos;
B. Lipazele independente de hormoni;
C. Tipul de acizi grasi care au esterificat glicerolul;
D. Prezenta in tesut a glicerol kinazei;
E. Corecte A. si C.
2. Sinteza ceramidelor are loc in:
A. Mitocondrii;
B. Citosol;
C. Nucleol;
D. Ribozomi;
E. Reticulul endoplasmatic.
3. Sinteza VLDL (very low density lipoproteins) se desfasoara la nivelul:
B. Legarea acizilor grasi eliberati din esteri de albumina serica;
C. Hidroliza lor, catalizata de catre lipaze specifice;
D. Eliberarea acizilor grasi in plasma sanguina;
E. Includerea lor in agregate lipoproteice.
5. Maladia Tay-Sachs se manifesta din punct de vedere biochimic prin:
A. Absenta patologica a receptorilor membranari pentru LDL;
Page 54 of 120
B. Deficitul de hexozaminidaza A, cu acumularea de lipide glicozilate in organism;
C. Deficitul de carbamoil fosfat sintaza I;
D. Acidemie arginino-succinica moderata si acidurie;
E. Nici o varianta corecta.
6. Maladia Niemann-Pick se manifesta din punct de vedere biochimic prin:
A. Deficitul de sfingomielinaza, cu acumularea de sfingomielina;
B. Deficitul de HMG CoA reductaza;
C. Deficitul de chilomicroni in plasma sanguina;
D. Triacilglicerolemie;
E. Deficitul de ceramide tisulare.
7. Dintre cele cinci tipuri de lipoproteine, in tesutul hepatic sunt sintetizate:
A. Chilomicronii si VLDL;
B. VLDL si HDL;
C. LDL si HDL;
D. IDL si LDL;
E. Toate.
8. Modificarile biochimice care survin in maladia Tangier constau in:
A. Deficiente in biosinteza sfingozinei;
B. Deficiente in biosinteza ceramidelor;
C. HDL reduse ca numar sau absente in plasma sanguina;
D. Mobilizarea excesiva a acizilor grasi din tesutul adipos de catre insulina si de subutilizarea lipidelor din chilomicroni sl LDL;
E. Sintetizarea unor chilomicroni de dimensiuni prea mari.
9. Dintre cele cinci agregate lipoproteice circulante, cele mai mari dimensiuni le au:
A. VLDL;
B. Chilomicronii;
C. IDL;
Page 55 of 120
D. HDL;
E. LDL.
10. Hipolipoproteinemie (a beta lipoproteinemie) are ca defect molecular:
A. Lipsa chilomicronilor, a VLDL, sau LDL datorita defectelor de “incarcare” cu lipide ale apo proteinei B;
B. Deficienta in receptori LDL sau mutatii aparute la nivelul ligandului (apo B-100);
C. Mobilizarea excesiva a acizilor grasi din tesutul adipos de catre insulina si de subutilizarea lipidelor din chilomicroni sl LDL;
D. Deficiente de inlaturare a agregatelor reziduale de catre ficat, datorita une apo E anormale;
E. Deficitul de sfingomielinaza, cu acumularea de sfingomielina.
Complement Multiplu
11. In cursul etapelor de sinteza a ceramidelor:
A. 3 cetosfinganina este transformata printr-un proces reductiv in dihidrosfingozina;
B. 3 cetosfinganina este transformata printr-un proces oxidativ in dihidrosfingozina;
C. Dihidrosfingozina este transformata in dihidroceramida de catre enzima dihidrosfingozin N-acil transferaza;
D. Dihidrosfingozina este transformata in dihidroceramida de catre enzima hidroximetil glutaril Co A reductaza;
E. se realizeaza sinteza ceramidelor pornindu-se de la metionina si hidroximetil glutaril Co A.
12. Sfingolipidele:
A. Sunt sintetizate pornind de la ceramide;
B. Au ca precursori de sinteza palmitoil Co A si serina;
C. Sunt sintetizate pe aceiasi cale ca si colesterolul;
D. Isi incep sinteza in reticulul endoplasmatic;
E. Au ca si precursori de sinteza metionina si malonil Co A.
13. Glicerol kinaza se gaseste in cantitati mari in:
A. Tesut adipos brun;
B. Muschi cardiac;
Page 56 of 120
C. Glanda mamara;
D. Tesut nervos;
E. Ficat.
14. Hipertriacilglicerolemia este disfunctia lipidica:
A. Cel mai des intalnita in insuficienta renala;
B. Cel mai des intalnita in diabetul zaharat;
C. Cauzata de mobilizarea excesiva a acizilor grasi din tesutul adipos de catre insulina si de sub-utilizarea lipidelor din chilomicroni sl LDL;
D. Cauzata de mobilizarea excesiva a acizilor grasi din tesutul adipos de catre insulina si de sub-utilizarea lipidelor din chilomicroni sl VLDL;
E. Specifica maladiei Niemann-Pick.
15. Deficienta familiala de lecitin-colesterol acil transferaza (LCAT):
A. Duce la acumularea de HDL mari, bogate in triacilgliceroli si de reziduuri VLDL;
B. Duce la supraproductie de VLDL deseori asociata cu intoleranta la glucoza;
C. Duce la pierderea capacitatii HDL de a prelua colesterolul si de a-l esterifica;
D. Are ca manifestare scaderea concentratiei plasmatice de colesterol esterificat;
E. Se asociaza de obicei cu maladii cardiace, diabet, obezitate, alcoolism.
16. Despre apoproteina Apo B-100 sunt adevarate:
A. Se gaseste in LDL;
B. Se gaseste in VLDL;
C. Se gaseste in chilomicroni;
D. Este ligand pentru receptorii LDL;
E. Este ligand pentru receptorii HDL.
17. Despe apoproteina Apo-A I sunt adevarate:
A. Se gaseste in LDL;
B. Se gaseste in chilomicroni;
C. Este activator al lecitin: colesterol acil transferazei (LCAT);
Page 57 of 120
D. Este prezentăîn exces în VLDL la pacienţii cu hiperlipoproteinimii ;
E. Este ligand pentru receptorii HDL.
18. In scleroza grasa a ficatului:
A. Se inregistreaza o ratădezechilibratăde formare şi eliberare a triacil glicerolilor la nivelul ţesutului hepatic;
B. In ficat se acumuleaza triacilgliceroli in cantitati mari;
C. Insulina cauzeaza o mobilizare excesivăa acizilor graşi liberi din ţesutul adipos;
D. Are loc o sub-utilizare a chilomicronilor şi VLDL;
E. Se observa hipolipemii.
19. IDL (lipoproteine cu densitate intermediară):
A. Sunt produse la nivel intestinal, in urma absorbtiei lipidelor din aport alimentar;
B. Sunt un produs al metabolismului lipoproteinelor în plasmă;
C. Au dimensiuni cuprinse intre 25-30 nm;
D. Au dimensiuni cuprinse intre 100-1000 nm;
E. Au dimensiuni cuprinse intre 7,5-10 nm.
20. Lipoproteinele plasmatice:
A. Transporta lipidele in torentul sanguin;
B. Contin aceiasi proportie de proteine si lipide, indiferent de tip si de dimensiuni;
C. Contin proteine identice, cu rol de transport;
D. Sunt sintetizate de tesutul adipos;
E. Cresc solubilitatea lipidelor şi permit transportul lor către toate celulele corpului 231 Viteza de eliberare a acizilor grasi din tesutul adipos in plasma este accelerata de:
21. Insulina:
A. Interfera in mica masura in mecanismele de reglare ale metabolismului tesutului adipos;
B. Inhiba eliberarea acizilor grasi liberi din tesutul adipos;
C. Amplifica sinteza acil glicerolilor;
D. Activeaza lipazele din tesutul adipos;
Page 58 of 120
E. Amplifica eliberarea acizilor grasi liberi din adipocite.
22. Insulina:
A. Reduce eliberarea de glicerol din adipocite;
B. Are ca efect cresterea concentratiei de acizi grasi liberi in plasma sanguina;
C. Actioneaza antagonist glucagonului in procesele de reglare a concentratiei de acizi grasi liberi in plasma sanguina;
D. Actioneaza sinergic glucagonului in procesele de reglare a concentratiei de acizi grasi liberi in plasma sanguina;
E. Actioneaza antagonist catecolaminelor in procesele de reglare a concentratiei de acizi grasi liberi in plasma sanguina.
23. Scleroza grasa a ficatului:
A. Poate fi asociata cu cresterea concentratiei de acizi grasi liberi in plasma;
B. Poate fi asociat si amplificat de deficitul de vitamina E;
C. Poate fi asociat si amplificat de deficitul de vitamina C;
D. Se datoreaza acumularii de triacil gliceroli in tesutul hepatic peste limitele de incorporare ale acestora in VLDL;
E. Este asociata cu excesul de insulina.
24. Perturbarile metabolismului lipidic in diabetul zaharat:
A. Constau in diminuarea eliberarii de acizi grasi liberi din tesuturile de depozit;
B. Se datoreaza diminuarii secretiei de insulina, principalul hormon responsabil de controlul metabolismului tesutului adipos;
C. Se datoreaza cresterii secretiei de cortisol, principalul hormon responsabil de controlul metabolismului tesutului adipos;
D. Pot consta in unele cazuri in afectarea capacitatii ficatului de a sintetiza VLDL;
E. Pot consta in unele cazuri in afectarea capacitatii ficatului de a sintetiza chilomicroni.
25. In procesul biosintezei acil glicerolilor:
A. Prima etapa implica sinteza unui acid fosfatidic;
B. Glicerolul supus procesului de esterificare este activat sub forma de glicerol 3 fosfat;
C. Glicerolul supus procesului de esterificare este activat sub forma de glicerol 2 fosfat;
Page 59 of 120
D. Glicerolul liber este esterificat cu doua molecule de acizi grasi;
E. Acizii grasi care participa la reactia de esterificare sunt activati sub forma de acil gras CoA.
26. Dintre dereglarile de sinteza ale sfingolipidelor datorate unor cauze genetice fac parte:
A. Maladia Cori;
B. Sindromul McArdle;
C. Neuropatia Leber;
D. Maladia Tay-Sachs;
E. Maladia Niemann-Pick
27. Despre VLDL sunt adevarate:
A. Sunt sintetizate exclusiv in intestinul subtire;
B. Au dimensiunile cele mai mici, comparativ cu celelealte lipoproteine;
C. Sunt sintetizate in ficat;
D. Au dimensiuni cuprinse intre 30-70 nm;
E. Contin cel mai redus procent de proteine dintre toate particulele lipoproteice plasmatice;
28. Despre chilomicroni sunt adevarate:
A. Sunt sintetizati in intestin;
B. Contin cel mai redus procent de proteine dintre toate particulele lipoproteice plasmatice;
C. Au dimensiuni cuprinse intre 30-70 nm;
D. Au dimensiunile cele mai mari, comparativ cu celelealte lipoproteine;
E. Sunt produsi prin metabolizarea VLDL.
29. Despre HDL sunt adevarate:
A. Sunt produse ale metabolizarii VLDL;
B. Contin cel mai mare procent de proteine dintre toate particulele lipoproteice plasmatice;
C. Contin apolipoproteina Apo A-I;
D. Nu contin colesterol liber sau esterificat;
E. Au cele mai mici dimensiuni dintre toate particulele lipoproteice plasmatice.
Page 60 of 120
30. Viteza de eliberare a acizilor grasi din tesutul adipos in plasma este accelerata de:
A. Hormonul adenocorticotrop (ACTH);
B. Catecolamine;
C. Insulina;
D. Hormonul de stimulare tiroidian;
E. Estradiol.
Capitolul 9. Metabolismul aminoacizilor si al proteinelor. Catabolismul
proteinelor si eliminarea azotului aminic; turn-overul proteic; ciclul ureogenetic-
etape, reactii, aspecte clinice. Consideratii generale privind catabolismul
scheletelor hidrocarbonate ale aminoacizilor, aspecte clinice
Complement Simplu
1. Prima etapa a biosintezei ureei este:
A. Deaminarea oxidativa a glutamatului;
B. Ciclul ureogenetic hepatic;
C. Transportul la ficat al amoniacului;
D. Transaminarea;
E. Glicozilarea proteinelor.
2. Viteza de degradare a unei molecule proteice este exprimatăprin:
A. Raportul dintre absorbţia totalăde azot şi eliminarea sa;
B. Continua degradare şi re-sintetizare a proteinelor celulare;
C. Perioada de timp necesarăpentru reducerea concentraţiei sale la 50% din valoarea iniţială;
D. Recircularea aminoacizilor din componenta sa;
E. Nici o varianta corecta.
3. Balanta azotului reprezinta:
A. Raportul dintre absorbţia totalăde azot şi eliminarea sa;
Page 61 of 120
B. Continua degradare şi re-sintetizare a proteinelor celulare;
C. Absorbtia totala de azot din aport alimentar;
D. Eliminarea de azot prin urina si fecale;
E. Corecte B. si D.
4. Apoptoza reprezinta:
A. O proteina produsa de o gena supresor;
B. O proteina virala;
C. Moartea genetic programata a celulei;
D. Un proces indus de genomul unor virusuri, ca urmare a infectarii celulei;
E. Nici o varianta corecta.
5. Secventele PEST din unele lanturi polipeptidice ale unor proteine:
A. Constituie ţinte specifice pentru enzimele proteolitice ;
B. Sunt specifice moleculelor proteice cu timp lung de injumatatire ;
C. Sunt prezente in toate proteinele ;
D. Sunt terminatii de acid sialic fixate pe lanturile polipeptidice ;
E. Nici o varianta corecta.
6. Ubiquitina este:
A. O proteina cu rolul de a prelungi timpul de injumatatire a moleculelor proteice;
B. O proteina cu rol de a lega şi derivatiza numeroase proteine intracelulare destinate degradării;
C. O proteina virala specifica, care impiedica procesul natural de apoptoza;
D. O proteina cu structura quaternara si secventa inalt variabila, in functie de specie;
E. Nici o varianta corecta.
7. Animalele terestre şi oamenii sunt organisme: A. Uricotele;
A. Amonotele;
B. Uricotele si amonotele;
C. Ureotele;
Page 62 of 120
D. Nici un raspuns corect.
E. 248 Prima etapa a ureogenezei este:
8. Prima reactie a ciclului ureogenetic constăîn:
A. Condensarea CO2, NH3 şi ATP cu formare de carbamoil fosfat;
B. Biosinteza glutaminei de către glutamin sintază;
C. Transferul gruparilor amino către αcetoglutarat cu formare de L glutamat;
D. Transferul grupărilor α-amino de la aminoacizii supuşi degradării către αcetoacizi;
E. Nici o varianta corecta.
9. Transferul gruparilor amino de pe moleculele aminoacizilor supusi deaminarii se face pe:
A. Moleculele glutamatului;
B. Moleculele alaninei;
C. Moleculele oxaloacetatului;
D. Moleculele asparaginei si glutaminei;
E. Moleculele αcetoglutaratului şi piruvatului.
10. Prima etapa a ureogenezei este:
A. Deaminarea glutamatului;
B. Transaminarea aminoacizilor supusi degradarii oxidative;
C. Transaminarea aminoacizilor din lanturile polipeptidice;
D. Deaminarea alaninei;
E. Nici un raspuns corect.
Complement Multiplu
11. Procesul de transaminare, ca etapa a biosintezei ureei:
A. Realizeaza sinteze de aminoacizi neproteinogeni la nivelul citoplasmei hepatocitelor;
B. Constituie etapa in care se sintetizeaza ureea la nivelul hepatocitului;
C. Realizeaza “colectarea” gruparilor αamino din moleculele aminoacizilor proteinogeni;
D. Este catalizat de enzime specifice denumite transaminaze sau aminotransferaze;
E. Este catalizat de enzime hepatice specifice din care face parte si glutamin sintaza.
Page 63 of 120
12. Etape ale ciclului ureogenetic sunt:
A. Sinteza carbamoil fosfatului, catalizata de carbamoil fosfat sintaza I;
B. Sinteza acidului L-aspartic pornind de la citrulina;
C. Sinteza ornitinei catalizata de ornitin-carbamoil transferaza;
D. Sinteza ureei din L-arginina, catalizata de arginaza;
E. Sinteza ureii din L-citrulina catalizata de citrinaza.
13. Simptomele clinice comune tuturor deficientelor metabolice ale ciclului ureogenetic cuprind:
A. Ataxie intermitenta, letargie si retardere metala;
B. Intoleranta fata de alimentele cu continut proteic ridicat;
C. Hiperglicemie si hipercolesterolemie;
D. Formarea de noduli de colesterol subcutani (xantomas);
E. Miros specific de mere putrede al respiratiei.
14. Alanin-transaminaza: A. Poate avea ca substrat glutamatul;
A. Poate avea ca substrat piruvatul;
B. Poate avea ca substrat alanina;
C. Poate avea ca substrat oxoglutaratul;
D. Are ca si coenzima tiamina.
E. 255 Eliberarea azotului din grupările αamino ale majorităţii aminoacizilor, captat in glutamat sub formăde amoniac:
15. Amoniacul produs în ţesuturi prin deaminarea aminoacizilor:
A. Este transportat ca atare prin sange pana la ficat;
B. Este transportat prin sange pana la rinichi, unde este convertit in uree;
C. Este rapid eliminat prin biosinteza glutaminei de către glutamin sintază;
D. Constituie un toxic foarte puternic;
E. Este transformat in glutamina cu formarea concomitenta a unei molecule de ATP.
16. Referitor la transportul ionului amoniu rezultat din deaminarea oxidativa a aminoacizilor, sunt adevarate:
Page 64 of 120
A. Vehiculele azotului prin plasma sanguina sunt alanina si glutamina;
B. Alanina este reţinutăin ficat;
C. Glutamina este reţinutăin sange;
D. Ionul amoniu circula liber in plasma sanguine;
E. Ionul amoniu circula in plasma sanguine complexat cu albumina serica.
17. Reactia catalizata de carbamoil fosfat sintaza in cadrul caii ureogenetice:
A. Are loc in mitocondrie;
B. Este o reactie reversibila;
C. Are loc in citosol;
D. Are loc cu aportul energetic rezultat din hidroliza a doua molecule ATP;
E. Conduce la formarea carbamoil fosfatului.
18. In ciclul ureogenetic, reactia catalizata de arginino succinat sintetaza:
A. Este reversibila;
B. Este ireversibila;
C. Are loc cu aportul energetic adus de ATP;
D. Are loc cu aport ul energetic adus de carbamoil fosfat;
E. Conduce la formarea de L-arginino-succinat.
19. In reactia de sinteza a citrulinei in cadrul ciclului ureogenetic:
A. Enzima care catalizeaza procesul este o enzima mitocondriala;
B. Enzima care catalizeaza procesul este o enzima citosolica;
C. Se transfera o grupare carbamoil activata;
D. Energia reactiei este furnizata de hidroliza unei legaturi macroergice din molecula carbamoil fosfatului;
E. Energia necesara reactiei provine din hidroliza unei molecule ATP. 261 In ciclul ureogenetic argininosuccinaza catalizeaza:
20. Arginaza:
A. Este o enzima specifica ciclului ureogenetic;
Page 65 of 120
B. Este o hidrolaza;
C. Are ca produs de reactie ureea;
D. Se gaseste exclusiv in tesutul hepatic;
E. Este o liaza.
21. Sunt aminoacizi exclusiv glicogenici (ale caror schelete hidrocarbonate se converteste in molecule glucidice):
A. Leucina;
B. Izoleucina;
C. Metionina;
D. Acidul glutamic;
E. Alanina.
22. Sunt aminoacizi glicogenici si cetogenici (ale caror schelete hidrocarbonate se converteste in molecule glucidice
si lipidice):
A. Leucina;
B. Izoleucna;
C. Valina;
D. Triptofanul;
E. Prolina.
23. Prin degradarea scheletului hidrocarbonat al diferitilor aminoacizi dupa deaminare rezulta:
A. Oxaloacetat din asparagina;
B. α cetoglutarat din acid aspartic;
C. α cetoglutarat din prolina;
D. Piruvat din alanina;
E. Piruvat din arginina.
24. Prin degradarea scheletului hidrocarbonat al diferitilor aminoacizi dupa deaminare rezulta:
A. α cetoglutarat din serina;
B. α cetoglutarat din histidina;
C. α cetoglutarat din glicina;
Page 66 of 120
D. Piruvat din cisteina;
E. Oxaloacetat din glutamina.
25. Sunt catabolizate la succinil CoA:
A. Triptofanul;
B. Fenil alanina;
C. Metionina;
D. Izoleucina;
E. Valina.
26. Desfăşurarea defectuoasăa catabolizării aminoacizilor cu catenăramificatăgenereazămaladii cum sunt:
A. Acidemia izovalerianică;
B. Tirozinoza;
C. Sindromul Tangier;
D. Alcaptonuria;
E. Neuropatia Warnike.
27. Fenilcetonuria:
A. Este o maladie genetica cauzata de acumularea de fenil piruvat;
B. Este cauzata de perturbarea caii biosintetice a fenil alaninei;
C. Poate fi tratata prin administrarea unei diete alimentare cu continut scazut in fenil alanina;
D. Poate fi tratata prin administrarea unei diete alimentare cu continut scazut in triptofan;
E. Este cauzata de perturbarea caii de catabolizare a triptofanului.
28. Boala urinei cu miros de sirop de artar:
A. Este consecinta a perturbarii catabolismului alaninei;
B. Este consecinta a perturbarii catabolismului leucinei;
C. Este consecinta a perturbarii biosintezei valinei;
D. Este consecinta a perturbarii catabolismului izoleucinei;
E. Este o maladie genetica autozaomala recesiva.
Page 67 of 120
29. Eliberarea azotului din grupările α amino ale majorităţii aminoacizilor, captat in glutamat
sub formă
de amoniac:
A. Este catalizată de L-glutamat dehidrogenază;
B. Este catalizată de glutamin sintaza;
C. Se poate face utilizand ca acceptor de hidrogen atât NAD+ cât şi NADP+;
D. Are loc utilizand ca acceptor de hidrogen FAD;
E. Este catalizata de o enzima care se gaseste in toate tesuturile.
30. In ciclul ureogenetic argininosuccinaza catalizeaza:
A. O reactie de transaminare;
B. O reactie ireversibila;
C. O reactie cu formare de acid fumaric;
D. O reactie cu formare de L-arginina;
E. O reactie cu formare de legaturi macroergice.
Capitolul 10. Metabolismul aminoacizilor si al proteinelor. Biosinteza proteica:
etape, reactii, modele de control
Complement Simplu
1. In cadrul biosintezei proteice, etapa de activare a aminoacizilor:
A. Necesita consumul a 2 legaturi macroergice/molecula aminoacid;
B. Necesita consumul unei legaturi macroergice/molecula aminoacid;
C. Incepe numai dupa atasarea aminoacidului la nivelul subunitatilor ribozomale;
D. Este catalizata de peptidil transferaza;
E. Presupune activarea initiala a metioninei formilate.
2. Etapa de terminare a biosintezei proteice:
A. Este semnalizata prin ocuparea situsului A ribosomal de catre un codon non-sens;
B. Este catalizata de peptidil transferaza;
Page 68 of 120
C. Este semnalizata prin ocuparea situsului P ribosomal de catre un codon non-sens;
D. Necesita consumul a doua legaturi macroergice;
E. Are loc exclusiv in mitocondrii.
3. Referitor la modalitatea de stocare a informatiei genetice la nivelul celulelor, este adevarat ca:
A. ADN este direct implicat in biosinteza proteica;
B. Fiecare aminoacid este reprezentat in molecula ARNm de un cuvant de cod (codon) alcatuit dintr-o secventa de trei nucleotide;
C. Catena ADN serveste ca matrita pentru sinteza unei catene complementare de ARNt;
D. Transcrierea informatiei genetice se face pe model neconservativ;
E. Fiecare aminoacid este reprezentat in molecula ADN de un cuvant de cod (codon) alcatuit dintr-o secventa de trei nucleotide.
4. Referitor la procesul de transcriptie a informatiei din ADN in ARN, este adevarat ca:
A. Lantul purtator de informatie din molecula ADN este transcris in ARN similar;
B. Lantul purtator de informatie din molecula ADN este transcris in ARN complementar;
C. La eucariote catena initiala de ARN copiaza exact catenele ADN;
D. O etapa a acestui proces o constituie transferul aminoacidului activat la ARNt;
E. Nici o afirmatie adevarata.
5. Ribozomii sunt:
A. Conglomerate moleculare citoplasmatice continand proteine si ADN;
B. Conglomerate moleculare citoplasmatice continand proteine si ARN;
C. Complexe lipoproteice;
D. Homoproteine cu structura cuaternara;
E. Complexe enzimatice transmembranare.
6. Enzima care catalizează formarea legăturii peptidice in procesul biosintezei proteice este localizată:
A. Pe ARNm;
B. Pe subunitatea mica a ribozomilor;
C. Pe subunitatea mare a ribozomilor;
Page 69 of 120
D. In citoplasma;
E. In mitocondrie.
7. Peptidil transferaza:
A. Necesita pentru functionare UTP;
B. Necesita pentru functionare GTP;
C. Necesita pentru functionare ATP;
D. Este o hidrolaza;
E. Este o oxidoreductaza.
8. Dintre antibioticele cu efect de inhibitori ai traducerii informatiei genetice face parte:
A. Penicilina;
B. Rifampicina;
C. Tetraciclina;
D. Cefalosporina;
E. Actinomicina D.
9. Definitia operonului este:
A. O gena care codifica trei proteine si se transcrie într-un singur ARNm;
B. Un grup de trei gene care codifica proteine similare;
C. Un grup de gene care realizeaza controlul procesului de biosinteza proteica prin intermediul ARNm;
D. Un grup de gene adiacente care se transcriu într-un singur ARNm;
E. Nici o definitie corecta.
10. Segmentul de ADN care conţine toatăinformaţia pentru producerea unui singur polipeptid este numit:
A. Operon;
B. Policistron;
C. Cistron;
D. Operon lac;
E. Nici o denumire corecta.
Page 70 of 120
Complement Multiplu
11. Etapa de elongare a lantului polipeptidic din cadrul procesului de biosinteza proteica:
A. Implica atasarea Aminoacil-ARNt pe situsul P ribosomal;
B. Presupune formarea unei legaturi peptidice printr-un atac nucleofil al gruparii αamino a unuia din aminoacizi asupra gruparii carboxil a celui de-al doilea aminoacid;
C. Presupune formarea unei legaturi peptidice printr-un atac electrofil al gruparii αamino a unuia din aminoacizi asupra gruparii carboxil a celui de-al doilea aminoacid;
D. Implica atasarea Aminoacil-ARNt pe situsul A ribosomal;
E. Implica participarea enzimei peptidil transferaza.
12. Etapa de initiere a sintezei lantului polipeptidic din cadrul procesului de biosinteza proteica:
A. Necesita la eucariote prezenta unor factori de initiere cu rolul de a pozitiona corect Nformil Metionina-ARNt pe situsul P ribosomal;
B. Necesita fixarea pe situsul P a ARNt initiator;
C. Este consecutiva etapei de activare a aminoacizilor;
D. Este consecutiva etapei de elongare a lantului polipeptidic;
E. Se mai numeste si sinteza ectopica.
13. Biosinteza proteinelor:
A. Presupune prezenta UTP ca furnizor de energie;
B. Cuprinde patru etape distincte: activarea aminoacizilor, activarea ribozomilor, initierea sintezei lantului polipeptidic si elongarea lantului polipeptidic;
C. Are loc la nivelul mitoribozomilor celulelor procariote;
D. Are loc la nivelul ribozomilor;
E. Presupune prezenta GTP ca furnizor de energie.
14. Despre modelul operon lac sunt adevarate urmatoarele afirmatii:
A. Operonul este definit ca un grup de gene situate pe cromozomi diferiti care se transcriu intr-un singur ARNm;
B. Constituie un model al controlului transcrierii ADN pentru producerea ARNm;
C. Presupune fixarea unei proteine represoare de o molecula-inductor;
D. Operonul contine informatie pentru sinteza exclusiva a proteinelor policistronice;
Page 71 of 120
E. Operonul cuprinde o gena reglatoare (i), si mai multe gene structurale.
15. Dintre premisele modelului de reglare a biosintezei proteice Jacob-Monod fac parte:
A. În timp ce proteina-represor este legatăde ADN, trascrierea nu poate avea loc pentru căsitusul de iniţiere a transcrierii este în segmentul promotor ;
B. Segmentul promotor P al ADN este punctul de iniţiere a sintezei ARNm policistronic;
C. Dacăinductorul nu este legat de proteina represoare, aceasta adoptăo structurăR, care nu se poate lega la segmentul operator (O) al ADN pentru a initia traducerea;
D. Spre capătul 3’ al ARNm se aflăsecvenţe specifice, numite secvenţe Shine-Delgarno, care permit ribozomilor săse desprindăireversibil de ARNm;
E. Operonul controleaza transcrierea ARNm pentru o singura molecula proteica.
16. Etapa de activare a aminoacizilor in vederea biosintezei proteinelor din aminoacizi liberi:
A. Necesita consumul unei legaturi macroergice din ATP;
B. Necesita consumul a doua legaturi macroergice din ATP;
C. Necesita cate o enzima specifica pentru fiecare din aminoacizii proteinogeni;
D. Are drept rezultat atasarea aminoacidului la capatul 5’ al ARNt;
E. Conduce la fixarea aminoacidului activat pe bratul anticodon al ARNt.
17. Etapa de activare a aminoacizilor in vederea biosintezei proteinelor din aminoacizi liberi:
A. Are loc prin activarea gruparii α-carboxil a aminoacidului;
B. Are loc prin activarea gruparii α-amino a aminoacidului;
C. Cuprinde trei reactii consecutive;
D. Cuprinde o singura reactie;
E. Cuprinde doua reactii consecutive.
18. Referitor la structura ribozomilor sunt adevarate:
A. Au structura similara la eucariote si procariote;
B. Au structura identica la eucariote si procariote;
C. Sunt formati din 2 subunităţi cu mărimi diferite;
D. Enzima care catalizează formarea legăturii peptidice este localizată într-o „vale” dintre douăprotuberanţe ale subunitatii mici;
Page 72 of 120
E. Peptidil transferaza este localizatăîntr-o „vale” dintre douăprotuberanţe ale subunitatii mari.
19. Referitor la structura ribozomilor sunt adevarate:
A. La eucariote subunitatea mare are coeficientul de sedimentare de 60S;
B. La eucariote subunitatea mare are masa moleculara 60;
C. La eucariote subunitatea mica are coeficientul de sedimentare 60S;
D. Cele doua subunitati ribozomale trebuie sa fie asamblate pentru ca ribozomul sa fie functional;
E. Pe subunitatea ribozomala mare este situat situsul de iesire al lantului polipeptidic in crestere.
20. Referitor la procesele din etapa de initiere a sintezei lantului polipeptidic:
A. Necesita la procariote mai multe proteine care au rolul de factori de iniţiere ai procesului;
B. Necesita la eucariote prezenta mai multor proteine care au rolul de a asigura o poziţionare corectă a ARNt-Met în situsul P al ribozomului (eIF);
C. Aminoacidul N-terminal al polipeptidelor nou iniţiate este la procariote metionina;
D. Aminoacidul N-terminal al polipeptidelor nou iniţiate este la eucariote metionina;
E. Factorii de initiere se asocieaza mai intai cu ARNm.
21. Etapa de initiere a sintezei lantului polipeptidic:
A. Incepe numai dupa ce ribozomul s-a asamblat;
B. Presupune si procesul de asamblare al ribozomului;
C. Necesita energie furnizata de hidroliza unei molecule ATP;
D. Necesita energie furnizata prin hidroliza unei molecule GTP;
E. Presupune fixarea complexului ARNt-Met pe situsul P din subunitatea ribozomala mica.
22. Etapa de elongare a lantului polipeptidic din procesul biosintezei proteice:
A. Implica fixarea complexului ARNt-aminoacid 2 pe situsul A ribozomal;
B. Necesita actiunea catalitica a unei translocaze pentru formarea legaturii peptidice;
C. Necesita energia cedata prin hidroliza unei molecule GTP;
D. Dupa formarea legatuii peptidice, peptidul nou format se muta pe situsul P;
E. Necesita energia cedata prin hidroliza unei molecule ATP.
Page 73 of 120
23. Etapa de elongare a lantului polipeptidic din procesul biosintezei proteice:
A. Necesita actiunea catalitica a peptidil transferazei;
B. Presupune formarea unei legaturi peptidice prin atacul nucleofil al gruparii αamino a aminoacidului 2 (de pe situsul ribozomal A);
C. Necesita prezenta factorului de eliberare;
D. Necesita la procariote prezenta a trei molecule proteice cu rol de factori de eliberare;
E. Incepe atunci cand in dreptul situsului A se gaseste codonul UAG.
24. Etapa de terminare a biosintezei proteice:
A. Necesita energie furnizata de GTP pentru dezasamblarea ribozomului;
B. Necesita la procariote doi factori de eliberare;
C. Este semnalizata de ocuparea situsului A de un codon non-sens;
D. Presupune hidroliza legaturii esterice peptidil-ARNt;
E. Presupune eliberarea ribozomului de pe ARNr.
25. Etapa de terminare a biosintezei proteice:
A. Necesita ATP pentru hidroliza legaturii esterice peptidil-ARNt;
B. Presupune disocierea ribozomului de ARNm;
C. Se incheie cu disocierea celor doua subunitati ribozomale;
D. Necesita la eucariote trei factori de eliberare;
E. Nu necesita aport energetic.
26. Referitor la actiunea antibioticelor asupra procesului de biosinteza a proteinelor, sunt adevarate:
A. Actiunile lor sunt in general indreptate inspre blocarea uneia dintre etapele biosintezei proteice;
B. Exista antibiotice cu efect de amplificatori ai traducerii informatiei genetice;
C. Penicilinele fac parte din antibioticele care se fixeaza la nivelul dublului helix ADN si interfera in procesul de replicare si transcriere a informatiei genetice;
D. Rifampicina face parte din antibioticele care se fixeaza la nivelul dublului helix ADN si interfera in procesul de replicare si transcriere a informatiei genetice;
E. Cefalosporinele fac parte din antibioticele care se fixeaza la nivelul dublului helix ADN si interfera in procesul de replicare si transcriere a informatiei genetice.
Page 74 of 120
27. Referitor la actiunea antibioticelor asupra procesului de biosinteza a proteinelor, sunt adevarate:
A. Antibioticele cu efect de inhibitori ai traducerii informatiei genetice inhiba sinteza peretelui celular al bacteriilor Gram-pozitive;
B. Rifampicina si cefalosporinele sunt inhibitori ai traducerii informatiei genetice;
C. Streptomicina si eritromicina sunt inhibitori ai traducerii informatiei genetice;
D. Rifampicina si actinomicina D au actiune de intercalatori;
E. Intercalatorii inhiba sinteza peretelui celular al bacteriilor Gram-pozitive.
28. Controlul sintezei proteice la eucariote:
A. Se poate realiza prin fosforilarea factorilor de iniţiere;
B. Se poate realiza prin intermediul hormonilor endocrini;
C. Este un proces identic cu cel de la procariote;
D. Se face la nivelul transcrierii de către porţiunea de ADN situatăîn aval faţăde situsul de transcriere;
E. Se face la nivelul transcrierii dupa modelul operonului lac.
29. Controlul sintezei proteice la eucariote include:
A. Existenta unor secvenţe speciale de ADN, numite „enhancers”, situate la distanţăde pânăla câteva sute de perechi de baze de promotor;
B. Recunoastera secventelor enhancers de catre proteine specifice denumite factori de transcriere;
C. Existenta unui mecanism unic;
D. Participarea unor antibiotice;
E. Actiunea de amplificare a sintezei exercitata de hormonii estrogeni.
30. Traducerea informatiei genetice la eucariote presupune:
A. Iesirea din nucleu a ARNm maturat;
B. Fixarea ARNm maturat pe ribozomi;
C. Replicarea ADN pe model semiconservativ;
D. Sinteza unui lant polipeptidic cu secventa conforma informatiei transmise de ARNm;
E. Sinteza ARNm dupa matrita ADN.
Page 75 of 120
Capitolul 11. Biosinteza hemului, catabolismul hemului, deficiente patologice si
aspecte clinice
Complement Simplu
1. Prima reactie in calea biosinteica a nucleuli hem este:
A. Sinteza porfobilinogenului;
B. Condensarea acetil CoA cu glicina;
C. Condensarea porfobilinogenului;
D. Sinteza δamino levulinatului;
E. Condensarea malonatului cu CO2.
2. Prima reactie in procesul de degradare al nucleului hem este:
A. Deschiderea inelului porfirinic si formarea biliverdinei catalizata de ferochelataza;
B. Deschiderea inelului porfirinic si formarea biliverdinei catalizata de hemoxigenaza;
C. Sinteza urobilinogenului prin reduceri successive catalizate de enzime bacteriene;
D. Conjugarea bilirubinei cu acidul glucuronic la nivelul tesutului hepatic;
E. Nici o varianta corecta.
3. Chelatarea unui ion Fe pentru sinteza nucleului hem:
A. Este catalizata de ferochelataza;
B. Are loc la nucleul uroporfirinogenului III;
C. Este catalizata de transferina;
D. Este catalizata de feritina;
E. Nici un raspuns corect.
4. Etapa-cheie in procesul biosintezei hemului este:
A. Reactia catalizata de porfobilinogen deaminaza;
B. Reactia catalizata de ferochelataza;
C. Reactia catalizata de protoporfirinogen oxidaza;
D. Reactia catalizata de δamino levulinat dehidrogenaza;
Page 76 of 120
E. Reactia catalizata de δamino levulinat sintaza.
5. Initierea reactiei de condensare a glicinei cu succinil CoA consta in:
A. Condensarea cap-coada a celor doi compusi;
B. Formarea unei baze Schiff între tiamin pirofosfat şi glicină;
C. Dehidrogenarea succinatului;
D. Formarea unei baze Schiff între piridoxal fosfat şi glicină;
E. Decarboxilarea spontana a a amino b ceto adipatului.
6. Porfiria eritropoeticăcongenitală:
A. Este datoratăactivităţii deficitare a cosintazei;
B. Este datoratăactivităţii deficitare a δamino levulinat sintazei;
C. Este datoratăactivităţii deficitare a δamino levulinat dehidrogenazei;
D. Este datoratăactivităţii deficitare a porfobilinogen deaminazei;
E. Este datoratăactivităţii deficitare a feritinei.
7. Modificarile care survin in porfiria eritropoetica congenitala se datoreaza:
A. Excesului de protoporfirina IX;
B. Deficitului de polipiril metan;
C. Acumularii de uroporfirinăI, coproporfirinăI şi derivaţi ai uroporfirinogenului I;
D. Acumularii de coproporfirinogen III;
E. Corecte B. si D.
Page 77 of 120
reprezinta
8. Structura:
A. Biliverdinei;
B. Bilirubinei libere;
C. Bilirubinei conjugate;
D. Stercobilinei;
E. Urobilinogenului.
9. In calea catabolizarii hemului, reactia catalizata de biliverdin reductaza necasita potential reducator adus de:
A. NADH;
B. FADH2;
C. Tetrahidrofolat;
D. NADPH;
E. Citocromul C.
10. In cursul procesului de biodegradare al nucleului hem, reactia care necesita O2 si NADPH este catalizata de:
A. Biliverdin reductaza;
B. Hemoxigenaza;
C. Cosintaza;
D. Ferochelataza;
E. Coproporfirinogen oxidaza.
Complement Multiplu
11. Porfiriile:
Page 78 of 120
A. Sunt maladii genetice;
B. Sunt cauzate de anomalii ale caii de degradare a nucleului porfirinic;
C. Au o mare incidenta in societatea moderna;
D. Pot induce complicatii dermatologice, hepatice, psihiatrice;
E. Pot induce icter post hepatic.
12. Biosinteza hemului se realizeaza cu precadere in:
A. Maduva hematoformatoare;
B. Tesutul muscular;
C. Splina;
D. Ileon;
E. Ficat.
13. δamino levulinat sintaza:
A. Este o enzima citoplasmatica;
B. Are ca si coenzima piridoxal fosfatul;
C. Catalizeaza reactia de sinteza a δamino levulinatului in procesul de catabolism al hemului;
D. Este o enzima allosterica;
E. Are ca inhibitor nuceul hem.
14. δamino levulinat sintaza:
A. Poate fi modulata prin blocarea transportului sau in mitocondrie atunci cand concentratia de hem creste peste necesar;
B. In hepatocit poate fi modulata prin intermediul barbituricelor, prin inducerea biosintezei ei;
C. Este activata de glucoza;
D. Este modulata pozitiv de monoglucide;
E. Catalizeaza reactia de condensare a succinil CoA cu glicina.
15. Reglarea procesului de biosinteza a hemului se poate realiza prin:
A. Reglarea allosterica a ferochelatazei;
Page 79 of 120
B. Desfasurarea primei si ultimei etape a caii metabolice in acelasi compartiment, dandu-se posibilitatea realizarii unui mecanism feed-back de inhibitie prin produs final;
C. Modularea biosintezei enzimei-cheie a procesului prin intermediul produsului final sau a unor compusi exogeni sau endogeni;
D. Modificarea concentratiei de piridoxal fosfat in citoplasma hepatocitelor;
E. Modularea procesului de condensare a porfobilinogenului prin mecanism cap-coada.
16. Structura
A. Reprezinta porfobilinogenul;
B. Reprezinta α-amino β-cetoadipatul;
C. Reprezinta δ-amino levulinatul;
D. Reprezinta produsul de condensare al succinil CoA cu glicina;
E. Reorezinta produsul reactiei catalizate de δ-amino levulinat dehidrogenaza.
17. Reactia catalizata de δamino levulinat dehidrogenaza:
A. Se desfasoara in citosol;
B. Se desfasoara in mitocondrie;
C. Are ca substrat δ-amino levulinatul;
D. Are ca produs de reactie uroporfirinogenul III;
E. Este prima reactie a caii metabolice de biosinteza a hemului.
18. Porfobilinogen deaminaza:
A. Catalizeaza conversia δamino levulinatului in porfobilinogen;
B. Catalizeaza condensarea cap-coada a patru molecule de porfobilinogen;
C. Catalizeaza ciclizarea polipirilmetanului impreuna cu cosintaza;
D. Catalizeaza a doua reactie din calea de biosinteza a hemului;
E. Catalizeaza reactia-cheie a procesului de sinteza a hemului.
Page 80 of 120
19. Ionul Fe necesar biosintezei hemului:
A. Este transportat in plasma de o proteina numita transferina;
B. Este depozitat in moleculele de feritina;
C. Este transportat in plasma de o proteina numita feritina;
D. Este depozitat in moleculele de transferina;
E. Este chelatat cu protoporfirina IX sub actiunea ferochelatazei.
20. Precursorii biosintezei hemului sunt:
A. Alanina;
B. Glicocolul;
C. Acetil CoA;
D. Succinil CoA;
E. Malonil Co A.
21. Nucleul hem din hemoproteine:
A. Are o durata de viata de 120 zile;
B. Este catabolizat prin conversie in bilirubina;
C. Este catabolizat printr-un mecanism unic;
D. Este catabolizat pe cai diferite, in functie de tipul de hemoproteina;
E. Este hirolizat la aminoacizii componenti.
22. Prima reactie în degradarea hemului:
A. Este catalizatăde bilirubin sintaza;
B. Este catalizata de o enzima aparţinând familiei citocromilor P450;
C. Necesita O2 si NADPH;
D. Genereaza CO;
E. Este catalizata de biliverdin reductaza.
23. In reactia catalizata de biliverdin reductaza din calea de degradare a hemului:
A. Bilirubina este transportata la ficat sub forma complexata cu albumina serica;
B. Bilirubina este transportata in canaliculele biliare printr-un mecanism de transport activ;
C. Bilirubina este esterificata cu una sau doua molecule de acid glucuronic la nivelul radicalilor propionat;
D. Bilirubina este redusa de bacteriile din intestinul gros;
E. Bilirubina este oxidata in ileonul terminal.
26. Bilirubina conjugata:
A. Este mai solubila in apa decat bilirubina libera;
B. Este transportata la ficat dupa complexare cu albumina serica;
C. Este transportata din hepatocit in canaliculele biliare printr-un mecanism de transport activ;
D. Este degradata in ileonul terminal si in intestinul gros;
E. Este excretata in totalitate pe cale renala.
27. Degradarea bilirubinei conjugate:
A. Incepe prin indepartarea moleculelor de acid glucuronic;
B. Este un proces patologic;
C. Are loc sub actiunea enzimelor bacteriene din intestinul gros;
Page 82 of 120
D. Se realizeaza prin oxidarea completa a moleculei;
E. Este conditionata de prozenta oxigenului molecular.
28. Urobilinogenul:
A. Rezulta prin oxidarea bilirubinei conjugate;
B. Rezulta prin reducerea bilirubinei libere;
C. Este reabsorbit in cea mai mare parte in intestin;
D. Este excretat in cea mai mare parte prin fecale;
E. Se elimina in cea mai mare parte pe cale renala.
29. Bilirubina:
A. Este un agent oxidant eficace;
B. Este un agent antioxidant eficace;
C. Sub forma complexata cu albumina serica este mai eficienta decat acidul ascorbic in procesul de anihilare al peroxizilor solubili;
D. Este unul din principalii agenti oxidanti din plasma sanguina;
E. Este un agent de protectie al biomembramelor alaturi de vitamina E.
30. In procesul degradarii hemului:
A. Ionul de fier este eliminat prin fecale;
B. Nucleul protoporfirinic este procesat intr-o molecula solubila aciclica si excretat;
C. Ionul de fier este reciclat;
D. Nucleul protoporfirinic este convertit int-un compus aciclic, care este complet reciclat;
E. Se obtine un compus cu actiune antioxidanta.
Capitolul 12. Metabolismul nucleotidelor purinice si pirimidinice. Biosinteza si
catabolizarea nucleotidelor purinice. Biosinteza si catabolizarea nucleotidelor
pirimidinice. Aspecte clinice
Complement Simplu
1. Precursorii de sinteza ai nucleului purinic sunt:
Page 83 of 120
A. Acidul glutamic, asparagina, glicina, dioxid de carbon;
B. Glutamina, acidul aspartic, dioxidul de carbon;
C. Acidul glutamic, acidul aspartic, glicina;
D. Glutamina, asparagina, glicina;
E. Glicina, tetrahidrofolatul, dioxidul de carbon, acidul aspartic.
2. Forma de eliminare a azotului purinic in cazul mamiferelor este:
A. Ureea;
B. Allopurinolul;
C. Acidul uric;
D. Acidul ascorbic;
E. Inozin monofosfatul.
3. Prima reacţie din cadrul căii metabolice a biosintezei purinelor constă în:
A. Sinteza 5 fosfo ribozil 1 pirofosfat;
B. Sinteza 5 fosfo rilbozil 1 aminei;
C. Condensarea glutaminei cu ribozo 1 pirofosfatul;
D. Sinteza 5’ fosfo ribozil aminoimidazolului;
E. Nici un raspuns corect.
4. Sinteza de novo a inozin monofosfatului:
A. Are un bilant usor exergonic;
B. Este puternic endergonica;
C. Conduce la un compus cu rol biologic important;
D. Este un proces reversibil;
E. Constituie calea generala de biosinteza a nucleotidelor pirimidinice.
5. Nucleotidele eliberate prin degradarea ARN împreunăcu cele obţinute prin sintezăde novo şi cu cele din aport
exogen:
A. Sunt in totalitate hidrolizate la nucleozide, degradate oxidativ si eliminate pe cale digestiva;
Page 84 of 120
B. Alcătuiesc un fond metabolic comun, accesibil tuturor celulelor, pentru sinteza de acizi nucleici;
C. Sunt utilizate pentru sinteza de acid uric;
D. Sunt excretate ca atare;
E. Nici un raspuns corect.
6. Transformarea IMP în AMP:
A. Debuteaza prin introducerea funcţiei oxigenate la C2 al ciclului purinic;
B. Implică înlocuirea grupării oxigenate din nucleul purinic cu o grupare amino provenită dintr
C. o moleculă de acid aspartic;
D. Necesita energie furnizata de moleculele de ITP;
E. Necesita energie furnizata de moleculele de UTP;
7. Transformarea IMP în GMP:
A. Debuteaza prin introducerea funcţiei oxigenate la C2 al ciclului purinic;
B. Implicăînlocuirea grupării oxigenate din nucleul purinic cu o grupare amino provenitădintr
C. o moleculăde acid aspartic;
D. Necesita energie furnizata de moleculele de ITP;
E. Necesita energie furnizata de moleculele de UTP;
8. Guanina şi hipoxantina rezultate din hidroliza nucleotidelor:
A. Sunt in totalitate reciclate;
B. Sunt partial catabolizate la acid uric;
C. Sunt excretate ca atare prin urina;
D. Se depun sub forma de cristale insolubile la articulatii;
E. Nici un raspuns corect.
9. Adenina rezultata din hidroliza nucleotidelor:
A. Este partial catabolizata la acid uric;
B. Este in totalitate reciclata;
C. Este convertita intr-o forma solubila prin conjugare cu acid glucuronic;
Page 85 of 120
D. Se depune sub forma de cristale putin solubile la nivelul articulatiilor;
E. Nici un raspuns corect.
10. Acidul uric este:
A. Un compus cu o buna hidrosolubilitate;
B. Produsul de degradare al nucleului pirimidinic;
C. Un compus cu actiune oxidanta;
D. Forma de eliminare a azotului aminic la mamifere;
E. Nici un raspuns corect.
Complement Multiplu
11. Sinteza de novo a inozin monofosfatului :
A. Consuma sase legaturi macroergice;
B. Este puternic exergonica;
C. Este ireversibila;
D. Debuteaza prin sinteza 5 fosfo ribozil 1 pirofosfat din ATP si riboza 5 fosfat;
E. Debuteaza prin sinteza 5 fosfo ribozil N-formil glicin amidei din ATP si riboza 5 fosfat.
12. Dintre precursorii de biosinteza ai ciclului purinic fac parte:
A. Apa;
B. Dioxidul de carbon;
C. Glicina;
D. Asparagina;
E. Alanina.
13. Hiperuricemiile sunt corectate prin administrarea de:
A. Analogi structurali ai hipoxantinei;
B. Analogi structurali ai bazelor azotate pirimidinice;
C. Analogi structurali ai nucleotidelor purinice;
D. Inhibitori ai xantinoxidazei;
Page 86 of 120
E. Acid uric.
14. Despre calea metabolic de biosinteza a nucleotidelor pirimidinice sunt adevarate afirmatiile:
A. Primul metabolit al caii este carbamil fosfatul;
B. In biosinteza nucleului pirimidinic sursa de azot este glutamina;
C. Prima reactie specifica biosintezei este catalizata de arginaza si conduce la formarea acidului oxalilacetic;
D. In cursul sintezei rezulta orotidin monofosfat, compus fara rol biologic;
E. Gruparea metil a tiaminei este furnizata de N5, N10-metilen tetra hidrofolat.
15. Despre acidul uric sunt adevarate urmatoarele:
A. Constituie un factor protector faţăde radicalii hiperreactivi ai oxigenului;
B. Este convertit la alantoina in organismul uman;
C. Constituie forma de eliminare a azotului purinic;
D. Constituie forma de eliminare a azotului pirimidinic;
E. Este o substanţă uşor oxidabilă.
16. Structura :
A. Reprezinta alantoina;
B. Reprezinta produsul de catabolism al guaninei;
C. Este un compus cu actiune antioxidanta;
D. Reprezinta allopurinolul;
E. Reprezinta produsul de catabolism al bazelor pirimidinice.
17. Guta poate fi cauzată de:
Page 87 of 120
A. Tratamentul cu allopurinol;
B. Un deficit metabolic ereditar prin care se sintetizeazăacid uric în exces;
C. Incetinirea eliminării renala a monouratului de sodiu;
D. Un deficit de eliminare al bilirubinei;
E. Un deficit de metabolizare a nucleotidelor pirimidinice.
18. Tratamentul gutei cu analogi structurali de hipoxantina are ca efecte:
A. Împiedicarea transformarii hipoxantinei în xantinăşi a acesteia în acid uric;
B. Accelerarea catabolizarii guaninei si hipoxantinei;
C. Scaderea concentratiei de 5 fosforibozil 1 pirofosfat si reducerea fluxului de metaboliti spre sinteza de novo a nucleotidelor pirimidinice;
D. Inhibitia xantinoxidazei;
E. Accelerarea biosintezei nucleotidelor cu guanina si hipoxantina.
19. In calea de biosinteza a nucleotidelor pirimidinice:
A. Prima reactie se desfasoara in mitocondrie;
B. Are loc sinteza carbamoil fosfatului;
C. Prima reactie este catalizata de o enzima din ciclul ureogenetic;
D. Sursa de azot pentru sinteza carbamoil fosfatului este glutamina;
E. Sursa de azot pentru sinteza carbamoil fosfatului este ionul amoniu.
20. In calea de biosinteza a nucleotidelor pirimidinice:
A. Exista un fond citoplasmatic de carbamoil fosfat, distinct de cel utilizat de ciclul ureogenetic;
B. Exista un fond mitocondrial de carbamoil fosfat, distinct de cel utilizat de ciclul ureogenetic;
C. Prima reactie are loc cu aportul energetic adus de hidroliza unei molecule de ATP la ADP;
D. Prima reactie are loc cu aportul energetic adus de hidroliza unei molecule de ATP la AMP;
E. Prima reactie este catalizata de o enzima citoplasmatica.
21. In calea de biosinteza a nucleotidelor pirimidinice:
A. Prima reactie specifica biosintezei este sinteza carbamoil fosfatului;
B. Prima reactie specifica biosintezei este catalizata de aspartat transcarbamilaza;
Page 88 of 120
C. Prima reactie specifica biosintezei conduce la sinteza acidului carbamil aspartic;
D. Reactia de condensare dintre carbamoil fosfat si acid aspartic conduce la un compus ciclic;
E. Reactia de condensare dintre carbamoil fosfat si acid aspartic se produce cu hidroliza unei molecule de ATP.
22. In calea de biosinteza a nucleotidelor pirimidinice:
A. Carbamil aspartatul este ciclizat la dihidro orotat;
B. Ciclizarea carbamil aspartatului are loc cu eliminarea unei molecule de apa;
C. Ciclizarea carbamil aspartatului are loc cu aditia unei molecule de apa;
D. La sinteza orotatului, potentialul reducator este transferat unei molecule FAD;
E. La sinteza orotatului, potentialul reducator este transferat unei molecule NADP.
23. Produsul propriu-zis al caii de biosinteza a nucleotidelor pirimidinice:
A. Nu are rol biologic;
B. Are un important rol biologic;
C. Este inozin monofosfatul;
D. Este orotidin monofosfatul;
E. Este uridin monofosfatul.
24. Uridin monofosfatul:
A. Este sintetizat prin decarboxilarea orotidin monofosfatului;
B. Este metilat cu N5, N10-metilen tetrahidrofolat pentru sinteza TMP;
C. Este nucleotidul functional pornind de la care sunt sintetizate celelalte nucleotide pirimidinice;
D. Este nucleotidul functional pornind de la care sunt sintetizate celelalte nucleotide purinice;
E. Este sintetizat prin decarboxilarea dihidro orotidin monofosfatului.
25. Ţesuturile în diviziune rapida:
A. Sunt foarte sensibile la acţiunea dihidrofolat reductazei, enzimăce reduce dihidrofolatul la tetrahidrofolat, deoarece la nivelul lor are loc o sinteza accelerata de TMP, sinteza care necesita N5, N10-metilen tetrahidrofolat;
B. Pot fi stopate din dezvoltare de compusi cu actiune de inhibitori ai dihidrofolat reductazei;
Page 89 of 120
C. Pot fi stopate din dezvoltare prin marirea concentratiei de NADPH;
D. Pot fi stopate din dezvoltare de analogi structurali ai acidului folic cu potential de inhibitori ai timidilat sintazei;
E. Pot fi stopate din dezvoltare de analogi structurali ai acidului folic cu potential de inhibitori ai dihidrofolat reductazei.
26. Principalele puncte de control in sinteza de novo a nucleotidelor pirimidinice sunt:
A. Inhibiţia enzimei care catalizeaza prima reactie specifica a caii de către produşii finali printr-un mecanism feed-back;
B. Inhibitia carbamil fosfat sintetazei mitocondriale de catre UMP;
C. Inhibitia sintezei 5 fosforibozil 1 pirofosfat de catre carbamil fosfat;
D. Inhibitia sintezei 5 fosforibozil 1 pirofosfat de catre nucleotide;
E. Inhibitia carbamil fosfat sintetazei mitocondriale de catre AMP.
27. Principalele puncte de control in sinteza de novo a nucleotidelor pirimidinice sunt:
A. Inhibiţia aspartat transcarbamilazei de către produşii finali;
B. Inhibitia carbamil fosfat sintetazei citoplasmatice de către UMP;
C. Inhibitia carbamil fosfat sintetazei citoplasmatice de către ATP;
D. Activarea sintezei carbamil fosfatului de catre 5 fosforibozil 1 pirofosfat;
E. Inhibitia sintezei carbamil fosfatului de catre 5 fosforibozil 1 pirofosfat.
28. Catabolizarea nucleotidelor pirimidinice:
A. Are loc prin reacţii similare celor descrise la purine;
B. Are loc cu degradarea bazelor pirimidinice libere la compusi cu molecula mica;
C. Nu se produce cu degradarea lor, ele fiind excretate ca atare;
D. Genereaza α-alanina si acid b aminoizobutiric;
E. Nu are loc, acestea fiind reciclate in organism.
29. Sunt precursori de biosinteza ai nucleului pirimidinic:
A. Acidul glutamic;
B. Glutamina;
C. Acidul aspartic;
Page 90 of 120
D. Asparagina;
E. Dioxidul de carbon.
30. 5 fosforibozil 1 pirofosfat:
A. Este un compus cu incarcatura energetica mare;
B. Este molecula pe care se asambleaza nucleul purinic;
C. Este molecula pe care se grefeaza acidul orotic in procesul sintezei pirimidinelor;
D. Este un compus care rezulta din dioxid de carbon si glutamina;
E. Este un compus care este sintetizat din ribozo 5 fosfat.
Capitolul 13. Notiuni de biochimie aplicata in clinica. Profilul metabolic al
organelor principale. Constituientii biochimici principali ai sangelui si urinei.
A. Producerea glucozei in cantitati excesive de catre ficat si sub-utilizarea ei de catre celelalte organe;
B. Distrugerea prin mecanisme autoimune a celulelor α-pancreatice;
C. Secretia excesiva de insulina, neadecvata necesitatilor organismului;
D. Inhibarea cetogenezei si sub utilizarea acizilor grasi din tesutul adipos;
E. Exacerbarea suntului pentozofosfatilor si a caii acidului glucuronic.
2. Principalul combustibil utilizat de catre tesutul nervos este:
A. Aminoacizii neesentiali;
B. Aminoacizii esentiali;
C. Trigliceridele;
D. Glucoza;
E. Colesterolul.
3. Caile metabolice principale care se desfasoara in celulele tesutului hepatic sunt:
Page 91 of 120
A. Gluconeogeneza,
B. β oxidarea acizilor grasi,
C. cetogeneza,
D. ureogeneza
E. corecte A., B., C., D.
4. Dintre particularitatile metabolice ale tesutului nervos nu fac parte:
A. Functionarea exclusiva pe baza metabolismului aerob;
B. Posibilitatea de a utiliza acizii grasi ca molecule-combustibil;
C. Posibilitatea de a utiliza glucoza ca molecula-combustibil;
D. Imposibilitatea de a-si stoca moleculele-combustibil;
E. Permeabilitatea foarte mare a celulelor pentru glucoza.
5. Dintre particularitatile metabolice ale tesutului muscular striat face parte:
A. Functionarea exclusiva pe baza metabolismului aerob;
B. Functionarea exclusiva pe baza metabolismului anaerob;
C. Stocarea glucozei sub forma de glicogen;
D. Posibilitatea de a elimina din celula glucoza in exces;
E. Posibilitatea de a utiliza exclusiv acizii grasi ca molecule-combustibil.
6. Unul din rolurile tesutului adipos este:
A. De a stoca glucoza sub forma de tracilgliceroli;
B. De a converti acizii grasi in triacilgliceroli;
C. De a elibera in sange enzime;
D. De a sintetiza hormoni liposolubili;
E. De a sintetiza ATP.
7. In tesutul adipos, eliberarea primului acid gras din triacilgliceroli prin hidroliza este:
A. O reactie spontana;
B. O reactie care necesita NADPH;
Page 92 of 120
C. Etapa limitatoare de viteza a procesului de hidroliza a triacilglicerolilor;
D. Etapa catalizata de insulina;
E. O reactie din care rezulta ATP.
8. Pentru propriile sale nevoi energetice tesutul hepatic utilizeaza preferential:
A. Glucoza;
B. Acizii grasi cu catena nesaturata;
C. Acizii grasi cu catena saturata;
D. Cetoacizii rezultaţi prin degradarea aminoacizilor;
E. Corpii cetonici.
9. Principala sursa de acizi grasi utilizatăde ţesutul adipos pentru sinteza triacilglicerolilor o constituie:
A. Chilomicronii;
B. Acizii grasi conjugati cu albumina serica;
C. VLDL;
D. IDL;
E. HDL.
10. Functia principala a ficatului pentru organism este aceea de a:
A. Stoca si descompune triacilglicerolii;
B. Excreta compusii toxici si biosintetiza glucoza;
C. Coordona sistemul endocrin;
D. Furniza glucoza eritrocitelor;
E. Efectua servicii metabolice pentru alte organe şi ţesuturi.
Complement Multiplu
11. Dintre caile metabolice care se desfasoara in creier fac parte:
A. Glicoliza;
B. Cetogeneza;
C. Lipoliza;
Page 93 of 120
D. Metabolizarea aminoacizilor;
E. Biosinteza triacilglicerolilor.
12. Dintre caile metabolice principale care se desfasoara in muschi nu fac parte:
A. Glicoliza aeroba;
B. Glicoliza anaeroba;
C. Biosinteza colesterolului;
D. Biosinteza nucleotidelor purinice;
E. Ciclul acizilor tricarboxilici.
13. Dintre caile metabolice principale care se desfasoara in rinichi nu fac parte:
A. Gluconeogeneza;
B. Biosinteza hemului;
C. Biosinteza acizilor grasi;
D. Lipoliza;
E. Cetogeneza.
14. Hemoglobina glicozilata:
A. Reprezinta un indicator pretios al nivelului sanguin al glucozei;
B. Este foarte utila pentru stabilirea fluctuatiei glicemiei unui pacient pe o perioada de cateva saptamani;
C. Are proprietati electroforetice modificate fata de hemoglobina normal;
D. Reprezinta un indicator al ratei de desfasurare a cetogenezei;
E. Inregistreaza cresteri pe termen scurt in toate afectiunile hepatice.
15. In eritrocitele mature nu se desfasoara urmatoarele cai metabolice:
A. β oxidarea acizilor grasi;
B. Ciclul acizilor tricarboxilici;
C. Catena respiratorie;
D. Glicoliza;
E. Suntul pentozofosfatilor.
Page 94 of 120
16. Dintre parametrii care diagnosticheaza diabetul fac parte:
A. Prezenta glucozei in sange;
B. Prezenta glucozei in urina;
C. Cresterea pH-ului sanguin;
D. Cresterea procentului de hemoglobina glicozilata in sange;
E. Modificarea valorii activitatilor transaminazelor in sange.
17. In cazul bolnavilor de diabet netratati:
A. Nivelul insulinei în sânge este mult scăzut fata de necesitatile organismulu;
B. Concentratia serica a colesterolului este mult crescuta;
C. Concentratia plasmatica de HDL-colesterol este mult crescuta;
D. Nivelul glucagonului in sange este mult mai ridicat decat necesitatile organismului;
E. Activitatea hepatica a transaminazelor este mult crescuta.
18. Glicemia creste in:
A. Diabet zaharat;
B. Pancreatite;
C. Hepatite virale acute;
D. Retentie renala de sodiu;
E. Distrofie musculara progresiva.
19. La diabeticul netratat:
A. Intrarea glucozei în celule este împiedicată datorita deficitului de insulina;
B. Intrarea glucozei în celule este împiedicată datorita deficitului de glucagon;
C. Glicoliza este inhibata de excesul de glucagon;
D. Glicoliza este inhibata de deficitul de insulina;
E. Gluconeogeneza este stimulata de excesul de glucagon.
20. In cazul pacientilor diabetici, imposibilitatea de a utiliza glucidele ca sursa de energie conduce la:
A. Exacerbarea β-oxidarii acizilor grasi;
Page 95 of 120
B. Exacerbarea gluconeogenezei;
C. Exacerbarea glicogenolizei;
D. Exacerbarea cetogenezei;
E. Oprirea completa a catenei respiratorii.
21. La pacientul diabetic cetonemia este ridicata deoarece:
A. Datorităcantităţii scăzute de oxaloacetat disponibile, acetil-CoA produsa in urma degradarii oxidative a acizilor grasi nu poate intra în ciclul acizilor tricarboxilici;
B. Excesul de acetil-CoA este orientat spre sinteza din corpi cetonici;
C. Catena respiratorie este decuplata de fosforilare;
D. Glicoliza este exacerbata in tesutul hepatic;
E. Creierul nu reuseste sa consume toti corpii cetonici pusi la dispozitie.
22. Excesul de corpi cetonici produsi de ficatul diabeticului:
A. Induce o exacerbare a metabolismului oxidativ la nivelul tesutului nervos;
B. Sparge sistemele-tampon din sange si contracareaza efortul de reechilibrare al rinichilor;
C. Conduce la dezechilibrarea balanţei acido-bazice;
D. Induce o accelerare a desfasurarii reactiilor ciclului acizilor tricarboxilici;
E. Conduce la utilizarea alternativa a aminoacizilor ca surse energetice.
23. Referitor la parametrii biochimici sanguini sunt adevarate afirmatiile:
A. Se recomanda a fi determinati imediat dupa recomandarea medicului sau farmacistului;
B. Se recomanda a fi determinati dupa un repaus alimentar de 8-12 ore;
C. Se recomanda a fi determinati dupa un repaus alimentar de 10-14 ore;
D. Se recomanda determinarea dupa un repaus alimentar de 10-14 ore numai in cazul triacilglicerolilor;
E. Pot fi alterati de o colectare improprie a probelor.
24. In urma separarii electroforetice a proteinelor plasmatice, in banda albuminelor se gasesc:
A. Transtiretina;
B. Albumina;
Page 96 of 120
C. Antitripsina;
D. Antichimotripsina;
E. Trombina.
25. In urma separarii electroforetice a proteinelor plasmatice, in banda alfa 1 globulinelor se gasesc:
A. Transcortina;
B. Transtiretina;
C. Lipoproteine HDL;
D. Protrombina;
E. Ceruloplasmina.
26. In urma separarii electroforetice a proteinelor plasmatice, in banda alfa 2 globulinelor se gasesc:
A. Plasmina;
B. Plasminogenul;
C. Antitrombina III;
D. Ceruloplasmina;
E. Proteina C-reactiva.
27. In urma separarii electroforetice a proteinelor plasmatice, in banda beta globulinelor se gasesc:
A. Fibrinogenul;
B. Imunoglobuline A;
C. Ceruloplasmina;
D. Transferina;
E. Proteina C-reactiva.
28. In urma separarii electroforetice a proteinelor plasmatice, in banda gamma globulinelor se gasesc:
A. Imunoglobuline G;
B. Fibrina;
C. Plasmina;
D. Imunoglobuline M;
Page 97 of 120
E. Imunoglobuline C.
29. Pentru acidul uric se observa valori sanguine crescute in urmatoarele afectiuni:
A. Hipoxie;
B. Obstructii biliare;
C. Guta;
D. Leucemii;
E. Infarct miocardic.
30. Valori crescute ale colesterolului se intalnesc in:
A. Insuficienta renala;
B. Hepatite;
C. Pancreatite;
D. Distrofii musculare;
E. Leucemii.
Capitolul 14. Notiuni de pato-biochimie. Perturbari metabolice in etilism.
Perturbari metabolice si parametri de diagnostic in afectiunile hepato-biliare.
Perturbari metabolice si parametri de diagnostic in afectiunile cardio-vasculare
Complement Simplu
1. In urma infarctului miocardic acut are loc cresterea activitatii serice a:
A. Gamma glutamil transpeptidazei;
B. Creatinkinazei;
C. 5’ nucleotidazei;
D. Somatomedinei;
E. Alcool dehidrogenazei.
2. Consumul excesiv de alcool pe o perioada indelungata conduce la:
A. Cresterea vitezei de oxidare a acizilor grasi la nivelul tesutului hepatic;
Page 98 of 120
B. Cresterea vitezei de desfasurare a ciclului acizilor tricarboxilici in citosolul hepatocitelor;
C. Acumularea acizilor graşi în ficat, proveniţi in principal din sintezăendogenă;
D. Perturbări ale biosintezei de proteine la nivelul ficatului;
E. Nici una din modificarile metabolice mentionate mai sus.
3. Etilismul:
A. Reprezinta consumul ocazional de cantitati excesive de alcool;
B. Reprezinta consumul cronic de cantitati mici de alcool;
C. Conduce la acumularea de lipide în ficat, hiperlipidemie şi în final la ciroză;
D. Conduce la o producţie excesivăde NADH la nivelul tesutului adipos;
E. Induce carenta de calciu.
4. Metabolizarea etanolului de catre alcool dehidrogenaza:
A. Are loc exclusiv in tesutul renal;
B. Are loc exclusiv in enterocite si eritrocite;
C. Are loc cu participarea sistemului microzomal de oxidare a etanolului;
D. Conduce la creşterea raportului [lactat]/[piruvat], datorita producţiei excesive de NADH;
E. Conduce la scaderea raportului [lactat]/[piruvat], datorita producţiei excesive de NADH.
5. Cauza probabila a agravarii gutei in urma consumului excesiv de alcool este:
A. Scaderea capacitatii rinichilor de a excreta acid uric datorita hiperlactacidemiei;
B. Scaderea raportului [NADH]/[NAD+] in hepatocite;
C. Metabolizarea etanolului cu participarea sistemului microzomal de oxidare a etanolului;
D. Inhibarii metabolizarii barbituricelor;
E. Nici o varianta corecta.
6. Citocromul P450:
A. Face parte din transportorii de electroni ai catenei respiratorii;
B. Conduce la conversia etanolului in acetaldehida cu generarea de NADH in exces;
C. Este coenzima alcool dehidrogenazei hepatice;
Page 99 of 120
D. Inhiba metabolizarea medicamentelor;
E. Face parte din sistemului microzomal de oxidare a etanolului (MEOS).
7. Flora bacteriană din ileonul terminal si din colon converteste acizii biliari din secretia biliara in:
A. Bilirubina;
B. Colesterol liber si esterificat;
C. Esteri primari ai bilirubinei;
D. Acizi biliari secundari;
E. Acizi biliari primari.
8. Acizii litocolic si deoxicolic din ileonul terminal si din colon:
A. Sunt absorbiţi şi intrăîn circulaţia portală, reîntorcându-se la ficat;
B. Sunt excretati pe cale renala;
C. Sunt utilizati ca molecule-combustibil de catre bacteriile florei intestinale;
D. Sunt acizi biliari primari;
E. Sunt degradati la bilirubina si excretati prin fecale.
9. Icterul hepatic:
A. Se produce în urma deficienţei de excreţie a bilirubinei cauzate de obstrucţia mecanicăa fluxului bilei în intestin;
B. Poate fi produs printr-un deficit al mecanismului de conjugare sau printr-o secreţie anormalăa bilirubinei de către hepatocit;
C. Se poate instala in urma anemiei hemolitice;
D. Reprezinta colorarea in rosu-brun a mucoaselor si tegumentelor;
E. Se datoreaza excretiei defectuase a acizilor biliari.
10. Când secreţia biliarănu este deversată în intestin:
A. Fecalele se decolorează;
B. Fecalele se coloreaza intens;
C. Urina se decoloreaza;
D. Plasma sanguina se coloreaza in rosu-brun;
Page 100 of 120
E. Nici o varianta corecta.
Complement Multiplu
11. Magnezemia:
A. Reprezinta concentratia serica de magneziu;
B. Reprezinta un indice al riscului producerii infarctului miocardic;
C. Inregistreaza cresteri usoare in infarctul miocardic acut;
D. Reprezinta un parametru exclusiv pentru diagnosticarea afectiunilor renale;
E. Creste in mod fiziologic la femeile insarcinate.
12. Etilismul:
A. Conduce la acumularea de lipide la nivelul tesutului hepatic;
B. Conduce la acumularea in ficat a acizilor grasi proveniti din sinteza endogena;
C. Conduce la acumularea in ficat a acizilor grasi proveniti exclusiv din hidroliza triacilglicerolilor depozitati in tesutul adipos;
D. Este cauza majora de crestere a concentratiei de glucoza in urina;
E. Conduce in final la ciroza hepatica.
13. Gamma glutamil transpeptidaza:
A. Se gaseste in concentratii mari in splina, hematii si tesut adipos;
B. Se gaseste in concentratii mari in rinichi si ficat;
C. Este un parametru care confera indicatii in neoplasmul hepatic in absenta icterului;
D. Este in mod frecvent primul parametru clinic care se modifica in cazul alcoolicilor;
E. Este strict specifica pentru diagnosticul cancerului de colon.
14. Lactat dehidrogenaza:
A. Reprezinta un parametru exclusiv pentru diagnosticarea afectiunilor renale;
B. Inregistreaza cresteri usoare in infarctul miocardic acut;
C. Isi mentine valorile crescute in ser timp de aproximativ 10 zile dupa infarctul miocardic acut;
D. Confera informatii pretioase pentru diagnostic in cazul analizei izoenzimelor ei;
E. Se gaseste exclusiv in tesutul muscular.
Page 101 of 120
15. Icterul prehepatic:
A. Este produs printr-o perturbare in procesul de eliminare a biliverdinei din hepatocite;
B. Este rezultatul prezentei unei cantitati de bilirubina in exces in hepatocite;
C. Se produce in urma deficientei de excretie a bilirubinei cauzata de obstructia mecanica a fluxului bilei in intestine;
D. Se caracterizeaza prin cresterea concentratiei de bilirubina libera;
E. Este u n simptom caracteristic etilismului.
16. Activitatea serica a fosfatazei alcaline:
A. Se determina in clinica pentru a diagnostica afectiuni hepatice si osoase;
B. Inregistreaza cresteri pe termen scurt in toate afectiunile hepatice;
C. Creste in metastazele hepatice, putand fi singurul parametru modificat pe buletinul de analiza;
D. Creste in infarctul miocardic acut;
E. Creste in mod fiziologic la femeile insarcinate.
17. Despre secreţia biliarăsunt adevarate urmatoarele informatii:
A. Este compusădin acizi biliari, liberi sau sub formăde săruri, pigmenţi biliari, colesterol şi alte substanţe provenite din sânge;
B. Producţia medie de secreţie biliarăa unui organism în decursul unei zile este de 3 L;
C. Participăla digestia şi absorbţia lipidelor la nivelul hepatocitelor;
D. Contine exclusiv acizi biliari secundari (deoxicolic şi litocolic);
E. Contine hemoglobina si alte proteine cu hem.
18. Determinarea activitatii serice a fosfatazei alcaline se foloseste in clinica pentru:
A. Diagnosticarea afectiunilor ososase, atunci cand ceilalti parametrii de diagnostic nu indica afectiuni hepatice;
B. Diagnosticarea timpurie a sarcinii;
C. Identificarea afectiunilor de natura hepatica;
D. Diagnosticarea complicatillor diabetului si a afectiunilor coronariene.
E. Diagnosticarea hepatitei virale C
19. Alanin aminotransferaza (ALT) sau glutamat piruvat transaminaza (GTP):
Page 102 of 120
A. Este o enzimăcare se găseşte în toate ţesuturile, cu precădere în ficat şi, în cantităţi mai mici, în rinichi şi muşchii scheletici;
B. Inregistreaza activitati crescute in hepatitele virale severe;
C. Este o enzimăa cărei activitate sericăcreşte în 6-8 ore de la producerea infarctului miocardic, atingând valoarea maximăla 24 ore;
D. Creste foarte mult in obstrucţiile biliare;
E. Creste spectaculos in afectiunile hepatice cronice.
20. Determinarea activitatii serice a 5’-nucleotidazei serveste in clinica pentru:
A. A se evidenţia dacă activitatea crescută a fosfatazei alcaline se datorează afecţiunilor hepatice sau celor osoase;
B. Certificarea unei afectiuni coronariene, prin determinarea simultana a alanin aminotransferazei serice;
C. Diagnosticarea metastazelor hepatice, constituind un parametru foarte sensibil de diagnostic;
D. Evidentierea hepatitelor virale;
E. Decelarea etilismului la conducatorii auto.
21. Despre creatin kinaza sunt adevarate urmatoarele:
A. Este o enzima care catalizeaza reactii de transaminare in tesutul hepatic;
B. Activiatea sa cea mai crescutăse inregistreaza în muşchii scheletici, miocard şi în creier;
C. Este primul parametru clinic ce se modifică în cazul alcoolicilor;
D. Este implicată în stocarea energiei în creatin-fosfat la nivelul tesutului muscular;
E. Inregistreaza valori crescute ale activitatii în disfuncţiile musculare, afecţiunile sistemului nervos central (accidente vasculare cerebrale), degenerări nervoase, şocuri ale sistemului nervos central.
22. O pacientăîn vârstăde 75 ani, cu diabet, prezintăo valoare a hemoglobinei glicozilate (HbA1C) de 8% (faţăde
mai puţin de 5% -valoarea normală). A relatat ca îşi ţine singurăboala sub control prin regim alimentar, fara medicatie
şi a declarat de asemenea că testarea glucozuriei folosind teste-stick colorimetrice conduce întotdeauna la rezultate negative.
Determinarea glicemiei a jeun in laboratorul clinicii timp de 5 zile a condus la inregistrarea unor valori usor crescute fata de limita de diagnosticare a diabetului (200 mg/dL). Nu s-a observat glucozurie in acest interval de timp. Din analiza cazului reiese ca:
A. Valorile hemoglobinei glicozilate diagnosticheaza in mod cert diabetul;
B. Valoarea medie crescuta pentru glicemie indica o scapare a bolii de sub control;
C. Pacienta nu sufera de diabet;
Page 103 of 120
D. Pacienta poate sa-si fi facut testul in mod incorect sau sa fi avut dificultati in a-l citi, datorita problemelor de vedere; E. Pacienta sufera de insuficienta renala.
23. Separarea şi identificarea electroforeticăa izoenzimelor creatin kinazei:
A. Constituie un indicator specific pentru diagnosticul infarctului miocardic;
B. Certifica diagnosticul de infarct miocardic la evidenţierea unei creşteri a activităţii serice CKMB maimaride6%;
C. Certifica diagnosticul de infarct miocardic la evidenţierea unei creşteri a activităţii serice CKBB mai mari de 6% ;
D. Certifica diagnosticul de infarct miocardic la evidenţierea unei creşteri a activităţii serice CKMM maimaride6%;
E. Nu constituie unparametru de diagnostic pentru afectiunile cardiovasculare.
24. Constituie parametri utili in dagnosticarea infarctului miocardic acut:
A. Aspartat aminotransferaza hepatica;
B. Analiza izoenzimelor lactat dehidrogenazei;
C. Analiza izoenzimelor creatin kinazei;
D. Gamaglutamil transpeptidaza;
E. Colesterolul total.
25. Aspartat aminotransferaza (AST) sau glutamat oxaloacetat transaminaza (GOT):
A. Este o enzimă care se găseşte în toate ţesuturile (ubicuitara);
B. Este o enzimă care se găseşte exclusiv in tesutul hepatic, modificarea activitatii sale conferind informatii despre functionarea acestui tesut;
C. Este o enzimă care se găseşte exclusiv in muschiul miocardic, modificarea activitatii sale constituind un parametru de diagnostic al infarctului miocardic;
D. Inregistreaza activitati crescute in hepatitele virale severe;
E. Este o enzimă a cărei activitate serică creşte în 6-8 ore de la producerea infarctului miocardic, atingând valoarea maximă la 24 ore.
26. O pacientă în vârstă de 80 ani a fost diagnosticată cu hipertensiune arterială, insuficienţă
cardiacă, anemie, prezumptiv diabet şi insuficienţă renală cronică. A fost tratată cu diuretice şi perfuzată cu soluţii de electroliţi. A fost externată după 4 zile. Rezultatele investigării biochimice sunt prezentate în tabelul următor.
Page 104 of 120
Parametrii Prima internare A doua internare Valori normale
Iniţial La externare Iniţial La externare
Uree serică mg/dL
58 - - 61.0 25-50
Creatinina mg/dL 6,2 6,2 6,4 6,0 0,5-1,1
Acid uric mg/dL 10.0 - - 9,2 2-5
Glucoza mg/dL 86 80 113 - 80-110
După 5 luni, a fost reinternată în spital pentru tratarea unor accese repetate de dispnee. A fost trecută pe dietă alimentară şi pe medicaţie antihipertensivă adecvată. După aceste măsuri simptomele s-au remis. A reieşit că pacienta nu-şi lua medicaţia prescrisă şi a fost consiliată privind importanţa respectării tratamentului medicamentos prescris.
Analizand cazul se pot trage urmatoarele concluzii:
A. Cauza valorii crescute a ureei serice la prima internare a fost insuficienta renala;
B. Cauza valorii crescute a ureei serice la prima internare a fost anemia;
C. Cauza valorii crescute a creatininei a fost hipertensiunea arteriala;
D. Valoarea crescuta a ureei serice nu se datoreaza insuficientei cardiace, deoarece nu s-au observat imbunatatiri semnificative ale parametrilor biochimici de diagnostic la dministrarea de antihipertensive;
E. Valorile mari ale acidului uric si creatininei se datoreaza capacitatii insuficiente de liminare
27. O fetiţă de 3 ani este internată cu diagnosticul de leucemie limfocitară acută. Rezultatele
investigării biochimice sunt prezentate în tabelul următor.
Param. Prima zi
A 2a zi
A 3a zi
A 4a zi
A 11a zi
A 12a zi
A 20a zi
Valori normale
Uree serică mg/dL
12,0 - - 15,0 4,0 2,0 - 25-50
Creatinina mg/dL
0,7 - - 1,0 0,7 - 0,7 0,5-1,1
Acid uric mg/dL
12,0 9,2 4,0 1,9 2,3 - 3,1 2-5
Limfocite/ mm3
56.300 - - - 3.700 2.800 3.700 4.000-10.000
A fost instituit tratamentul chimioterapic adecvat din a doua zi de spitalizare. Din analiza cazului se pot desprinde urmatoarele concluzii:
A. Cresterea valorii acidului uric se datoreste cresterii semnificative a distructiei nucleelor limfocitelor,
aflate in numar mult mai mare fata de normal;
B. Cresterea valorii acidului uric are o cauza renala;
C. Cresterea valorii acidului uric nu are o cauza renala, deoarece valorile creatininei si ureei sunt
Page 105 of 120
normale;
D. Parametrii biochimici certifica diagnosticul de leucemie;
E. Parametrii biochimici nu certifica diagnosticul de leucemie
28. Un băiat de 19 ani a fost internat de urgenţă, aflându-se în comă, fără semne de lovire sau
criză comiţială. Examenul medical nu a dus la concluzii relevante. S-a remarcat doar un suflu scurt şi gâfâit, precum şi puls slab şi foarte rapid. Rezultatele investigării biochimice sunt prezentate în tabelul următor.
Parametrii Valori Valori normale
Teste pentru detectarea drogurilor (proba de urină)
negativ -
pH urinar 5.0 4,5-8,0
Glucoză urinară 4+ negativ
Cetonurie 3+ negativ
Proteine în urină 2+ negativ
pH sanguin 7,15 7,35-7,45
Cetonemie 3+ negativ
Uree serică mg/dL 60 25-50
Creatinină mg/dL 1,7 0,5-1,4
Glicemie mg/dL 723 65-110
Din analiza cazului se pot desprinde urmatoarele concluzii:
A. Cel mai probabil diagnostic este insuficienta renala;
B. Cel mai probabil diagnostic este cetoacidoza diabetica;
C. Pacientul sufera de diabet netratat;
D. Pacientul nu sufera de diabet, hiperglicemia inregistrata fiind recent instalata si tranzitorie;
E. Pacientul necesita investigatii hematologice suplimentare pentru diagnosticare.
29. Un pacient prezentând icter sever, durere sub rebordul costal drept, febra şi frisoane are
următorul tablou biochimic sanguin:
Parametrii Valori
Forfatază alcalină De 4 ori valoarea normală
Colesterol total Crescut
GOT Normal
5’-nucleotidaza Crescută
Bilirubina totală 25 mg/dL (valori normale 0,2-0,8 mg/dL)
B. Pacientul sufera de icter mecanic datorat unei obstructii extrahepatice;
C. Pacientul nu sufera cu certitidine de metastaza hepatica;
D. Pacientul prezinta suspiciune de metastaza hepatica;
E. Valoarea bilirubinei totale nu justifica diagnosticul de obstructie extrahepatica
30. In cazul unui pacient care acuza o uşoară pierdere în greutate, senzaţie de vomă şi greaţă
s-au obţinut următoarele valori ale parametrilor biochimici:
Parametrii Valori Valori normale
Bilirubina totală 20 mg/dL 0,2-0,8 mg/dL
Bilirubia conjugată 10 mg/dL 0-0,2 mg/dL
Fosfataza alcalină Uşor crescută
GPT Semnificativ crescută
GOT Uşor crescută
Albumina Scăzută
Gamma-globuline Crescute
Ulterior, pacientul a dezvoltat icter şi hepatomegalie. Analizand cazul, putem deduce ca:
A. Procesul morbid care s-a dezvoltat cel mai probabil este distructia hepatocitelor;
B. Cauzele cele mai probabile ale acestor dereglari pot fi: hepatita virala, hepatita alcoolica, sau o intoxicatie cu toxici hepatici severi de tipul tetraclorurii de carbon sau al fosforului;
C. Pacientul nu prezinta afectiuni hepatice;
D. Cauzele cele mai probabile ale acestor dereglari pot fi: infarctul miocardic acut, calculoza biliara,
dislipidemiile;
E. Sunt necesare determinari suplimentare pentru diagnosticul diabetului.
31. O pacienta in varsta de 60 ani, cu un cancer mamar tratat chirurgical in urma cu 3 ani, se plange de o stare de slabiciune generala si de dureri ososase. Examenul biochimic al sangelui au relevat valori normale ale electrolitilor (Ca, Mg, PO3), ale proteinei totale serice si albuminei. Alti parametrii biochimici masurati in ser au avut urmatoarele valori:
valori normale
Bilirubina totala- 7 μmol/L 3- 22 μmol/L
Page 107 of 120
AST- 33 U/L 12-48 U/L
ALT- 38 U/L 3-55 U/L
Fosfataza alcalina- 890 U/L 80-280 U/L
Γ Glutamil transpeptidaza- 32 U/L < 36 U/L
Considerate ca pacienta:
A. Are o disfunctie biliara;
B. Are osteoporoza;
C. Poate avea o metastaza osoasa;
D. Poate avea o metastaza hepatica;
E. Este sanatoasa.
32. O pacienta in varsta de 60 ani, cu un cancer mamar tratat chirurgical in urma cu 3 ani, se plange de o stare de slabiciune generala si de dureri ososase. Examenul biochimic al sangelui au relevat valori normale ale electrolitilor (Ca, Mg, PO3), ale proteinei totale serice si albuminei. Alti parametrii biochimici masurati in ser au avut urmatoarele valori:
valori normale
Bilirubina totala- 7 μmol/L 3- 22 μmol/L
AST- 33 U/L 12-48 U/L
ALT- 38 U/L 3-55 U/L
Fosfataza alcalina- 890 U/L 80-280 U/L
Glutamil transpeptidaza- 32 U/L < 36 U/L
Ce recomandari ati face pacientei?
A. Sa ia vitamine;
B. Sa faca un examen RMN al oaselor;
C. Sa se odihneasaca mai mult;
D. Sa continue examinarea biochimica a sangelui cu parametrii tumorali;
E. Sa bea mai putin alcool.
33. Un pacient in varsta de 65 ani soseste la Urgenta cu semne vizibile de icter. Nu acuza dureri dar relateaza o scadere a greutatii, precum si o decolorare a fecalelor. Consuma de obicei alcool in cantitati moderate si nu ia nici un fel de medicamente. Rezultatele examenului biochimic seric sunt: valori normale
Bilirubina 250 μmol/L 3- 22 μmol/L
Page 108 of 120
AST 87 U/L 12-48 U/L
ALT 92 U/L 3-55 U/L
Fosfataza alcalina 850 U/L 80-280 U/L
In acest caz:
A. Exista supozitia unei litiaze colecistice
B. Este foarte probabil un diagnostic de carcinom de cap de pancreas
C. Valorile ALT si AST nu indica injurii serioase ale hepatocitului
D. Valorile ALT si AST indica injurii serioase ale hepatocitului
E. Se recomanda de urgenta o ecografie abdominal
34. Un pacient in varsta de 65 ani soseste la Urgenta cu semne vizibile de icter. Nu acuza dureri dar relateaza o scadere a greutatii, precum si o decolorare a fecalelor. Consuma de obicei alcool in cantitati moderate si nu ia nici un fel de medicamente. Rezultatele examenului biochimic seric sunt: valori normale
Bilirubina 250 μmol/L 3- 22 μmol/L
AST 87 U/L 12-48 U/L
ALT 92 U/L 3-55 U/L
Fosfataza alcalina 850 U/L 80-280 U/L
Aceste valori se pot datora:
A. Unei cresteri a volumului ganglionilor limfatici, care obstructioneaza astfel ductul biliar;
B. Consumului de alcool;
C. Unei hepatite virale acute;
D. Unei tumori pancreatice;
E. Varstei inaintate.
35. La o petrecere campeneasca a fost organizat un pavilion sanitar, in cadrul caruia s-au masurat si diversi parametri biochimici sanguini prin teste rapide.
Un baietel de 11 ani a avut o glicemie de 14,4 mmol/L (valoare normala maxima 5,4 mmol/L). Familia sa a fost foarte ingrijorata si a dorit repetarea testului cu un alt aparat, care a confirmat primul rezultat. Varul sau primar, de varsta apropiata, sufera de diabet.
Recomandati in acest caz:
A. Inceperea terapiei cu insulina;
Page 109 of 120
B. Testarea intregii familii;
C. Repetarea determinarii a doua zi de dimineata pe nemancate;
D. Un consum mai redus de dulciuri concentrate;
E. Repetarea determinarilor in urmatoarea ora.
36. La o petrecere campeneasca a fost organizat un pavilion sanitar, in cadrul caruia s-au masurat si diversi parametri biochimici sanguini prin teste rapide.
Un baietel de 11 ani a avut o glicemie de 14,4 mmol/L (valoare normala maxima 5,4 mmol/L). Familia sa a fost foarte ingrijorata si a dorit repetarea testului cu un alt aparat, care a confirmat primul rezultat. Varul sau primar, de varsta apropiata, sufera de diabet.
Care ar putea fi cauzele acestui rezultat?
A. Diabetul zaharat;
B. Testarea glicemiei imediat dupa consumul de alimente;
C. In general, consumul de alimente bogate in glucide;
D. Realizarea determinarii in conditii neadecvate pentru pacient;
E. Ereditatea copilului.
37. Un pacient in varsta de 52 ani ajunge la sectia de Urgenta prezentand dureri puternice in capul pieptului cu radiere pe bratul stang, care persista de mai mult de doua ore. Are un istoric de anghina pectorala si este fumator.
Ce teste biochimice recomandati pentru confirmarea diagnosticului?
A. Bilirubina serica si colesterolul;
B. Creatin kinaza serica;
C. Glicemia si colesterolul;
D. Izoenzimele creatin kinazei;
E. Lactat dehidrogenaza serica.
38. Un pacient in varsta de 52 ani ajunge la sectia de Urgenta prezentand dureri puternice in capul pieptului cu radiere pe bratul stang, care persista de mai mult de doua ore. Are un istoric de anghina pectorala si este fumator.
Diagnosticul de infarct miocardic poate fi confirmat de:
A. Dozarea magneziului seric;
Page 110 of 120
B. Dozarea creatin kinazei serice;
C. Dozarea bilirubinei serice;
D. Dozarea colesterolului seric;
E. Evidentierea electroforetica a izoenzimelor creatin kinazei.
39. Un adolescent in varsta de16 ani, epileptic, cu crize de epilepsie din ce in ce mai frecvente si mai violente este investigat biochimc.
Valoarea activitatii γ glutamil transpeptidazei este 82 U/L (valori normale < 36 U/L) iar cea a fosfatazei alcaline 520 U/L (valori normale 80-280 U/L).
Referitor la rezultatele obtinute se poate afirma ca:
A. Adolescentul are o tumora osoasa;
B. Adolescentul are ciroza hepatica;
C. Nu se confirma abuzul de alcool;
D. Adolescentul este in crestere;
E. Nu exista o afectiune hepatica.
40. Un adolescent in varsta de16 ani, epileptic, cu crize de epilepsie din ce in ce mai frecvente si mai violente este investigat biochimc.
Valoarea activitatii γ glutamil transpeptidazei este 82 U/L (valori normale < 36 U/L) iar cea a fosfatazei alcaline 520 U/L (valori normale 80-280 U/L).
Referitor la acest caz, sunt corecte afirmatiile:
A. O valoare crescuta a γ glutamil transpeptidazei nu constituie in sine un indiciu pentru alcolism;
B. Valoarea fosfatazei alcaline este fireasca pentru un adolescent in plina dezvoltare;
C. Valoarea fosfatazei alcaline poate fi considerata un indice al unei suferinte hepatice;
D. Valoarea fosfatazei alcaline poate fi considerata un indice al unei metastaze osoase;
E. Valoarea fosfatazei alcaline confirma supozitia de alcoholism.
41. Un barbat in varsta de 35 ani se prezinta la serviciul de urgenta cu dureri puternice in zona lombara. Examenul biochimic al sangelui releva o valoare a creatininei de 150 μ moli/L (valori normale 62-132 μ moli/L). In urina colectata timp de 24 ore valoarea creatininei este 7.5 m moli/L (valori normale 6-16,5 m moli/L).
Referitor la acest caz, sunt corecte afirmatiile:
A. Pacientul sufera de insuficienta renala;
Page 111 of 120
B. Pacientul nu are afectiuni renale;
C. Pentru diagnosticarea eventualelor afectiuni renale se recomanda completarea tabloului cu determinarea altor parametri;
D. In lipsa altor date, nu poate fi diagnosticata o litiaza renala;
E. Pacientul sufera de o complicatie a diabetului zaharat.
42. O pacienta de 49 ani se prezinta la medicul de familie acuzand o anorexie durand de 8 zile, greata si simptome similar celor gripale. De asemenea, relateaza ca urina are culoare bruna de doua zile. La palpare, abdomenul este moale si nedureros.
Examenul biochimic al sangelui a condus la urmatoarele rezultate:
Valori normale
Bilirubina totala 63 μ moli/L 3- 22 μmol/L
AST 939 U/L 12-48 U/L
ALT 2700 U/L 3-55 U/L
Fosfataza alcalina 410 U/L 80-280 U/L
Γ glutamil transpeptidaza 312 U/L < 36 U/L
Proteina totala 68 g/L 60-80 g/L
Albumina serica 42 g/L 35-55 g/L
Referitor la cele prezentate, sunt adevarate afirmatiile:
A. Cele mai alterate valori le prezinta fosfataza alcalina;
B. Cele mai alterate valori le prezinta γ glutamil transpeptidaza;
C. Valorile transaminazelor indica un grad foarte ridicat de distructie a hepatocitelor;
D. Este un tablou tipic de hepatita acuta;
E. Diagnosticul probabil este de litiaza biliara.
43. O pacienta de 49 ani se prezinta la medicul de familie acuzand o anorexie durand de 8 zile, greata si simptome similar celor gripale. De asemenea, relateaza ca urina are culoare bruna de doua zile. La palpare, abdomenul este moale si nedureros.
Examenul biochimic al sangelui a condus la urmatoarele rezultate:
Valori normale
Bilirubina totala 63 μ moli/L 3- 22 μmol/L
AST 939 U/L 12-48 U/L
ALT 2700 U/L 3-55 U/L
Fosfataza alcalina 410 U/L 80-280 U/L
Γ glutamil transpeptidaza 312 U/L < 36 U/L
Proteina totala 68 g/L 60-80 g/L
Page 112 of 120
Albumina serica 42 g/L 35-55 g/L
Referitor la cele prezentate, nu sunt adevarate urmatoarele:
A. Este vorba de o ciroza hepatica;
B. Este vorba de o insuficienta hepatica cronica;
C. Este vorba de o suferinta hepatica acuta;
D. Cele mai alterate sunt valorile transaminazelor;
E. Tabloul biochimic, corelat cu simptomatologia, exclud un diagnostic de litiaza biliara.
44. Dintre pacientii de mai jos, care pot fi diagnosticati cu diabet zaharat?
1. Barbat de 65 ani, obez, glicemie a jeun 16.7 mmol/L, glicozurie;
2. Femeie 62 ani, glicemie a jeun 11,7 mmol/L, glicozurie absenta;
3. Barbat 41 ani, constitutie astenica, glicemie a jeun 10.1 mmol/L,, hemoglobina glicozilata 4%;
4. Femeie 65 ani, obeza si hipertensiva, glicemie a jeun 15 mmol/L, corpi cetonici 4 mg/dL
45. O pacienta de 22 ani, diabetica, soseste la serviciul de urgent, cu dureri abdominale si spasme de voma de aproximativ 2 zile. Este apatica iar respiratia ii este adanca si rapida. Halena respiratiei are iz de mere putrede.
Care din urmatoarele variante sunt corecte?
A. Pacienta are o criza de apendicita;
B. Se recomanda determinarea de urgenta a glicemiei si cetonuriei pentru verificarea diagnosticului;
C. Pacienta manifesta o cetoacidoza diabetica;
D. Pacienta are o forma incipienta de hepatita;
E. Se recomanda masurarea pH-ului sanguine.
Page 113 of 120
46. O pacienta de 22 ani, diabetica, soseste la serviciul de urgent, cu dureri abdominale si spasme de voma de aproximativ 2 zile. Este apatica iar respiratia ii este adanca si rapida. Halena respiratiei are iz de mere putrede.
Care din urmatoarele variante nu sunt corecte?
A. Pacienta si-a neglijat tratamentul pentru diabet;
B. Pacienta are acidoza metabolica;
C. Pacienta are alcaloza metabolica;
D. Pacienta manifesta o blocare a cetogenezei;
E. Se recomanda investigarea urinei pacientei, precum si masurarea glicemiei.
47. O pacienta de 66 ani se prezinta la medicul specialist acuzand dureri puternice in piciorul drept si in pelvis. Examenul radiologic a relevant leziuni specific maladiei Paget.
Care din urmatoarele variante sunt corecte?
A. Examenul biochimic va releva o scadere semnificativa a activitatii fosfatazei alcaline serice;
B. Examenul biochimic va releva o crestere semnificativa a activitatii fosfatazei alcaline serice;
C. Fosfataza alcalina va creste datorita amplificarii activitatii osteoblastice;
D. Fosfataza alcalina va scadea datorita amplificarii activitatii osteoblastice;
E. Nu se vor observa modificari ale activitatii fosfatazei alkaline serice.
48. La investigarea unei paciente obeze de 63 ani s-au determinat urmatoarele valori ale unor parametri biochimici sanguini:
Valori normale
Colesterol total 9,3 mmol/L 3,9-5,7 mmol/L
Creatinkinaza 290 U/L 10-50 U/L
AST 35 U/L 0-41 U/L
Care din urmatoarele aprecieri sunt adevarate?
A. Se recomanda de urgenta un examen EKG;
B. Pacienta prezinta un risc sever de infarct miocardic sau chiar a suferit un infarct;
C. Pacienta este la menopauza si de aceea valorile acestor parametrii sunt alterate;
D. Se recomanda explorarea de urgent a functiei renale;
E. Pacienta nu prezinta risc de infarct.
Page 114 of 120
49. O pacienta de 69 ani, cu un cancer mamar tratat chirurgical in urma cu 8 ani, se plange de slabiciune, lipsa de energie, slabire accentuata in ultimele 3 luni si dureri osoase.
Care din urmatoarele variante sunt adevarate, referitor la acest caz?
A. Se recomanda determinarea activitatii fosfatazei alkaline serice;
B. Se recomanda determinarea activitatii γ glutamil transpeptidazei;
C. Se recomanda determinarea transaminazelor serice;
D. Este suspicionata metastazarea tumorii;
E. Pacienta sufera de hipomagnezemie.
50. Un tanar de 20 ani, diabetic insulinodependent, este gasit fara cunostinta langa bicicleta sa de apartament. Istoricul releva faptul ca pacientul a consumat excesiv alcool cu o ora inainte.
Ce investigatii considerate ca sunt necesare in acest caz?
A. Determinarea corpilor cetonici din urina;
B. Determinarea glicemiei;
C. Determinarea fosfatazei alkaline;
D. Determinarea creatin kinazei;
E. Determinarea AST si LDH.
51. Un tanar de 20 ani, diabetic insulinodependent, este gasit fara cunostinta langa bicicleta sa de apartament. Istoricul releva faptul ca pacientul a consumat excesiv alcool cu o ora inainte.
Ce investigatii considerate ca sunt irelevante in acest caz?
A. Determinarea cetonuriei;
B. Determinarea glucozuriei;
C. Masurarea concentratiei sanguine de glucoza;
D. Determinarea activitatii γ glutamil transpeptidazei;
E. Determinarea transaminazelor serice.
52. Un pacient de 53 ani , nefumator, obez, hipertensiv, a fost investigat intr-un control de rutina. Rezultatele sunt:
Valori normale
C olesterol total 8,4 mmol/L 3,9-5,7 mmol/L
Triglyceride 6,8 mmol/L < 2,5 mmol/L
Page 115 of 120
Glucoza 9,8 mmol/L 3,6-6,1 mmol/L
γ glutamil transpeptidaza 138 U/L < 36 U/L
Ce alte informatii si investigatii dintre cele mentionate mai jos considerati ca sunt necesare pentru un diagnostic corect?
A. Hemoglobina glicozilata;
B. Fosfataza alcalina;
C. Prezenta glucozei in urina;
D. LDH;
E. Creatin kinaza
53. Un pacient de 53 ani , nefumator, obez, hipertensiv, a fost investigat intr-un control de rutina. Rezultatele sunt:
Valori normale
C olesterol total 8,4 mmol/L 3,9-5,7 mmol/L
Triglyceride 6,8 mmol/L < 2,5 mmol/L
Glucoza 9,8 mmol/L 3,6-6,1 mmol/L
γ glutamil transpeptidaza 138 U/L < 36 U/L
Ce alte informatii si investigatii dintre cele mentionate mai jos considerati ca nu sunt necesare pentru un diagnostic corect?
A. Transaminazele serice;
B. Creatinina
C. Urea
D. HDL-colesterol;
E. LDL-colesterol.
54. Un pacient de 72 ani se plange de dureri in epigastru si in abdomenul inferior de 2 luni. Examenul echografic a relevant un ficat marit. Pacientul a consumat mari cantitati de alcool in decursul vietii sale. Rezultatele examenului biochimic au fost:
23 iunie
Valori normale
Bilirubina 24 μmoli/L 3,5-20,5 μmoli/L
Fosfataza alkaline 1540 U/L 80-280 U/L
AST 83 U/L 0-41 U/L
ALT 93 U/L 0-45 U/L
LDH 430 U/L 55-140 U/L
Page 116 of 120
γ glutamil transpeptidaza 719 U/L < 36 U/L
1 iulie
Bilirubina 25 μmoli/L
Fosfataza alkaline 2179 U/L
AST 80 U/L
ALT 107 U/L
LDH 440 U/L
γ glutamil transpeptidaza 1020 U/L
Ce consideratii sunt plauzibile in acest caz?
A. Pacientul sufera de icter hepatic;
B. Valorile crescute ale fosfatazei alkaline si ale γ glutamil transpeptidazei, impreuna cu cresterile moderate ale transaminazelor sugereaza colestaza;
C. In lipsa altor simptome, colestaza se datoreaza cirozei hepatice;
D. Pacientul sufera de hepatita virala;
E. Pacientul sufera de complicatii ale diabetului zaharat.
55. Un pacient de 72 ani se plange de dureri in epigastru si in abdomenul inferior de 2 luni. Examenul echografic a relevant un ficat marit. Pacientul a consumat mari cantitati de alcool in decursul vietii sale. Rezultatele examenului biochimic au fost:
23 iunie
Valori normale
Bilirubina 24 μmoli/L 3,5-20,5 μmoli/L
Fosfataza alkaline 1540 U/L 80-280 U/L
AST 83 U/L 0-41 U/L
ALT 93 U/L 0-45 U/L
LDH 430 U/L 55-140 U/L
γ glutamil transpeptidaza 719 U/L < 36 U/L
1 iulie
Bilirubina 25 μmoli/L
Fosfataza alkaline 2179 U/L
AST 80 U/L
ALT 107 U/L
LDH 440 U/L
γ glutamil transpeptidaza 1020 U/L
Ce consideratii nu sunt plauzibile conform datelor prezentate?
Page 117 of 120
A. Pacientul sufera de afectiuni cardiace;
B. Pacientul are un icter prehepatic, datorita unei presumptive anemii hemolitice;
C. Pacientul este suspectat de o ciroza hepatica la care a contribuit consumul ridicat de alcool din decursul vietii;
D. Valorile parametrilor hepatici sunt profund alterate;
E. Pacientul prezinta simptome de guta.
56. Un pacient de 50 ani s-a sculat din somn in timpul noptii din cauza unei dureri intense la nivelul degetelor de la picioare, care erau inflamate si fierbinti.
Care dintre examenele biochimice de mai jos le considerati irelevante pentru stabilirea diagnosticului?
A. Acid uric;
B. Bilirubina;
C. Examenul sumar de urina;
D. Transaminazele serice;
E. Hemoglobina glicozilata.
57. Un pacient in varsta de 41 ani este internat in spital in urma unui colaps la locul de munca. Relateaza ca are simptome similare cu cele gripale de 4 zile (mialgie, dureri de cap, greata, voma si diaree). La sosire valorile enzimelor serice au fost:
Valori normale
AST 149 U/L 0-41 U/L
ALT 88 U/L 0-45 U/L
LDH 1330 U/L 55-140 U/L
Creatin kinaza 6000 U/L <150 U/L
La aceste valori alterate ar fi putut contribui:
A. Muschii scheletici;
B. Muschiul cardiac;
C. Pancreasul;
D. Rinichii;
E. Tiroida.
Page 118 of 120
58. Un pacient in varsta de 41 ani este internat in spital in urma unui colaps la locul de munca. Relateaza ca are simptome similare cu cele gripale de 4 zile (mialgie, dureri de cap, greata, voma si diaree). La sosire valorile enzimelor serice au fost:
Valori normale
AST 149 U/L 0-41 U/L
ALT 88 U/L 0-45 U/L
LDH 1330 U/L 55-140 U/L
Creatin kinaza 6000 U/L <150 U/L
Dintre testele de mai jos, care ar putea contribui la identificarea surselor de enzime deversate in sange?
A. glutamil tranpeptidaza;
B. Izoenzimele LDH;
C. Hemoglobina glicozilata;
D. Creatinina;
E. Colesterolul.
59. Un pacient in varsta de 72 ani se prezinta la medical de familie intr-o stare confuza. Prezinta deficiente de mers si de echilibru si respiratia are un slab miros de alcool.
Ce teste biochimice recomandati?
A. glutamil tranpeptidaza;
B. Hemoglobin glicozilata;
C. Bilirubina;
D. Creatin kinaza;
E. Fosfataza alcalina.
60. O pacienta in varsta de 48 ani prezinta o stare alterata de sanatate, cu greata, balonari, lipsa de apetit. Rezultatele examenului biochimic se prezinta astfel:
Valori normale
Bilirubina totala 3,1 mg/dL 0,2-1,2 mg/dL
ALT 48 U/L 0-45 U/L
Fosfataza alcalina 1800 U/L 90-280 U/L
Γ glutamil tranpeptidaza 840 U/L < 36 U/L
Uree 2,2 mmol/L 2-8,3 mmol/L
Ce diagnostice prezumati?
Page 119 of 120
A. Hepatita virala acuta;
B. Calculi biliari;
C. Diskinezie biliara;
D. Ciroza hepatica;
E. Insuficienta renala.
61. Un pacient de 47 ani, obez, soseste la serviciul de urgenta cu deshidratare si oligurie.
Rezultatele examenului biochimic se prezinta astfel:
Valori normale
Glicemie 98 mg/dL 65-110 mg/dL
AST 24 U/L 0-41 U/L
Cholesterol 387 mg/dL 150-220 mg/dL
ALT 32 U/L 0-45 U/L
Creatinina 2,6 mg/dL 0,7-1,5 mg/dL
Uree 11 mmol/L 2-8,3 mmol/L
In acest caz:
A. Este foarte probabil un diagnostic de insuficienta renala;
B. Este probabil un diagnostic de diabet;
C. Este necesar un examen de urina;
D. Pacientul are o disfunctie hepatica, confirmata de valoarea colesterolului total;
E. Este suspectata o pancreatita.
62. Un pacient de 50 ani, cu un istoric de diabet neglijat, se prezinta la medic acuzand oligurie si febra. Examenul biochimic al sangelui se prezinta astfel:
Valori normale
Glicemie 498 mg/dL 65-110 mg/dL
Cholesterol 387 mg/dL 150-220 mg/dL
Creatinina 3,6 mg/dL 0,7-1,5 mg/dL
Uree 22,9 mmol/L 2-8,3 mmol/L
In acest caz, parametrii sugereaza ca:
A. Diabetul netratat a condus la scleroza grasa a ficatului;
B. Diabetul a devenit insulinodependent;
C. Diabetul a condus la insuficienta renala;
Page 120 of 120
D. Se recomanda examenul urinei;
E. Se recomanda explorarea functional a ficatului.
63. Urmatorul tablou biochimic al sangelui sugereaza:
Sange Valori normale
Glicemie 80 mg/dL 65-110 mg/dL
Cholesterol 187 mg/dL 150-220 mg/dL
Creatinina 4,1 mg/dL 0,7-1,5 mg/dL
Uree 18,2 mmol/L 2-8,3 mmol/L
Albumina 1,8 g/dL 3,5-5,5 g/dL
A. Ciroza ;
B. O disfunctie renala;
C. Insuficienta hepatica;
D. Dislipidemie;
E. Insuficienta renala.
Page 1 of 14
Biochimie medicala, generala si aplicata, Anul III
Farmacie, sem. II - (14)
Capitolul 1. Catena respiratorie si fosforilarea oxidativa: componente,
functionare, control, inhibitie, aspecte clinice
Complement Simplu
1 - C. 2 - A. 3 - C. 4 - A. 5 - C. 6 - A. 7 - C. 8 - B. 9 - B. 10 - C.
Complement Multiplu
11 - A. B. D. E. 12 - A. D. E. 13 - B. C. 14 - C. E. 15 - A. B. E. 16 - A. C. D. E. 17 - A. D. 18 - B. D. E. 19 - B. D. 20 - A. B. C. 21 - A. C. E. 22 - A. C. E. 23 - A. C. D. 24 - A. D. E. 25 - C. D. 26 - B. C. 27 - B. C. E. 28 - A. D. E. 29 - B. C. E.
1 - C. 2 - B. 3 - A. 4 - C. 5 - D. 6 - C. 7 - B. 8 - C. 9 - A. 10 - E.
Complement Multiplu
11 - A. C. 12 - B. C. E. 13 - B. C. E. 14 - B. C. D. E. 15 - C. D. E. 16 - A. D. E. 17 - C. E. 18 - B. C. E. 19 - B. C. 20 - B. C. E. 21 - A. B. 22 - B. C. D. 23 - B. D. 24 - A. B. C. 25 - C. D. 26 - B. D. 27 - A. C. E. 28 - B. D. 29 - A. B. C. 30 - B. E.
1 - D. 2 - C. 3 - D. 4 - D. 5 - C. 6 - A. 7 - B. 8 - A. 9 - B. 10 - B.
Complement Multiplu
11 - A. B. 12 - A. B. E. 13 - A. C. E. 14 - A. C. 15 - A. B. 16 - B. C. E. 17 - C. D. 18 - B. D. 19 - A. D. E. 20 - C. E. 21 - A. C. D. 22 - A. D. 23 - B. C. 24 - A. E. 25 - A. C. D. 26 - A. D. 27 - A. C. E. 28 - C. D. 29 - A. D. 30 - B. C. D.
Capitolul 4. Metabolismul glucidelor. Gluconeogeneza si controlul glicemiei: reactii, reglaj enzimatic, reglaj hormonal, aspecte clinice
Complement Simplu
1 - A. 2 - B.
Page 4 of 14
3 - C. 4 - C. 5 - A. 6 - C. 7 - D. 8 - D. 9 - E. 10 - A.
Complement Multiplu
11 - B. C. D. 12 - B. C. D. 13 - A. B. D. 14 - A. B. C. 15 - A. C. E. 16 - A. B. 17 - A. D. E. 18 - B. D. E. 19 - A. E. 20 - A. D. 21 - B. C. E. 22 - B. C. D. 23 - A. C. 24 - B. C. 25 - A. C. E. 26 - C. D. E. 27 - A. B. 28 - B. C. 29 - B. D. 30 - A. D. E.
Capitolul 5. Metabolismul glucidelor. Metabolismul glicogenului: glicogeno-neogeneza si glicogenoliza, controlul sintezei si degradarii glicogenului, aspecte clinice.Suntul pentozo-fosfatilor. Calea acidului glucuronic
Complement Simplu
1 - A. 2 - C. 3 - A. 4 - B. 5 - B.
Page 5 of 14
6 - A. 7 - C. 8 - B. 9 - C. 10 - B.
Complement Multiplu
11 - C. D. 12 - A. C. 13 - A. B. E. 14 - A. B. C. E. 15 - B. D. 16 - A. B. 17 - A. B. C. 18 - B. C. 19 - A. C. 20 - B. C. 21 - A. B. E. 22 - A. D. 23 - C. D. 24 - C. D. 25 - A. D. 26 - A. C. D. 27 - B. C. 28 - B. D. E. 29 - A. B. 30 - B. C. E.
1 - C. 2 - D. 3 - C. 4 - A. 5 - C. 6 - B. 7 - D. 8 - E. 9 - B.
Page 6 of 14
10 - A.
Complement Multiplu
11 - A. B. 12 - A. B. 13 - A. B. D. 14 - A. B. C. 15 - A. C. 16 - A. D. 17 - A. C. D. 18 - B. D. 19 - A. D. 20 - C. D. 21 - A. C. 22 - C. D. E. 23 - A. B. 24 - B. C. E. 25 - C. D. E. 26 - B. C. D. 27 - A. C. E. 28 - A. B. E. 29 - A. C. E. 30 - A. D.
Capitolul 7. Metabolismul lipidelor. Biosinteza acizilor grasi; biosinteza colesterolului, transportul si excretia sa. Aspecte clinice
Complement Simplu
1 - B. 2 - C. 3 - C. 4 - D. 5 - E. 6 - C. 7 - A. 8 - B. 9 - A. 10 - E.
Complement Multiplu
Page 7 of 14
11 - C. D. E. 12 - A. B. D. 13 - A. D. 14 - A. B. E. 15 - A. C. 16 - A. B. D. 17 - A. B. D. 18 - B. C. 19 - C. D. E. 20 - A. B. C. 21 - A. C. D. 22 - B. C. E. 23 - A. C. 24 - A. C. 25 - A. C. D. 26 - C. D. 27 - B. D. E. 28 - B. D. E. 29 - A. B. E. 30 - B. C. E.
Capitolul 8. Metabolismul lipidelor. Notiuni privind metabolismul acil glicerolilor si al sfingolipidelor, aspecte clinice. Schema generala a repartizarii si utilizarii lipoproteinelor in organism, aspecte clinice privind dislipidemiile
Complement Simplu
1 - D. 2 - E. 3 - D. 4 - C. 5 - B. 6 - A. 7 - B. 8 - C. 9 - B. 10 - A.
Complement Multiplu
11 - A. C. 12 - A. B. D. 13 - A. C. E.
Page 8 of 14
14 - B. D. 15 - A. C. D. 16 - A. B. D. 17 - B. C. E. 18 - A. B. 19 - B. C. 20 - A. E. 21 - B. C. 22 - A. C. E. 23 - A. B. D. 24 - B. D. 25 - B. E. 26 - D. E. 27 - C. D. 28 - A. B. D. 29 - B. C. E. 30 - A. B. D.
Capitolul 9. Metabolismul aminoacizilor si al proteinelor. Catabolismul proteinelor si eliminarea azotului aminic; turn-overul proteic; ciclul ureogenetic-etape, reactii, aspecte clinice. Consideratii generale privind catabolismul scheletelor hidrocarbonate ale aminoacizilor, aspecte clinice
Complement Simplu
1 - D. 2 - C. 3 - A. 4 - C. 5 - A. 6 - B. 7 - D. 8 - A. 9 - E. 10 - B.
Complement Multiplu
11 - C. D. 12 - A. D. 13 - A. B. 14 - A. B. C. D. 15 - C. D.
Page 9 of 14
16 - A. B. C. 17 - A. D. E. 18 - B. C. E. 19 - A. C. D. 20 - A. B. C. 21 - C. D. E. 22 - B. D. 23 - A. C. D. 24 - B. D. 25 - C. D. E. 26 - A. B. D. 27 - A. C. 28 - B. D. E. 29 - A. C. E. 30 - C. D.
Capitolul 10. Metabolismul aminoacizilor si al proteinelor. Biosinteza proteica: etape, reactii, modele de control
Complement Simplu
1 - A. 2 - A. 3 - E. 4 - B. 5 - A. 6 - C. 7 - B. 8 - C. 9 - D. 10 - C.
Complement Multiplu
11 - B. D. E. 12 - B. C. 13 - D. E. 14 - B. C. E. 15 - A. B. 16 - B. C. 17 - A. C. 18 - A. C. E. 19 - A. D. E.
Page 10 of 14
20 - B. D. E. 21 - B. D. E. 22 - A. C. D. 23 - A. B. 24 - C. D. 25 - B. C. 26 - A. D. 27 - C. D. 28 - A. B. E. 29 - A. B. E. 30 - A. B. D.
1 - D. 2 - B. 3 - A. 4 - E. 5 - D. 6 - A. 7 - C. 8 - B. 9 - D. 10 - B.
Complement Multiplu
11 - A. D. 12 - A. C. E. 13 - B. D. E. 14 - A. B. E. 15 - B. C. 16 - C. D. 17 - A. C. 18 - B. C. 19 - A. B. E. 20 - B. D. 21 - B. C. 22 - B. C. D. 23 - C. D.
Page 11 of 14
24 - A. D. E. 25 - A. C. 26 - A. C. 27 - A. C. 28 - B. C. 29 - B. C. E. 30 - B. C. E.
Capitolul 12. Metabolismul nucleotidelor purinice si pirimidinice. Biosinteza si catabolizarea nucleotidelor purinice. Biosinteza si catabolizarea nucleotidelor pirimidinice. Aspecte clinice
Complement Simplu
1 - B. 2 - C. 3 - A. 4 - B. 5 - B. 6 - B. 7 - A. 8 - B. 9 - E. 10 - E.
Complement Multiplu
11 - A. C. D. 12 - B. C. 13 - A. D. 14 - A. B. D. E. 15 - A. C. E. 16 - B. C. 17 - B. C. 18 - A. C. D. 19 - B. D. 20 - A. C. E. 21 - B. C. 22 - A. B. 23 - A. D. 24 - A. B. C. 25 - A. B. D. 26 - A. D.
Page 12 of 14
27 - A. B. D. 28 - A. B. 29 - B. C. E. 30 - A. B. C. E.
Capitolul 13. Notiuni de biochimie aplicata in clinica. Profilul metabolic al organelor principale. Constituientii biochimici principali ai sangelui si urinei. Disfunctii metabolice caracteristice diabetului zaharat
Complement Simplu
1 - A. 2 - D. 3 - E. 4 - B. 5 - C. 6 - B. 7 - C. 8 - D. 9 - C. 10 - E.
Complement Multiplu
11 - A. D. 12 - C. D. 13 - B. C. D. E. 14 - A. B. C. 15 - A. B. C. 16 - B. D. 17 - A. D. 18 - A. B. 19 - A. C. E. 20 - A. D. 21 - A. B. 22 - B. C. 23 - B. D. E. 24 - A. B. 25 - A. C. D. 26 - B. C. D. 27 - A. D. E. 28 - A. D. 29 - C. D. E.
Page 13 of 14
30 - B. C.
Capitolul 14. Notiuni de pato-biochimie. Perturbari metabolice in etilism. Perturbari metabolice si parametri de diagnostic in afectiunile hepato-biliare. Perturbari metabolice si parametri de diagnostic in afectiunile cardio-vasculare
Complement Simplu
1 - B. 2 - C. 3 - C. 4 - D. 5 - A. 6 - E. 7 - D. 8 - A. 9 - B. 10 - A.
Complement Multiplu
11 - A. B. 12 - A. B. E. 13 - B. C. D. 14 - B. C. D. 15 - B. D. 16 - A. B. C. E. 17 - A. B. 18 - A. C. 19 - A. B. 20 - A. D. 21 - B. D. E. 22 - A. B. D. 23 - A. B. 24 - B. C. 25 - A. D. E. 26 - A. D. E. 27 - A. C. D. 28 - B. C. 29 - B. D. 30 - A. B. 31 - C. D. 32 - B. D.
Page 14 of 14
33 - B. C. 34 - A. D. 35 - C. D. 36 - B. D. 37 - B. D. 38 - B. E. 39 - D. E. 40 - A. B. 41 - C. D. 42 - C. D. 43 - A. B. 44 - A. C. 45 - B. C. E. 46 - C. D. 47 - B. C. 48 - A. B. 49 - A. B. D. 50 - A. B. 51 - B. D. E. 52 - A. C. 53 - A. B. C. 54 - B. C. 55 - A. B. E. 56 - B. D. E. 57 - A. B. 58 - A. B. 59 - A. D. E. 60 - B. C. 61 - A. C. 62 - C. D. 63 - B. E.